1. Introduction
The status and dynamics of vegetation leaf area, often reported in terms of leaf area index (LAI), can be a critical determinant of regional air and water quality [
1]. LAI is commonly used as a surrogate of photosynthetically active area when photosynthesis is the principle process controlling chemical exchange between the atmosphere and underlying land surfaces. Leaf area influences the sequestration of carbon from carbon emissions [
2], the removal of pollutant species through deposition [
3], the biogenic emission of volatile organic compounds (BVOC) that contribute to tropospheric ozone formation [
4], and the emission of greenhouse gases [
5]. Temporally resolved leaf area and canopy heights for natural forest stands are critical inputs for process-based meteorological models. Leaf area estimates contribute to the calculation of surface evapotranspiration and albedo, and canopy height largely determines surface roughness, which contributes to mechanical mixing of the atmosphere [
6]. Calculation of gaseous air pollutant deposition velocity (V
d) frequently requires values for leaf area and surface roughness [
7,
8,
9]. The Clean Air Status and Trends Network (CASTNET), for instance, measures atmospheric concentrations and then estimates water vapor, ozone (O
3), sulfur dioxide (SO
2), and nitric acid (HNO
3) fluxes using the Multilayer Model (MLM) [
8,
10,
11]. The MLM inputs a generalized annual LAI time-series developed from measured values of maximum LAI for each plant species and typical phenology. Estimates of deposition velocity calculated by MLM were seen to be highly sensitive to LAI time-series parameters [
12] with differences in V
d of about 25% for sulfur dioxide and nitric acid and greater than 60% for ozone. On a regional scale, the United States Environmental Protection Agency’s (US EPA) Community Multiscale Air Quality Model (CMAQ) [
8] relies on output from the Weather Research Model (WRF) [
13] and the Pleim-Xiu (PX) land surface scheme [
14]. PX LAI estimates are based on deep soil temperature, an LAI response function based on soil temperature and a specified minimum and maximum LAI for each land use classification which results in known biases [
15].
