Effects of Warming and Precipitation Manipulation on Fine Root Dynamics of Pinus densiflora Sieb. et Zucc. Seedlings


Department of Environmental Science and Ecological Engineering, Graduate School, Korea University, Seoul 02841, Korea
*
Author to whom correspondence should be addressed.
Forests 2018, 9(1), 14; https://doi.org/10.3390/f9010014
Submission received: 14 November 2017
/
Revised: 19 December 2017
/
Accepted: 22 December 2017
/
Published: 25 December 2017
(This article belongs to the Special Issue Forest Fine Roots in Changing Climate)
Abstract
:Air warming (TC: control; TW: +3 °C) and precipitation manipulation (PC: control; PD: −30%; PI: +30%) were established to examine effects of these treatments on fine root production (FRP), fine root mortality (FRM), and total root (coarse and fine root) biomass in 33- to 59-month-old Pinus densiflora Sieb. et Zucc. seedlings for two years. We hypothesized that warming and altered precipitation would affect the growth, death, and biomass of fine roots by changing soil temperature and soil water availability. Mean annual FRP and total root biomass were significantly altered by only precipitation manipulation: they were 29.3% (during the two-year period) and 69.0% (after the entire two years) higher, respectively, in PD plots than in PC plots, respectively. In contrast, only warming had a significant effect on mean annual FRM, being 13.2% lower in TW plots than TC plots during the two-year period. Meanwhile, fine root biomass was affected negatively and simultaneously by both soil temperature and soil moisture. It seemed that fine root dynamics have changed so that they maintain their systems in response to the altered soil temperature and moisture. The current study adds significant knowledge for understanding the fine root dynamics of P. densiflora seedlings under altered temperature and precipitation regimes.
1. Introduction
It is expected that global mean temperature and precipitation are going to increase by +1.1 to +4.8 °C, and decrease or increase −50 to +50%, respectively, by the turn of this century [1]. Soil temperature may also increase by about 70–120% as a result of the increase in air temperature [2,3]. The interactions between changed air or soil temperatures and precipitation levels are expected to have significant consequences for below-ground dynamics, such as root dynamics, root respiration, and microbial decomposition, as well as for above-ground ecophysiological processes [4].
Plant roots are a crucial structure connecting above- and below-ground carbon (C) systems, and perform a critical role in plant productivity and the control of below-ground C dynamics [5]. Specifically, fine roots are the main structures linking nutrient and water absorption, C process, and microbial communities in below-ground ecosystems [6]. A large amount of net primary production (7–76%) is assigned to fine root dynamics, such as fine root production and fine root mortality, in forests, though fine root biomass comprises a small portion of total biomass [7,8].
Pinus densiflora Sieb. et Zucc. forests dominate more than 23% of the South Korean forest area [9] and are broadly distributed across the East Asia region. Seedlings of P. densiflora are extensively used for reforestation due to its cultural and economic importance in Korea [10]. However, seedlings may have low ability to adapt to environmental stress in plantations and the responses occurring at the seedling stage may further influence growth at subsequent life stages [11]. Because of this, climate change experiments on the seedlings of P. densiflora are necessary and important.
Climatic factors directly or indirectly affect fine root dynamics. For the purpose of predicting the responses in below-ground C cycling, it is necessary to understand fine root responses to climate change [12]. Because of this, several climate change experiments have been conducted in temperate and tropical forest ecosystems, in order to examine the effect on fine root dynamics, such as fine root biomass [13,14], production [15,16], mortality [17], and decomposition [13].
Fine root dynamics under warming or precipitation manipulation may depend on soil water availability and soil nutrient availability [18,19]. Warming indirectly increases soil nitrogen availability [20] and decreases soil water availability, and this can influence fine root biomass, production, and mortality [15]. Precipitation manipulation directly changes soil water availability. Fine root responses to climate change experiments vary with ecosystems and regions [6,12]. In ecosystems where water is not limited, temporarily reduced soil water availability stimulates fine root dynamics to ameliorate water stress in general [13]. In addition to changes in soil environmental conditions, physiological responses of plants under warming conditions can affect fine root dynamics in below-ground processes. Air warming can stimulate or hamper physiological parameters, depending on the temperature stress of leaves [21], and these effects can further alter below-ground carbon allocation [22,23]. A previous study in the same experimental system has confirmed a tendency to decrease in net photosynthetic rates under warming conditions [23]. Therefore, we hypothesized that warming would enhance or reduce fine root production, morality, and root biomass because warming can accelerate evaporation of soil water and hinder carbon uptake by altering plant physiology, such as stomatal conductance and transpiration rate. Given that shifts in precipitation change soil moisture availability, we secondly hypothesized that elevated precipitation would reduce fine root production and mortality, and root biomass, and that reduced precipitation would enhance these parameters.
Climate change experiments, such as warming and precipitation manipulation, have mostly focused on each independent effect, while there were few studies that examined the combination effect of two treatments on below-ground dynamics [3]. This study aimed to examine the fine root dynamics to climate changes by exposing P. densiflora seedlings to altered temperatures and precipitation, and a mixture of the two treatments. We thirdly hypothesized that there would be a combination effect between altered temperature and precipitation on fine root production and mortality, and root biomass. Water stress caused by warming with decreased precipitation would stimulate responses among these parameters more than a single treatment. Because of potential combination effects, changes in soil temperature and moisture under the warming and precipitation manipulations in this study would simultaneously affect fine root parameters.
2. Materials and Methods
2.1. Experimental Site
The open-field experimental system was located in the Environmental Ecology Arboretum of Korea University, Seoul, Korea (37°35′36′′ N, 127°1′31′′ E). Annual precipitation and average annual temperature were 1450.5 mm and 12.5 °C, respectively, during 30 years (1981 to 2010) in Seoul, Korea [24]. During the study period, the average annual temperature at the study site was 12.9 °C in 2014 and 13.4 °C in 2015. The annual precipitation at the study site was 704.0 mm in 2014 and 683.5 mm in 2015.
2.2. Experimental Design
In April 2013, 18 experimental plots (1.5 m × 1.5 m) were established, each of which contained 45 24-month-old P. densiflora seedlings. The density was chosen to be in accordance with the values proposed by the Creation and Management of Forest Resources Act of South Korea [25]. Soil pH was 6.52 and the soil was classified as loamy sand [26]. The experiment had six treatments containing three replicates: two levels of air warming (temperature control (TC), +3 °C (TW)) were combined with three levels of precipitation (precipitation control (PC), 30% decreased (PD), 30% increased (PI)). We designed these treatment levels, expected in Korea over the next 50 years with reference to RCP 8.5 climate change, to simulate effects of climate change [27].
