The Impact of Water Content on Sources of Heterotrophic Soil Respiration


Department of Forest Resources & Environmental Conservation, Virginia Tech, Blacksburg, VA 24061, USA
*
Author to whom correspondence should be addressed.
Forests 2017, 8(8), 299; https://doi.org/10.3390/f8080299
Submission received: 13 June 2017
/
Revised: 7 August 2017
/
Accepted: 11 August 2017
/
Published: 16 August 2017
(This article belongs to the Collection Forests Carbon Fluxes and Sequestration)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Heterotrophic respiration (RH) is a major flux of CO2 from forest ecosystems and represents a large source of uncertainty in estimating net ecosystem productivity (NEP) using regional soil respiration (RS) models. RH from leaf litter (RHL) may contribute greatly to annual RH estimates, but its contribution may be misrepresented due to the logistical and technical challenges associated with chamber-based field measurements of RHL. The purpose of this study was to evaluate the sensitivity of sources of RH (mineral soil-derived heterotrophic respiration [RHM] and leaf litter-derived heterotrophic respiration [RHL]) of a loblolly pine plantation (Pinus taeda L.) to varying soil and litter water content over the course of a dry down event. Additionally, we investigated whether fertilization influenced RHL and RHM to understand how forest nutrient management may impact forest soil carbon (C) dynamics. RHL was measured under dry conditions and at field capacity to evaluate water content controls on RHL, determine the duration of increased CO2 release following wetting, and evaluate the potential contribution to total RH. We also measured RHM inside collars that excluded plant roots and litter inputs, from field capacity until near-zero RHM rates were attained. We found that RHL was more sensitive to water content than RHM, and increased linearly with increasing litter water content (R2 = 0.89). The contribution of RHL to RH was greatest immediately following the wetting event, and decreased rapidly to near-zero rates between 3 and 10 days. RHM also had a strong relationship with soil water content (R2 = 0.62), but took between 200 and 233 days to attain near-zero RHM rates. Fertilization had no effect on RHM (p = 0.657), but significantly suppressed RHL rates after the wetting event (p < 0.009). These results demonstrate that there is great temporal variability in both CO2 released and the water content of differing sources of RH, and forest fertilization may largely impact forest floor C stocks. This variability may not be captured reliably using conventional weekly to monthly chamber-based field sampling efforts and could lead to over- or underestimation of RH. In the context of climate change, changes in the frequency and intensity of wetting and drying events will likely alter RHL and its contribution to RS. Separate consideration of RH sources and controls, along with increased field sampling frequency using chamber-based methodology under a broader range of specific environmental conditions, are likely needed to reduce variability in RH estimates and improve the accuracy of forest NEP predictions.
1. Introduction
Current climate models predict a change in annual precipitation regimes for the southeastern United States, and it is important to understand how these changes will affect the carbon (C) balance of forest ecosystems in this region [1,2]. Southeastern forests currently have among the highest potential for C storage, as they represent nearly 60% of the regional land area, are aggrading biomass with age and with intensive management practices [3,4], and have high rates of net primary productivity (NPP) [1,2,5]. Altered regional precipitation and evapotranspiration, or hydroclimate, is posited to have the largest consequences for productivity and C sequestration potential. Water balance of a site can attenuate or exacerbate effects on NPP as a result of elevated atmospheric CO2, warming temperatures, and consequently longer growing seasons [2,6]. While increases in annual temperature of 1–4 °C [1,7], along with increases in annual precipitation of up to 6 percent, are generally predicted for this region [2,7,8], intensification of annual precipitation regimes is expected, meaning exacerbated wet and dry periods, increases in the frequency of summer droughts, and larger rainfall events between drought periods [9,10,11]. Drier summers can counter early season growth in the southeast by increasing evapotranspiration and decreasing soil moisture, which is a limiting factor controlling NPP, the decomposition of organic matter pools, and thus C and nutrient cycling [10]. The interaction of increasing growing season temperatures and decreasing growing season precipitation may potentially decrease the C sequestration capacity of southern pine forests by 10% [12], and increase the variability of ecosystem C flows.
To understand how changes in moisture dynamics may influence the net C balance or net ecosystem production (NEP) of southeastern forests, the separation of total soil respiration (RS) into components of heterotrophic, microbial respiration (RH; i.e., decomposition) and autotrophic, root respiration (RA) is necessary [13,14]. NEP is estimated from the balance between net primary production (NPP) and RH [15]. Recently, considerable effort has been devoted to quantifying fluxes of CO2 from southeastern pine forests to develop regional respiration models [16], and to improve predictions of NEP by quantifying the RH contribution to RS [17,18,19,20]. These efforts have yielded highly variable predicted contributions of RH to RS in these ecosystems, ranging from of 55–84 percent [19,20,21]. This variability may be attributed, in part, to the fact that sources of RS have different sensitivities to controlling factors such as water content, soil temperature, and substrate availability and composition [22,23]. Heterotrophic sources in particular have been shown to be more sensitive to water content than RA [24,25]. Within heterotrophic sources, RHL is more dynamic than RHM, as surface litter layers are more susceptible to extremes and rapid changes in temperature and water content than subsurface mineral layers, and have a greater dependency on moisture than temperature [25,26]. Additionally, RHL and RHM rates differ temporally due to differences in substrate type and availability, and microbial community abundance and composition [27,28,29].
RHL may be a large source of uncertainty when estimating RS and NEP due to the transient nature of this C pool and its properties. RHL may contribute from 11–51% of RS [26,30,31,32,33], suggesting that the forest floor could be a primary source of C loss from forest ecosystems. However, the contribution of RHL to RS is largely dependent upon litter water content and the frequency and amount of precipitation [26,32]. Our understanding of the controls over RHL is limited by the lack of direct RHL measurements and the inability to non-destructively and continuously measure leaf litter water content. Attempted seasonal in situ spot measurements of RHL using chamber-based methods have yielded near-zero rates in loblolly pine forests of Virginia. These near-zero rates may be a result of the dynamic nature of litter moisture content and the rapid and transient CO2 pulse following surface wetting events (i.e., Birch effect) [24,34,35], which may be missed with non-continuous spot measurements. The difference in temporal resolution of moisture control over litter compared to mineral soil is important to rectify to improve estimates of RH.
To understand how RHL and RHM respond independently to water content, we measured these variables on samples collected from a mid-rotation loblolly pine forest over the course of a dry down event. Additionally, we compared RHL and RHM from samples collected from fertilized and unfertilized plots, as forest fertilization is a common management practice in southeastern pine forests [36], with the potential to affect C turnover and respiration rates with separate studies showing everything from increases to decreases, to even no effect [37]. This ambiguity has been attributed, in part, to the divergent and potentially counteracting response of sources of RA and RH to mineral nutrient additions, with RA often increasing with fertilization due to increased biomass [38,39] and RH decreasing due to the decline in microbial activity [40]. RHL was measured first under dry conditions and then at field capacity to evaluate the temporal patterns occurring as a result of a wetting event. We hypothesized that RHL would be more sensitive to water content than RHM, in part, due to greater availability of labile C and greater microbial biomass in litter relative to mineral soil [41,42], and that fertilization would result in suppressed rates of respiration in both the mineral soil and leaf litter. Partitioning RH into RHM and RHL can enhance our understanding of controls over these components, and identify sources of variability in ecosystem RS and NEP estimates.
2. Materials and Methods
2.1. Site Description
This research is a component of the Pine Integrated Network: Education, Mitigation, and Adaptation project (PINEMAP; http://pinemap.org/). This regional study was designed to evaluate the effects of decreased rainfall and nutrient additions on loblolly pine plantation productivity and physiology [43]. Four locations were established to span the full temperature and precipitation range of the species. Leaf litter samples and soil cores used in this incubation study were collected from a 12-year-old loblolly pine (Pinus taeda L.) plantation located in the Appomattox-Buckingham State Forest, Virginia, USA (37°27′37″ N, 78°39′50″ W), an existing 1.3-ha, long-term throughfall reduction x fertilization study installed during spring 2012. Additional information on study installation and site characteristics can be found in Will et al. [43].
Mean annual precipitation for this region is 109 cm. The average growing season temperature (April through September) is 22.9 °C, and the average winter temperature (December through February) is 3.8 °C [44]. Soils are well drained, formed in residuum from metamorphic rocks (sericite schist, graphitic schist, and/or phyllite). Soils are mapped as Spears Mountain (fine, mixed, semiactive, mesic Typic Hapludult) and Littlejoe (fine, mixed, subactive, mesic, Typic Hapludult) series [45].
2.2. Soil and Litter Sampling
The field study was designed as a randomized complete block with four replications of a 2 × 2 factorial of throughfall reduction (0 and 30% reduction) and fertilization with either no addition, or a complete suite of essential nutrients to represent optimum nutrition, randomly assigned to each block. However, for this incubation study, litter and soil samples were collected from the four control and fertilization treatment plots only. The fertilization treatment was broadcast applied once in spring 2012. Nitrogen (N) was applied at a rate of 224 kg N ha−1 as urea, and phosphorus (P) was applied at a rate of 27 kg P ha−1 as diammonium phosphate. Elemental potassium (K) was applied at a rate of 56 kg K ha−1 as potash, and a granular oxysulfate micronutrient mix (Southeast Mix, Cameron Chemicals, Inc., Virginia Beach, VA, USA) consisting of 6% sulfur (S), 5% boron (B), 2% copper (Cu), 6% manganese (Mn), and 5% zinc (Zn) was applied at a rate of 22.4 kg ha−1. Treatment plots measured 14.6 × 16.8 m, surrounded by a 6.1-m buffer on all sides.