LAI inputs into air quality applications that rely on model estimates of chemical flux include periodic in situ point sampled and Light Detection and Ranging (LIDAR) LAI measurements, static look-up values, and satellite-derived LAI values. A number of studies have reported that simulating both accurate leaf- and canopy-scale fluxes is not possible when leaf-scale fluxes are scaled using an inaccurate LAI [
16,
17,
18,
19]. While more temporally and spatially detailed LAI inputs are likely to improve model estimates of meteorological conditions, biogenic emissions, and O
3 precursor estimates, interspecies competition, age and spatial distribution make temporally resolved LAI estimates for minimally managed or natural forest stands particularly difficult to develop. Satellite-based LAI estimates hold promise for retrospective analyses, e.g., [
20,
21], but we are still learning how best to make use of these data, and they will never be available for future meteorological alternative management applications. Therefore, we must continue to supplement temporally resolved, remotely sensed vegetative leaf area estimates with numerical model estimates.
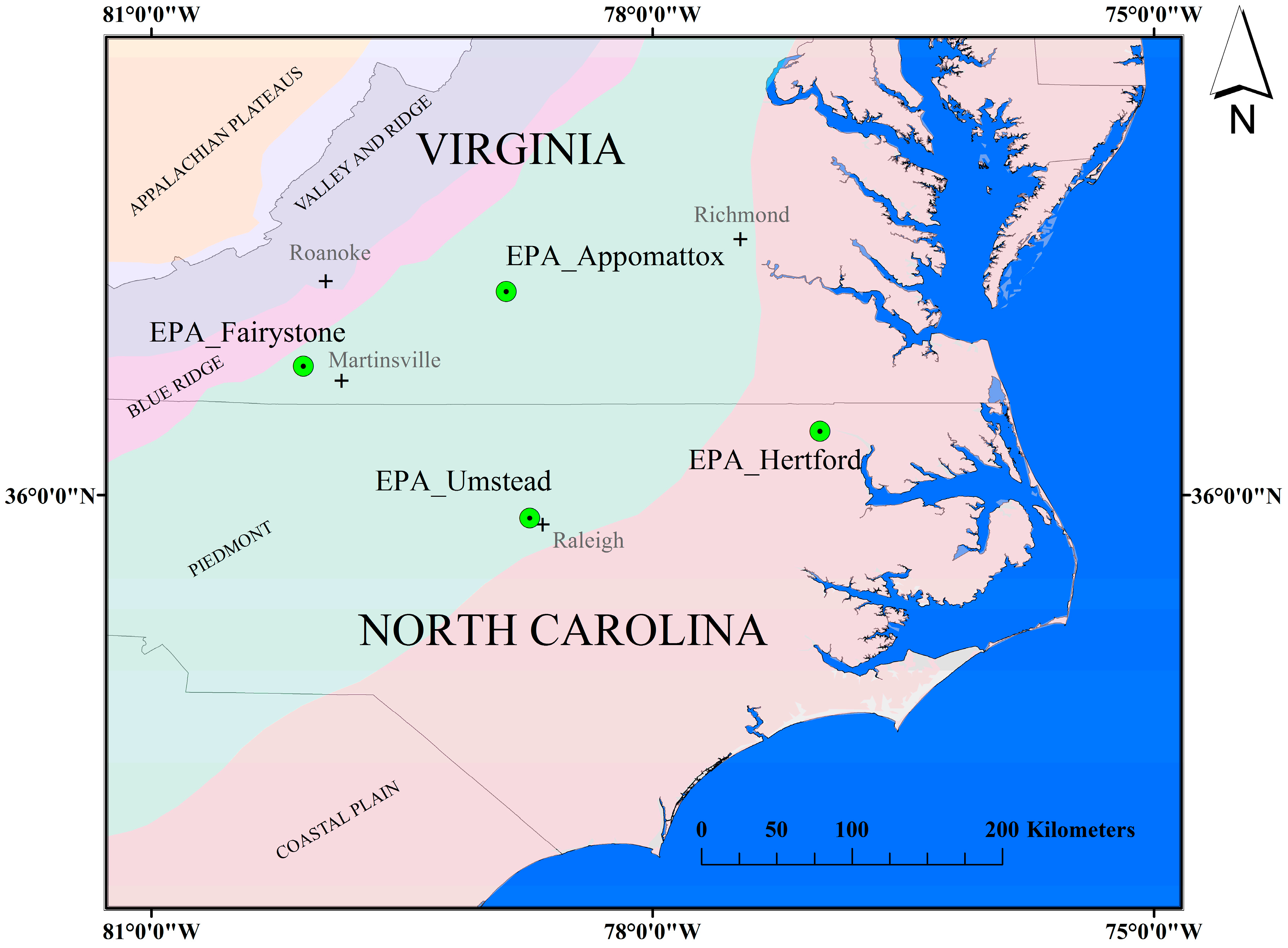
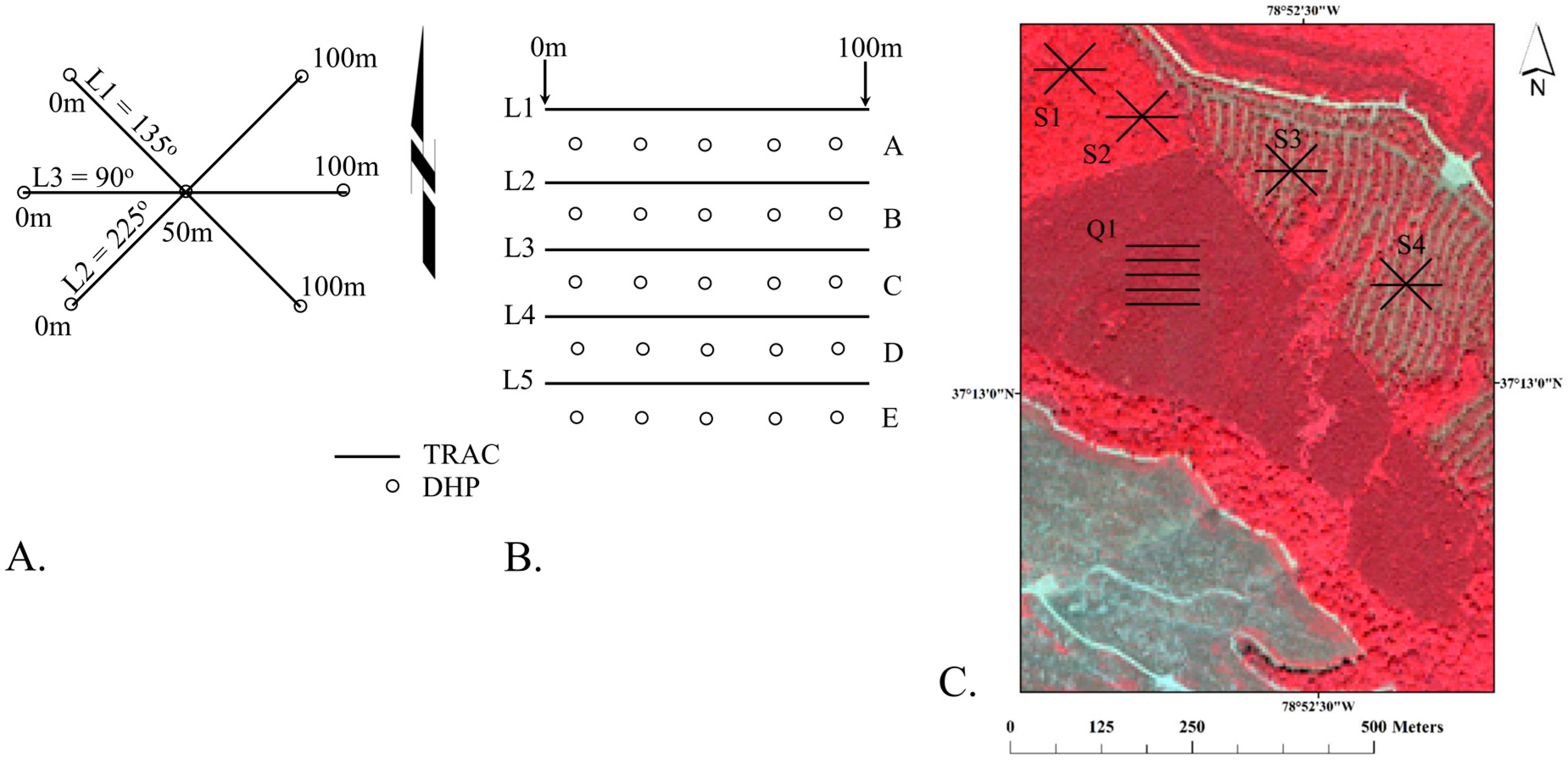
The objective of this study is to evaluate the capacity of a biogeochemical agroecosystems model to generate more biologically (process) representative, accurate and temporally resolved forest stand-level LAI estimates than existing numerical methods. We calibrate the USDA Environmental Policy Integrate Climate (EPIC) model from LAI estimates evaluated at four mixed forest stands in the southeastern U.S. We then demonstrate the practical areal application of this approach to represent mixed-age, mixed forest stands that could support nitrogen flux estimates for air quality assessment and supplement remotely sensed observations.
4. Discussion
While the results presented in
Section 3 suggest EPIC has an ability to simulate mixed forest stand LAI at the experimental plot scale, and previous EPIC applications have been performed for small watersheds, regional air quality models require simulations for much larger areas for which detailed information regarding stand characteristics needed by the EPIC model is often lacking. In addition, both experimental sites and SWAT land units often reflect relatively even-aged stands, i.e., the site is cleared at the start of an experiment. This will not be the case on a larger, regional basis and unmanaged or minimally managed stands. The discussion that follows illustrates one means of addressing these challenges.