One FTE-1000 infrared heater (Mor Electric Heating Association, Comstock Park, MI, USA) was installed in each warmed (TW) plot at a height of 60 cm above the seedling canopy. Air temperature was set to be 3 °C higher in the TW plot, compared to TC plots. If the difference in air temperature between the TC and TW was less than 3.0 °C, the data logger and relay system operated the infrared heater, and if the difference was greater than 3.0 °C, the heater shut off automatically. Dummy heaters, i.e., without infrared heaters, were used in control plots to guarantee the other environmental conditions were the same as in TW plots. PD manipulation was achieved by covering 30% of the area of the plot using transparent panels. PI manipulation accumulated the intercepted precipitation from the PD plots in a water tank, and it then automatically operated a drip-irrigation system using an automatic pump. For the PC and PI, transparent panel covers which did not obstruct precipitation were set to guarantee that the other environmental factors remained the same as for the PD manipulation. Detailed information of the experimental design is provided in [3].
Air temperature was measured by SI-111 infrared temperature sensors (Campbell Scientific, Logan, UT, USA) in each plot (n = 18). Soil temperature and soil moisture were measured in each plot (n = 18) by temperature sensors (107-L34, Campbell Scientific, Logan, UT, USA) and water content sensors (CS616, Campbell Scientific, Logan, UT, USA), respectively. The environmental data were measured with a CR3000 data logger (Campbell Scientific, Logan, UT, USA).
2.3. Fine Root Production and Mortality
On 29 July 2013, a 50 cm transparent acrylic tube was set in the soil of each plot, at a 45° angle to the ground. We covered 20 cm tube remaining above-ground using black tape and aluminum foil, to block out light. Observations began on 27 January 2014, about six months after the installation of the acrylic tubes. Minirhizotron images were taken on 27 January, 29 April, 31 July, 9 November 2014, and 26 January, 26 April, 17 July, 12 October, and 29 December 2015 (33- to 56-month-old seedlings), using a minirhizotron system (CI-600, CID, Camas, WA, USA). The size of each image was 19.56 × 14.14 cm, which covered a 15 cm soil depth. Fine root data (length) on the collected images were measured with WinRHIZO Tron MF software (Regent Instruments, Quebec City, QC, Canada). Fine root length production (FRP) for each collection period was calculated as the sum of emerging root lengths and the length growth of preexisting roots. Fine root length mortality (FRM) for each collection period was calculated as sum of lengths of disappeared roots from minirhizotron images, including dead roots and existing roots. Annual FRP and FRM (μm mm−2 year−1) in 2014 were the sum of FRP and FRM between January 2014 and January 2015. Similarly, annual FRP and FRM in 2015 were the sum of FRP and FRM between January 2015 and December 2015.
2.4. Root Biomass
To estimate root biomass, two or three seedlings, located near the minirhizotron tubes, were sampled in each plot in March 2016 after finalizing the minirhizotron observation (59-month-old seedlings). The root samples were washed, and then sorted into coarse roots (higher than 2 mm) and fine roots (less than 2 mm). After sorting, coarse roots and fine roots were weighed after drying at 65 °C for 48 h. Total root biomass (TRB) was calculated by summing coarse and fine root biomass (CRB and FRB).
2.5. Statistical Analysis
The treatment effects on air temperature, soil temperature, soil moisture, annual FRP, annual FRM, TRB, CRB, and FRB were examined by two-way ANOVA. The differences in air temperature, soil temperature, soil moisture, annual FRP, annual FRM, TRB, CRB, and FRB between the treatments were performed using an LSD test. Linear regression was performed for determining the correlations between warming-effect on annual FRM and warming-induced differences in soil temperature, and between precipitation-effect on annual FRP and precipitation-induced differences in soil moisture content. Warming-effect on annual FRP was calculated as the percentage difference in values from TW × PC, TW × PD, and TW × PI plots relative to TC × PC, TC × PD, and TC × PI plots, respectively. Precipitation-effect on annual FRP was calculated as the percentage difference in values from TC × PD or TC×PI plots relative to TC × PC plots, and from TW × PD or TW × PI plots relative to TW × PC plots, respectively. Similarly, warming or precipitation-effect on the annual FRM was calculated. Warming or precipitation-induced differences in soil temperature and moisture were based on the difference in measurements between the treatments. Multiple regression analysis was performed to determine the relationships between soil temperature and soil moisture with annual FRP, annual FRM, TRB, CRB, and FRB. Path analysis also was used to test the causal model for the effects of soil temperature and soil moisture on annual FRP, annual FRM, FRB, and CRB. SAS 9.4 (SAS Institute Inc., Cary, NC, USA) was used for statistical analyses.
3. Results
3.1. Air Temperature, Soil Temperature, and Soil Moisture
Warming significantly affected air temperature and soil temperature in 2014 and 2015 (Table 1). Specifically, warming increased average air temperature by 2.73 °C, 2.83 °C, and 2.97 °C, and by 2.81 °C, 2.69 °C, and 2.76 °C, in the PC, PD, and PI plots in 2014 and 2015, respectively (Table 2). Besides, warming increased average soil temperature by 2.66 °C, 2.09 °C, and 2.19 °C, and by 1.36 °C, 0.54 °C, and 0.79 °C, in the PC, PD, and PI plots in 2014 and 2015, respectively (Table 2). Warming and precipitation manipulation affected soil moisture in 2014 and 2015 (Table 1). Warming decreased average soil moisture by 0.25 to 1.9 vol %. Compared to the PC plots, average soil moisture content decreased/increased in the PD plots (0.12 to 0.54 vol %) and PI plots (0.54 to 2.10 vol %) respectively, during the study period (Table 2).