To facilitate independent measures of RHM, root exclusion cores constructed from steel conduit pipe (11.43 cm diameter × 35 cm length) were installed in the summer of 2013 in three locations within each treatment plot to sever roots for a preceding study [17]. Cores were driven vertically into the soil until the top was flush with the mineral soil surface. Prior to core installation, the O horizon (forest floor) was temporarily removed from the installation location to avoid driving organic matter into the soil profile or inside the core, and replaced after installation. Carbohydrate supply to roots is assumed to be depleted 90 days post-installation, and root-derived, autotrophic respiration (RA) inside the pipe is assumed to be zero, thereby providing an independent measure of RHM [46]. This laboratory study utilized the existing intact root-severing field cores to isolate RHM from RA in a controlled environment. Cores were extracted from the field locations on four separate occasions from June through July 2015, loosely wrapped with plastic to prevent soil and moisture loss, and transported to a temperature-controlled lab within four hours of collection. Mean air temperature and relative humidity (RH) were measured daily throughout the duration of the experiment using an Amprobe THWD-5 RH and temperature meter (Danaher Corporation USA, Everett, Washington, DC, USA). Mean lab air temperature was 22.5 ± 0.07 and mean RH was 48.7 ± 0.04. On each of the four field sampling occasions, three replicate cores were collected from one control and one fertilization treatment plot located within one block, with the exception of block two, where only two replicates were collected due to the inability to locate one previously installed root-severing core. This sampling resulted in a total of 11 replicate cores from each treatment. Cores remained root-free while in the field, as no visible root ingrowth from the bottom up occurred, and any vegetation growth inside the pipe was clipped in advance of extraction. Once in the lab, cores were placed upright with their bottoms in 10 cm of oven-dried (105 °C) sterilized sand to maintain their upright position and minimize the likelihood of ambient air entering through the bottom of the core during subsequent respiration measurements. Cores were allowed to equilibrate for 48 h prior to initiating respiration measurements.
Leaf litter from the Oi horizon was collected concurrently with the root-exclusion core extractions, for a total of 11 replicate samples from each treatment. The Oi horizon at this site was approximately 0.04 m thick. Litter was collected adjacent to each of the three root-severing cores from a 10 cm × 10 cm area, homogenized, and composited into one sample from each of the eight (2 fertilized treatments × 4 blocks) treatment plots. Litter samples were placed into paper bags and transported to the lab, then removed from bags and placed on trays to air-dry for 48 h before initiating the wetting event and respiration measurements.
2.3. RHM and RHL Laboratory Measurements
Eleven replicate microcosms for each treatment, three each from blocks one, two, and four, and two from block three, were constructed using the extracted root-severing cores and PVC collars placed atop the cores to contain corresponding litter samples. Air-dry litter from each treatment plot was homogenized and 20 g was added to a 10-cm diameter × 10-cm height PVC collar to simulate the quantity and depth of the Oi horizon from the field site. One layer of 2-mm mesh size gray fiberglass screen was applied to the bottom of each collar to prevent litter loss and allow for water drainage. Microcosms (litter collar + soil core) remained intact during the duration of the incubation, except when independent respiration measurements were being made. The initial weight of the litter collars and soil cores were measured before the start of the dry down, and immediately after each respiration measurement during the course of the experiment for later determination of gravimetric water content (GWC) at the time of each measurement. Soil and litter water content was expressed by weight as the ratio of the mass of water present to the dry weight of the samples. The water mass was determined by drying the soil core or litter microcosm after the incubation was completed to a constant weight and measuring the sample before and after drying. Soil cores were dried to a constant weight in a drying oven at 105 °C and litter was dried to a constant weight at 65 °C. Respiration measurements were made using a LI-8100A automated soil gas flux system and LI-8100-102 10-cm Survey Chamber (LI-COR Bioscience Inc., Lincoln, NE, USA). Measurements were logged for 90 s following CO2 equilibration within the chamber. Laboratory air temperature and relative humidity were controlled for the duration of the incubation; air temperature remained between 21 and 23 °C, and relative humidity remained between 46 and 49 percent.
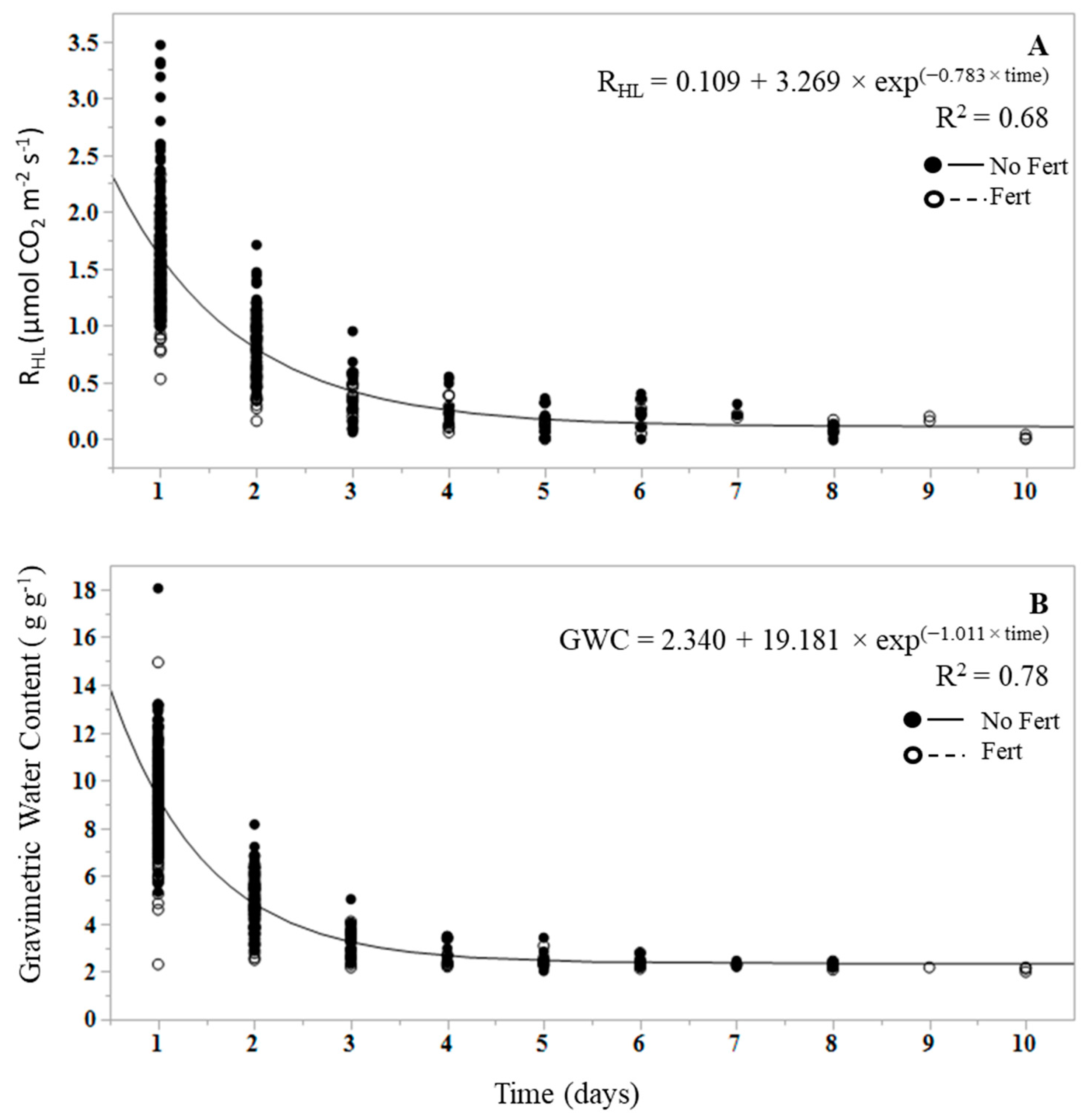
One wetting event was initiated for RHL measurements over the course of the dry down to bring litter to field capacity. Prior to wetting, mass of the sample and an initial respiration measurement were taken to confirm the respiration rates were zero or near-zero (i.e., rates less than 0.20 µmol m−2 s−1) prior to the wetting event. Litter was brought to field capacity by layering the bottom of a 6 L capacity polypropylene container with 50 mL of deionized (DI) water (2 cm depth), then placing the collars in the container and irrigating the tops of the collars with 200 mL of DI water to fully saturate the litter. The container was sealed to bring the container relative humidity to 100%, as measured using the Amprobe THWD-5 RH meter, and litter was allowed to equilibrate for 5 min. The collars were then removed from the container and allowed to drain and the weight was allowed to stabilize. Once excess water was removed, the starting weights were recorded and sequential respirations measurements were initiated. The bottom of the collar was sealed during active respiration measurements using closed cell rubber pipe insulation. Two samples, one from each treatment, were irrigated at a single time, and respiration measurements and sample weights were recorded systematically for 12 h each day until the respiration rate reached 0, or reached a 95% rate reduction from the initial peak rate. The duration of the incubation varied by sample and ranged from 3 to 10 days to achieve near-zero rates.
