
Differential Responses in Non-structural Carbohydrates of Machilus ichangensis Rehd. et Wils. and Taxus wallichiana Zucc. Var. chinensis (Pilg.) Florin Seedlings to Elevated Ozone

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Ozone Fumigation Treatment
2.3. Measurements
2.4. Statistical Analysis
3. Results
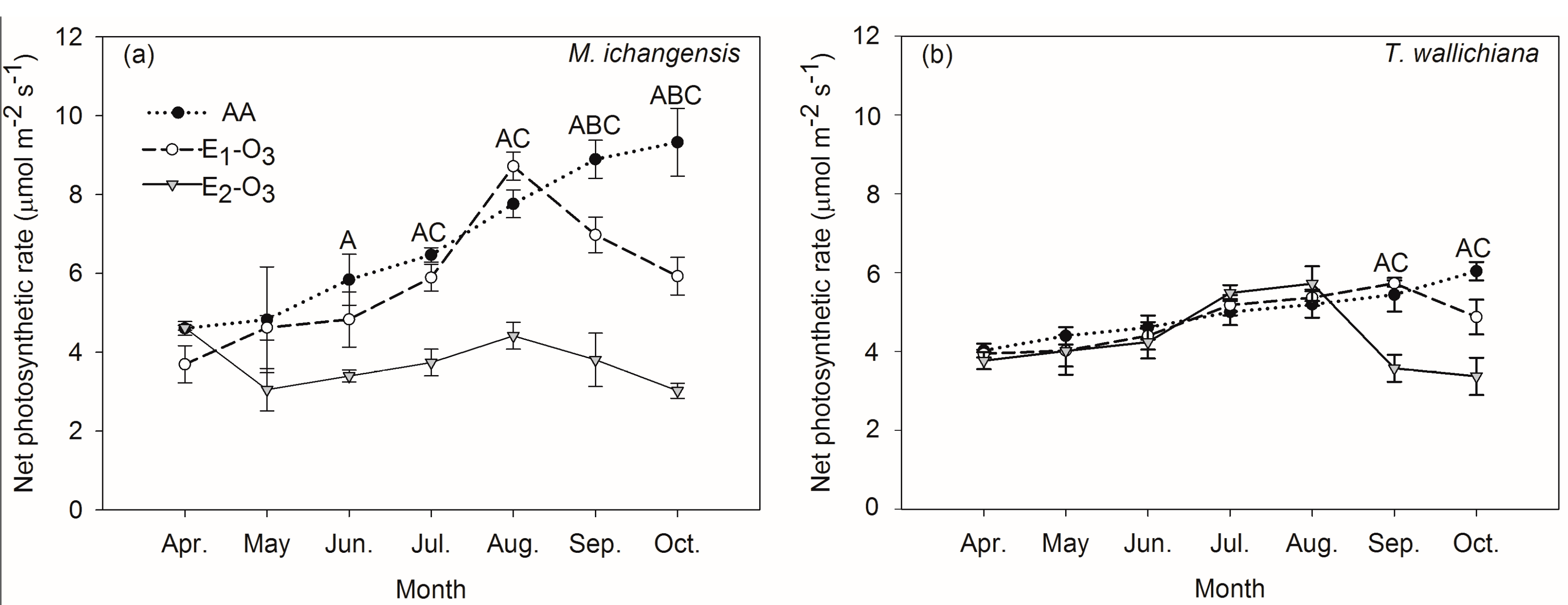
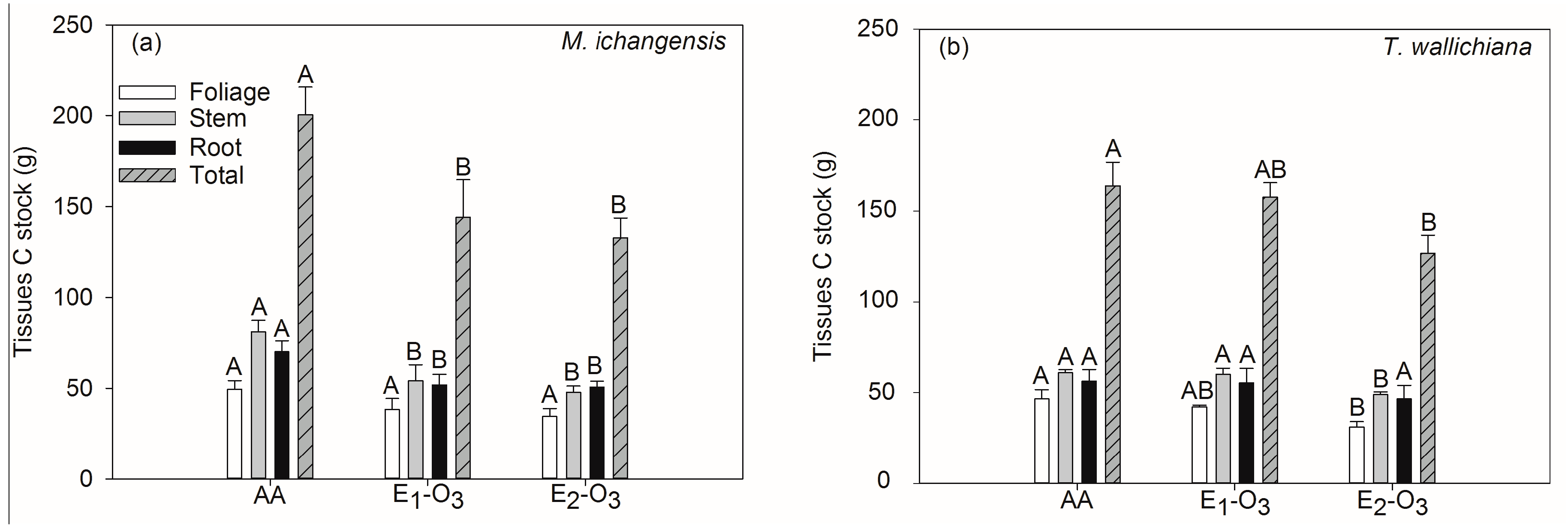
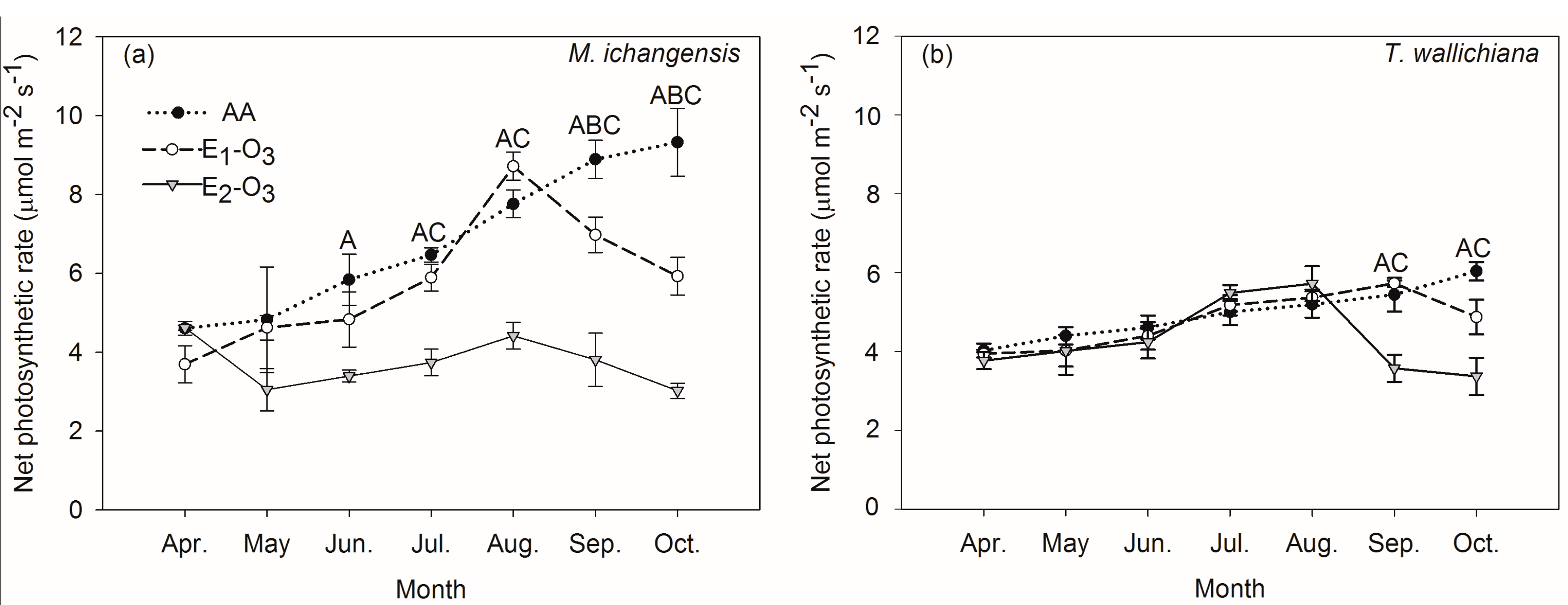
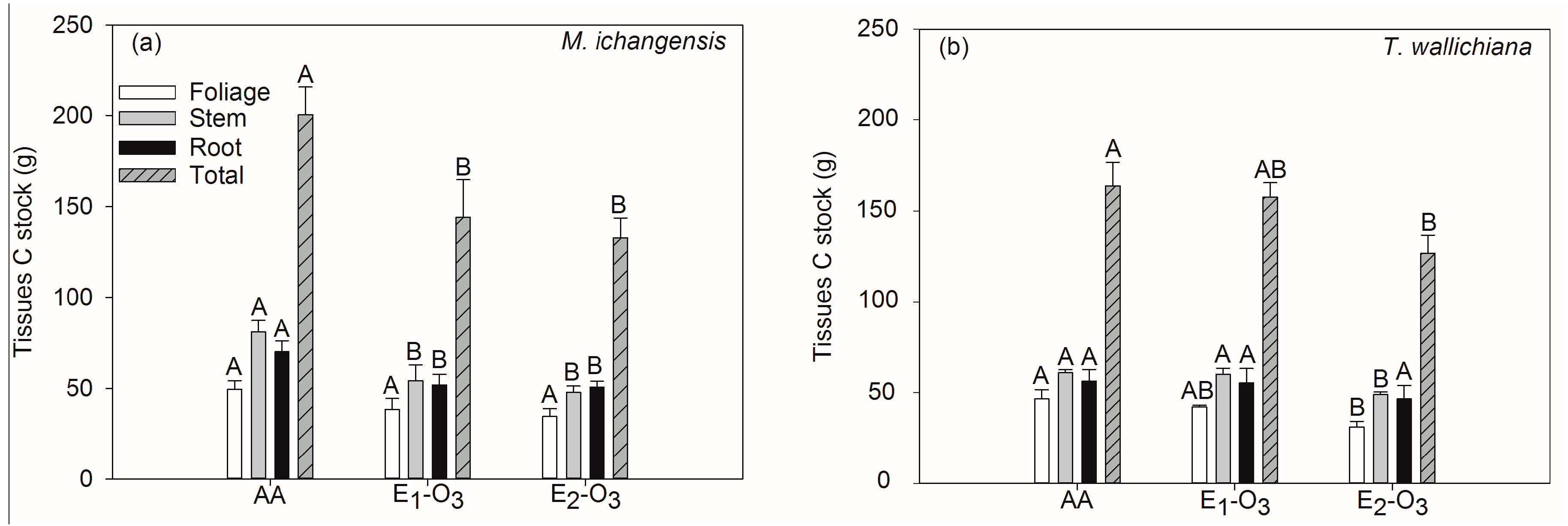
3.1. Effects of Elevated Ozone on Net Photosynthetic Rate and Carbon Stock
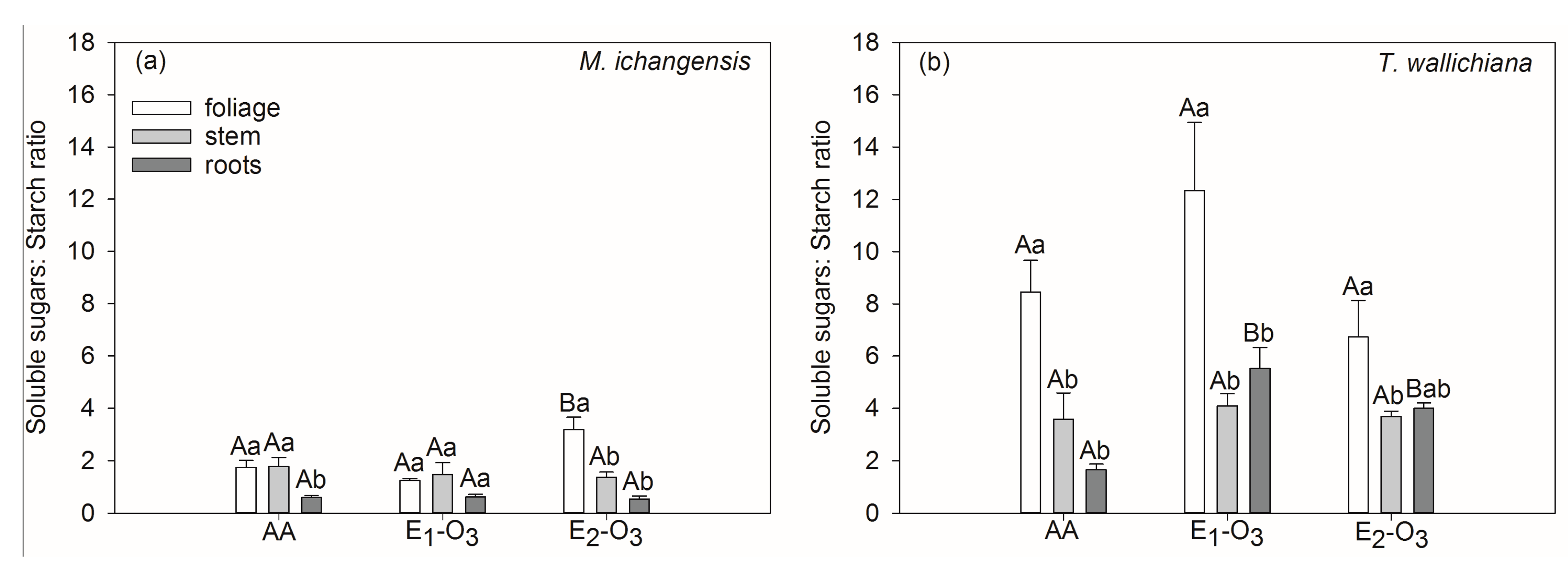
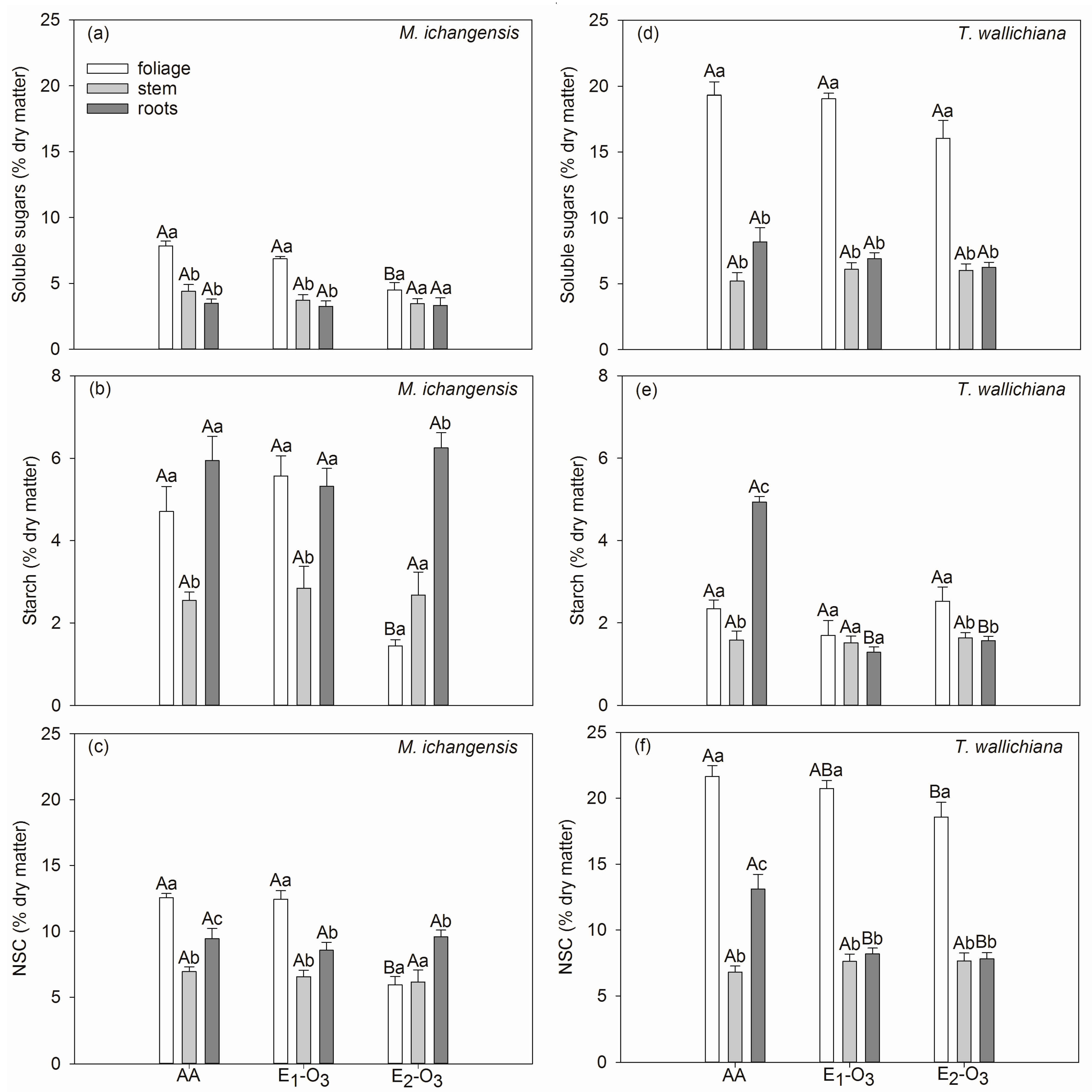
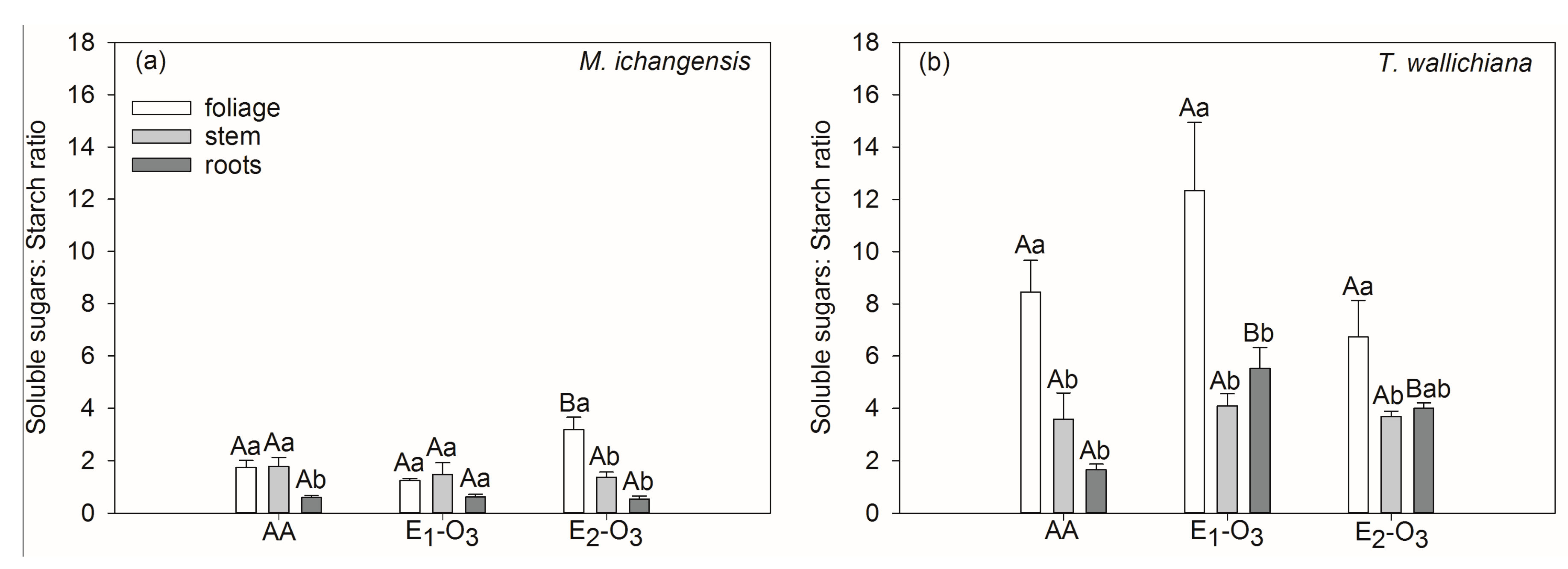
3.2. Effects of Elevated O3 on Non-Structural Carbohydrates of Different Seedling Tissues
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feng, Z.; Pang, J.; Kobayashi, K.; Zhu, J.; Ort, D.R. Differential responses in two varieties of winter wheat to elevated ozone concentration under fully open-air field conditions. Glob. Chang. Biol. 2011, 17, 580–591. [Google Scholar] [CrossRef]
- Pacifico, F.; Folberth, G.A.; Sitch, S.; Haywood, J.M.; Rizzo, L.V.; Malavelle, F.F.; Artaxo, P. Biomass burning related ozone damage on vegetation over the Amazon forest: A model sensitivity study. Atmos. Chem. Phys. 2015, 15, 2791–2804. [Google Scholar] [CrossRef] [Green Version]
- Vingarzan, R. A review of surface O3 background levels and trends. Atmos. Environ. 2004, 38, 3431–3442. [Google Scholar] [CrossRef]
- Fowler, D.; Amann, M.; Anderson, R.; Ashmore, M.; Cox, P.; Depledge, M.; Derwent, D.; Grennfelt, P.; Hewitt, N.; Hov, O.; et al. Ground-Level Ozone in the 21st Century: Future Trends, Impacts and Policy Implications; Society Science Policy Report, 15/08; The Royal Society: London, UK, 2008; pp. 1–4. [Google Scholar]