3.2. Annual FRP and FRM
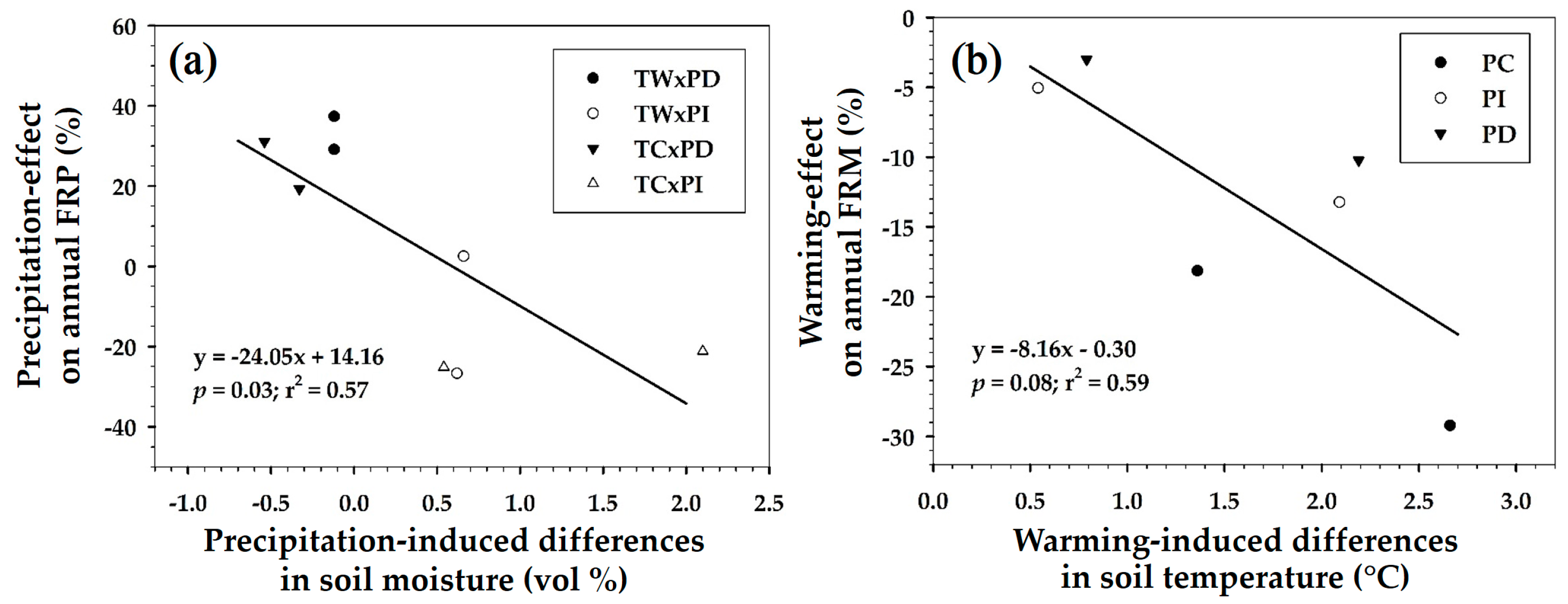
The annual FRPs of 2014 and 2015 were significantly altered by precipitation manipulation; however, they were not influenced by the warming treatment (Table 1). Compared to the PC plots, the annual FRP was 28.4 to 30.1% higher in PD plots and 9.3 to 25.9% lower in PI plots across the two years (Table 2). The annual FRM in the TW plots was 17.6 to 8.7% lower than that in the TC plots across the two years (Table 2). PD and PI manipulation decreased annual FRM (8.6 to 9.5% in PD plots and 8.0–25.4% in PI plots) (Table 2); however, annual FRM did not show significant differences due to the precipitation manipulations (Table 1). Across the two years, there were negative linear relationships between precipitation-effects on annual FRP and precipitation-induced differences in soil moisture (p = 0.03 and r2 = 0.57; Figure 1a), and between warming-effect on annual FRM and warming-induced differences in soil temperature (p = 0.08 and r2 = 0.59; Figure 1b). Meanwhile, multiple regression analyses for annual FRP and FRM and soil temperature and soil moisture were not significant (p > 0.05).
3.3. Root Biomass
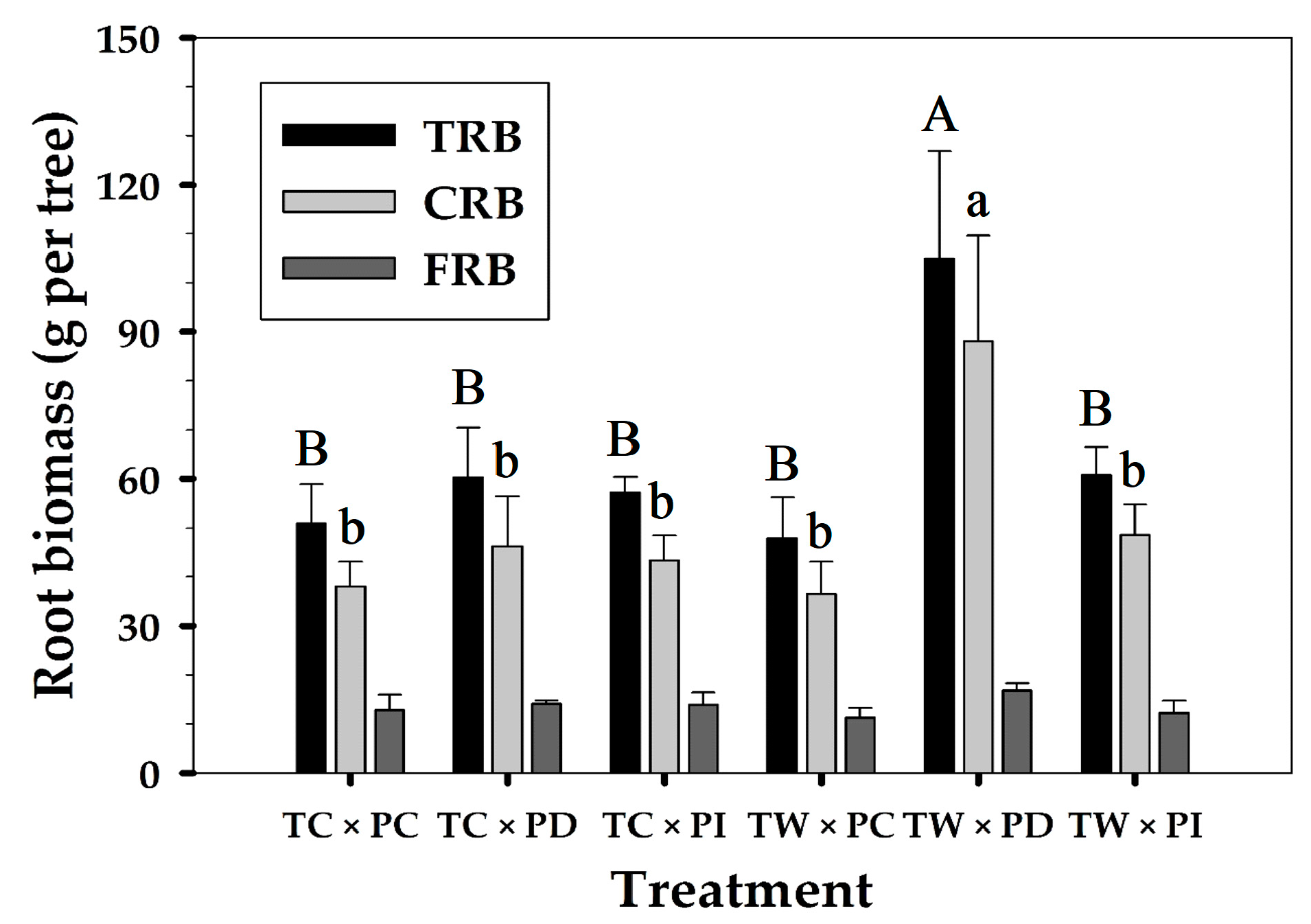
Only precipitation manipulation had a significant effect on TRB and CRB (Table 3). PD was 69.0 to 81.3% higher compared to the PC. CRB (g per tree) in TW × PD (88.1 ± 21.5) was significantly higher than those from other treatments (TC × PC: 38.1, TC × PD: 46.2, TC × PI: 43.3, TW × PC: 36.5, and TW × PI: 48.6) (Figure 2). FRB was higher in PD than PC and PI, regardless of the warming treatment; however, there was no significant difference in FRB among the precipitation manipulations (TC × PC: 12.8, TC × PD: 14.1, TC × PI: 13.8, TW × PC: 11.3, TW × PD: 16.9, and TW × PI: 12.2). A multiple regression model of FRB with soil temperature and soil moisture was significant (p = 0.05) (Table 4), but the regression models of TRB and CRB with soil temperature and soil moisture were not significant (p > 0.05). There was a linear but negative relationship between FRB and soil temperature and soil moisture (r2 = 0.31).
