Effective Seed Dispersal and Fecundity Variation in a Small and Marginal Population of Pinus pinaster Ait. Growing in a Harsh Environment: Implications for Conservation of Forest Genetic Resources


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
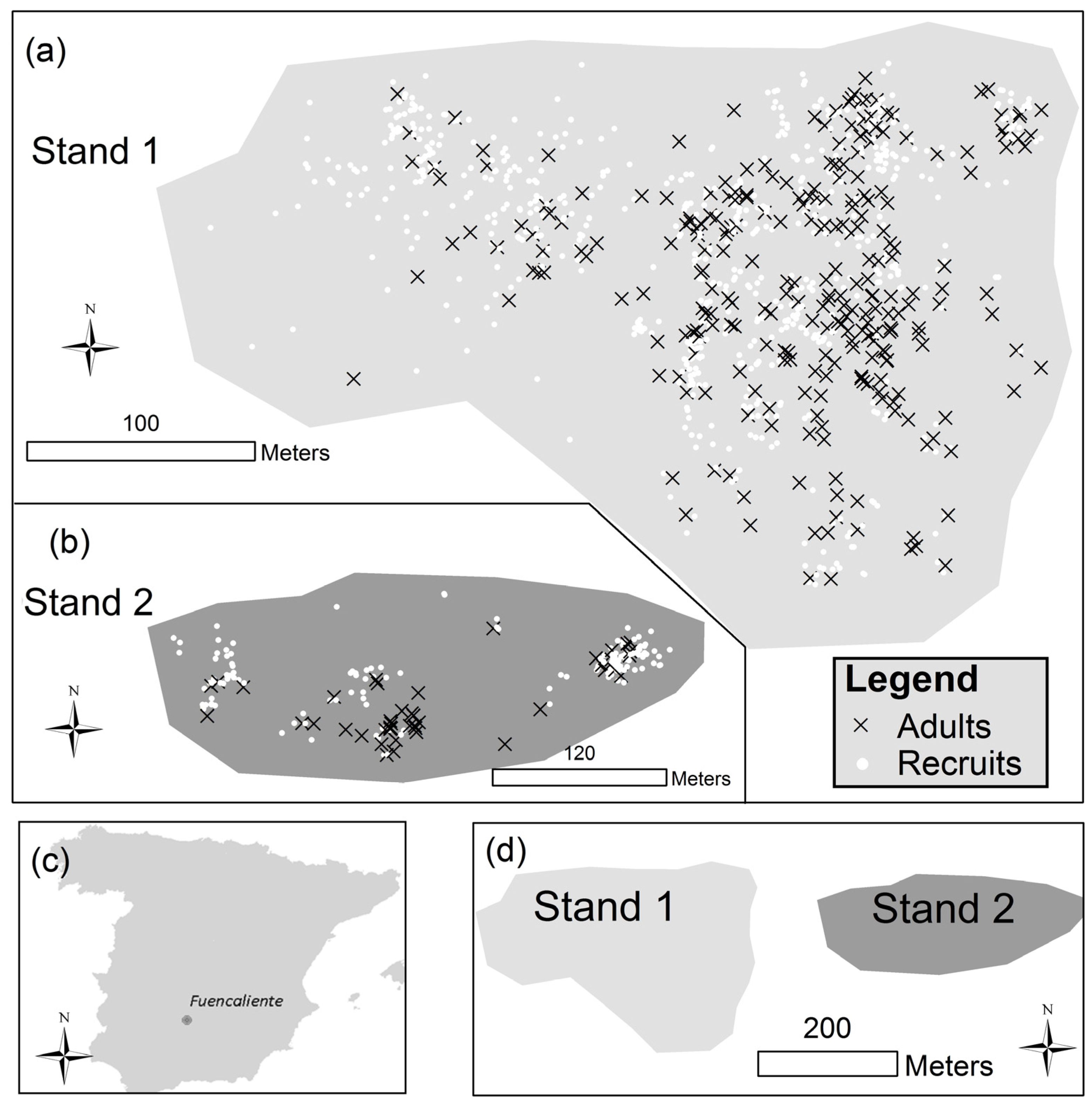
2.2. Forest Inventory
- -
- Adults: trees with at least one cone on their crown
- -
- Recruits: the rest of the individuals
2.3. Modeling Effective Dispersal
2.4. Models for Fecundity
- Tree size covariates (basal area, BA, and tree height, H). From an ecological perspective, these models inherently assume that reproductive success is a linear function of tree size.