- Stevenson, D.S.; Dentener, F.J.; Schultz, M.G.; Ellingsen, K.; van Noije, T.P.C.; Wild, O.; Zeng, G.; Amann, M.; Atherton, C.S.; Bell, N.; et al. Multimodel ensemble simulations of present-day and near-future tropospheric ozone. J. Geophys. Res. 2006, 111, D08301. [Google Scholar] [CrossRef]
- Avnery, S.; Mauzerall, D.L.; Liu, J.; Horowitz, L.W. Global crop yield reductions due to surface ozone exposure: 2. Year 2030 potential crop production losses and economic damage under two scenarios of O3 pollution. Atmos. Environ. 2011, 45, 2297–2309. [Google Scholar] [CrossRef]
- De Leeuw, F.A.A.M.; van Zantvoort, E.D.G. Mapping of exceedances of ozone critical levels for crops and forest trees in the Netherlands: Preliminary results. Environ. Pollut. 1997, 96, 89–98. [Google Scholar] [CrossRef]
- Zhang, Q.; He, K.; Huo, H. Cleaning China’s air. Nature 2012, 484, 161–162. [Google Scholar] [PubMed]
- Li, J.; Lu, K.; Lv, W.; Li, J.; Zhong, L.; Ou, Y.; Chen, D.; Huang, X.; Zhang, Y. Fast increasing of surface ozone concentrations in Pearl River Delta characterized by a regional air quality monitoring network during 2006–2011. J. Environ. Sci. 2014, 26, 23–36. [Google Scholar] [CrossRef]
- Yuan, X.; Calatayud, V.; Jiang, L.; Manning, W.J.; Hayes, F.; Tian, Y.; Feng, Z. Assessing the effects of ambient ozone in China on snap bean genotypes by using ethylenediurea (EDU). Environ. Pollut. 2015, 205, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Anger, A.; Dessens, O.; Xi, F.; Barker, T.; Wu, R. China’s air pollution reduction efforts may result in an increase in surface ozone levels in highly polluted areas. Ambio 2016, 45, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Curtis, P.S.; Wang, X. A meta-analysis of elevated CO2 effects on woody plant mass, form, and physiology. Oecologia 1998, 113, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Zak, D.R.; Holmes, W.E.; Pregitzer, K.S. Atmospheric CO2 and O3 alter the flow of 15N in developing forest ecosystems. Ecology 2007, 88, 2630–2639. [Google Scholar] [CrossRef] [PubMed]
