Predicting Shifts in the Suitable Climatic Distribution of Walnut (Juglans regia L.) in China: Maximum Entropy Model Paves the Way to Forest Management


Abstract
:1. Introduction
2. Materials and Methods
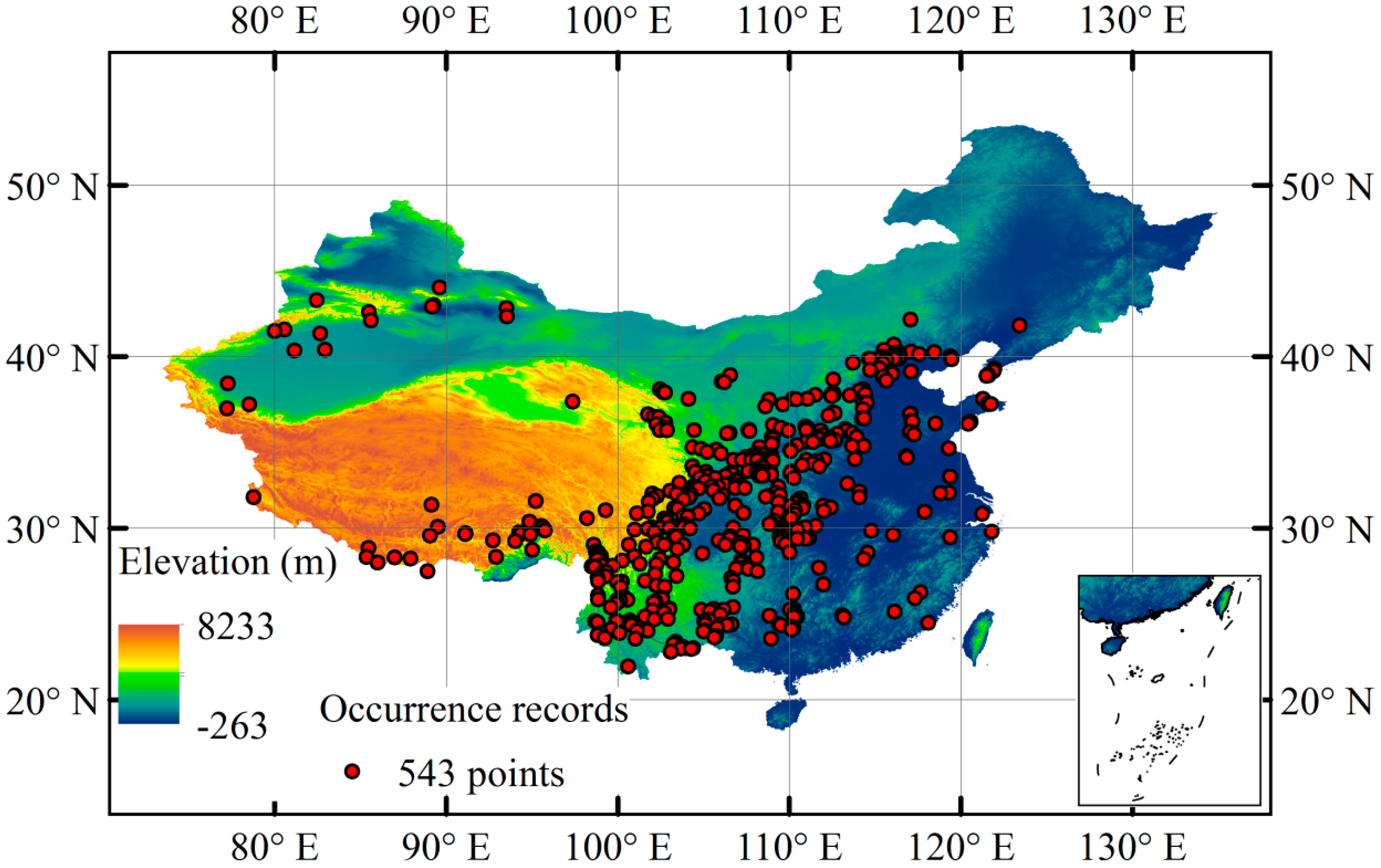
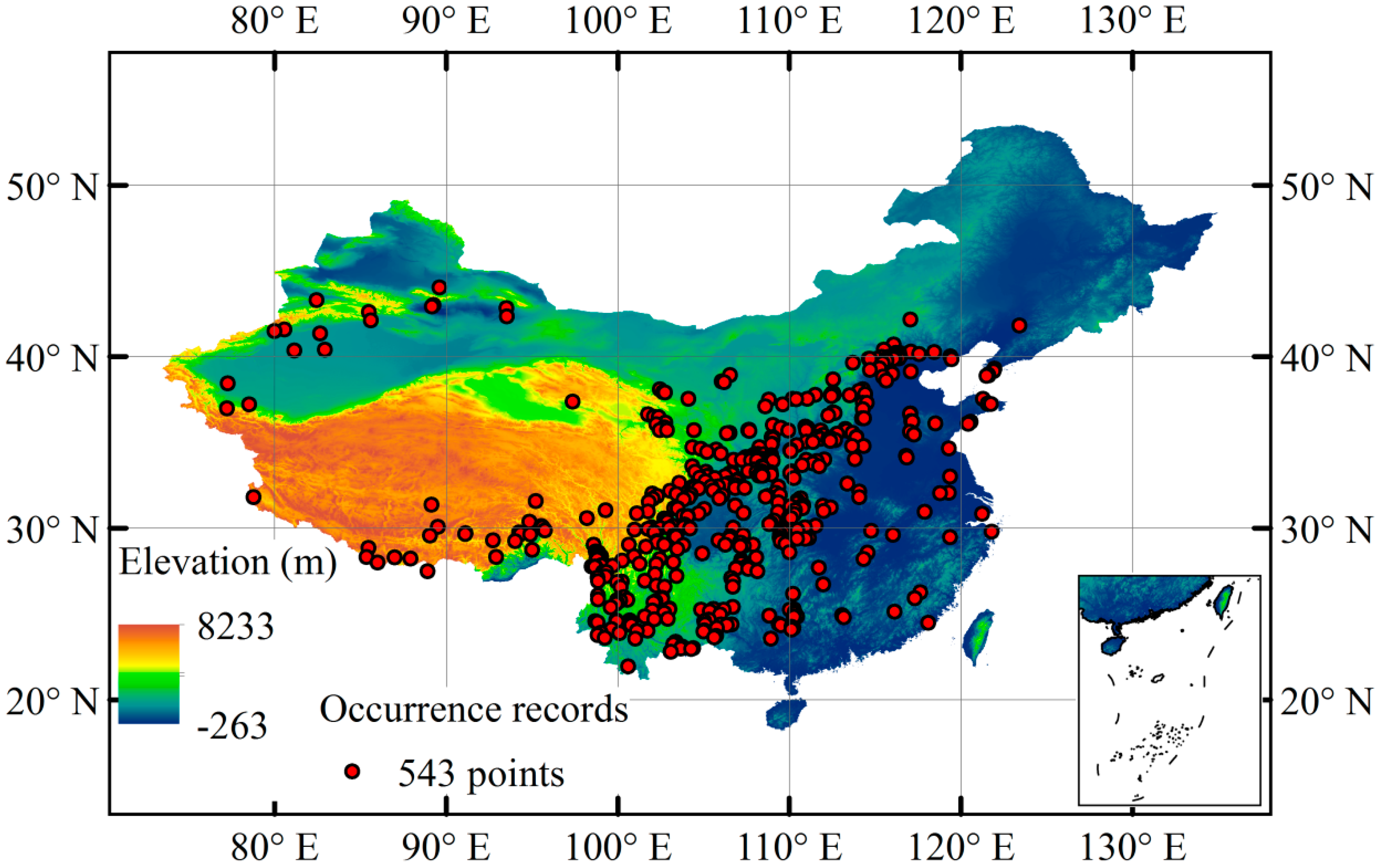
2.1. Species Occurrence Records
2.2. Environmental Variables
2.3. Correlation Analysis and Principal Component Analysis
2.4. MaxEnt Models
3. Results
3.1. Model Evaluation and Variables’ Contribution
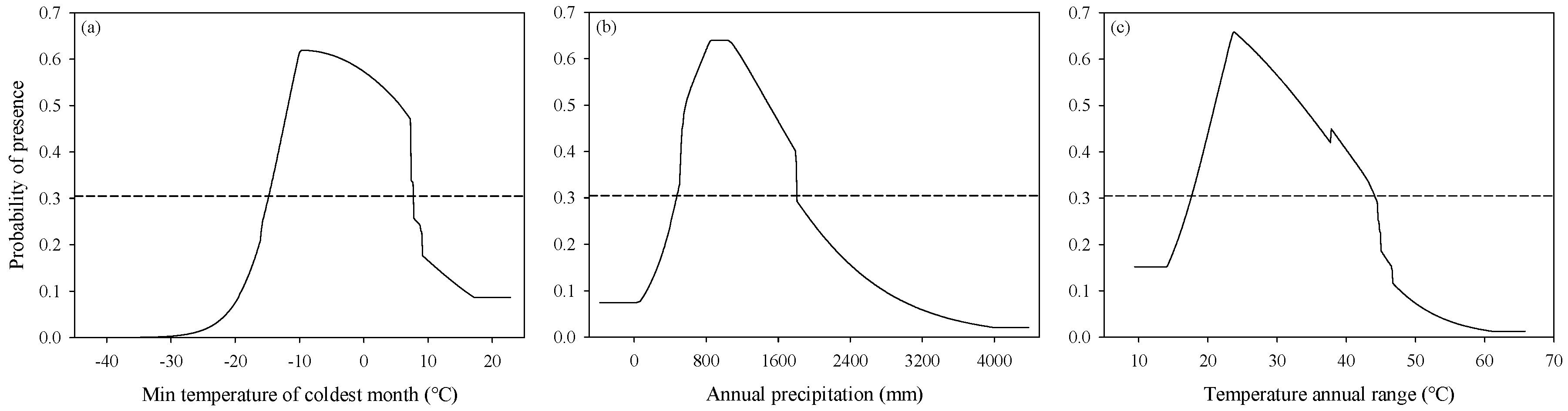
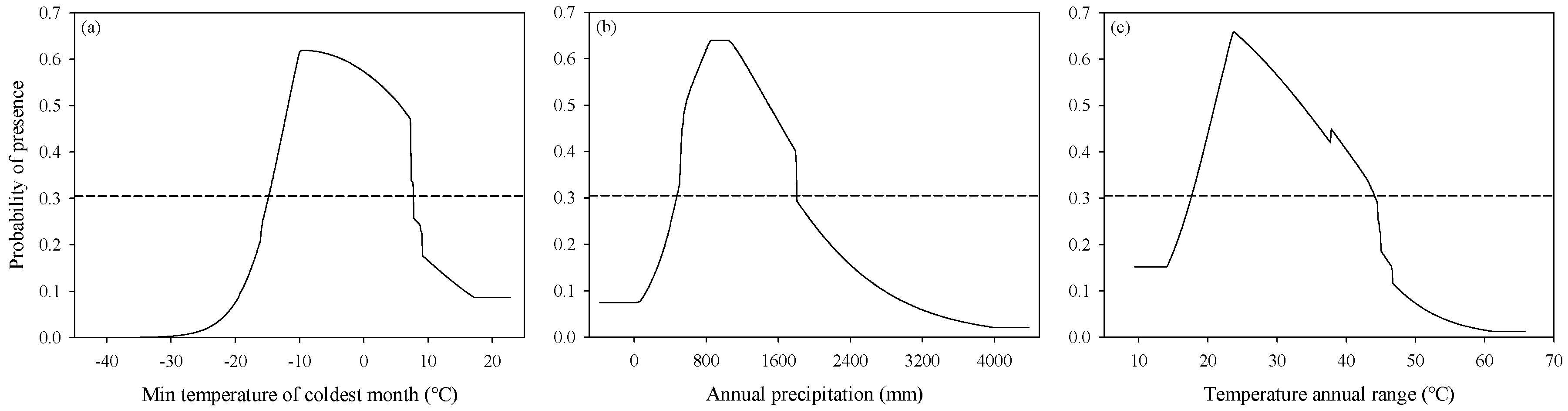
3.2. Response of Variables to Suitability
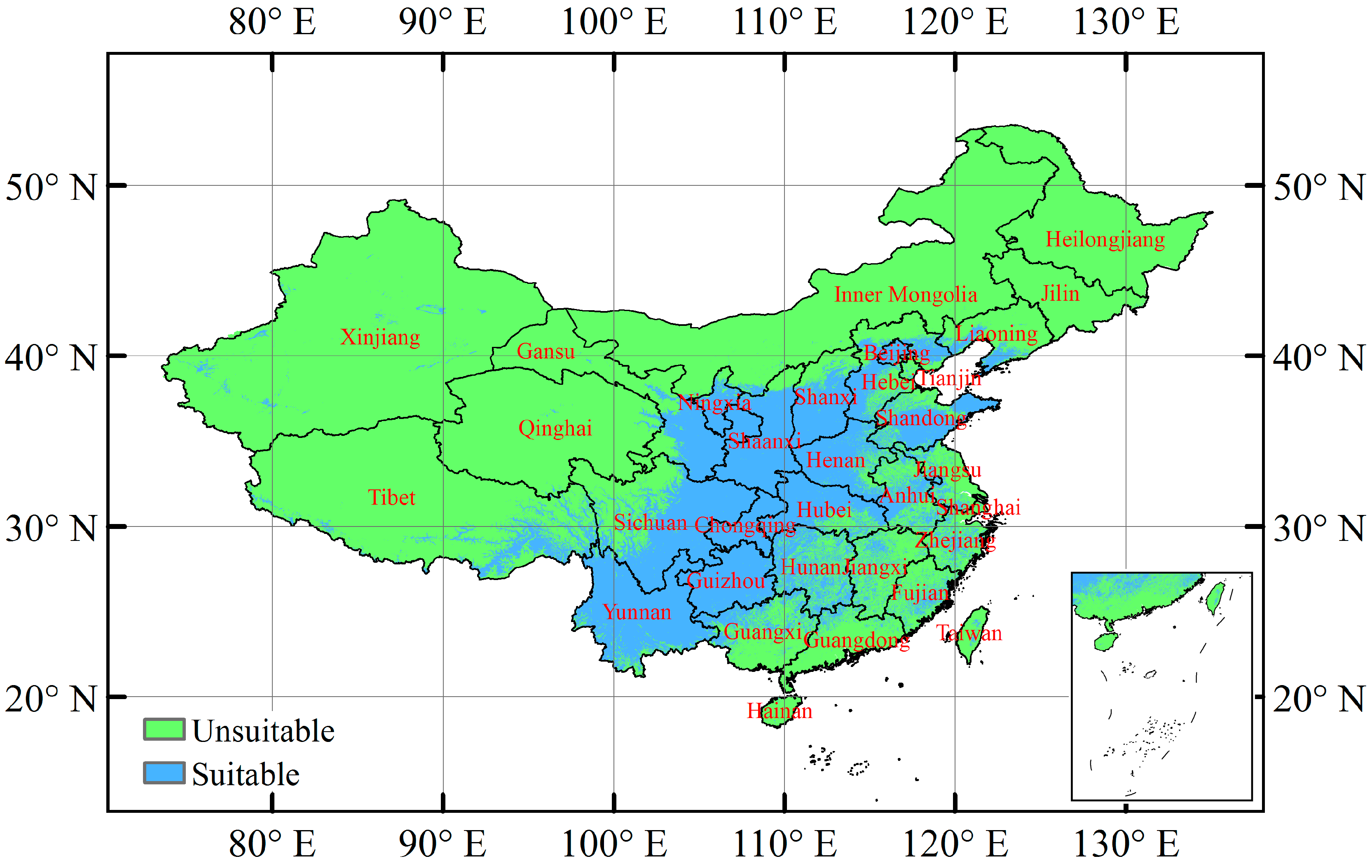
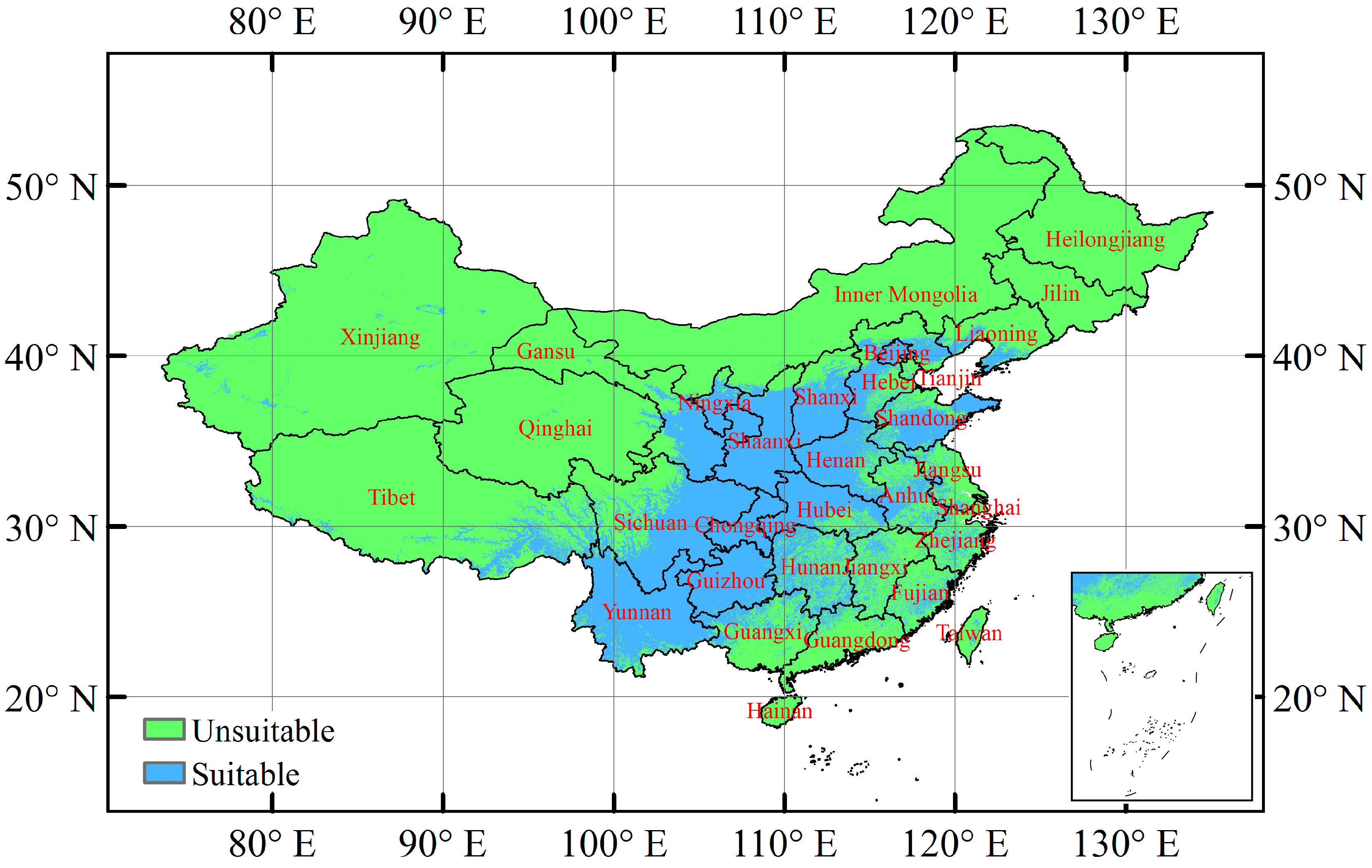
3.3. Current Potentially Suitable Climatic Distribution
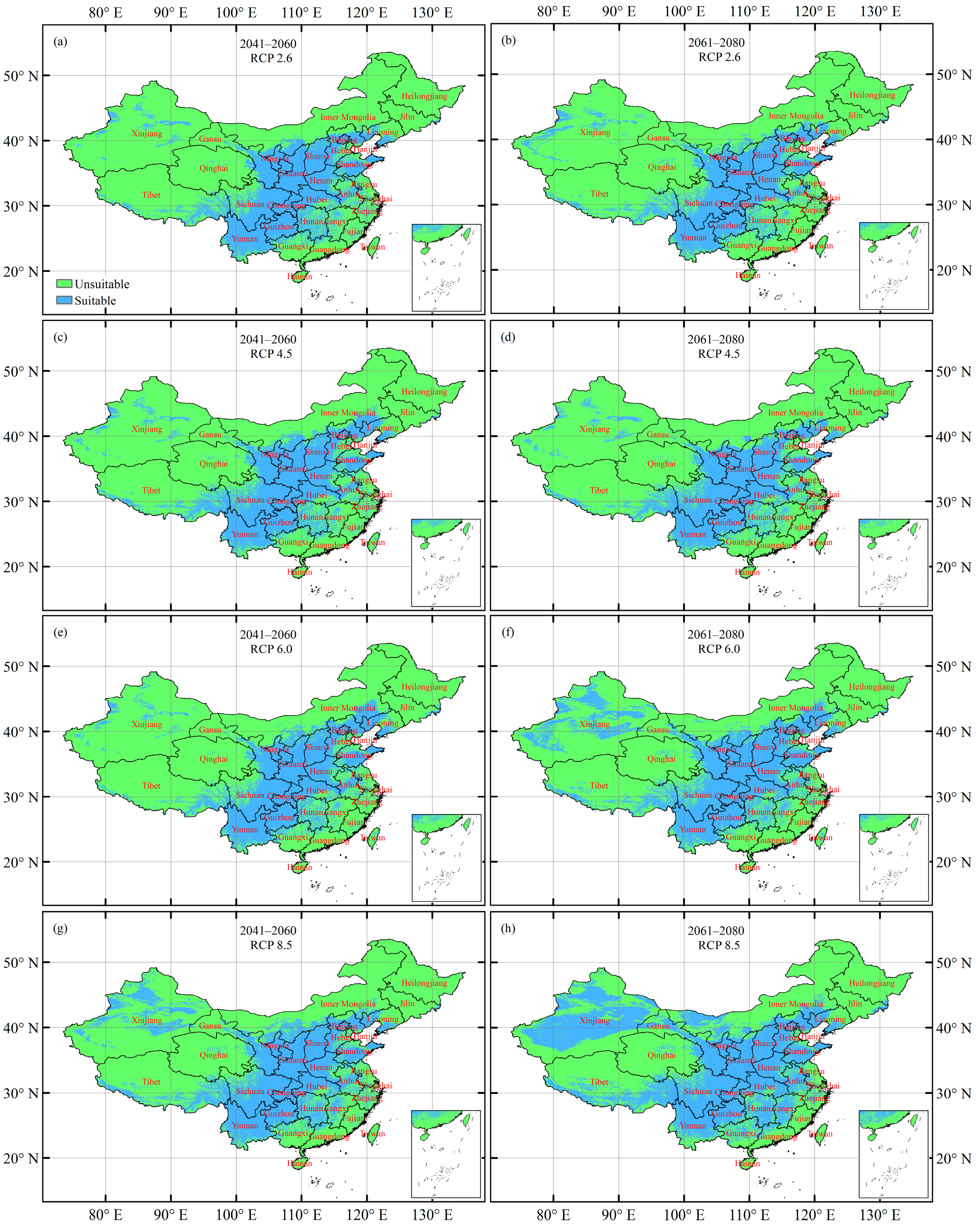
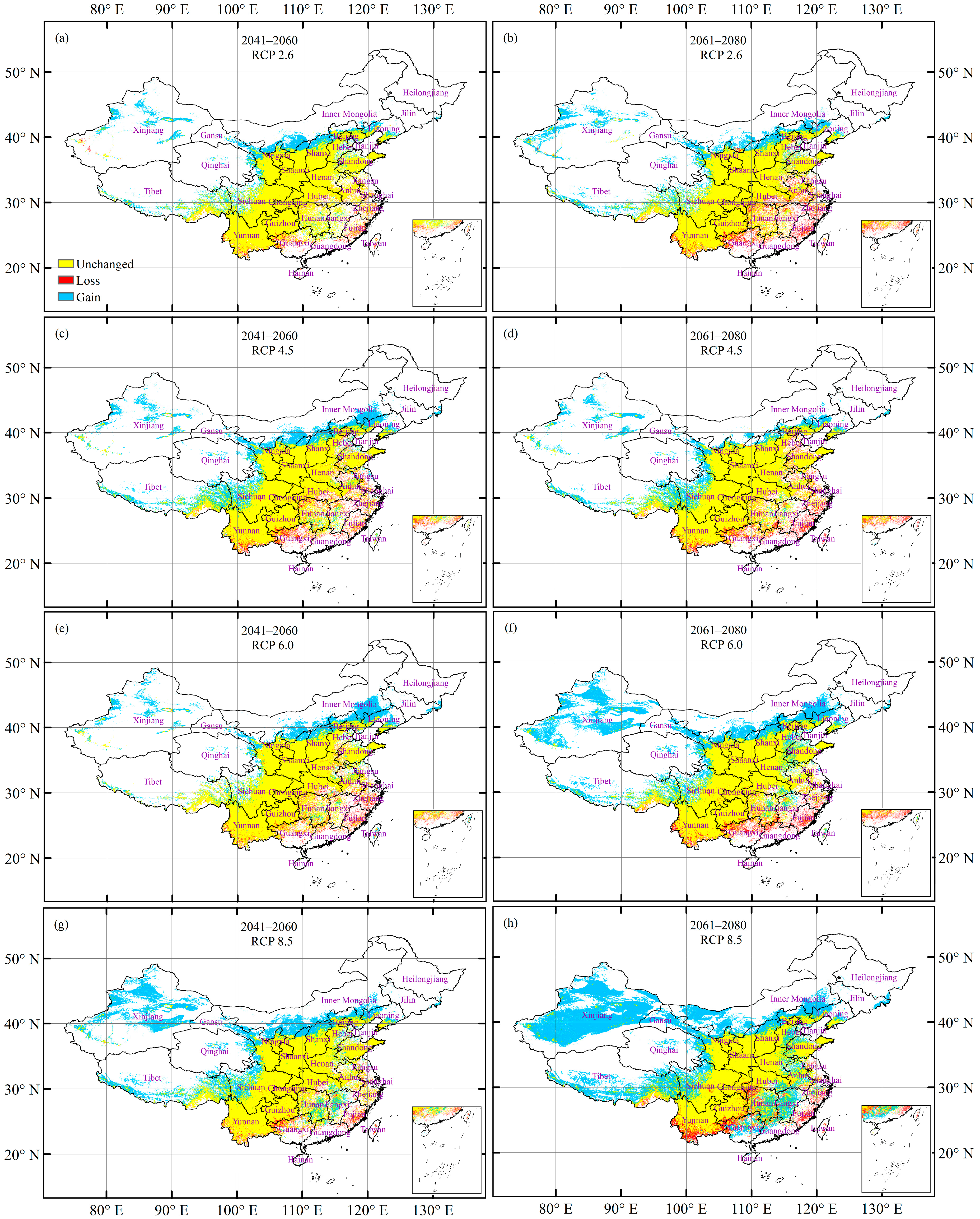
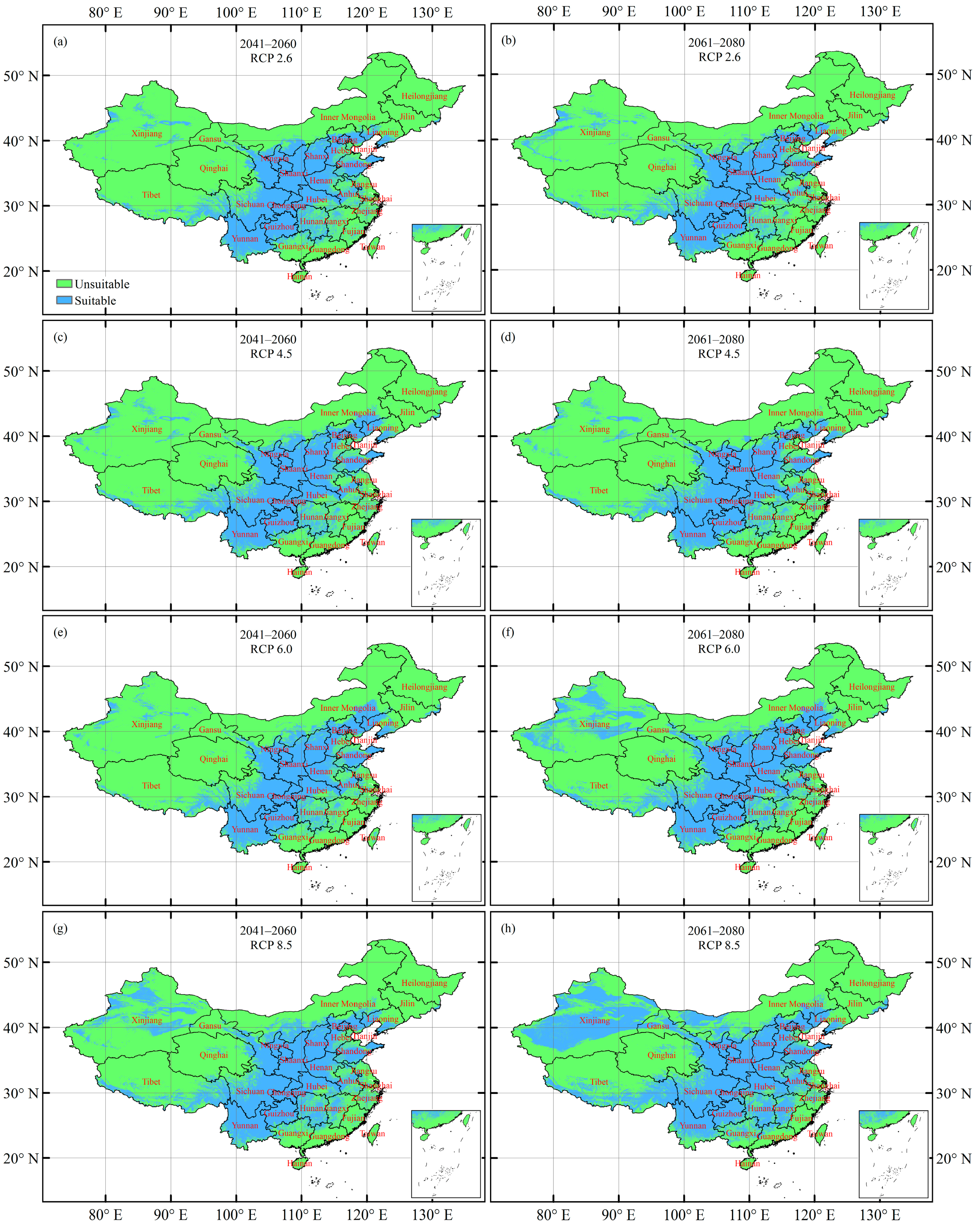
3.4. Future Potentially Suitable Climatic Distribution
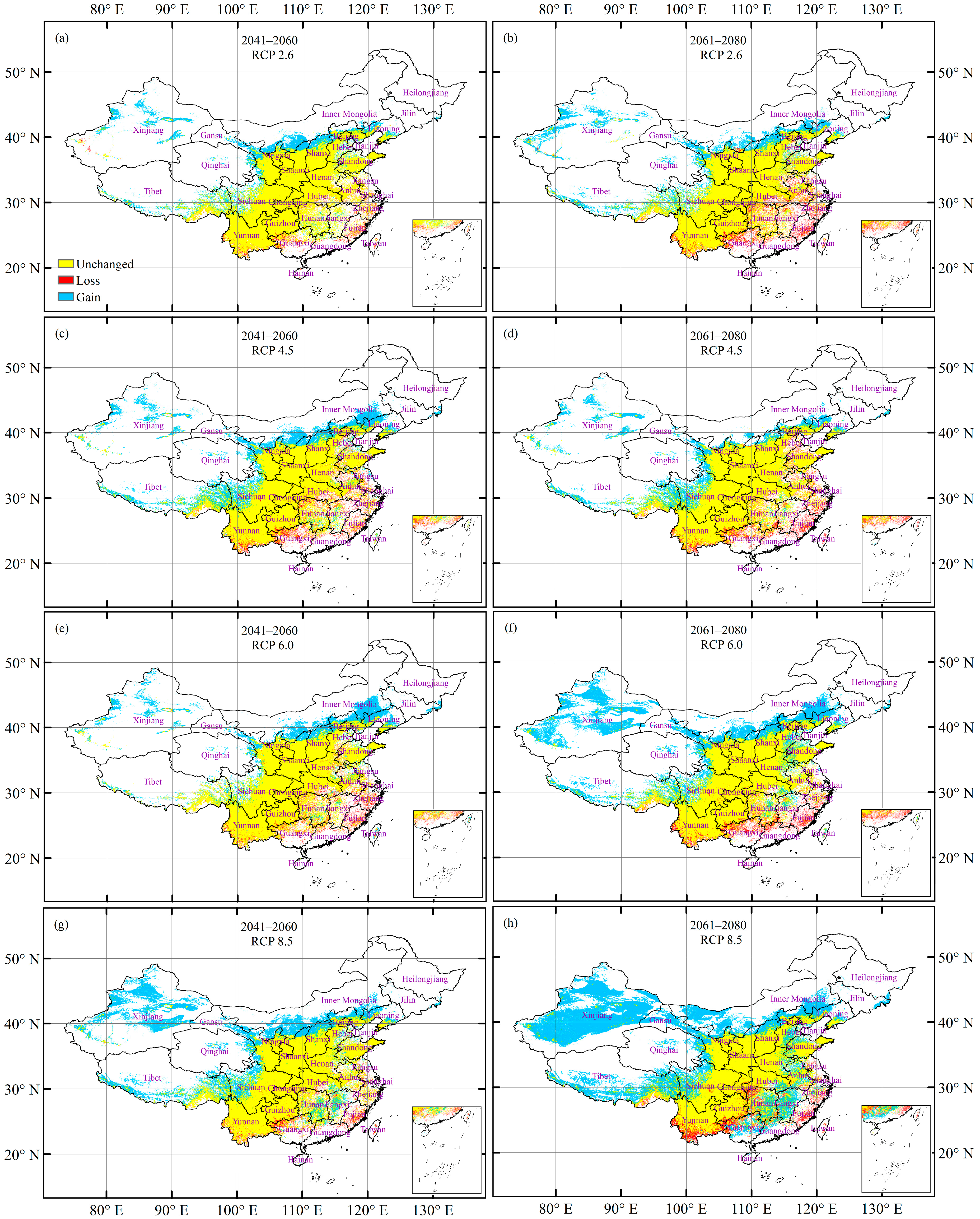
3.5. Future Changes in the Climatically Suitable Habitat Area
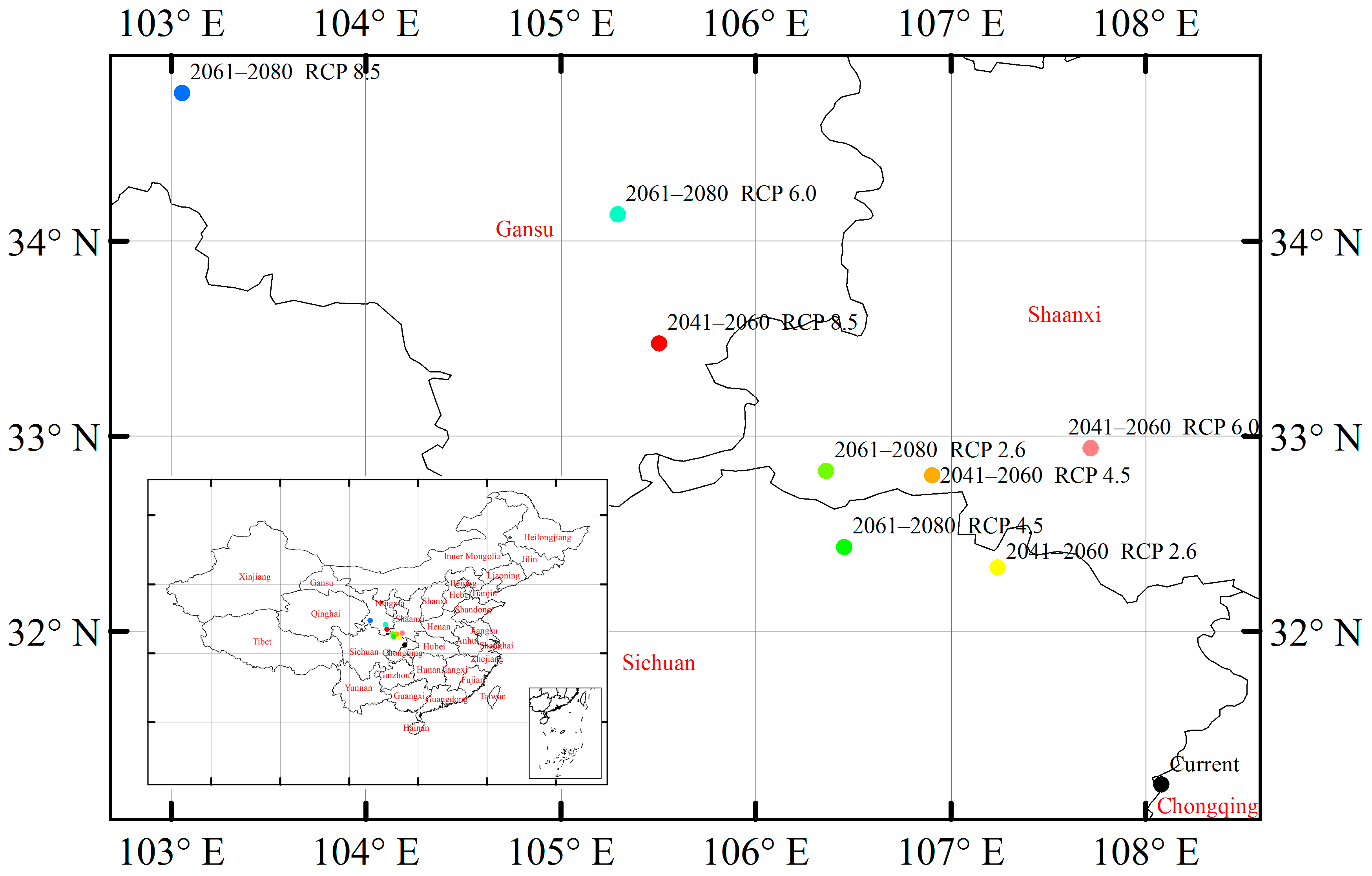
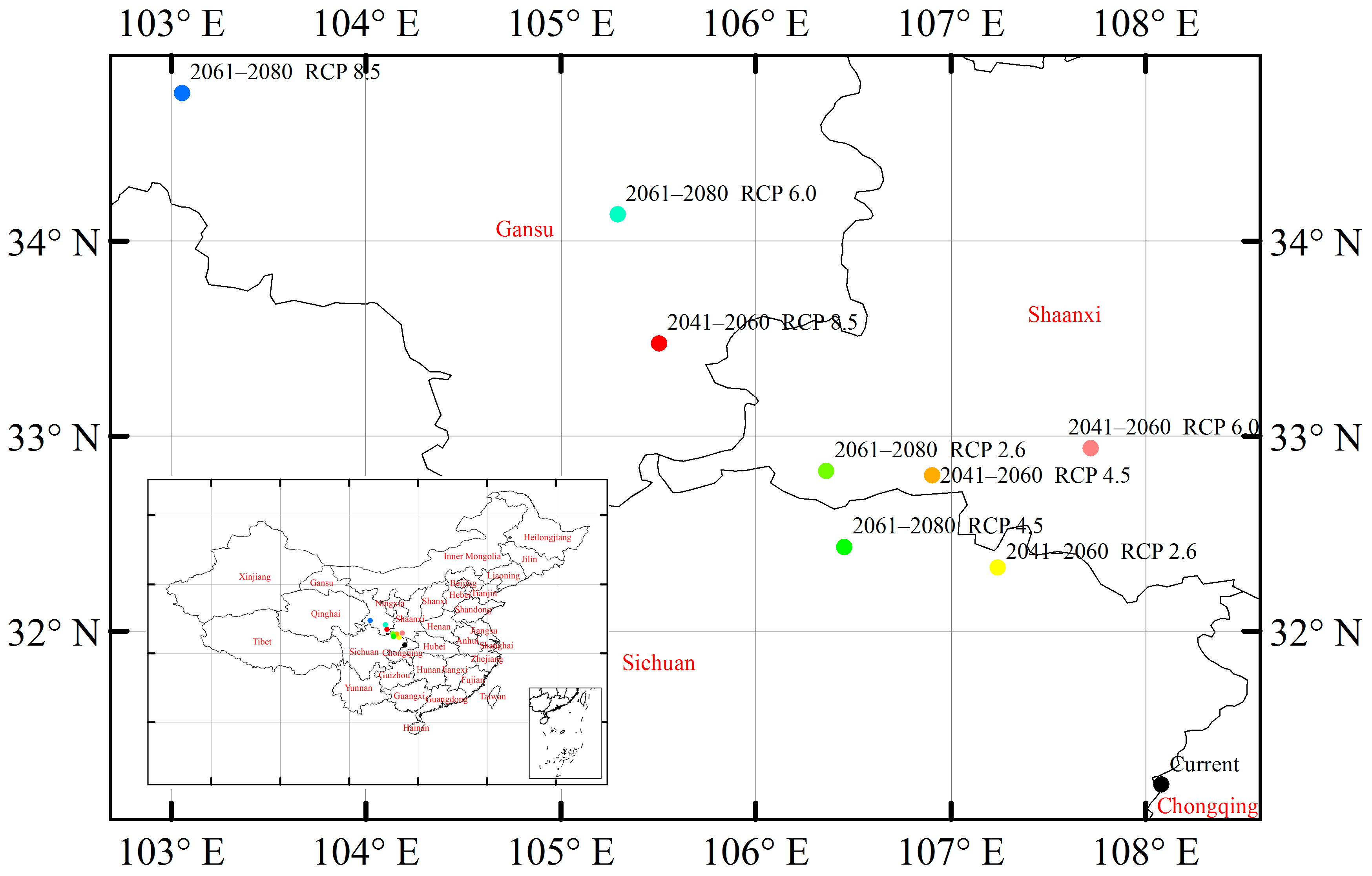
3.6. Suitable Climatic Habitat Shift
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| Bio1 | 0.282 | –0.234 | –0.214 |
| Bio2 | –0.199 | ≈0 | –0.276 |
| Bio4 | –0.196 | –0.374 | 0.246 |
| Bio5 | 0.102 | –0.491 | ≈0 |
| Bio6 | 0.316 | ≈0 | –0.171 |
| Bio7 | –0.239 | –0.325 | 0.137 |
| Bio8 | 0.149 | –0.444 | –0.153 |
| Bio9 | 0.302 | ≈0 | –0.248 |