Soil cores did not receive a wetting event, as they were near field capacity (between 32 and 45 percent volumetric water content) [47,48] at the start of the dry down. Volumetric water content was estimated using a 12-cm HydroSense soil-water sensor (Campbell Scientific USA, Logan, UT, USA) at the time of core extraction, and by converting GWC to VWC (volumetric water content) by multiplying GWC by the bulk density of soil (site-wide mean bulk density from 0–30 cm depth was 1.32 ± 0.16 g cm−3) divided by the density of water (assuming 1.0 g cm−3) [49]. RHM and weight measurements began after the initial 48-h equilibration period following sample collection, and continued until the respiration rates were near zero, or reduced by 95% from the peak measurement rate. The duration of the RHM dry down to achieve near-zero rates varied by core but ranged from 200 to 223 days.
2.4. Statistical Analysis
The strengths of the relationships between RHL and RHM and gravimetric water content for litter and mineral soil for each treatment were analyzed using linear regression with a significance level of α = 0.05 (n = 11). Variables were log transformed for fit or as necessary to achieve normal distribution before the regressions, and log(x + 1) was applied to the efflux values due to the presence of values of zero in the dataset. The difference of the slopes and intercepts of RHM or RHL and moisture between the two treatment types (unfertilized and fertilized) was compared using ANOVA. An exponential decay function was fit to the relationship between RHL and RHM, and GWC over time to derive the decay constant, or k. The three-parameter exponential decay equation is fit using the formula:
All statistical analyses were performed using JMP® 11 software system (SAS Institute, Cary, NC, USA).
3. Results
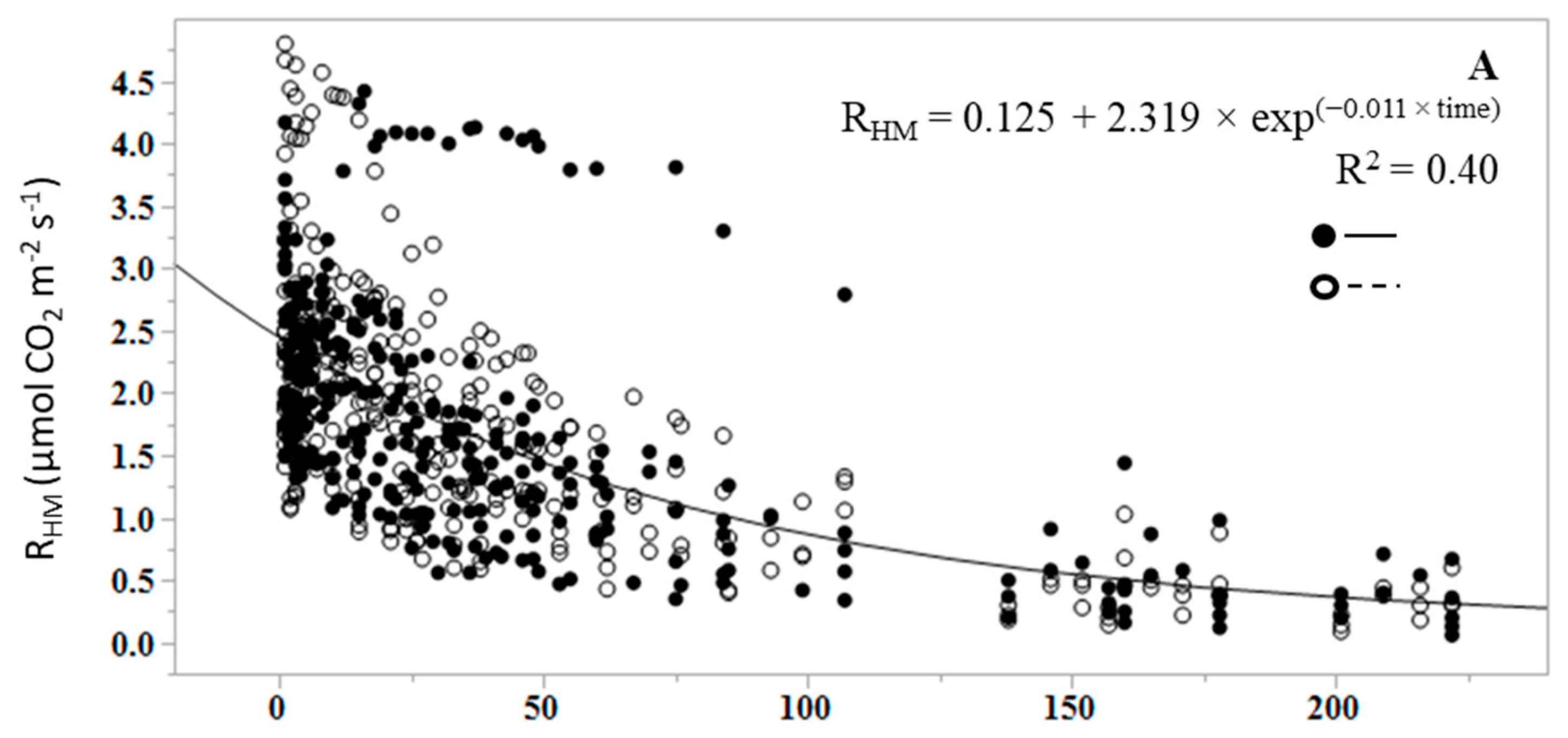
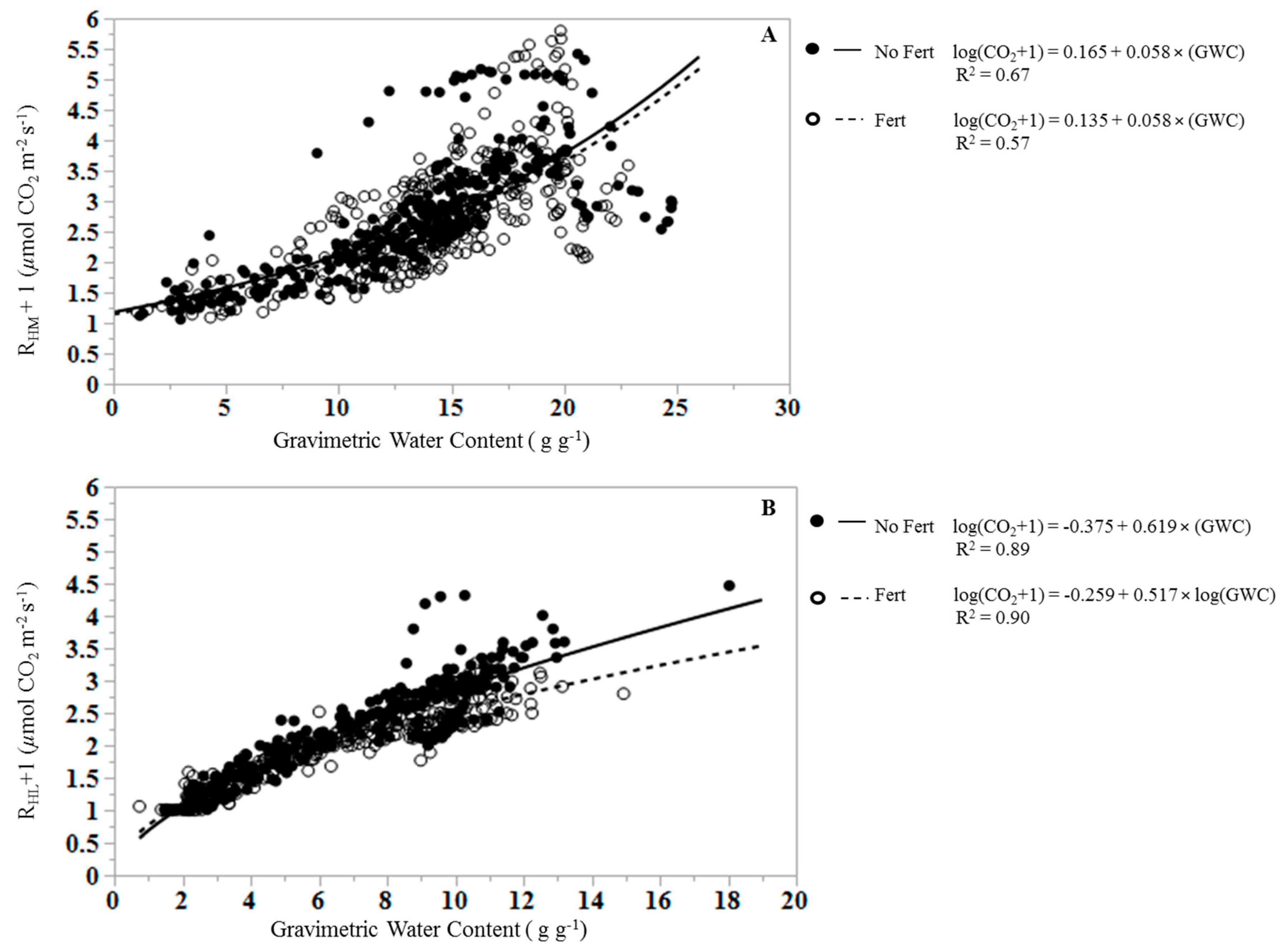
RHL and RHM decreased over the course of the dry down experiment, and with decreasing GWC (Figure 1 and Figure 2). Prior to the wetting event, RHLs of all treatments were near zero and reached a peak rate in the first measurement following the wetting event that brought the litter water content to field capacity (Figure 2). GWC of litter and soil was a significant predictor of RHL and RHM (p < 0.0001), but had greater predictive value and a stronger linear relationship for RHL than for RHM. Across all samples, litter GWC explained 89% of the variance in cumulative RHL (i.e., R2 = 0.89) using the Equation:
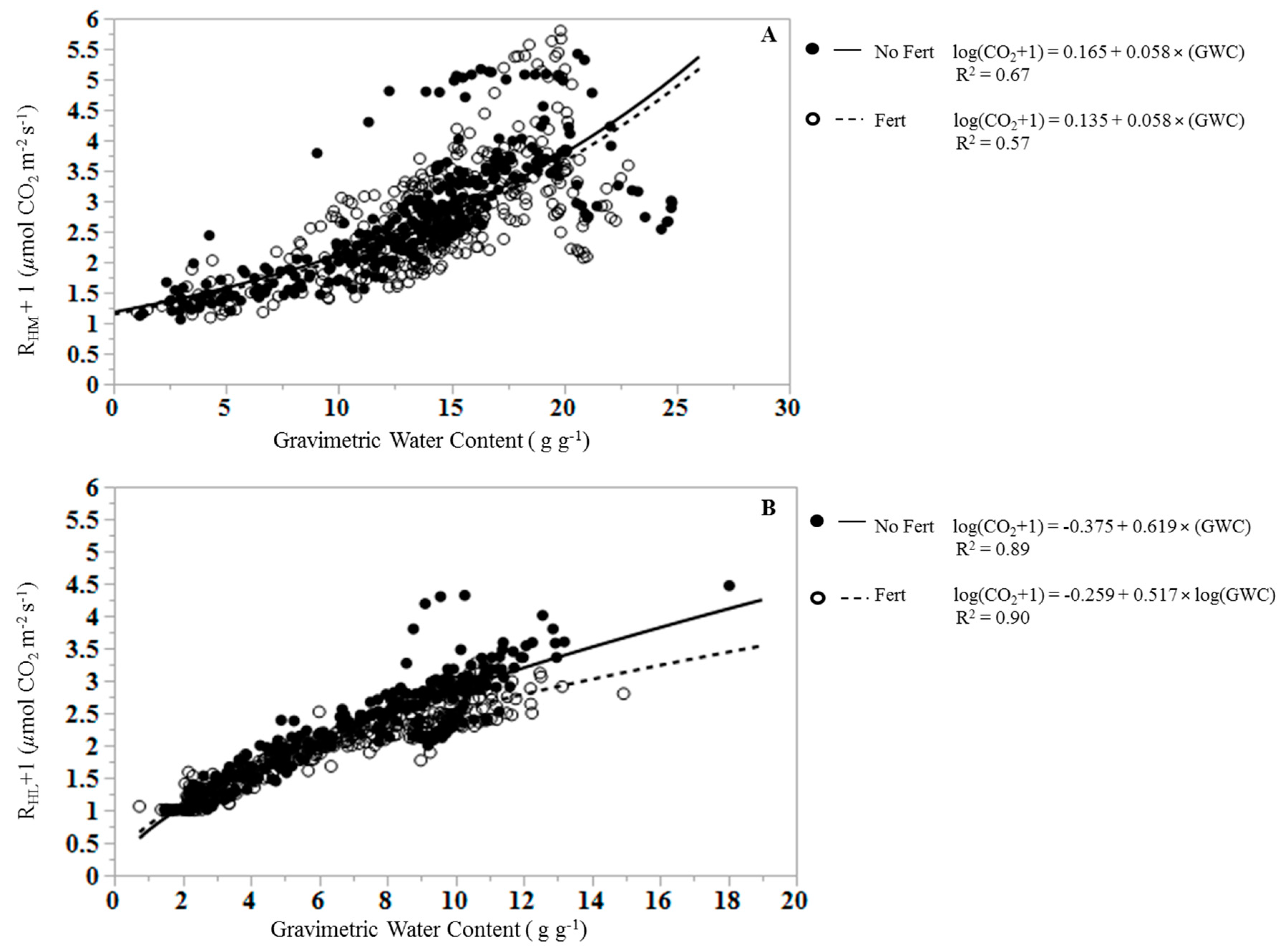
The regression of RHL on GWC differs between fertilized and unfertilized plots (Figure 3) as the slopes of the regression equations differed significantly (p < 0.0001; e.g., 0.619 for unfertilized and 0.517 for fertilized RHL), and the mean response for fertilization differs at zero (p < 0.0001; e.g., intercepts of −0.375 for unfertilized and −0.259 for fertilized RHL). The mean RHL differs by treatment (p = 0.009), with lower mean RHL in fertilized treatments (0.93 ± 0.040 µmol CO2 m−2 s−1) than in unfertilized treatments (1.27 ± 0.042 µmol m−2 s−1).
Soil GWC explained 62% of the variance in cumulative RHM using the equation:
There was no significant difference between the slopes of fertilized and unfertilized regression equations for RHM (p = 0.825), and the mean response for fertilization did not differ at zero (p = 0.604). Fertilization had no effect on RHM (p = 0.657), with an overall mean RHM of 1.705 ± 0.054 µmol CO2 m−2 s−1.