- Karnosky, D.F.; Skelly, J.M.; Percy, K.E.; Chappelka, A.H. Perspectives regarding 50 years of research on effects of tropospheric ozone air pollution on US forests. Environ. Pollut. 2007, 147, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M.D.; Dickson, R.E.; Isebrands, J.G.; Karnosky, D.F. Carbon allocation and partitioning in aspen clones varying in sensitivity to tropospheric ozone. Tree Physiol. 1995, 15, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Battistelli, A.; Pasqualini, S.; Moscatello, S.; Ederli, L.; Proietti, S.; Antonielli, M. Effects of short-term ozone fumigation on carbohydrates in darkened tobacco leaves. Plant Physiol. Biochem. 2001, 39, 539–543. [Google Scholar] [CrossRef]
- Chen, Z.; Shang, H.; Cao, J.; Yu, H. Effects of ambient ozone concentrations on contents of nonstructural carbohydrates in Phoebe bournei and Pinus massoniana seedlings in subtropical China. Water Air Soil Pollut. 2015, 226, 1–8. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, S. Plant Physiology, 1st ed.; China Agriculture Press: Beijing, China, 2004; pp. 167–187. [Google Scholar]
- Hoch, G. Cell wall hemicelluloses as mobile carbon stores in non-reproductive plant tissues. Funct. Ecol. 2007, 21, 823–834. [Google Scholar] [CrossRef]
- Rook, F.; Hadingham, S.A.; Li, Y.; Bevan, M.W. Sugar and ABA response pathways and the control of gene expression. Plant Cell Environ. 2006, 29, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Körner, C.; Hoch, G. End of season carbon supply status of woody species near the treeline in western china. Basic Appl. Ecol. 2006, 7, 370–377. [Google Scholar] [CrossRef]