| Bio10 | 0.152 | –0.460 | ≈0 |
| Bio11 | 0.308 | ≈0 | –0.272 |
| Bio12 | 0.307 | ≈0 | 0.206 |
| Bio13 | 0.274 | ≈0 | ≈0 |
| Bio16 | 0.293 | 0.114 | ≈0 |
| Bio17 | 0.239 | ≈0 | 0.526 |
| Bio18 | 0.279 | 0.134 | |
| Bio19 | 0.232 | ≈0 | 0.530 |
| Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| t_bulk_den | –0.389 | –0.11 | ≈0 |
| s_bulk_den | –0.422 | ≈0 | –0.185 |
| t_clay | –0.21 | 0.341 | –0.444 |
| s_clay | –0.21 | 0.341 | –0.444 |
| t_gravel | ≈0 | –0.112 | 0.137 |
| s_gravel | –0.177 | ≈0 | ≈0 |
| t_ph_h20 | –0.343 | ≈0 | 0.301 |
| s_ph_h20 | –0.419 | ≈0 | ≈0 |
| t_esp | –0.106 | –0.151 | 0.243 |
| s_esp | ≈0 | ≈0 | 0.242 |
| t_sand | –0.15 | –0.557 | –0.142 |
| s_sand | –0.263 | –0.447 | –0.197 |
| t_silt | –0.202 | 0.318 | 0.468 |
| s_silt | –0.321 | 0.311 | 0.239 |
| Variable | Bio5 | Bio6 | Bio7 |
|---|---|---|---|
| Bio6 | 0.530 | ||
| Bio7 | 0.128 | –0.773 | |
| Bio12 | 0.286 | 0.758 | –0.673 |
| Variable | t_bulk_den | s_bulk_den | t_clay | t_gravel | s_gravel | t_ph_h20 | t_esp | s_esp | t_sand | s_sand | t_silt |
|---|---|---|---|---|---|---|---|---|---|---|---|
| s_bulk_den | 0.415 | ||||||||||
| t_clay | 0.181 | 0.648 | |||||||||
| t_gravel | 0.177 | –0.320 | –0.277 | ||||||||
| s_gravel | 0.175 | 0.550 | 0.341 | 0.364 | |||||||
| t_ph_h20 | 0.707 | 0.345 | 0.108 | –0.033 | 0.023 | ||||||
| t_esp | 0.083 | 0.189 | 0.141 | –0.084 | 0.064 | 0.305 | |||||
| s_esp | 0.058 | 0.192 | 0.176 | –0.220 | –0.020 | 0.295 | 0.772 | ||||