3.4. Path Analysis
Path analysis confirmed that annual FRP and annual FRM were strongly related to changes in soil temperature and soil moisture (Figure 3). Soil temperature had a negative effect on the annual FRM, but little effect on the annual FRP (Figure 3). Soil moisture had a negative effect on annual FRP, but little effect on the annual FRM (Figure 3). Annual FRP had positive effects on CRB and FRB, while annual FRM had negative effects on CRB and FRB (Figure 3). Meanwhile, FRB was directly affected by soil temperature and soil moisture simultaneously. CRB was less affected by soil temperature and soil moisture than FRB.
4. Discussion
During the study period, warming had no effect on annual FRP, TRB, CRB, and FRB in 2014 and 2015. Only annual FRM in 2014 was affected by warming. Annual FRM was decreased by increasing the warming effect. Reduced precipitation showed a strong positive effect on annual FRP, TRB, and CRB. FRB was affected by soil temperature and soil moisture simultaneously, although the effect of warming and precipitation was not significant.
4.1. Effect of Warming
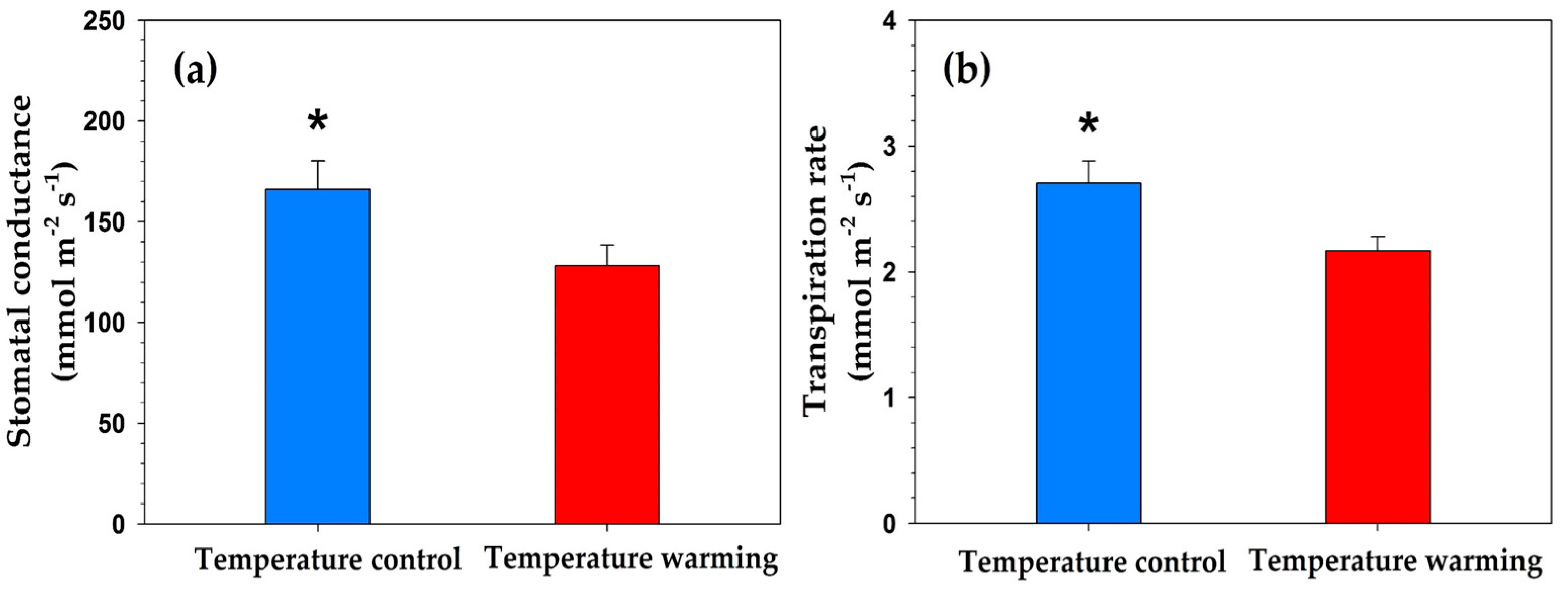
Contrary to our hypothesis 1, warming-induced reductions in soil water content did not affect FRP (including annual FRP) or root biomass (TRB, CRB, and FRB). These results were analogous to previous findings at the same study site, which found that warming had no effect on TRB and FRP of P. densiflora seedlings [23,28]. The absence of the warming effects on FRP, FRM, and root biomass might result from physiological adaptations of leaves to heat and drought stresses. Warming has effects on stomatal conductance, water use efficiency (WUE), and transpiration rate, as well as soil moisture [29,30]. When heat above an appropriate temperature is applied to leaves, the stomatal conductance is lowered, and therefore the transpiration rate is reduced; these changes increase WUE [31,32]. The increase of WUE in the condition of lack of water supply inhibits plant growth, such as root production [33]. Actually, in this study, stomatal conductance and transpiration rates in 2014 (Table A1; Figure A1) and 2015 [34] were significantly lower in the TW plots. Therefore, seedlings might offset the effects of a decrease in soil moisture availability by reducing stomatal conductance and transpiration rate, instead of promoting root growth.