- Yu, L.; Wang, C.; Wang, X. Allocation of nonstructural carbohydrates for three temperate tree species in Northeast China. Chin. J. Plant Ecol. 2011, 35, 1245–1255. [Google Scholar] [CrossRef]
- Zhang, W. Effects of Elevated O3 Level on the Native Tree Species in Subtropical China. Ph.D. Thesis, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing, China, 2011. [Google Scholar]
- Niu, J. Effects of Elevated Ozone and Nitrogen Deposition on the Growth and Physiology of Cinnamomum Camphora Seedlings. Ph.D. Thesis, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing, China, 2012. [Google Scholar]
- Cao, J.; Shang, H.; Chen, Z.; Tian, Y.; Yu, H. Effects of elevated ozone on stoichiometry and nutrient pools of Phoebe bournei (Hemsl.) Yang and Phoebe zhennan S. Lee et F. N. Wei seedlings in subtropical China. Forests 2016, 7, 78. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Z.; Shang, H.; Cao, J. Physiological and biochemical responses of Machilus ichangensis Rehd. et Wils and Taxus chinensis (Pilger) Rehd. to elevated O3 in subtropical China. Environ Sci. Pollut. Res. 2017. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Yu, X.; Yang, M.; Ke, L.; Liu, Y.; Lan, X. Effects of recommended fertilization on growth and photosynthetic physiology of Machilus ichangensis Rehd. et Wils seedlings. China For. Sci. Tech. 2009, 23, 46–49. [Google Scholar]
- Yang, B.; Ju, T.; Cao, C.; Li, B.; Zhang, S.; Ma, J. Dynamic analysis of Taxus wallichiana var. chinensis population in Xiaolong Mountain National Nature Reserve, Gansu Province. J. Gansu Agric. Univ. 2015, 50, 88–93. [Google Scholar]
- Zhang, X.; Fu, X.; Wang, G.; Cheng, F.; Chen, J. Anti-inflammatory activity in vitro of n-BuOH extract from Machilus ichangensis. J. China Three Gorges Univ. 2013, 35, 98–102. [Google Scholar]