| t_sand | 0.460 | –0.021 | –0.451 | 0.273 | –0.027 | 0.132 | –0.051 | –0.109 | |||
| s_sand | 0.363 | 0.748 | 0.133 | –0.175 | 0.399 | 0.219 | 0.127 | 0.098 | 0.535 | ||
| t_silt | 0.251 | -0.016 | 0.121 | 0.017 | 0.003 | 0.440 | 0.057 | 0.145 | –0.534 | –0.479 | |
| s_silt | 0.241 | 0.732 | 0.603 | –0.311 | 0.399 | 0.429 | 0.197 | 0.232 | –0.522 | 0.190 | 0.569 |
References
- IPCC. IPCC Climate Change 2013: The Physical Science Basis. Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change—Summary for Policymakers; IPCC: Geneva, Switzerland, 2013. [Google Scholar]
- Despland, E.; Houle, G. Climate influences on growth and reproduction of Pinus banksiana (Pinaceae) at the limit of the species distribution in eastern North America. Am. J. Bot. 1997, 84, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; De, R.P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofríodillon, G.; De, R.P.; Vidal, C.; Pierrat, J.C.; Gégout, J.C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517. [Google Scholar] [CrossRef] [PubMed]
- Dion, P.-P.; Bussières, J.; Lapointe, L. Sustainable leaf harvesting and effects of plant density on wild leek cultivation plots and natural stands in Southern Quebec, Canada. Agrofor. Syst. 2016, 90, 979–995. [Google Scholar] [CrossRef]
- Lu, C.Y.; Gu, W.; Dai, A.H.; Wei, H.Y. Assessing habitat suitability based on geographic information system (GIS) and fuzzy: A case study of Schisandra sphenanthera Rehd. et Wils. in Qinling Mountains, China. Ecol. Model. 2012, 242, 105–115. [Google Scholar] [CrossRef]