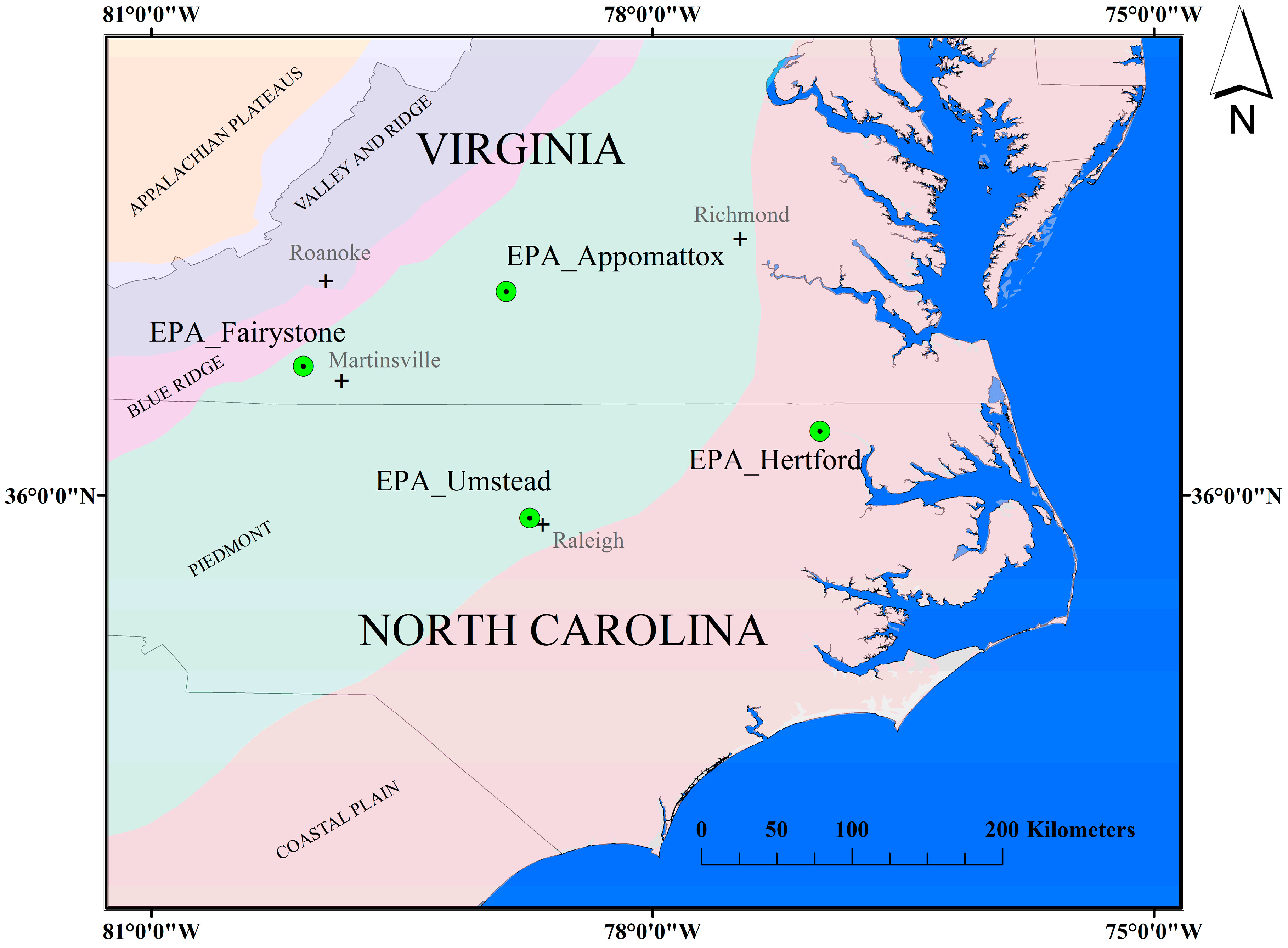
The EPIC model requires information regarding species, species density and species age. It then uses this information to model a single, homogeneous mixture of competing forest species. Neither characterization is likely to precisely represent any “real world” natural stand, but can serve to bound the range of possible stand-level LAI estimates. Both areal coverage and species density are rarely available for the same experimental site, but the US Forest Service Forest Inventory and Analysis (FIA) plot calculator (
http://apps.fs.fed.us/Evalidator/evalidator.jsp) can provide a consistent, reproducible estimate of species area and density information for a ~50 mi
2 (~130 km
2) area (4 mi or 6.4 km radius circle) surrounding the Appomattox field site (
Table 5). Approximately 57% of the area within this radius is reported as being forested.
The FIA summary data are aggregated into three species groupings: (1) Loblolly Pine and Virginia Pine (
Pinus virginiana) represented by Loblolly Pine; (2) mixed oak species comprised of Chestnut, Black and Red Oaks, represented by Black Oak; and (3) mixed upland hardwoods, represented by Red Maple. Stem densities for the groupings and stem fraction in trees per hectare (TPH) are 1349 (76%), 90 (5%) and 339 (19%). This compares to the Appomattox experimental plot represented by Loblolly Pine (26%), White Oak (34%), Red Maple (37%) and Sweetgum (3%). Stand densities are 1778 TPH for the FIA area stand compared to 4856 TPH for the Appomattox experimental plot. The FIA species age distribution suggests a 60-year simulation period with Black Oaks “planted” in year 1 (1942) of the simulation, Red Maples “planted” in year 20 (1961) of the simulation and Loblolly Pine “planted” in year 40 (1981) of the 60-year simulation. The FIA summary suggests relatively even ages within each species. If this were not the case, a range of ages within a species could be simulated by introducing (“planting”) new saplings each year within a desired establishment window. With some simplification, this use of FIA data is similar to that described for boreal forests [
29]. For instance, this research introduces a temporally dynamic function to simulate population dynamics that is missing from our example [
29] and other research suggests the use of temporally variable growth parameters to represent rapidly developing or “short-rotation” species [
39].