- Bao, W.; Chen, Q. Present status, problems, and further development strategies on natural Taxus resource and their exploitation in China. J. Nat. Resour. 1998, 13, 375–380. [Google Scholar]
- Dong, W.; Zhang, X.; Wang, H.; Dai, X.; Sun, X.; Qiu, W.; Yang, F. Effect of different fertilizer application on the soil fertility of paddy soils in red soil region of southern China. PLoS ONE 2012, 7, e44504. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Tang, X.; Zhang, Y.; Li, W. City clusters in China: Air and surface water pollution. Front. Ecol. Environ. 2006, 4, 353–361. [Google Scholar] [CrossRef]
- Manual on Methodologies and Criteria for Modelling and Mapping Critical Loads and Levels and Air Pollution Effects, Risks and Trends. Available online: http://www.icpmapping.org./Latest_update_Mapping_ Manual (accessed on 15 July 2017).
- Ma, Z.; Wang, Y.; Zhang, X.; Xu, J. Comparison of ozone between Beijing and downstream Area. Environ. Sci. 2011, 32, 924–929. [Google Scholar]
- Anav, A.; De Marco, A.; Proietti, C.; Alessandri, A.; Dell’Aquila, A.; Cionni, I.; Friedlingstein, P.; Khvorostyanov, D.; Menut, L.; Paoletti, E.; et al. Comparing concentration-based (AOT40) and stomatal uptake (PODY) metrics for ozone risk assessment to European forests. Glob. Chang. Biol. 2015, 22, 1608–1627. [Google Scholar] [CrossRef] [PubMed]
- Eggleston, H.S.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K. 2006 IPCC Guidelines for National Greenhouse Gas Inventories (Volume 4) Agriculture, Forestry and Other Land Use, 1st ed.; Institute for Global Environmental Strategies: Hayama, Japan, 2006; p. 48. [Google Scholar]