- Guo, Y.; Wei, H.; Lu, C.; Gao, B.; Gu, W. Predictions of potential geographical distribution and quality of Schisandra sphenanthera under climate change. PeerJ 2016, 4, e2554. [Google Scholar] [CrossRef] [PubMed]
- Hakl, J.; Fuksa, P.; Konečná, J.; Pacek, L.; Tlustoš, P. Effect of applied cultivation technology and environmental conditions on lucerne farm yield in the Central Europe. Plant Soil Environ. 2014, 60, 475–480. [Google Scholar] [CrossRef]
- Lamb, J.F.S.; Jung, H.-J.G.; Riday, H. Growth environment, harvest management and germplasm impacts on potential ethanol and crude protein yield in alfalfa. Biomass Bioenergy 2014, 63, 114–125. [Google Scholar] [CrossRef]
- Anderson, R.P. A framework for using niche models to estimate impacts of climate change on species distributions. Ann. N. Y. Acad. Sci. 2013, 8–28. [Google Scholar] [CrossRef] [PubMed]
- Martin, Y.; Titeux, N. Testing instead of assuming the importance of land use change scenarios to model species distributions under climate change. Glob. Ecol. Biogeogr. 2013, 22, 1204–1216. [Google Scholar] [CrossRef]
- Qin, Z.; Zhang, J.E.; Ditommaso, A.; Wang, R.L.; Liang, K.M. Predicting the potential distribution of Lantana camara L. under RCP scenarios using ISI-MIP models. Clim. Chang. 2016, 134, 193–208. [Google Scholar] [CrossRef]