An exponential decay function was fit to quantify and compare the rate (k) at which RHL and RHM, and litter GWC and mineral soil GWC, declined over the duration of the dry down (Figure 1 and Figure 2). Initial CO2 was deemed to be the first respiration measurement at RHL or RHM field capacity. In the case of RHL, the function was fit from the first efflux measurement following the wetting event, as the first measurement of the dry litter was effectively zero. Dry down measurements were terminated once all efflux rates reached 95% of the initial peak efflux rate at the start of the study. The mean decay rate, defined as k, is commonly used in decomposition experiments as the decomposition constant [50]. The mean k values for RHL and RHM were significantly different from each other (p < 0.0001), with a RHL k mean of 0.78 ± 0.04 and a RHM k mean of 0.01 ± 0.003 (Figure 1A and Figure 2A). The k values did not vary with fertilization treatment in either RHL (p = 0.649) or RHM (p = 0.262). Similarly, litter and mineral soil mean GWC k values were significantly different from each other (p < 0.0001), with a litter GWC k of 1.011 and a mineral soil GWC k of 0.009 (Figure 1A,B), but did not differ by fertilization treatment in litter (p = 0.622) or mineral soil (p = 0.379). The change in RHL and litter GWC over time was more rapid than in mineral soil, resulting in higher k values; near-zero RHL rates were reached between 3 and 10 days, while the dry down of mineral soil to near-zero RHM rates under the same conditions required between 200 and 233 days.
4. Discussion
This study demonstrates that water content is a controlling factor for both RHL and RHM, but more strongly influences the magnitude of RHL. Prior to the addition of water, dry leaf litter from fertilized and unfertilized litter samples had RHL rates of zero or near zero. RHL immediately increased following the wetting event, bringing the litter layer to field capacity, to an average rate of 2.65 μmol m−2 s−1 in unfertilized samples and 1.89 μmol m−2 s−1 in fertilized samples, and decreased to pre-wetting levels between 3 and 10 days following the decline in litter water content (Figure 2). Based on our results, RHL increases linearly with litter water content (Figure 3). The rapid decline in RHL as litter water content decreased can be attributed to the osmotic stress experienced by microorganisms within a short time period as litter becomes dry [51]. These results are consistent with previous findings that show that dry litter contributed proportionally little to RS, but following wetting, can contribute 11 to 51 percent of RS, demonstrating the regulatory role of water content on RHL [30,32,42]. While the contribution of RHL to RS was not investigated in this study, we can approximate the RHL contribution to RH (RHL + RHM) when both sample types were at field capacity. Immediately following litter wet-up, RHL contributed between 38 and 69 percent to total RH. Peak RHM values ranged from 1.63 to 4.80 μmol m−2 s−1. Unlike RHL, peak RHM values did not always occur at the highest water content, and in some cases, occurred in days following the start of the dry down. This may be due to the displacement of CO2 from soil pores and aggregates as the soil begins to dry. While the contribution of RHL to RH declined markedly within a day and over the course of the dry down, it does suggest that RHL is an important and dynamic source of C loss from forest soils after wetting events, and the contribution may be important to understanding spatial and temporal variability of RS and RH estimates in forest ecosystems.