- Riikonen, J.; Kontunen-Soppela, S.; Ossipov, V.; Tervahauta, A.; Tuomainen, M.; Oksanen, E.; Vapaavuori, E.; Heinonen, E.; Kivimäenpää, M. Needle metabolome, freezing tolerance and gas exchange in Norway spruce seedlings exposed to elevated temperature and ozone concentration. Tree Physiol. 2012, 32, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Kostiainen, K.; Kaakinen, S.; Warsta, E.; Kubiske, M.E.; Nelson, N.D.; Sober, J.; Karnosky, D.; Saranpaa, P.; Vapaavuori, E. Wood properties of trembling aspen and paper birch after 5 years of exposure to elevated concentrations of CO2 and O3. Tree Physiol. 2008, 28, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Barnes, R.L. Effects of chronic exposure to ozone on soluble sugar and ascorbic acid contents of pine seedlings. Can. J. Bot. 1972, 50, 215–219. [Google Scholar] [CrossRef]
- Landolt, W.; Günthardt-Goerg, M.; Pfenninger, I.; Scheidegger, C. Ozone-induced microscopical changes and quantitative carbohydrate contents of hybrid poplar (Populus × euramericana). Trees 1994, 8, 183–190. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, X.G.; Xue, L.; Chen, H.Y.; Liang, Z.Y. Effects of ozone stress on physiological characteristics of Elaeocarpus sylvestris seedlings. J. Cent. South Univ. Technol. 2015, 35, 97–103. [Google Scholar]
- Meier, S.; Grand, L.F.; Schoeneberger, M.M.; Reinert, R.A.; Bruck, R.I. Growth, ectomycorrhizae and nonstructural carbohydrates of loblolly pine seedlings exposed to ozone and soil water deficit. Environ. Pollut. 1990, 64, 11–17. [Google Scholar] [CrossRef]
- Paynter, V.A.; Reardon, J.C.; Shelburne, V.B. Carbohydrate changes in shortleaf pine (Pinus echinata) needles exposed to acid rain and ozone. Can. J. Res. 1991, 21, 666–671. [Google Scholar] [CrossRef]
- Braun, S.; Zugmaier, U.; Thomas, V.; Flückiger, W. Carbohydrate concentrations in different plant parts of young beech and spruce along a gradient of ozone pollution. Atmos. Environ. 2004, 38, 2399–2407. [Google Scholar] [CrossRef]