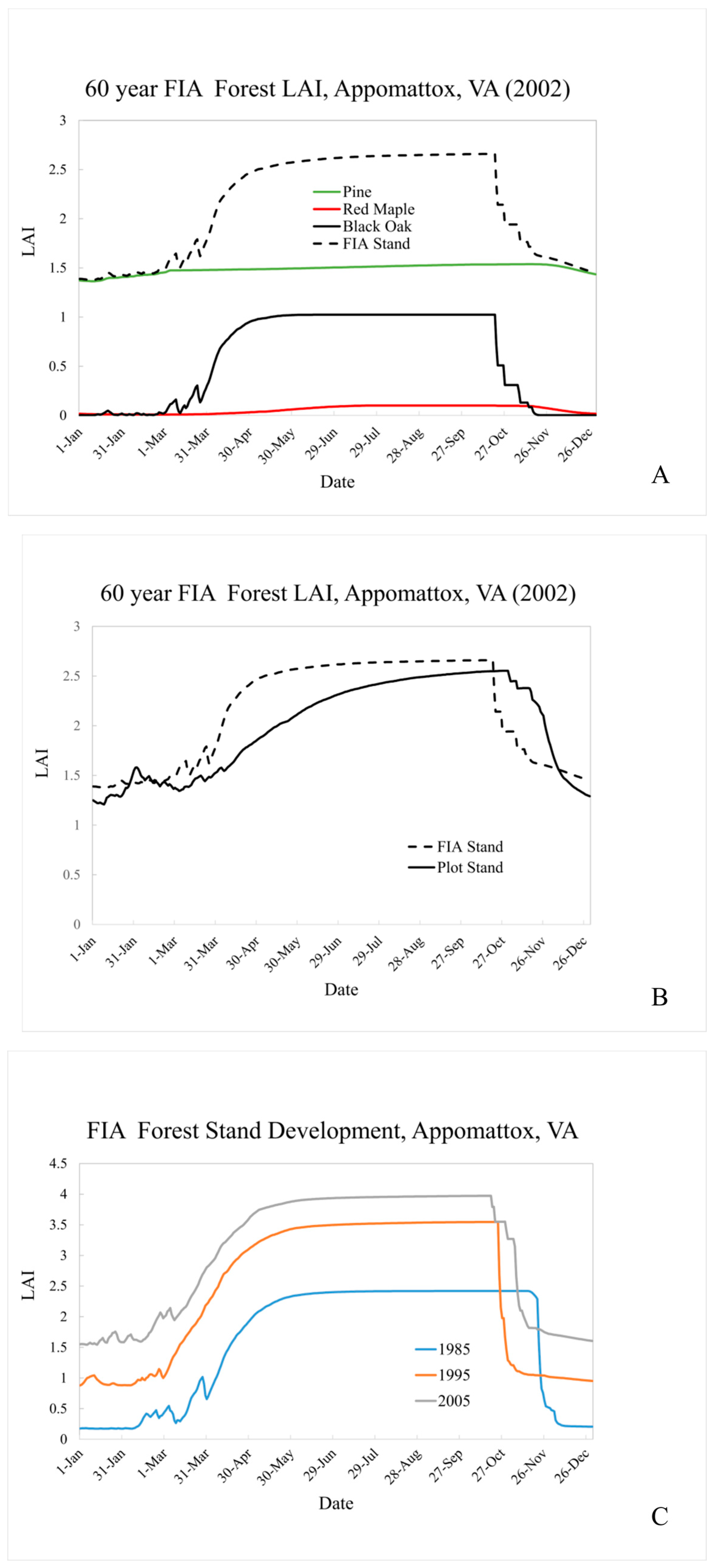
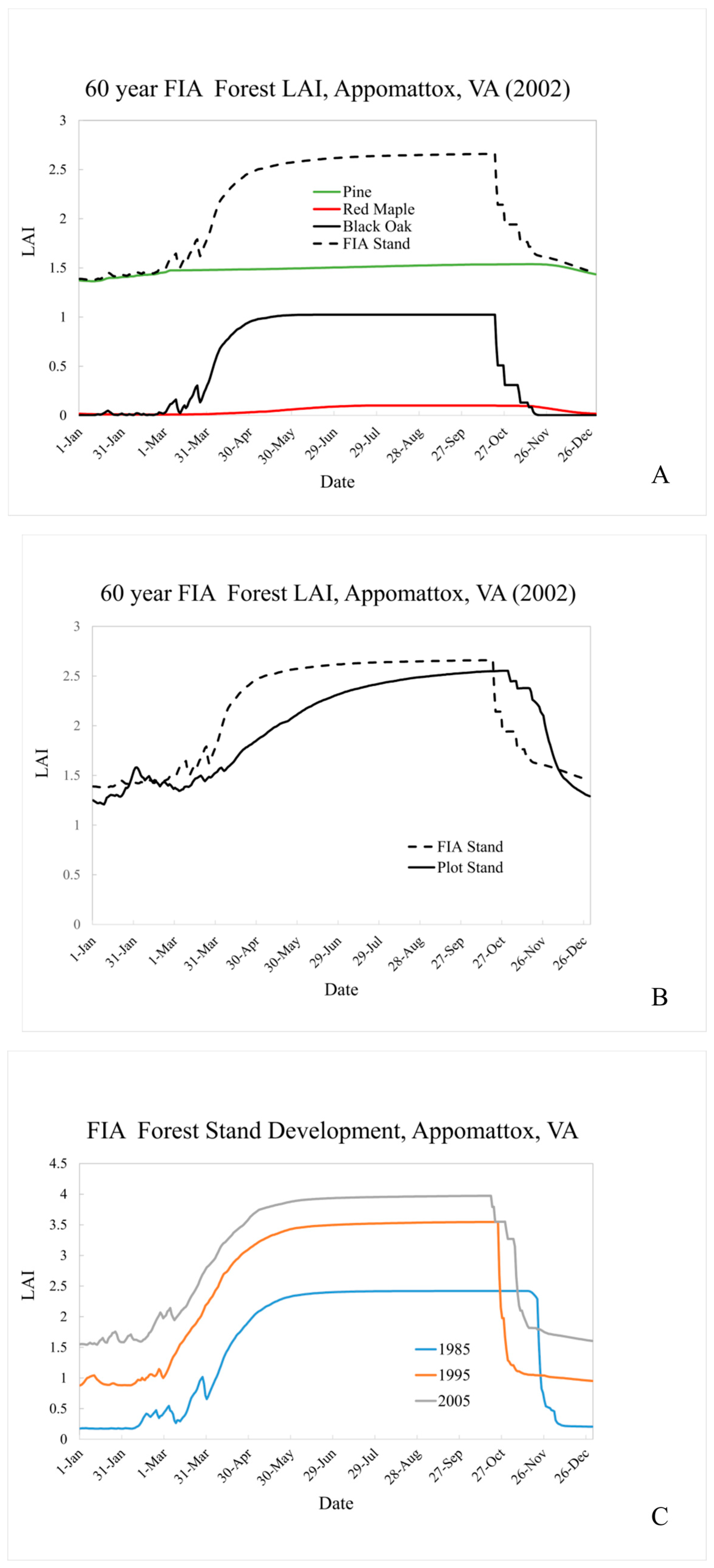
Figure 6A shows simulated LAI for the FIA area surrounding the Appomattox field site. For 2002 conditions, the substantial number of Pine provides a relatively constant base LAI of 1.5 throughout the winter months. Black Oak LAI provides the bulk of the stand seasonal signature, with green-up well under-way by April 1 and rapid brown-down (senescence) in late October.
Figure 6B compares our plot stand LAI in 2002 to our FIA representation. The seasonal peak LAI value is similar across stands, with the FIA stand’s greater maturity compensating for its lower overall stem density. The simulated Pine LAI is similar across the two simulations (plot value of ~1.2 and FIA value of ~1.4). This is, perhaps a bit surprising since pine are introduced into an existing hardwood stand for the FIA simulation but all species are “planted” simultaneously in the plot simulation. This result, however, appears to be supported by [
40] who report that hardwoods may facilitate longleaf pine seedling establishment, as opposed to suppressing it, at hardwood densities as high as 1400 stems·ha
−1. This simulated species interaction is explored further in
Figure 6C, which follows the FIA stand development from 1985 through 2005 (post pine introduction). Changes in stand level LAI magnitude and overall seasonal shape over time reflect simulated stand evolution and weather-driven variability. For instance, mean March minimum temperatures increase slightly between 1985 (~ 1°C) and 2005 (2.4°C). The simulated LAI seasonal pattern is guided by the accumulation of heat units above a species-specific “base” until a fixed annual total is reached. More rapid heat unit accumulation in the spring means that the annual total may be reached earlier in the year, effectively shifting brown-down initiation earlier as well. Changes such as these could be important for regional simulation of air quality since, as mentioned previously, LAI influences pollutant removal through gas phase deposition, and the magnitude and timing of pollutant precursor emissions can influence subsequent pollutant production and destruction.
An emerging research area to which improved areal LAI estimates could make a significant contribution is episodic land use change associated with wildland fires and prescribed burns used as part of land and resource management (
https://www.fs.fed.us/fire/).