The increase in respiration rates following a wetting event, as observed in RHL, has been attributed to a rapid increase in microbial activity and availability of previously unavailable substrates with increased water availability [52,53,54]. These substrates can be made available by the release of osmolytes from microorganisms that accumulated during the dry period as a result of microbial stress prior to a wetting event, from release of some of the cytoplasmic solutes accumulated during the dry period into the soil solution from lysed cells upon wetting [41,55,56,57], and from a change in the kinetics of enzyme transport and microbial uptake with increased water availability [51]. The duration and magnitude of elevated respiration rates is strongly related to water content [31,53], microbial physiology, and community structure as influenced by the physical and chemical characteristics of the environment [51], and therefore likely differs between organic and mineral soil horizons [58]. In mineral soil, the structural alterations in soil aggregates upon wetting may also cause the release of previously physically protected organic matter [24,59].
While the mineral soil in our study did not receive a wetting event, as it was initially within the range of VWC values determined to be field capacity for the soil textural class (i.e., 32–45%) [47,48], we did observe marked differences in sensitivity to water content, and in the duration of the dry down. RHM was more stable over the course of the dry down and less sensitive to decreasing soil water content than RHL, which supports our initial hypothesis. Soil water content explained less of the variation in RHM than litter water content in RHL (R2 0.62 and 0.89, respectively), and as expected, took far longer (>200 days) to dry down to water contents that induced microbial stress, yielding significantly lower mean k values than RHL. However, the duration of this dry down was influenced by the controlled lab environment, and there were no roots present, no transpiration, and no lateral drainage in the mineral soil microcosms. Previous studies have also found respiration from organic horizons to be more sensitive to rainfall and moisture than mineral soil respiration in temperate forests [26,30,42,60]. The reason may be that compared with the mineral soil, litter physical and chemical properties differ, and the water conditions of litter layers are more dynamic [26]. Litter and mineral soil heterotrophic community composition and biomass also differ, which may result in varying physiological responses to wetting and drying events [41,61], and C utilization strategies [62]. Additionally, the C supply in the soil may remain active at lower water potentials, and microbes can persist in microsites where conditions are more suitable [63,64]. These results demonstrate the need for separate measurements of RH sources and environmental controls (water content, temperature, etc.), as mineral soil is protected from extreme changes in environment by the litter layer [29], and there is large spatial and temporal variability of water content and respiration rates.
We expected fertilization to suppress RHL and RHM, as previous studies have reported reduced RH with mineral nutrient additions [39,40,65]. These reductions may be associated with a direct influence on the soil microbial community, such as decreases in fungal biomass following fertilization [66,67,68]. Fertilization lessened the magnitude of the pulse of CO2 released after wetting and resulted in suppressed RHL compared to unfertilized samples. Additionally, the slopes and intercepts of the linear relationship between RHL and litter water content were significantly different (Figure 3). There was no effect of fertilization on RHM or the duration of increased CO2 release following wetting for RHM and RHL, as the exponential curve function values were not significantly different. Previous studies have reported declines in RH with fertilization in temperate ecosystems [39,40,69,70,71], but increases have also been reported in soils with inherently low nutrient status [72]. Declines have been attributed to direct effects on the microbial community associated with osmotic changes and reduced metabolic activity per unit biomass [40]. The variable response to fertilization observed in the litter and mineral soil in this study may be a result of multiple factors as reported in previous studies, such as differences in substrate quality [73,74], nutrient availability, microbial community composition, and microbial biomass [75,76,77]. However, in a previous study conducted at this same study location, the fertilized treatment resulted in improved litter quality (lower C:N ratios), increased potential extracellular enzyme activities, and increased microbial biomass, which does not directly support the suppressed RHL rates reported in this study. Based on the results in this study, reduced RHL with fertilization could lead to the accumulation of litter and higher soil C concentrations as a result of reduced microbial C turnover [78,79].
Improved temporal resolution in relationships between litter water content and RHL measurements are needed to improve estimates of RH in soil respiration models. Field measurements of soil respiration may fail to adequately capture the annual RH contribution to RS due to dynamic moisture conditions, logistical constraints associated with sampling under wet conditions, and the shortage of direct measurements of RHL and litter water content [32]. To remedy this deficiency in current ecosystem C accounting efforts, measurements of heterotrophic respiration components should occur at greater frequencies, and at times that capture a range of moisture conditions and time following precipitation events. Respiration from litter has been demonstrated to have a greater dependency on moisture than temperature, but RHL contributions depend on the frequency and amount of precipitation [26,58]. The southeastern region is predicted to experience greater frequency of summer droughts, and more intense precipitation events between drought periods [9,10,11]. Drier soils have been shown to have a greater magnitude of C loss following wetting events [64,80], as the pulse of CO2 is dependent upon pre-wetting water content. These findings suggest that more severe drought may increase the magnitude of CO2 losses [57,81]. When under wet conditions, however, CO2 losses could be depressed by subsequent rainfall by bringing soils to oxygen-limited levels that inhibit respiration [82,83]. Additionally, the intensity of rainfall events could alter microbial biomass and activity with long-term effects on RH, as rapid changes in water content could contribute to greater incidence of cell lysis [64]. Given the range of predicted changes in regional hydroclimate and the varying sensitivities of RH sources to water content, a better understanding of the mechanisms that influence the control of water content on RH is needed to understand how changes in precipitation regimes will influence the C balance of these ecosystems.
5. Conclusions
The results of this study suggest that routine chamber-based measurements of forest soil respiration components performed in the field on a weekly to monthly sampling regime may not adequately capture the contribution of RHL to RH, and may contribute largely to the variability in soil respiration models used to estimate NEP, particularly under future precipitation scenarios. Established regional respiration models exhibit large amounts of variability (~40%) that cannot be explained by soil temperature, moisture, and other site parameters [16]. Sampling during periods when litter is either wet or dry, and failure to parameterize models with litter water content in addition to soil moisture, may influence this variability. We found that RH sensitivity to water content and fertilization varies among heterotrophic sources (RHL vs. RHM), resulting in great vertical and temporal heterogeneity among RH fluxes. RHL rapidly increased with increases in litter water content, but this response was short-lived and may be missed during field sampling efforts without direct and more frequent measures of RHL and litter water content. Further, failure to directly measure RHL and litter water content could mask management effects on RH, as fertilization led to suppressed RHL but not RHM. These results indicate that greater sampling frequency under varying moisture and environmental conditions is needed to improve our estimates of RH in ecosystem C accounting models.
Acknowledgments
This research was supported, in part, by the Virginia Agricultural Experiment Station and the McIntire-Stennis Program of the National Institute of Food and Agriculture (NIFA), U.S. Department of Agriculture (USDA), and by the Pine Integrated Network: Education, Mitigation, and Adaptation Project (PINEMAP), a Coordinated Agricultural Project funded by the USDA NIFA (Award No. 2011-68002-30185). Thanks to Brian Parr for providing field and lab assistance.
Author Contributions
K.M.M. designed and performed the experiment, collected and analyzed the data, and wrote the manuscript. J.R.S. and B.D.S. assisted with data analysis and interpretation, and critically revised the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mitchell, R.J.; Liu, Y.; O’Brien, J.J.; Elliott, K.J.; Starr, G.; Miniat, C.F.; Hiers, J.K. Future climate and fire interactions in the southeastern region of the United States. For. Ecol. Manag. 2014, 327, 316–326. [Google Scholar] [CrossRef]
- Rocca, M.E.; Miniat, C.F.; Mitchell, R.J. Editorial. Introduction to the regional assessments: Climate change, wildfire, and forest ecosystem services in the USA. For. Ecol. Manag. 2014, 327, 265–268. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J. Climate change and forest disturbances: Climate change can affect forests by altering the frequency, intensity, duration, and timing of fire, drought, introduced species, insect and pathogen outbreaks, hurricanes, windstorms, ice storms, or landslides. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Jokela, E.J.; Martin, T.A.; Vogel, J.G. Twenty-five years of intensive forest management with southern pines: Important lessons learned. J. For. 2010, 108, 338–347. [Google Scholar]
- Wear, D.N.; Greis, J.G. Southern Forest Resource Assessment—Technical Report; General Technical Report SRS-53; USDA Forest Service; Southern Research Station: Asheville, NC, USA, 2002.