- Neufeld, H.S.; Peoples, S.J.; Davison, A.W.; Chappelka, A.H.; Somers, G.L.; Thomley, J.E.; Booker, F.L. Ambient ozone effects on gas exchange and total non-structural carbohydrate levels in cutleaf coneflower (Rudbeckia laciniata L.) growing in Great Smoky Mountains National Park. Environ. Pollut. 2012, 160, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.P.; Scagel, C.F. Nutrient availability alters below-ground respiratory responses of Ponderosa Pine to ozone. Tree Physiol. 1997, 17, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.P. Source-sink balance and carbon allocation below ground in plants exposed to ozone. New Phytol. 2003, 157, 213–218. [Google Scholar] [CrossRef]
- Tingey, D.T.; Laurence, J.A.; Weber, J.A.; Greene, J.; Hogsett, W.E.; Brown, S.; Lee, E.H. Elevated CO2 and temperature alter the response of Pinus ponderosa to ozone: A simulation analysis. Ecol. Appl. 2001, 11, 1412–1424. [Google Scholar]
- Wang, X.; Zheng, Q.; Feng, Z.; Xie, J.; Feng, Z.; Ouyang, Z.; Manning, W.J. Comparison of a diurnal vs steady-state ozone exposure profile on growth and yield of oilseed rape (Brassica napus L.) in open-top chambers in the Yangtze Delta, China. Environ. Pollut. 2008, 156, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Ögren, E.; Nilsson, T.; Sunfblad, L.G. Relationship between respiratory depletion of sugars and loss of cold hardiness in coniferous seedlings over-wintering at raised temperatures: Indications of different sensitivities of spruce and pine. Plant Cell Environ. 1997, 20, 247–253. [Google Scholar] [CrossRef]
- Morin, X.; Améglio, T.; Ahas, R.; Kurz-besson, C.; Lanta, V.; Lebourgeois, F.; Miglietta, F.; Chuine, I. Variation in cold hardiness and carbohydrate concentration from dormancy induction to bud burst among provenances of three European oak species. Tree Physiol. 2007, 27, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.M.; Basahi, J.M.; Hassan, I.A. Gas exchange and chlorophyll fluorescence of pea (Pisum sativum L.) plants in response to ambient ozone at a rural site in Egypt. Sci. Total Environ. 2014, 498, 585–593. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissues | NSC Components | Pn | Total C Stock | |