Figure 7 illustrates the magnitude of the wildfire issue in terms of number of fires and total area burned. Air quality modeling research scientists are responding by explicitly including particulate emissions generated by these fires in their simulations (e.g., [
41]). The majority of these fire events are small, but 59 wildfire events, each of which were responsible for burning more than 100,000 acres (247,105 ha) were reported from January 2010 through December 2015. As our ability to simulate these events improves, it becomes important to include associated landscape changes such as surface exposure and forest stand re-growth in our estimates of subsequent pollutant deposition and precursor emission.
EPIC is an attractive option for capturing large-scale interactions between forest land covers and air quality because it is relatively easy to implement and has already been successfully coupled with a regional air quality model [
42]. There are, however, other forest ecosystem options that could be considered such as the LANDIS and LANDIS-II models [
43,
44]. It is always challenging to balance the desire for ecological and biogeochemical realism against the additional resources required for their inclusion. Additional research is needed to determine the level of process detail needed to adequately characterize these complex air and land surface interactions for regional air quality applications.
Finally, modeled LAI has the potential to augment satellite-derived estimates of this same parameter. In a perfect world, satellite estimates would provide reliable LAI in magnitude and without data drops, however this is almost never the case across all forested types and biomes. As an example, the Bigfoot MODIS Validation Project found agreement between validation data and MODIS LAI at low levels of LAI but was problematic at higher biomass levels [
45]. Also, they found significant differences dependent on the algorithm pathway chosen, which was a product of atmospheric interference (i.e., cloud contamination) and the number of quality scenes acquired in an 8-day chronosequence. EPIC or any other validated biomass model may work in combination with the satellite feed by constraining or inflating LAI values to reasonable figures based on biases observed in these validation studies [
46]. Thus, the modeled LAI could provide bias corrections where the satellite-derived LAI could provide the timing of green-up and senescence and relative seasonal changes in LAI.