- McNulty, S.; Vose, J.; Swank, W. Loblolly pine hydrology and productivity across the southern United States. For. Ecol. Manag. 1996, 86, 241–251. [Google Scholar] [CrossRef]
- Mearns, L.O.; Gutowski, W.; Jones, R.; Leung, R.; McGinnis, S.; Nunes, A.; Qian, Y. A regional climate change assessment program for North America. Eos 2009, 90, 311. [Google Scholar] [CrossRef]
- Ingram, K.T. Climate of the Southeast United States: Variability, Change, Impacts, and Vulnerability; Ingram, K.T., Dow, K., Carter, L., Anderson, J., Eds.; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Groisman, P.Y.; Knight, R.W.; Karl, T.R.; Easterling, D.R.; Sun, B.; Lawrimore, J.H. Contemporary changes of the hydrological cycle over the contiguous United States: Trends derived from in situ observations. J. Hydrometeorol. 2004, 5, 64–85. [Google Scholar] [CrossRef]
- Angert, A.; Biraud, S.; Bonfils, C.; Henning, C.; Buermann, W.; Pinzon, J.; Tucker, C.; Fung, I. Drier Summers Cancel Out the CO2 Uptake Enhancement Induced by Warmer Springs. Proc. Natl. Acad. Sci. USA 2005, 102, 10823–10827. [Google Scholar] [CrossRef] [PubMed]
- Laseter, S.H.; Ford, C.R.; Vose, J.M.; Swift, L.W. Long-term temperature and precipitation trends at the Coweeta Hydrologic Laboratory, Otto, North Carolina, USA. Hydrol. Res. 2012, 43, 890–901. [Google Scholar] [CrossRef]
- Noormets, A.; Gavazzi, M.J.; McNulty, S.G.; Domec, J.C.; Sun, G.; King, J.S.; Chen, J. Response of carbon fluxes to drought in a coastal plain loblolly pine forest. Glob. Chang. Biol. 2010, 16, 272–287. [Google Scholar] [CrossRef]
- Hanson, P.; Edwards, N.; Garten, C.; Andrews, J. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Subke, J.-A.; Inglima, I.; Francesca Cotrufo, M. Trends and methodological impacts in soil CO2 efflux partitioning: A metaanalytical review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Woodwell, G.M.; Whittaker, R.H. Primary production in terrestrial ecosystems. Am. Zool. 1968, 8, 19–30. [Google Scholar] [CrossRef]
- Templeton, B.S.; Seiler, J.R.; Peterson, J.A.; Tyree, M.C. Environmental and stand management influences on soil CO2 efflux across the range of loblolly pine. For. Ecol. Manag. 2015, 355, 15–23. [Google Scholar] [CrossRef]
- Heim, B.C. Partitioning Soil Respiration in Response to Drought and Fertilization in Loblolly Pine: Laboratory and Field Approaches. Ph.D. Thesis, Virginia Polytechnic Institute & State University, Blacksburg, VA, USA, 2014. [Google Scholar]
- Noormets, A.; Epron, D.; Domec, J.; McNulty, S.; Fox, T.; Sun, G.; King, J. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef]
- ArchMiller, A.; Samuelson, L. Partitioning longleaf pine soil respiration into its heterotrophic and autotrophic components through root exclusion. Forests 2016, 7, 39. [Google Scholar] [CrossRef]
- McElligott, K.M.; Seiler, J.R.; Strahm, B.D. Partitioning soil respiration across four age classes of loblolly pine (Pinus taeda L.) on the Virginia Piedmont. For. Ecol. Manag. 2016, 378, 173–180. [Google Scholar] [CrossRef]
- Brown, R.M. Soil Heterotrophic Respiration in Loblolly Pine Plantations: Measuring and Modeling Seasonal Variation and Silvicultural Impacts. Ph.D. Thesis, University of Georgia, Athens, GA, USA, 2016. [Google Scholar]
- Luo, Y.; Zhou, X. Soil Respiration and the Environment; Academic Press: Burlington, MA, USA, 2006. [Google Scholar]
- Suseela, V.; Conant, R.T.; Wallenstein, M.D.; Dukes, J.S. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Glob. Chang. Biol. 2012, 18, 336–348. [Google Scholar] [CrossRef]
- Inglima, I.; Alberti, G.; Bertolini, T.; Vaccari, F.; Gioli, B.; Miglietta, F.; Cotrufo, M.; Peressotti, A. Precipitation pulses enhance respiration of Mediterranean ecosystems: The balance between organic and inorganic components of increased soil CO2 efflux. Glob. Chang. Biol. 2009, 15, 1289–1301. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Piao, S.; Janssens, I.A.; Tang, J.; Liu, W.; Chi, Y.; Wang, J.; Xu, S. Soil respiration under climate warming: Differential response of heterotrophic and autotrophic respiration. Glob. Chang. Biol. 2014, 20, 3229–3237. [Google Scholar] [CrossRef] [PubMed]
- Hanson, P.J.; O’Neill, E.G.; Chambers, M.L.S.; Riggs, J.S.; Joslin, J.D.; Wolfe, M.H. Soil respiration and litter decomposition. In North American Temperate Deciduous Forest Responses to Changing Precipitation Regimes; Hanson, P.J., Wullschleger, S.D., Eds.; Springer: New York, NY, USA, 2003; pp. 163–189. [Google Scholar]
- Tewary, C.; Pandey, U.; Singh, J. Soil and litter respiration rates in different microhabitats of a mixed oak-conifer forest and their control by edaphic conditions and substrate quality. Plant Soil 1982, 65, 233–238. [Google Scholar] [CrossRef]
- Boone, R.D.; Nadelhoffer, K.J.; Canary, J.D.; Kaye, J.P. Roots exert a strong influence on the temperature sensitivityof soil respiration. Nature 1998, 396, 570–572. [Google Scholar] [CrossRef]
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in forest ecosystem and landscape ecology variations in local climate can be used to monitor and compare the effects of different management regimes. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef]
- Cisneros-Dozal, L.M.; Trumbore, S.E.; Hanson, P.J. Effect of moisture on leaf litter decomposition and its contribution to soil respiration in a temperate forest. J. Geophys. Res. 2007, 112. [Google Scholar] [CrossRef]
- DeForest, J.L.; Chen, J.; McNulty, S.G. Leaf litter is an important mediator of soil respiration in an oak-dominated forest. Int. J. Biometeorol. 2009, 53, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ataka, M.; Kominami, Y.; Jomura, M.; Yoshimura, K.; Uematsu, C. CO2 efflux from leaf litter focused on spatial and temporal heterogeneity of moisture. J. For. Res. 2014, 19, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Ge, X.; Zeng, L.; Huang, Z.; Lei, J.; Zhou, B.; Li, M. Rates of litter decomposition and soil respiration in relation to soil temperature and water in different-aged Pinus massoniana forests in the Three Gorges Reservoir area, China. PLoS ONE 2014, 9, e101890. [Google Scholar] [CrossRef] [PubMed]
- Birch, H. The effect of soil drying on humus decomposition and nitrogen availability. Plant Soil 1958, 10, 9–31. [Google Scholar] [CrossRef]
- Jarvis, P.; Rey, A.; Petsikos, C.; Wingate, L.; Rayment, M.; Pereira, J.; Banza, J.; David, J.; Miglietta, F.; Borghetti, M. Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: The “Birch effect”. Tree Physiol. 2007, 27, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Fox, T.R.; Allen, H.L.; Albaugh, T.J.; Rubilar, R.; Carlson, C.A. Tree nutrition and forest fertilization of pine plantations in the southern United States. South. J. Appl. For. 2007, 31, 5–11. [Google Scholar]
- Knorr, W.; Prentice, I.; House, J.; Holland, E. Long-term sensitivity of soil carbon turnover to warming. Nature 2005, 433, 298–301. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Mattson, K.G.; Zaerr, J.B.; Marshall, J.D. Root respiration of Douglas-fir seedlings: Effects of N concentration. Soil Biol. Biochem. 1998, 30, 331–336. [Google Scholar] [CrossRef]
- Gough, C.; Seiler, J.; Maier, C.A. Short-term effects of fertilization on loblolly pine (Pinus taeda L.) physiology. Plant Cell Environ. 2004, 27, 876–886. [Google Scholar] [CrossRef]
- Thirukkumaran, C.M.; Parkinson, D. Microbial respiration, biomass, metabolic quotient and litter decomposition in a lodgepole pine forest floor amended with nitrogen and phosphorous fertilizers. Soil Biol. Biochem. 2000, 32, 59–66. [Google Scholar] [CrossRef]
- Van Gestel, M.; Merckx, R.; Vlassak, K. Microbial biomass responses to soil drying and rewetting: The fate of fast-and slow-growing microorganisms in soils from different climates. Soil Biol. Biochem. 1993, 25, 109–123. [Google Scholar] [CrossRef]
- Borken, W.; Davidson, E.; Savage, K.; Gaudinski, J.; Trumbore, S.E. Drying and wetting effects on carbon dioxide release from organic horizons. Soil Sci. Soc. Am. J. 2003, 67, 1888–1896. [Google Scholar] [CrossRef]
- Will, R.E.; Fox, T.; Akers, M.; Domec, J.C.; Gonzalez-Benecke, C.; Jokela, E.J.; McGuire, M.A. A range-wide experiment to investigate nutrient and soil moisture interactions in loblolly pine plantations. Forests 2015, 6, 2014–2028. [Google Scholar] [CrossRef]
- NCDC.NOAA.gov. Monthly Summaries. Lynchburg, VA, USA. Available online: http://www.ncdc.noaa.gov/cdo-web/search (accessed on 27 November 2013).
- Soil Series Classification Database, USDA Natural Resources Conservation Service. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/survey/class/?cid=nrcs142p2_053583 (accessed on 27 November 2015).
- Heim, B.C.; Seiler, J.R.; Strahm, B.D. Root nonstructural carbohydrates and their relationship with autotrophic respiration of loblolly pine (Pinus taeda L.). Commun. Soil Sci. Plan. 2015, 46, 888–896. [Google Scholar] [CrossRef]
- Saxton, K.; Rawls, W.J.; Romberger, J.; Papendick, R. Estimating generalized soil-water characteristics from texture. Soil Sci. Soc. Am. J. 1986, 50, 1031–1036. [Google Scholar] [CrossRef]
- Van Genuchten, M.T.; Leij, F.J.; Yates, S.R. The RETC Code for Quantifying the Hydraulic Functions of Unsaturated Soils; U.S. Environmental Protection Agency: Washington, DC, USA, 1991.