Only the annual FRM of 2014 was affected by warming (Table 3); warming-effect on annual FRM decreased with an increase in the warming effect (Figure 1b). In general, warmer soil temperatures stimulate the rapid death of roots in warming treatment of 1.3 to 1.7 °C [35] and 5.0 °C [36]. However, in some cases, no change in FRM with 2.8 °C warmer treatment [37] or decreases in FRM with relatively weak warmer (0.3 to 1.4 °C) system [12,38] was observed. The contradictory results of previous studies might be explained by the water availability in each ecosystem [39]. The inhibitory effect resulted in root mortality to maintain the root system, because soil water content is a key factor in dry conditions as found in our site [12], and seedlings might be different from mature trees in this matter.
4.2. Effect of Precipitation Manipulation
Our findings support our second hypothesis that altered precipitation negatively affects fine root dynamics. Responses of FRP and root biomass to precipitation changes have been reported in forest ecosystems, however, there are no apparent trends. For example, in previous studies, a drought effect (reduced precipitation) on root growth was positive [13,18,40], or negative [16,19], or insignificant [15]. Specifically, the response of FRP and root biomass to drought appear to vary depending on the soil water condition [18,19]. FRP in forest ecosystems with limited precipitation is more sensitive to soil water content [41]. In our study site, annual precipitation during the study period was half the 30-year average amount of precipitation, due to severe drought. Decreased soil water content increases C allocation to the root system to some extent, because a larger root system is required to satisfy tree water demand in low water conditions [13,40]. In other words, fine roots tend to extend vertically or horizontally to soil in dry conditions [42]. The results in Figure 1a and Table 4, which show negative correlations between precipitation-effect on annual FRP and soil moisture, and between FRB and soil moisture, support the above discussion.
4.3. Interaction between Warming and Precipitation Manipulation
The results of this study speculated that the interaction of warming with reduced precipitation leads to increased TRB and CRB (Figure 2). It might be that warming treatment suppresses fine root death, and reduced precipitation stimulates fine root growth, in the TW × PD. FRB increased with decreasing soil temperature and soil moisture (Figure 3; Table 4), but there were no significant differences across treatments. It seems that FRB differed depending on effects of warming and precipitation manipulations on soil nitrogen availability [43] and soil microbial biomass [13], as well as soil temperature and soil moisture. In previous studies, conducted in various ecosystems, interactions between warming and reduced precipitation significantly decreased soil water availability, resulting in the stimulation of a water stress response [3,44]. These changes in soil water availability might cause an increased growth of the root collar diameter [3], and a decreased FRB [13]. However, there were few studies that examined the interaction effect between warming and reduced precipitation [4].
4.4. Implications for Changes in Fine Root Dynamics
Contrary to a previous study [45], which reported that P. densiflora forests are vulnerable to climate change, P. densiflora seedlings might adapt to increased temperature and reduced precipitation by stimulating rootage and initial growth. The observed changes in fine root dynamics and root biomass could lead to further shifts in P. densiflora forests because the responses occurring at the seedling stage connect to the subsequent growth [3,11]. Based on the relationship between soil water content and fine root dynamics, drought and soil drying by warming can stimulate below-ground net primary production and biomass of P. densiflora seedlings, with an increase in autotrophic respiration originating from plant roots. This autotrophic respiration is directly influenced by changes in below-ground productivity [12,46]. Warming-induced changes in soil temperature can suppress fine root mortality, and subsequently can reduce the return of C and nutrients to soil [35]. These results represent only the initial changes to equilibrate between altered conditions and uptake organs at the beginning of the experiment. The treatment effects (warming and precipitation manipulation) can fade in the long term because the fine roots may gradually adapt to the altered conditions [47]. Meanwhile, natural drought stress might be unexpectedly stronger than precipitation manipulation effects because the study occurred during a period of abnormal drought compared to the average precipitation during the last 30 years (1980–2010). In order to investigate the intact treatment effect (warming or precipitation manipulation) on fine root dynamics or physiology responses, further studies should be conducted under normal precipitation conditions.
5. Conclusions
This study examined initial changes in the fine root dynamics of P. densiflora seedlings under warming and precipitation manipulation over a two-year period. We found significant shifts in fine root productivity and total root biomass following precipitation manipulation; conversely, there was a significant effect on fine root death following the warming treatment. There were negative relationships between FRP and soil moisture, and FRM and soil temperature, respectively. However, TRB and FRB remained unchanged by warming. Seedlings might offset the decrease in soil moisture availability by reducing physiological factors such as stomatal conductance and transpiration rate. These results will be of great value for predicting the important aspect of the below-ground dynamics of P. densiflora seedlings in response to altered temperature and precipitation regimes in Korea.
Acknowledgments
This study was supported by the National Research Foundation of Korea (NRF-2013R1A1A2012242) and Korea Forest Service (2017058A00-1719-AB01).
Author Contributions
Seung Hyun Han, Soon Jin Yun, and Yowhan Son designed the study; Guanlin Li, Hanna Chang, and Jiae An collected field data; Seung Hyun Han and Seongjun Kim analyzed the data; Seung Hyun Han led writing of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
F-ratios of two-way ANOVA on the effects of warming (W) and precipitation manipulation (P) on stomatal conductance and transpiration rates of Pinus densiflora seedlings.
Table A1.
F-ratios of two-way ANOVA on the effects of warming (W) and precipitation manipulation (P) on stomatal conductance and transpiration rates of Pinus densiflora seedlings.
| Stomatal Conductance 1 | Transpiration Rates 1 | ||
|---|---|---|---|
| 2014 | W | 3.90 * | 5.43 ** |
| P | 0.21 | 0.17 | |
| W × P | 0.37 | 0.30 |
1 Stomatal conductance and transpiration rates were measured in May, July, September, and November in 2014, using CIRAS-2 (PP-Systems, Hitchin, UK). * = significant (p < 0.1); ** = significant (p < 0.05).
Figure A1.
Effect of warming on (a) stomatal conductance and (b) transpiration rate of Pinus densiflora seedlings in 2014.
Figure A1.
Effect of warming on (a) stomatal conductance and (b) transpiration rate of Pinus densiflora seedlings in 2014.