- Cone number covariates (total cones, Tc, open cones, Oc, and serotinous cones, Sc). Models using covariates related to the cone number assume that the number of seeds and number of recruits produced by adults is linearly related.
- Spatial covariates (the east-west, Xco, and the north-south, Yco, coordinates of adults). Inherently, these models assume the reproductive success has some relation to the microhabitat conditions surrounding the adult tree.
2.5. Parameter Estimation
2.6. Model Comparison
- The difference between the AICc for the k-th model and the one with the smallest AICc (AICcmin):
- The correlation coefficient between observed and predicted counts in quadrats of the k-th model
3. Results
3.1. Descriptive Results
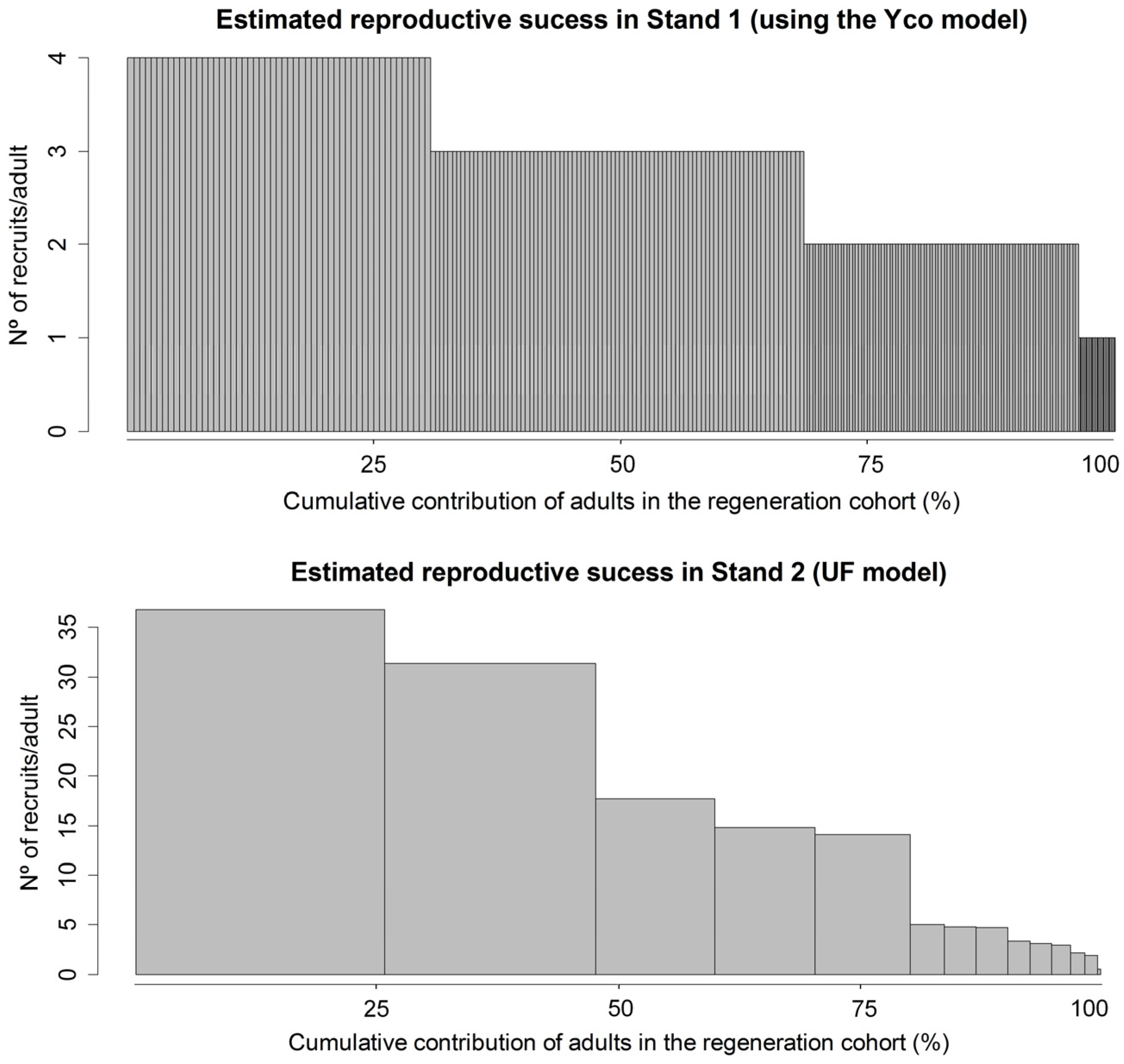
3.2. Choosing the Best Model for Fecundity
3.3. Dispersal and Fecundity Parameter Estimates
4. Discussion
4.1. Fecundity Dynamics
4.2. Effective Dispersal Distances
4.3. Management Implications
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganopoulos, I.; Aravanopoulos, F.A.; Argiriou, A.; Kalivas, A.; Tsaftaris, A. Is the genetic diversity of small scattered forest tree populations at the southern limits of their range more prone to stochastic events? A wild cherry case study by microsatellite-based markers. Tree Genet. Genom. 2011, 7, 1299–1313. [Google Scholar] [CrossRef]
- Lowe, A.J.; Boshier, D.; Ward, M.; Bacles, C.F.E.; Navarro, C. Genetic resource impacts of habitat loss and degradation; reconciling empirical evidence and predicted theory for neotropical trees. Heredity 2005, 95, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Ribbens, E.; Silander, J.A.; Pacala, S.W. Seedling recruitment in forests—Calibrating models to predict patterns of tree seedling dispersion. Ecology 1994, 75, 1794–1806. [Google Scholar] [CrossRef]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef]
- Moran, E.V.; Clark, J.S. Estimating seed and pollen movement in a monoecious plant: A hierarchical Bayesian approach integrating genetic and ecological data. Mol. Ecol. 2011, 20, 1248–1262. [Google Scholar] [CrossRef] [PubMed]
- Petit, R.J.; Bialozyt, R.; Garnier-Géré, P.; Hampe, A. Ecology and genetics of tree invasions: From recent introductions to Quaternary migrations. For. Ecol. Manag. 2004, 197, 117–137. [Google Scholar] [CrossRef]