- Pikul, J., Jr. Soil water, gravimetric measurement. In Encyclopedia of Water Science; Stewart, B.A., Howell, T., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 2003; pp. 879–881. [Google Scholar]
- Gholz, H.L.; Wedin, D.A.; Smitherman, S.M.; Harmon, M.E.; Parton, W.J. Long-term dynamics of pine and hardwood litter in contrasting environments: Toward a global model of decomposition. Glob. Chang. Biol. 2000, 6, 751–765. [Google Scholar] [CrossRef]
- Fierer, N.; Allen, A.S.; Schimel, J.P.; Holden, P.A. Controls on microbial CO2 production: A comparison of surface and subsurface soil horizons. Glob. Chang. Biol. 2003, 9, 1322–1332. [Google Scholar] [CrossRef]
- Schnürer, J.; Clarholm, M.; Boström, S.; Rosswall, T. Effects of moisture on soil microorganisms and nematodes: A field experiment. Microbial Ecol. 1986, 12, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Brookes, P. The proportional mineralisation of microbial biomass and organic matter caused by air-drying and rewetting of a grassland soil. Soil Biol. Biochem. 2005, 37, 507–515. [Google Scholar] [CrossRef]
- Borken, W.; Matzner, E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob. Chang. Biol. 2009, 15, 808–824. [Google Scholar] [CrossRef]
- Kieft, T.L. Microbial biomass response to a rapid increase in water potential when dry soil is wetted. Soil Biol. Biochem. 1987, 19, 119–126. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil. Soil Sci. Soc. Am. J. 2003, 67, 798–805. [Google Scholar] [CrossRef]
- Unger, S.; Máguas, C.; Pereira, J.S.; David, T.S.; Werner, C. The influence of precipitation pulses on soil respiration—Assessing the “Birch effect” by stable carbon isotopes. Soil Biol. Biochem. 2010, 42, 1800–1810. [Google Scholar] [CrossRef]
- Wu, H.-J.; Lee, X. Short-term effects of rain on soil respiration in two New England forests. Plant Soil 2011, 338, 329–342. [Google Scholar] [CrossRef]
- Griffiths, E.; Birch, H. Microbiological changes in freshly moistened soil. Nature 1961, 189, 424. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.-L.; Wang, H.; Guo, C.; Bao, W. Rainfall pulse primarily drives litterfall respiration and its contribution to soil respiration in a young exotic pine plantation in subtropical China. Can. J. For. Res. 2012, 42, 657–666. [Google Scholar] [CrossRef]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Herinková, J.; Cajthaml, T.; Baldrian, P. Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil. Soil Biol. Biochem. 2008, 40, 2068–2075. [Google Scholar] [CrossRef]
- Butterly, C.; Bünemann, E.; McNeill, A.; Baldock, J.; Marschner, P. Carbon pulses but not phosphorus pulses are related to decreases in microbial biomass during repeated drying and rewetting of soils. Soil Biol. Biochem. 2009, 41, 1406–1416. [Google Scholar] [CrossRef]
- Foster, R. Microenvironments of soil microorganisms. Biol. Fertil. Soils 1988, 6, 189–203. [Google Scholar] [CrossRef]
- Lado-Monserrat, L.; Lull, C.; Bautista, I.; Lidón, A.; Herrera, R. Soil moisture increment as a controlling variable of the “Birch effect”: Interactions with the pre-wetting soil moisture and litter addition. Plant Soil 2014, 379, 21–34. [Google Scholar] [CrossRef]
- Tyree, M.C.; Seiler, J.R.; Maier, C.A. Contrasting genotypes, soil amendments, and their interactive effects on short-term total soil CO2 efflux in a 3-year-old Pinus taeda L. plantation. Soil Biol. Biochem. 2014, 69, 93–100. [Google Scholar] [CrossRef]
- Lilleskov, E.A.; Fahey, T.J.; Horton, T.R.; Lovett, G.M. Belowground ectomycorrhizal fungal community change over a nitrogen deposition gradient in Alaska. Ecology 2002, 83, 104–115. [Google Scholar] [CrossRef]
- Nilsson, L.O.; Wallander, H. Production of external mycelium by ectomycorrhizal fungi in a Norway spruce forest was reduced in response to nitrogen fertilization. New Phytol. 2003, 158, 409–416. [Google Scholar] [CrossRef]
- Bittman, S.; Forge, T.A.; Kowalenko, C.G. Responses of the bacterial and fungal biomass in a grassland soil to multi-year applications of dairy manure slurry and fertilizer. Soil Biol. Biochem. 2005, 37, 613–623. [Google Scholar] [CrossRef]
- Burton, A.J.; Pregitzer, K.S.; Crawford, J.N.; Zogg, G.P.; Zak, D.R. Simulated chronic NO3− deposition reduces soil respiration in northern hardwood forests. Glob. Chang. Biol. 2004, 10, 1080–1091. [Google Scholar] [CrossRef]
- Swanston, C.; Homann, P.S.; Caldwell, B.A.; Myrold, D.D.; Ganio, L.; Sollins, P. Long-term effects of elevated nitrogen on forest soil organic matter stability. Biogeochemistry 2004, 70, 229–252. [Google Scholar] [CrossRef]
- Olsson, P.; Linder, S.; Giesler, R.; Högberg, P. Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration. Glob. Chang. Biol. 2005, 11, 1745–1753. [Google Scholar] [CrossRef]
- Gallardo, A.; Schlesinger, W.H. Factors limiting microbial biomass in the mineral soil and forest floor of a warm-temperate forest. Soil Biol. Biochem. 1994, 26, 1409–1415. [Google Scholar] [CrossRef]
- Lagomarsino, A.; Moscatelli, M.C.; De Angelis, P.; Grego, S. Labile substrates quality as the main driving force of microbial mineralization activity in a poplar plantation soil under elevated CO2 and nitrogen fertilization. Sci. Total Environ. 2006, 372, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Gartner, T.B. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef] [PubMed]
- Ajwa, H.; Dell, C.; Rice, C. Changes in enzyme activities and microbial biomass of tallgrass prairie soil as related to burning and nitrogen fertilization. Soil Biol. Biochem. 1999, 31, 769–777. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; Burton, S.D.; Firestone, M.K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology 2011, 92, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Han, X.; Wang, Y.; Han, M.; Shi, H.; Liu, N.; Bai, H. Influence of long-term fertilization on soil microbial biomass, dehydrogenase activity, and bacterial and fungal community structure in a brown soil of northeast China. Ann. Microbiol. 2015, 65, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Nohrstedt, H.-Ö.; Arnebrant, K.; Bååth, E.; Söderström, B. Changes in carbon content, respiration rate, ATP content, and microbial biomass in nitrogen-fertilized pine forest soils in Sweden. Can. J. For. Res. 1989, 19, 323–328. [Google Scholar] [CrossRef]
- Cusack, D.F.; Torn, M.S.; McDowell, W.H.; Silver, W.L. The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils. Glob. Chang. Biol. 2010, 16, 2555–2572. [Google Scholar] [CrossRef]
- Orchard, V.A.; Cook, F. Relationship between soil respiration and soil moisture. Soil Biol. Biochem. 1983, 15, 447–453. [Google Scholar] [CrossRef]
- Carbone, M.S.; Still, C.J.; Ambrose, A.R.; Dawson, T.E.; Williams, A.P.; Boot, C.M.; Schaeffer, S.M.; Schimel, J.P. Seasonal and episodic moisture controls on plant and microbial contributions to soil respiration. Oecologia 2011, 167, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wan, S.; Su, B.; Hui, D.; Luo, Y. Response of soil CO2 efflux to water manipulation in a tallgrass prairie ecosystem. Plant Soil 2002, 240, 213–223. [Google Scholar] [CrossRef]
- McIntyre, R.E.; Adams, M.A.; Ford, D.J.; Grierson, P.F. Rewetting and litter addition influence mineralisation and microbial communities in soils from a semi-arid intermittent stream. Soil Biol. Biochem. 2009, 41, 92–101. [Google Scholar] [CrossRef]
Figure 1.
Change in (A) mineral soil heterotrophic respiration (RHM) and water content of the root-free mineral soil (B) over the duration of the dry down (days) in unfertilized (closed symbols) and fertilized samples (open symbols). The line represents the mean fit of the exponential decay function. Data represent actual values for each replicated treatment.
Figure 1.
Change in (A) mineral soil heterotrophic respiration (RHM) and water content of the root-free mineral soil (B) over the duration of the dry down (days) in unfertilized (closed symbols) and fertilized samples (open symbols). The line represents the mean fit of the exponential decay function. Data represent actual values for each replicated treatment.