- Wang, H.H.; Wonkka, C.L.; Grant, W.E.; Rogers, W.E. Potential range expansion of Japanese honeysuckle (Lonicera japonica thunb.) in southern U.S. forestlands. Forests 2012, 3, 573–590. [Google Scholar] [CrossRef]
- Wang, W.; Tang, X.; Zhu, Q.; Pan, K.; Hu, Q.; He, M.; Li, J. Predicting the impacts of climate change on the potential distribution of major native non-food bioenergy plants in China. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.-S.; Wei, Y.-L.; Wang, X.-G. Maxent modeling for predicting the potential distribution of Sanghuang, an important group of medicinal fungi in China. Fungal Ecol. 2015, 17, 140–145. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Mapping the global potential geographical distribution of black locust (Robinia Pseudoacacia L.) using herbarium data and a maximum entropy model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, H.; Xie, T.; Xu, Y.; Zhao, L.; Tian, W. Effects of climate change on the potentially suitable climatic geographical range of Liriodendron chinense. Forests 2017, 8, 399. [Google Scholar] [CrossRef]
- Rundel, P.W. Modeled shifts in Polylepis species ranges in the Andes from the Last Glacial Maximum to the present. Forests 2017, 8, 232. [Google Scholar]
- Deb, J.C.; Phinn, S.; Butt, N.; Mcalpine, C.A. Climatic-induced shifts in the distribution of Teak (Tectona grandis) in tropical Asia: Implications for forest management and planning. Environ. Manag. 