References
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; 1535p. [Google Scholar]
- An, J.; Han, S.; Chang, H.; Park, M.J.; Kim, S.; Hwang, J.; Cho, M.S.; Chung, H.; Son, Y. Physiological and growth responses to experimental warming in first-year seedlings of deciduous tree species. Turk. J. Agric. For. 2017, 41, 175–182. [Google Scholar] [CrossRef]
- Yun, S.J.; Han, S.; Han, S.H.; Kim, S.; Li, G.; Park, M.; Son, Y. Short-term effects of warming treatment and precipitation manipulation on the ecophysiological responses of Pinus densiflora seedlings. Turk. J. Agric. For. 2016, 40, 621–630. [Google Scholar] [CrossRef]
- Wu, Z.; Dijkstra, P.; Koch, G.W.; Penuelas, J.; Hungate, B.A. Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation. Glob. Chang. Biol. 2010, 17, 927–942. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, X.; Zi, H.; Hu, L.; Ade, L.; Wang, G.; Lerdau, M. The effect of simulated warming on root dynamics and soil microbial community in an alpine meadow of the Qinghai-Tibet Plateau. Appl. Soil Ecol. 2017, 116, 30–41. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Chen, H.Y.H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Gower, S.T.; Isebrands, J.G.; Sheriff, D.W. Carbon allocation and accumulation in conifers. In Resource Physiology of Conifers; Smith, W.K., Hinckley, T.M., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 217–254. [Google Scholar]
- Vogt, K.A. Carbon budgets of temperate forest ecosystems. Tree Physiol. 1991, 9, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korea, 2011. [Google Scholar]
- Li, X.; Son, Y.M.; Lee, K.H.; Kim, R.H.; Jin, G.; Son, Y.; Park, P.S.; Yi, M.J. Biomass and carbon storage in an age-sequence of Japanese red pine (Pinus densiflora) forests in central Korea. For. Sci. Technol. 2013, 9, 39–44. [Google Scholar] [CrossRef]
- Mardani, Z.; Rabiei, B.; Sabouri, H.; Sabouri, A. Identification of molecular markers linked to salt-tolerant genes at germination stage of rice. Plant Breed. 2014, 133, 196–202. [Google Scholar] [CrossRef]
- Bai, W.; Wan, S.; Niu, S.; Liu, W.; Chen, Q.; Wang, Q.; Zhang, W.; Han, X.; Li, L. Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: Implications for ecosystem C cycling. Glob. Chang. Biol. 2010, 16, 1306–1316. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, S.; Wan, S.; Wang, J.; Wang, H.; Liu, K. Effects of experimental throughfall reduction and soil warming on fine root biomass and its decomposition in a warm temperate oak forest. Sci. Total Environ. 2017, 574, 1448–1455. [Google Scholar] [CrossRef] [PubMed]
- Sotta, E.D.; Veldkamp, E.; Schwendenmann, L.; Guimaraes, B.R.; Paixao, R.K.; Ruivo, M.L.P.; Lola Da Costa, A.C.; Meir, P. Effects of an induced drought on soil carbon dioxide (CO2) efflux and soil CO2 production in an eastern Amazonian rainforest, Brazil. Glob. Chang. Biol. 2007, 13, 2218–2229. [Google Scholar] [CrossRef]
- Joslin, J.D.; Wolfe, M.H.; Hanson, P.J. Effects of altered water regimes on forest root systems. New Phytol. 2000, 147, 117–129. [Google Scholar] [CrossRef]
- Sayer, M.A.S.; Haywood, J.D. Fine root production and carbohydrate concentrations of mature longleaf pine (Pinus palustris P. Mill.) as affected by season of prescribed fire and drought. Trees 2006, 20, 165–175. [Google Scholar] [CrossRef]
- Wan, S.; Norby, R.J.; Pregitzer, K.S.; Ledford, J.; O’Neill, E.G. CO2 enrichment and warming of the atmosphere enhance both productivity and mortality of maple tree fine roots. New Phytol. 2004, 162, 437–446. [Google Scholar] [CrossRef]
- Leuschner, C.; Backes, K.; Hertel, D.; Schipka, F.; Schmitt, U.; Terborg, O.; Runge, M. Drought responses at leaf, stem and fine root levels of competitive Fagus sylvatica L. and Quercus petraea (Matt.) Liebl. trees in dry and wet years. For. Ecol. Manag. 2001, 149, 33–46. [Google Scholar] [CrossRef]
- Jones, R.H.; Mitchell, R.J.; Stevens, G.N.; Pecot, S.D. Controls of fine root dynamics across a gradient of gap sizes in a pine woodland. Oecologia 2003, 134, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J.; GCTE-NEWS. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [PubMed]
- Saxe, H.; Cannell, M.G.R.; Johnsen, Ø.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef]
- Lewis, J.D.; Lucash, M.; Olszyk, D.; Tingey, D.T. Seasonal patterns of photosynthesis in Douglas fir seedlings during the third and fourth year of exposure to elevated CO2 and temperature. Plant Cell Environ. 2001, 24, 539–548. [Google Scholar] [CrossRef]
- Park, M.J.; Yun, S.J.; Yun, H.M.; Chang, H.; Han, S.H.; An, J.; Son, Y. Effects of open-field artificial warming and precipitation manipulation on physiological characteristics and growth of Pinus densiflora seedlings. J. Clim. Chang. Res. 2016, 7, 9–17, (In Korean with English abstract). [Google Scholar] [CrossRef]
- Korea Meteorological Administration. Climatological Normals of Korea; Korea Meteorological Administration: Seoul, Korea, 2011. [Google Scholar]
- Korea Forest Service. Creation and Management of Forest Resources Act; Korea Forest Service: Daejeon, Korea, 2012. (In Korean) [Google Scholar]
- Yun, S.J.; Han, S.; Han, S.H.; Lee, S.J.; Jung, Y.; Kim, S.; Son, Y. Open-field experimental warming and precipitation manipulation system design to simulate climate change impact. J. Korean For. Soc. 2014, 103, 159–164, (In Korean with English abstract). [Google Scholar] [CrossRef]
- Climate Information Portal. Available online: http://www.climate.go.kr/ (accessed on 4 December 2017).