|---|---|---|---|---|
| M. ichangensis | Foliage | NSC | 0.807 ** | 0.610 |
| Soluble sugars | 0.848 ** | 0.721 * | ||
| Starch | 0.680 * | 0.518 | ||
| Stem | NSC | 0.395 | 0.389 | |
| Soluble sugars | 0.578 | 0.435 | ||
| Starch | −0.072 | 0.077 | ||
| Root | NSC | −0.031 | 0.133 | |
| Soluble sugars | 0.221 | 0.175 | ||
| Starch | −0.225 | 0.030 | ||
| T. wallichiana | Foliage | NSC | 0.858 ** | 0.729 * |
| Soluble sugars | 0.871 ** | 0.745 * | ||
| Starch | −0.121 | −0.099 | ||
| Stem | NSC | −0.449 | −0.079 | |
| Soluble sugars | −0.358 | 0.034 | ||
| Starch | −0.294 | −0.386 | ||
| Root | NSC | 0.720 * | 0.420 | |
| Soluble sugars | 0.580 | 0.365 | ||
| Starch | 0.685 * | 0.379 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Chen, Z.; Yu, H.; Shang, H. Differential Responses in Non-structural Carbohydrates of Machilus ichangensis Rehd. et Wils. and Taxus wallichiana Zucc. Var. chinensis (Pilg.) Florin Seedlings to Elevated Ozone. Forests 2017, 8, 323. https://doi.org/10.3390/f8090323
Cao J, Chen Z, Yu H, Shang H. Differential Responses in Non-structural Carbohydrates of Machilus ichangensis Rehd. et Wils. and Taxus wallichiana Zucc. Var. chinensis (Pilg.) Florin Seedlings to Elevated Ozone. Forests. 2017; 8(9):323. https://doi.org/10.3390/f8090323
Chicago/Turabian StyleCao, Jixin, Zhan Chen, Hao Yu, and He Shang. 2017. "Differential Responses in Non-structural Carbohydrates of Machilus ichangensis Rehd. et Wils. and Taxus wallichiana Zucc. Var. chinensis (Pilg.) Florin Seedlings to Elevated Ozone" Forests 8, no. 9: 323. https://doi.org/10.3390/f8090323