2017, 60, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Dai, G.; Yang, J.; Huang, C.; Sun, C.; Jia, L.; Ma, L. The effects of climate change on the development of tree plantations for biodiesel production in China. Forests 2017, 8, 207. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Zhang, Y.; Yuan, S.; Wang, X.; Feng, B.; Zhang, R.; Han, J. Analysis and reflection on development strategy of walnut processing industry in China. Trans. Chin. Soc. Agric. Eng. 2015, 31, 1–8, (in Chinese with English abstract). [Google Scholar]
- Crews, C.; Hough, P.; Godward, J.; Brereton, P.; Lees, M.; Guiet, S.; Winkelmann, W. Study of the main constituents of some authentic walnut oils. J. Agric. Food Chem. 2005, 53, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Xi, R.T. The textual research on the origin of Chinese walnut (Juglans regia L.). J. Hebei Univ. Agric. 1990, 13, 89–94, (in Chinese with English abstract). [Google Scholar]
- USDA-FAS. Oilseeds: World Markets and Trade, 07-10-2015. 2015. Available online: http://usda.mannlib.cornell.edu/usda/fas/oilseed-trade//2010s/2015/oilseed-trade-07-10-2015.pdf.
- Yuan, S.J. Developing situation and countermeasures of Shaanxi walnut industry. Shaanxi For. 2009, 6, 10, (In Chinese with English abstract). [Google Scholar]
- Chinese Virtual Herbarium. Available online: http://www.cvh.ac.cn/ (accessed on 24 July 2017).
- Global Biodiversity Information Facility. Available online: http://www.gbif.org/ (accessed on 24 July 2017).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Varela, S.; Lima-Ribeiro, M.S.; Terribile, L.C. A short guide to the climatic variables of the last glacial maximum for biogeographers. PLoS ONE 2015, 10, e0129037. [Google Scholar] [CrossRef] [PubMed]