- Gerzabek, G.; Oddou-Muratorio, S.; Hampe, A. Temporal change and determinants of maternal reproductive success in an expanding oak forest stand. J. Ecol. 2017, 105, 39–48. [Google Scholar] [CrossRef]
- Hampe, A.; Arroyo, J. Recruitment and regeneration in populations of an endangered South Iberian Tertiary relict tree. Biol. Conserv. 2002, 107, 263–271. [Google Scholar] [CrossRef]
- Nathan, R.; Safriel, U.N.; Noy-Meir, I.; Schiller, G. Spatiotemporal variation in seed dispersal and recruitment near and far from Pinus halepensis trees. Ecology 2000, 81, 2156–2169. [Google Scholar] [CrossRef]
- Lefèvre, F.; Boivin, T.; Bontemps, A.; Courbet, F.; Davi, H.; Durand-Gillmann, M.; Fady, B.; Gauzere, J.; Gidoin, C.; Karam, M.-J. Considering evolutionary processes in adaptive forestry. Ann. For. Sci. 2014, 71, 723–739. [Google Scholar] [CrossRef]
- Alía, R.; Martín, S.; de Miguel, J.; Galera, R.; Agúndez, D.; Gordo, J.; Salvador, L.; Catalán, G.; Gil, L. Las Regiones de Procedencia de Pinus pinaster Aiton en España; Organismo Autónomo Parques Nacionales-ETSI de Montes: Madrid, Spain, 1996. [Google Scholar]
- Gil, L. Consideraciones históricas sobre Pinus pinaster Aiton en el paisaje vegetal de la Península Ibérica. Estudios Geogr. 1991, 202, 5–27. [Google Scholar]
- Díaz-Fernández, P.M. Relations between modern pollen rain and Mediterranean vegetation in Sierra Madrona (Spain). Rev. Palaeobot. Palynol. 1994, 82, 113–125. [Google Scholar] [CrossRef]
- Rubiales, J.M.; García-Amorena, I.; Álvarez, S.G.; Morla, C. Anthracological evidence suggests naturalness of Pinus pinaster in inland southwestern Iberia. Plant Ecol. 2009, 200, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Zapata, B.; Gil-García, M.J.; de Bustamante, I. Paleoenvironmental reconstruction of Las Tablas de Daimiel and its evolution during the Quaternary period. In Ecology of Threatened Semi-Arid Wetlands: Long-Term Research in Las Tablas de Daimiel; Sánchez-Carrillo, S., Angeler, D.G., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2010; pp. 23–43. [Google Scholar]
- Valbuena-Carabaña, M.; López de Heredia, U.; Fuentes-Utrilla, P.; González-Doncel, I.; Gil, L. Historical and recent changes in the Spanish forests: A socio-economic process. Rev. Palaeobot. Palinol. 2010, 162, 492–506. [Google Scholar] [CrossRef]
- Charco, J. Evolución Histórica de los Bosques en Sierra Madrona y Valle de Alcudia (Ciudad Real) y Dinámica del Pinar Relicto de Navalmanzano. Ph.D. Thesis, E.T.S.I. de Montes, Universidad Politécnica de Madrid, Madrid, Spain, 2016. [Google Scholar]