- Lee, S.J.; Han, S.; Yoon, T.K.; Han, S.H.; Jung, Y.; Yun, S.J.; Son, Y. Growth and physiological characteristics of Pinus densiflora seedlings in response to open-field experimental warming using the infrared lamp. J. Korean For. Soc. 2013, 102, 522–529, (In Korean with English abstract). [Google Scholar]
- Loik, M.E.; Harte, J. Changes in water relations for leaves exposed to a climate-warming manipulation in the Rocky Mountains of Colorado. Environ. Exp. Bot. 1997, 37, 115–123. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, M.; Hou, R.; Shen, R.; Qiu, S.; Ouyang, Z. Effects of experimental warming on stomatal traits in leaves of maize (Zea may L.). Ecol. Evol. 2013, 3, 3095–3111. [Google Scholar] [CrossRef] [PubMed]
- Hamerlynck, E.P.; Knapp, A.K. Early season cuticular conductance and gas exchange in two oaks near the western edge of their range. Trees 1996, 10, 403–409. [Google Scholar] [CrossRef]
- Penuelas, J.; Filella, I.; Llusia, J.; Siscart, D.; Pinol, J. Comparative field study of spring and summer leaf gas exchange and photobiology of the Mediterranean trees Quercus ilex and Philyrea latifolia. Exp. Bot. 1998, 319, 229–238. [Google Scholar]
- Lim, J.H.; Woo, S.Y.; Kwon, M.J.; Chun, J.H.; Shin, J.H. Photosynthetic capacity and water use efficiency under different temperature regimes on healthy and declining Korean fir in Mt. Halla. J. Korean For. Soc. 2006, 95, 705–710, (In Korean with English abstract). [Google Scholar]
- Han, S.H.; Son, Y.; Kim, S.; Li, G.; Chang, H. Effect of soil moisture on fine root production under experimental warming and precipitation manipulation. In Proceedings of the 7th International Symposium on Physiological Processes in Roots of Woody Plants, University of Tartu, Tartu, Estonia, 26–29 June 2017; Kriiska, K., Rosenvald, K., Meitern, A., Ostonen, I., Eds.; University of Tartu: Tartu, Estonia, 2017. [Google Scholar]
- Hendrick, R.L.; Pregitzer, K.S. Patterns of fine root mortality in two sugar maple forests. Nature 1993, 361, 59–61. [Google Scholar] [CrossRef]
- Majdi, H.; Öhrvik, J. Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden. Glob. Chang. Biol. 2004, 10, 182–188. [Google Scholar] [CrossRef]
- Fitter, A.H.; Self, G.K.; Brown, T.K.; Bogie, D.S.; Graves, J.D.; Benham, D.; Ineson, P. Root production and turnover in an upland grassland subjected to artificial soil warming respond to radiation flux and nutrients, not temperature. Oecologia 1999, 120, 575–581. [Google Scholar] [PubMed]
- Bai, W.M.; Xia, J.Y.; Wan, S.Q.; Zhang, W.H.; Li, L.H. Day and night warming have different effect on root lifespan. Biogeosciences 2012, 9, 375–384. [Google Scholar] [CrossRef]
- Wan, S.; Hui, D.; Wallace, L.L.; Luo, Y. Direct and indirect warming effects on ecosystem carbon processes in a tallgrass prairie. Glob. Biogeochem. Cycles 2005, 19, 159–160. [Google Scholar] [CrossRef]
- Hertel, D.; Strecker, T.; Muller-Haubold, H.; Leuschner, C. Fine root biomass and dynamics in beech forests across a precipitation gradient—Is optimal resource partitioning theory applicable to water-limited mature trees? J. Ecol. 2013, 101, 1183–1200. [Google Scholar] [CrossRef]
- Johnson, M.G.; Rygiewicz, P.T.; Tingey, D.T.; Phillips, D.L. Elevated CO2 and elevated temperature have no effect on Douglas-fir fine root dynamics in nitrogen-poor soil. New Phytol. 2006, 170, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Gieger, T.; Thomas, F.M. Effects of defoliation and drought stress on biomass partitioning and water relations of Quercus robur and Quercus petraea. Basic Appl. Ecol. 2002, 3, 171–181. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J. The potential effects of nitrogen deposition on fine-root production in forest ecosystems. New Phytol. 2000, 147, 131–139. [Google Scholar] [CrossRef]
- Brando, P.M.; Nepstad, D.C.; Davidson, E.A.; Trumbore, S.E.; Ray, D.; Camargo, P. Drought effect on litterfall, wood production and belowground carbon cycling in an Amazon forest: Results of a throughfall reduction experiment. Philos. Trans. R. Soc. B 2008, 363, 1839–1848. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.; Lee, W.; Nor, D.; Kim, S.; Choi, J.; Lee, Y. The relationship between tree radial growth and topographic and climate factors in red pine and oak in central regions of Korea. J. Korean For. Soc. 2010, 99, 908–913, (In Korean with English abstract). [Google Scholar]
- Noh, N.J.; Lee, S.J.; Jo, W.; Han, S.; Yoon, T.K.; Chung, H.; Son, Y. Effects of experimental warming on soil respiration and biomass in Quercus variabilis Blume and Pinu sdensiflora Sieb. et Zucc. seedlings. Ann. For. Sci. 2016, 73, 533–545. [Google Scholar] [CrossRef]
- Atkin, O.K.; Edwards, E.J.; Loveys, B.R. Response of root respiration to changes in temperature and its relevance to global warming. New Phytol. 2000, 147, 141–154. [Google Scholar] [CrossRef]
Figure 1.
(a) Linear regression between precipitation-effect on annual fine root length production (FRP) of Pinus densiflora seedlings and precipitation-induced differences in soil moisture; (b) between warming-effect on annual fine root length mortality (FRM) and warming-induced differences in soil temperature. The treatment abbreviations are the same as in Table 2.
Figure 1.
(a) Linear regression between precipitation-effect on annual fine root length production (FRP) of Pinus densiflora seedlings and precipitation-induced differences in soil moisture; (b) between warming-effect on annual fine root length mortality (FRM) and warming-induced differences in soil temperature. The treatment abbreviations are the same as in Table 2.

Figure 2.
Total root biomass (TRB), coarse root biomass (CRB), and fine root biomass (FRB) of Pinus densiflora seedlings for each treatment in March 2016. Vertical bar indicates one standard error (n = 3). Values followed by a different letter are significantly different (p < 0.05). The treatment abbreviations are the same as in Table 2.
Figure 2.
Total root biomass (TRB), coarse root biomass (CRB), and fine root biomass (FRB) of Pinus densiflora seedlings for each treatment in March 2016. Vertical bar indicates one standard error (n = 3). Values followed by a different letter are significantly different (p < 0.05). The treatment abbreviations are the same as in Table 2.

Figure 3.
Causal models developed using path analysis for soil temperature (ST) and soil moisture (SM) effects on annual fine root production (FRP), annual fine root mortality (FRM), fine root biomass (FRB), and coarse root biomass (CRB) of Pinus densiflora seedlings. Numbers on arrows represent standardized path coefficients (the width of arrows is proportional to the absolute value of each coefficient).
Figure 3.
Causal models developed using path analysis for soil temperature (ST) and soil moisture (SM) effects on annual fine root production (FRP), annual fine root mortality (FRM), fine root biomass (FRB), and coarse root biomass (CRB) of Pinus densiflora seedlings. Numbers on arrows represent standardized path coefficients (the width of arrows is proportional to the absolute value of each coefficient).

Table 1.
F-ratios of two-way ANOVA on the effects of warming (W) and precipitation manipulation (P) on air temperature (AT), soil temperature (ST), soil moisture (SM), annual fine root length production (FRP), and annual fine root length mortality (FRM) of Pinus densiflora Sieb. et Zucc. seedlings.
Table 1.