- Bedia, J.; Herrera, S.; Gutiérrez, J.M. Dangers of using global bioclimatic datasets for ecological niche modeling. Limitations for future climate projections. Glob. Planet. Chang. 2013, 107, 1–12. [Google Scholar] [CrossRef]
- Fischer, G.; Nachtergaele, F.; Prieler, S.; Van Velthuizen, H.T.; Verelst, L.; Wiberg, D. Global Agro-Ecological Zones Assessment for Agriculture (GAEZ 2008); IIASA: Laxenburg, Austria; FAO: Rome, Italy, 2008. [Google Scholar]
- Xin, X.; Zhang, L.; Zhang, J.; Wu, T.; Fang, Y. Climate change projections over East Asia with BCC_CSM1.1, climate model under RCP scenarios. J. Meteorol. Soc. Japan 2013, 91, 413–429. [Google Scholar] [CrossRef]
- Rana, S.K.; Rana, H.K.; Ghimire, S.K.; Shrestha, K.K.; Ranjitkar, S. Predicting the impact of climate change on the distribution of two threatened Himalayan medicinal plants of Liliaceae in Nepal. J. Mt. Sci. 2017, 14, 558–570. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, Justicia adhatoda L. in Lesser Himalayan foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Manel, S.; Williams, H.C.; Ormerod, S.J. Evaluating presence-absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold criteria for conversion of probability of species presence to either—or Presence—Absence. Acta Oecol. 