- Chauchard, S.; Carcaillet, C.; Guibal, F. Patterns of land-use abandonment control tree-recruitment and forest dynamics in Mediterranean mountains. Ecosystems 2007, 10, 936–948. [Google Scholar] [CrossRef]
- González-Martínez, S.; Salvador, L.; Agúndez, D.; Alía, R.; Gil, L. Geographical variation of gene diversity of Pinus pinaster Ait. in the Iberian Peninsula. In Genetic Response of Forest Systems to Changing Environmental Conditions; Muller-Starck, G., Schubert, R., Eds.; Springer: Dordrecht, the Netherlands, 2001; pp. 161–171. [Google Scholar]
- Salvador, L.; Alía, R.; Agúndez, D.; Gil, L. Genetic variation and migration pathways of maritime pine (Pinus pinaster Ait) in the Iberian Peninsula. Theor. Appl. Genet. 2000, 100, 89–95. [Google Scholar] [CrossRef]
- González-Martínez, S.G.; Gil, L.; Alía, R. Genetic diversity estimates of Pinus pinaster in the Iberian Peninsula: A comparison of allozymes and quantitative traits. For. Syst. 2005, 14, 3–12. [Google Scholar] [CrossRef]
- Ramírez-Valiente, J.A.; Robledo-Arnuncio, J.J. Adaptive consequences of human-mediated introgression for indigenous tree species: The case of a relict Pinus pinaster population. Tree Physiol. 2014, 34, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Unger, G.M.; Vendramin, G.G.; Robledo-Arnuncio, J.J. Estimating exotic gene flow into native pine stands: Zygotic vs. gametic components. Mol. Ecol. 2014, 23, 5435–5447. [Google Scholar] [CrossRef] [PubMed]
- Unger, G.M.; Heuertz, M.; Vendramin, G.G.; Robledo-Arnuncio, J.J. Assessing early fitness consequences of exotic gene flow in the wild: A field study with Iberian pine relicts. Evol. Appl. 2016, 9, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Charco, J.; Perea, R.; Gil, L.; Nanos, N. Impact of deer rubbing on pine forests: Implications for conservation and management of Pinus pinaster populations. Eur. J. For. Res. 2016, 135, 719–729. [Google Scholar] [CrossRef]
- García-Rayego, J.L. Modelados de detalle en roquedos cuarcíticos de áreas de montaña media apalachense de la Meseta sur y Sierra Morena oriental. Ería 2006, 71, 269–282. [Google Scholar]
- Clark, J.S.; Silman, M.; Kern, R.; Macklin, E.; HilleRisLambers, J. Seed dispersal near and far: Patterns across temperate and tropical forests. Ecology 1999, 80, 1475–1494. [Google Scholar] [CrossRef]