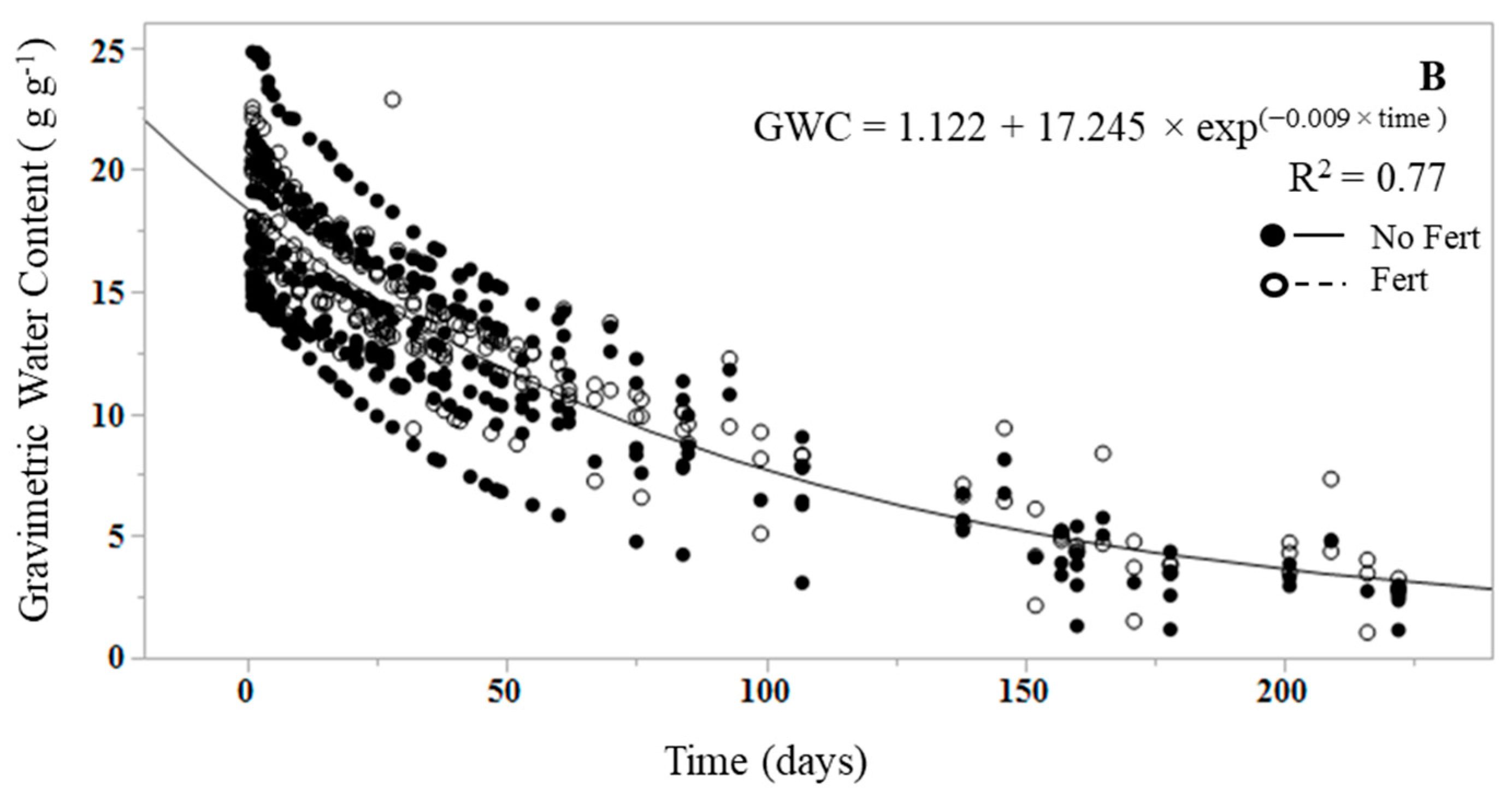
Figure 2.
Change in (A) litter heterotrophic respiration (RHL) and litter water content (B) following the initial wetting event over the duration of the dry down (days) in unfertilized (closed symbols) and fertilized samples (open symbols). The line represents the fit of the exponential decay function. Data represent actual values for each replicated treatment.
Figure 2.
Change in (A) litter heterotrophic respiration (RHL) and litter water content (B) following the initial wetting event over the duration of the dry down (days) in unfertilized (closed symbols) and fertilized samples (open symbols). The line represents the fit of the exponential decay function. Data represent actual values for each replicated treatment.
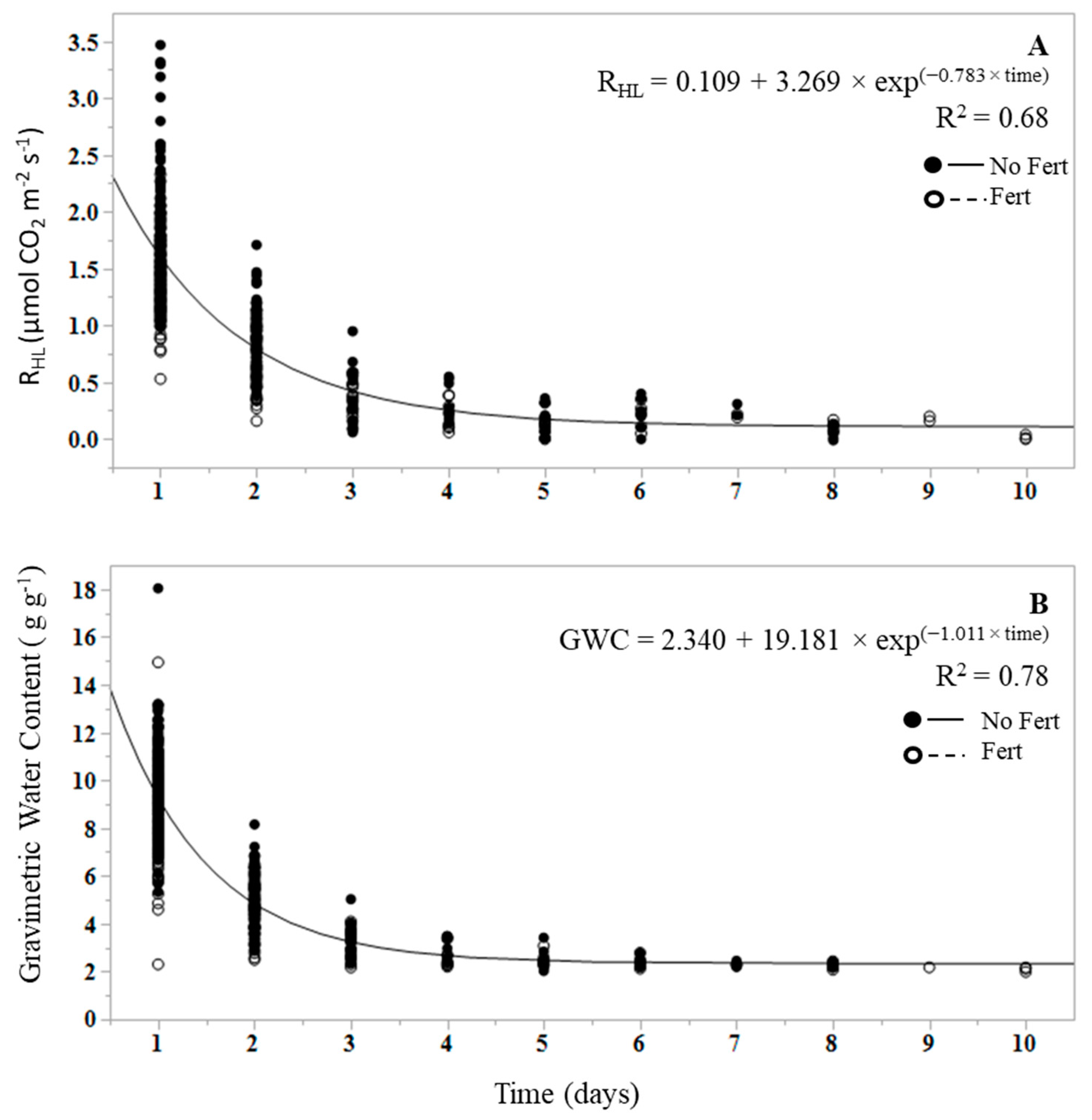
Figure 3.
Relationships between (A) mineral soil heterotrophic respiration (RHM) and soil gravimetric water content (GWC), and (B) leaf litter heterotrophic respiration (RHL) and leaf litter gravimetric water content (GWC) in unfertilized (closed symbols, solid line) and fertilized samples (open symbols, dashed line).
Figure 3.
Relationships between (A) mineral soil heterotrophic respiration (RHM) and soil gravimetric water content (GWC), and (B) leaf litter heterotrophic respiration (RHL) and leaf litter gravimetric water content (GWC) in unfertilized (closed symbols, solid line) and fertilized samples (open symbols, dashed line).
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McElligott, K.M.; Seiler, J.R.; Strahm, B.D. The Impact of Water Content on Sources of Heterotrophic Soil Respiration. Forests 2017, 8, 299. https://doi.org/10.3390/f8080299
AMA Style
McElligott KM, Seiler JR, Strahm BD. The Impact of Water Content on Sources of Heterotrophic Soil Respiration. Forests. 2017; 8(8):299. https://doi.org/10.3390/f8080299
Chicago/Turabian StyleMcElligott, Kristin M., John R. Seiler, and Brian D. Strahm. 2017. "The Impact of Water Content on Sources of Heterotrophic Soil Respiration" Forests 8, no. 8: 299. https://doi.org/10.3390/f8080299
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.