F-ratios of two-way ANOVA on the effects of warming (W) and precipitation manipulation (P) on air temperature (AT), soil temperature (ST), soil moisture (SM), annual fine root length production (FRP), and annual fine root length mortality (FRM) of Pinus densiflora Sieb. et Zucc. seedlings.
| AT | ST | SM | Annual FRP | Annual FRM | ||
|---|---|---|---|---|---|---|
| 2014 | W | 1071.11 *** | 200.21 *** | 7.76 ** | 0.02 | 5.00 * |
| P | 2.39 | 1.37 | 10.71 *** | 6.80 ** | 0.69 | |
| W × P | 0.64 | 1.16 | 2.85 | 0.70 | 0.84 | |
| 2015 | W | 91.29 *** | 13.77 ** | 17.77 ** | 0.46 | 0.34 |
| P | 0.07 | 0.50 | 5.81 ** | 9.93 ** | 0.99 | |
| W × P | 0.05 | 1.01 | 0.34 | 0.02 | 0.13 |
* = significant (p < 0.05); ** = significant (p < 0.01); *** = significant (p < 0.001).
Table 2.
Air temperature (AT, °C), soil temperature (ST, °C), soil moisture (SM, vol %), annual fine root length production (FRP, mm cm−2 year−1), and annual fine root length mortality (FRM, mm cm−2 year−1) of Pinus densiflora seedlings under six treatments, expressed as mean value ± standard error.
Table 2.
Air temperature (AT, °C), soil temperature (ST, °C), soil moisture (SM, vol %), annual fine root length production (FRP, mm cm−2 year−1), and annual fine root length mortality (FRM, mm cm−2 year−1) of Pinus densiflora seedlings under six treatments, expressed as mean value ± standard error.
| TC × PC | TC × PD | TC × PI | TW × PC | TW × PD | TW × PI | ||
|---|---|---|---|---|---|---|---|
| 2014 | AT | 13.11 ± 0.06 b | 13.26 ± 0.12 b | 12.98 ± 0.19 b | 15.84 ± 0.12 a | 16.09 ± 0.03 a | 15.95 ± 0.01 a |
| ST | 14.43 ± 0.26 b | 14.61 ± 0.06 b | 14.34 ± 0.22 b | 17.09 ± 0.31 a | 16.70 ± 0.11 a | 16.53 ± 0.12 a | |
| SM | 6.41 ± 0.33 b | 6.08 ± 0.02 b | 8.51 ± 0.79 a | 5.95 ± 0.31 b | 5.83 ± 0.08 b | 6.61 ± 0.12 b | |
| Annual FRP | 1.56 ± 0.16 ab | 1.87 ± 0.17 a | 1.23 ± 0.16 b | 1.36 ± 0.21 b | 1.86 ± 0.04 a | 1.39 ± 0.14 ab | |
| Annual FRM | 1.48 ± 0.09 a | 1.21 ± 0.01 ab | 1.20 ± 0.16 ab | 1.05 ± 0.15 b | 1.05 ± 0.16 b | 1.08 ± 0.14 ab | |
| 2015 | AT | 13.19 ± 0.17 b | 13.29 ± 0.26 b | 13.10 ± 0.10 b | 16.00 ± 0.73 a | 15.88 ± 0.23 a | 15.86 ± 0.20 a |
| ST | 14.22 ± 0.39 b | 14.74 ± 0.20 ab | 14.32 ± 0.21 b | 15.58 ± 0.36 a | 15.28 ± 0.34 a | 15.11 ± 0.20 ab | |
| SM | 6.65 ± 0.44 ab | 6.11 ± 0.03 bc | 7.19 ± 0.27 a | 5.56 ± 0.39 c | 5.44 ± 0.10 c | 6.18 ± 0.12 bc | |
| Annual FRP | 0.99 ± 0.03 ab | 1.30 ± 0.24 a | 0.74 ± 0.07 b | 1.07 ± 0.15 ab | 1.39 ± 0.10 a | 0.79 ± 0.07 b | |
| Annual FRM | 0.91 ± 0.04 a | 0.77 ± 0.12 a | 0.62 ± 0.30 a | 0.74 ± 0.16 a | 0.73 ± 0.12 a | 0.60 ± 0.02 a | |
TC = temperature control; TW = temperature warming; PC = precipitation control; PD = precipitation decreased; PI = precipitation increased; Values followed by a different letter are significantly different (p < 0.05).
Table 3.
F-ratios of two-way ANOVA on the effects of warming (W) and precipitation manipulation (P) on total root biomass (TRB), coarse root biomass (CRB), and fine root biomass (FRB) of Pinus densiflora seedlings.
Table 3.
F-ratios of two-way ANOVA on the effects of warming (W) and precipitation manipulation (P) on total root biomass (TRB), coarse root biomass (CRB), and fine root biomass (FRB) of Pinus densiflora seedlings.
| TRB | CRB | FRB | |
|---|---|---|---|
| W | 2.70 | 2.98 | 0.00 |
| P | 4.65 * | 4.06 * | 1.25 |
| W × P | 2.65 | 2.35 | 0.65 |
* = significant (p < 0.05).
Table 4.
Results of multiple regression analysis of fine root biomass of Pinus densiflora seedlings as a function of soil temperature (ST) and soil moisture (SM).
Table 4.
Results of multiple regression analysis of fine root biomass of Pinus densiflora seedlings as a function of soil temperature (ST) and soil moisture (SM).
| Model | Partial r2 | Model r2 | p Value |
|---|---|---|---|
| Fine root biomass −2.18 ST − 2.79 SM + 64.53 | 0.19 (ST), 0.12 (SM) | 0.31 | 0.05 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, S.H.; Kim, S.; Li, G.; Chang, H.; Yun, S.J.; An, J.; Son, Y. Effects of Warming and Precipitation Manipulation on Fine Root Dynamics of Pinus densiflora Sieb. et Zucc. Seedlings. Forests 2018, 9, 14. https://doi.org/10.3390/f9010014
AMA Style
Han SH, Kim S, Li G, Chang H, Yun SJ, An J, Son Y. Effects of Warming and Precipitation Manipulation on Fine Root Dynamics of Pinus densiflora Sieb. et Zucc. Seedlings. Forests. 2018; 9(1):14. https://doi.org/10.3390/f9010014
Chicago/Turabian StyleHan, Seung Hyun, Seongjun Kim, Guanlin Li, Hanna Chang, Soon Jin Yun, Jiae An, and Yowhan Son. 2018. "Effects of Warming and Precipitation Manipulation on Fine Root Dynamics of Pinus densiflora Sieb. et Zucc. Seedlings" Forests 9, no. 1: 14. https://doi.org/10.3390/f9010014
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.