5. Conclusions
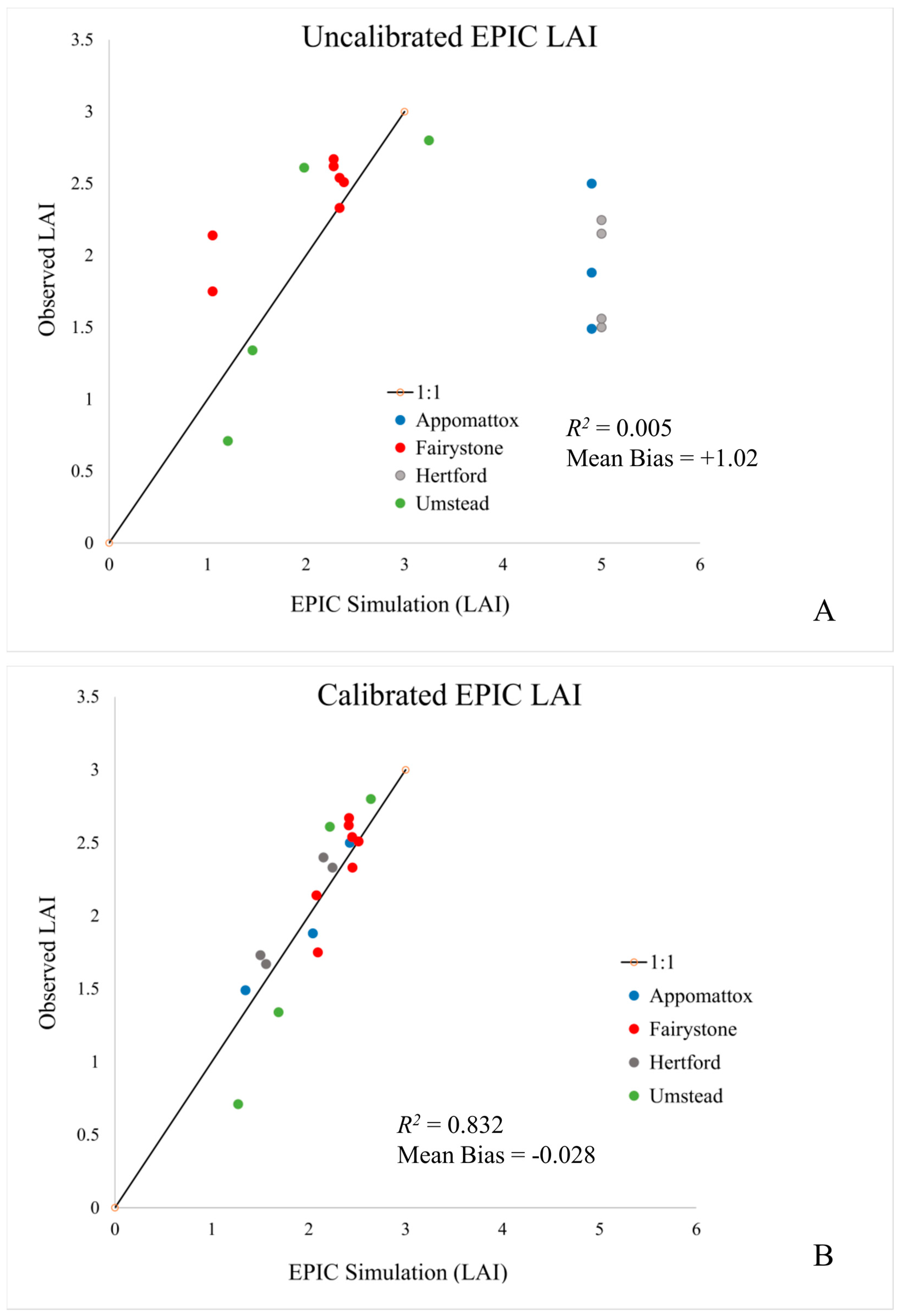
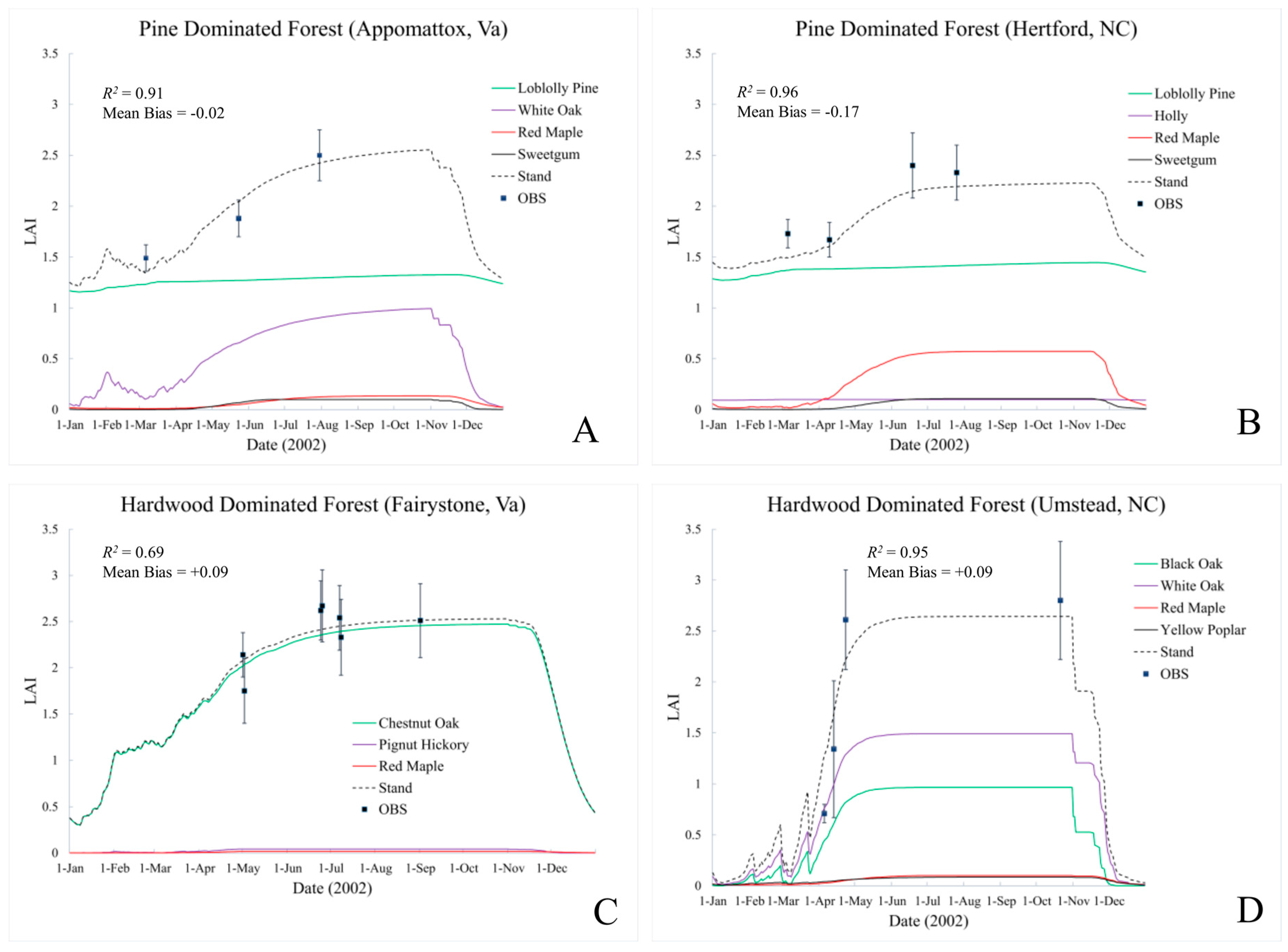
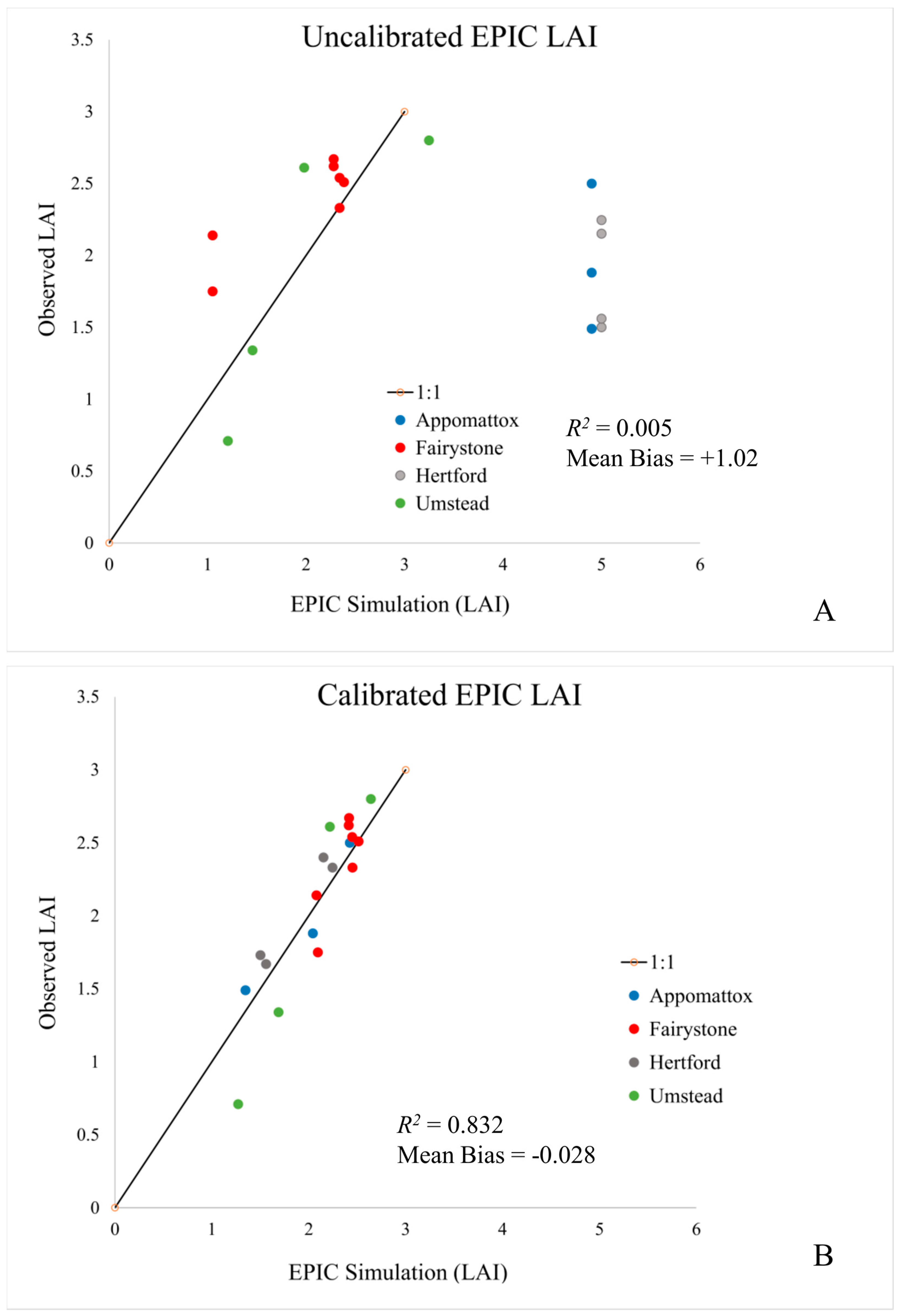
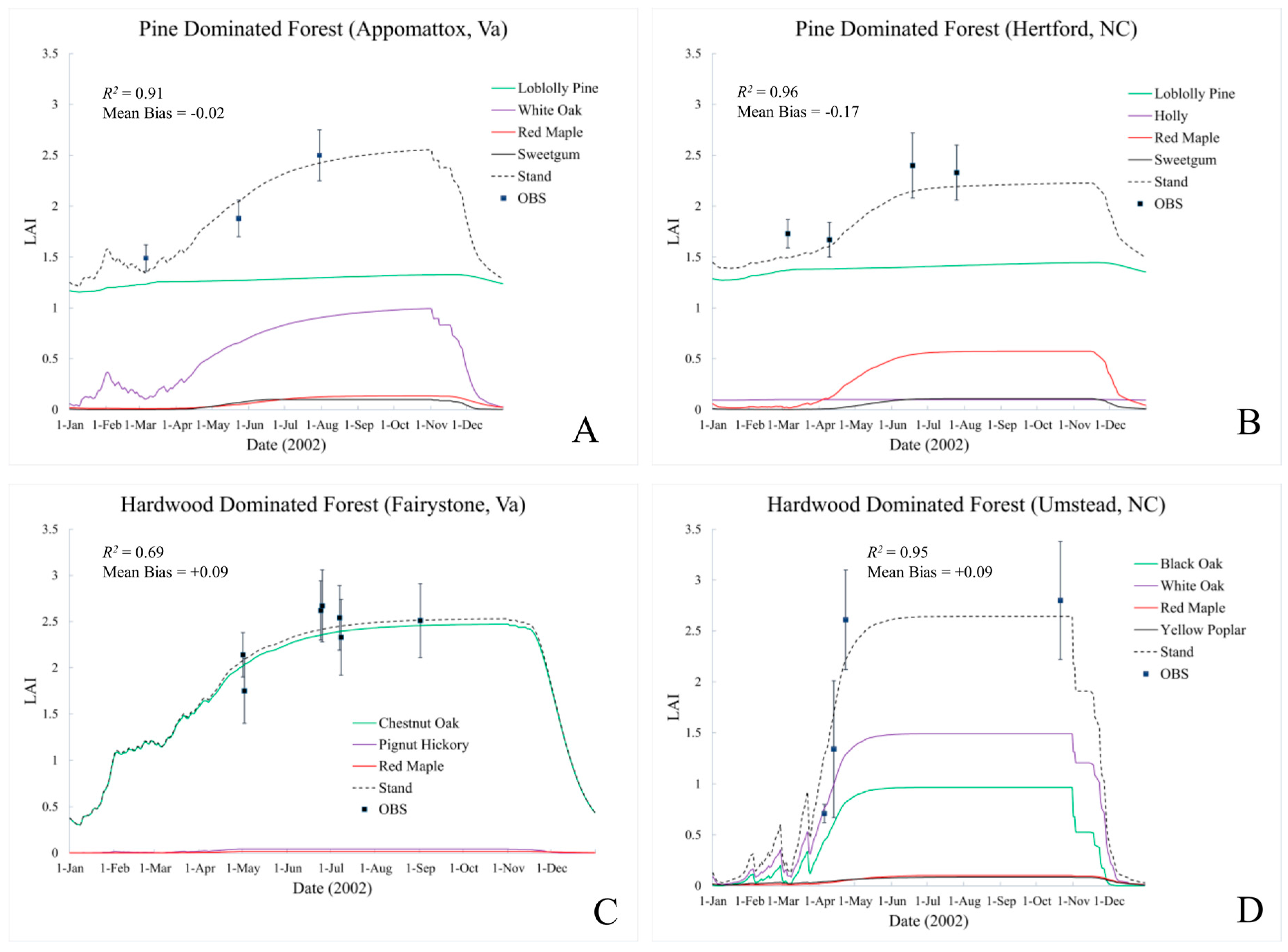
This study has explored the calibration of a semi-empirical, process-based biogeochemical model (EPIC) to estimate stand-level LAI and canopy height in four unfertilized mixed forest sites located in North Carolina and Virginia. Measurements of forest composition (species and number), LAI, diameter (DBH), height and basal area were recorded at each site during the 2002 field season. Calibration and verification results are good at these sites, with modeled to observed LAI correlations ranging from 0.61 to 0.96, correct identification of dominant species evolution, and dominant species (canopy) height estimate within 10% of observation (
Table 6). Across all sites and observations, calibration produces an