- Nanos, N.; Larson, K.; Millerón, M.; Sjöstedt-deLuna, S. Inverse modeling for effective dispersal: Do we need tree size to estimate fecundity? Ecol. Model. 2010, 221, 2415–2424. [Google Scholar] [CrossRef]
- Dempster, A.P.; Laird, N.M.; Rubin, D.B. Maximum likelihood from incomplete data via EM algorithm. J. R. Stat. Soc. B 1977, 39, 1–38. [Google Scholar]
- R Development Core Team. 2008 R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.R-project.org (accessed on 12 January 2017).
- Muller-Landau, H.C.; Wright, S.J.; Calderón, O.; Condit, R.; Hubbell, S.P. Interspecific variation in primary seed dispersal in a tropical forest. J. Ecol. 2008, 96, 653–667. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of south-eastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- González-Martínez, S.C.; Burczyk, J.; Nathan, R.; Nanos, N.; Gil, L.; Alía, R. Effective gene dispersal and female reproductive success in Mediterranean maritime pine (Pinus pinaster Aiton). Mol. Ecol. 2006, 15, 4577–4588. [Google Scholar] [CrossRef] [PubMed]
- De Heredia, U.L.; Nanos, N.; García-del-Rey, E.; Guzmán, P.; López, R.; Venturas, M.; Gil, L. High seed dispersal ability of Pinus canariensis in stands of contrasting density inferred from genotypic data. For. Syst. 2015, 24, 015. [Google Scholar] [CrossRef]
- Millerón, M.; de Heredia, U.L.; Lorenzo, Z.; Perea, R.; Dounavi, A.; Alonso, J.; Nanos, N. Effect of canopy closure on pollen dispersal in a wind-pollinated species (Fagus sylvatica L.). Plant Ecol. 2012, 213, 1715–1728. [Google Scholar] [CrossRef]
- Millerón, M.; Lopez de Heredia, U.; Lorenzo, Z.; Alonso, J.; Dounavi, A.; Gil, L.; Nanos, N. Assessment of spatial discordance of primary and effective seed dispersal of European beech (Fagus sylvatica L.) by ecological and genetic methods. Mol. Ecol. 2013, 22, 1531–1545. [Google Scholar] [CrossRef] [PubMed]
- Juez, L.; González-Martínez, S.C.; Nanos, N.; de-Lucas, A.I.; Ordóñez, C.; del Peso, C.; Bravo, F. Can seed production and restricted dispersal limit recruitment in Pinus pinaster Aiton from the Spanish Northern Plateau? For. Ecol. Manag. 2014, 313, 329–339. [Google Scholar] [CrossRef]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Sagnard, F.; Pichot, C.; Dreyfus, P.; Jordano, P.; Fady, B. Modelling seed dispersal to predict seedling recruitment: Recolonization dynamics in a plantation forest. Ecol. Model. 2007, 203, 464–474. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Model | Model Name | Abbr. | Formula | Number of Parameters | |
|---|---|---|---|---|---|
| Stand 1 | Stand 2 | ||||
| Full model | Unrestricted Fecundity | UF | 268 | 44 | |
| Null model | Mean fecundity | MF | 1 | 1 | |
| Tree size covariates | Basal area | BA | 1 | 1 | |
| Height | H | 1 | 1 | ||
| Cone number covariates | Total cones | Tc | 1 | 1 | |
| Open cones | Oc | 1 | 1 | ||
| Serotinous cones | Sc | 1 | 1 | ||
| Spatial covariates | X coordinate of adult | Xco | 1 | 1 | |
| Y coordinate of adult | Yco | 1 | 1 | ||
| Dbase (cm) | DBH (cm) | Height (m) | Total Cones | ||||||
|---|---|---|---|---|---|---|---|---|---|
| St1 | St2 | St1 | St2 | St1 | St2 | St1 | St2 | ||
| Adults | Min. | 5.7 | 7.6 | 0.0 | 0.0 | 0.5 | 1.2 | 1.0 | 1.0 |
| 1st Qu. | 16.5 | 18.9 | 8.8 | 13.1 | 3.5 | 3.5 | 5.0 | 3.8 | |
| Median | 23.2 | 27.4 | 17.2 | 19.8 | 5.0 | 5.8 | 17.0 | 11.5 | |
| Mean | 27.3 | 28.1 | 19.2 | 20.1 | 5.4 | 5.4 | 58.2 | 33.9 | |
| 3rd Qu. | 35.7 | 35.5 | 28.7 | 27.5 | 7.0 | 6.8 | 67.5 | 52.0 | |
| Max. | 68.8 | 65.6 | 56.3 | 44.6 | 16.5 | 10.0 | 587.0 | 193.0 | |
| Recruits | Min. | 0.1 | 0.3 | 0.0 | 0.0 | 0.04 | 0.07 | 0.0 | 0.0 |
| 1st Qu. | 2.1 | 4.1 | 0.0 | 0.0 | 0.3 | 0.5 | 0.0 | 0.0 | |
| Median | 4.7 | 7.6 | 0.0 | 0.0 | 0.6 | 0.8 | 0.0 | 0.0 | |
| Mean | 5.7 | 7.7 | 0.5 | 0.3 | 0.8 | 0.8 | 0.0 | 0.0 | |
| 3rd Qu. | 8.2 | 11.3 | 0.0 | 0.0 | 1.0 | 1.0 | 0.0 | 0.0 | |
| Max. | 34.3 | 31.5 | 19.0 | 11.1 | 6.6 | 4.3 | 0.0 | 0.0 | |
| Model | Ln(L) | AICc | cor | ||
|---|---|---|---|---|---|
| Stand 1 | Yco | −2753.7 | 5513.4 | 0.0 | 0.20 |
| Tc | −2762.5 | 5531.0 | 17.6 | 0.20 | |
| Sc | −2764.1 | 5534.3 | 20.9 | 0.19 | |
| MF-null | −2771.3 | 5548.7 | 35.3 | 0.18 | |
| BA | −2823.4 | 5652.8 | Nc | 0.18 | |
| UF | −2431.8 | 5749.5 | Nc | 0.30 | |
| H | −2883.6 | 5773.2 | Nc | 0.15 | |
| Xco | −2911.9 | 5829.8 | Nc | 0.16 | |
| Oc | −2982.4 | 5970.8 | Nc | 0.16 | |
| Stand 2 | UF | −546.2 | 1220.9 | 0.0 | 0.34 |
| Yco | −628.1 | 1262.3 | 41.4 | 0.22 | |
| Xco | −642.0 | 1290.2 | 69.3 | 0.20 | |
| MF-null | −642.8 | 1291.8 | 70.9 | 0.18 | |
| Tc | −661.8 | 1329.8 | Nc | 0.18 | |
| Sc | −663.8 | 1333.8 | Nc | 0.18 | |
| Oc | −667.5 | 1341.1 | Nc | 0.17 | |
| BA | −691.0 | 1388.1 | Nc | 0.11 | |