R2 value of 0.83 and mean bias of +0.008. These results suggest that EPIC offers an LAI estimate that agrees with observations while, at the same time better representing ecosystem processes and dynamic temporal patterns that respond to current and future meteorological conditions. All model calibrations reported here assume a uniform mixture of tree species. This appears to be a reasonable assumption for these stands, but more heavily managed mono-species stands are also easily simulated.
EPIC LAI estimates in regional air quality models such as CMAQ, but additional work is needed to operationalize this approach. For instance, we were able to calibrate and validate only a limited subset of species for a single geographic region. Calibration of additional species and validation across more geographically diverse settings and forest landscapes is needed, e.g., [
27]. While application of the EPIC LAI model across the continental U.S., a common CMAQ modeling domain, could require significant resources, this preliminary analysis suggests that it is technically feasible to define general eco-system-level stand composition profiles appropriate for larger geographic or ecological areas. Validation and evaluation of such areal estimates will be needed, but even satellite estimates of areal LAI are prone to error, so that a combination of remotely sensed data and measured atmospheric chemical fluxes may be appropriate. More temporally and spatially detailed characterization of vegetation cover should lead to more realistic atmospheric flux (precursor emission and chemical deposition) and ecosystem exposure estimation. Additional model evaluation is needed in order to better address long-term air quality trends and inter-annual variability that include emerging landscape drivers such as wildland and managed fires.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}