| H | −692.0 | 1390.2 | Nc | 0.12 |
| Model | Median | Mean | Mode | ||||
|---|---|---|---|---|---|---|---|
| Stand 1 | Yco | 2.66 | 1.02 | 0.01 | 14.30 | 24.05 | 5.05 |
| Tc | 2.87 | 0.77 | 0.04 | 17.67 | 23.74 | 9.79 | |
| Sc | 2.95 | 0.78 | 0.08 | 19.11 | 25.90 | 10.40 | |
| MF-null | 2.57 | 0.96 | 2.58 | 13.07 | 20.71 | 5.20 | |
| BA | 2.71 | 0.86 | 35.33 | Nc | Nc | Nc | |
| UF | 2.26 | 0.87 | - | Nc | Nc | Nc | |
| H | 2.72 | 0.99 | 0.00 | Nc | Nc | Nc | |
| Xco | 2.77 | 1.16 | 0.01 | Nc | Nc | Nc | |
| Oc | 4.25 | 1.45 | 0.14 | Nc | Nc | Nc | |
| Stand 2 | UF | 2.27 | 0.76 | - | 9.69 | 12.94 | 5.43 |
| Yco | 2.31 | 0.89 | 0.02 | 10.07 | 14.97 | 4.56 | |
| Xco | 2.32 | 0.92 | 0.004 | 10.18 | 15.54 | 4.36 | |
| MF-null | 2.32 | 0.91 | 3.32 | 10.18 | 15.40 | 4.45 | |
| Tc | 2.67 | 0.80 | 0.10 | Nc | Nc | Nc | |
| Sc | 2.67 | 0.80 | 0.24 | Nc | Nc | Nc | |
| Oc | 2.69 | 0.81 | 0.16 | Nc | Nc | Nc | |
| BA | 2.46 | 0.86 | 44.50 | Nc | Nc | Nc | |
| H | 2.38 | 0.97 | 0.01 | Nc | Nc | Nc |
| Model | Ave | Min | Max | Var | Sum | |
|---|---|---|---|---|---|---|
| Stand 1 | Yco | 2.6 | 0.1 | 4.3 | 0.9 | 699 |
| Tc | 2.5 | 0.04 | 26.0 | 18.3 | 692 | |
| Sc | 2.5 | 0.0 | 47.6 | 24.3 | 693 | |
| MF-null | 2.5 | 2.5 | 2.5 | 0.0 | 693 | |
| Stand 2 | UF | 3.2 | 0.0 | 36.8 | 63.1 | 143 |
| Yco | 3.3 | 2.5 | 4.5 | 0.3 | 145 | |
| Xco | 3.3 | 2.5 | 4.0 | 0.2 | 146 | |
| MF-null | 3.3 | 3.3 | 3.3 | 0.0 | 146 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charco, J.; Venturas, M.; Gil, L.; Nanos, N. Effective Seed Dispersal and Fecundity Variation in a Small and Marginal Population of Pinus pinaster Ait. Growing in a Harsh Environment: Implications for Conservation of Forest Genetic Resources. Forests 2017, 8, 312. https://doi.org/10.3390/f8090312
Charco J, Venturas M, Gil L, Nanos N. Effective Seed Dispersal and Fecundity Variation in a Small and Marginal Population of Pinus pinaster Ait. Growing in a Harsh Environment: Implications for Conservation of Forest Genetic Resources. Forests. 2017; 8(9):312. https://doi.org/10.3390/f8090312
Chicago/Turabian StyleCharco, Jesús, Martin Venturas, Luis Gil, and Nikos Nanos. 2017. "Effective Seed Dispersal and Fecundity Variation in a Small and Marginal Population of Pinus pinaster Ait. Growing in a Harsh Environment: Implications for Conservation of Forest Genetic Resources" Forests 8, no. 9: 312. https://doi.org/10.3390/f8090312