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Kissling, W.D.; Carvalheiro, L.G.; WallisDeVries, M.F.; Franzén, M.; Biesmeijer, J.C. Functional traits help to explain half-century long shifts in pollinator distributions. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.J.; Purvis, A.; Gittleman, J.L. Quaternary climate change and the geographic ranges of mammals. Am. Nat. 2009, 174, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cao, B.; Bai, C.; Li, G.; Mao, M. Predicting suitable cultivation regions of medicinal plants with Maxent modeling and fuzzy logics: A case study of Scutellaria baicalensis in China. Environ. Earth Sci. 2016, 75, 1–12. [Google Scholar] [CrossRef]
- Pearce, J.; Lindenmayer, D. Bioclimatic analysis to enhance reintroduction biology of the endangered Helmeted Honeyeater (Lichenostomus melanops cassidix) in Southeastern Australia. Restor. Ecol. 1998, 6, 238–243. [Google Scholar] [CrossRef]
- Aguilar-Soto, V.; Melgoza-Castillo, A.; Villarreal-Guerrero, F.; Wehenkel, C.; Pinedo-Alvares, C. Modeling the potential distribution of Picea chihuahuana Martínez, an endangered species at the Sierra Madre Occidental, Mexico. Forests 2015, 6, 692–707. [Google Scholar] [CrossRef]
- Nabout, J.C.; Magalhães, M.R.; Ma, D.A.G.; Da, C.H. The impact of global climate change on the geographic distribution and sustainable harvest of Hancornia speciosa Gomes (Apocynaceae) in Brazil. Environ. Manag. 2016, 57, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Noulèkoun, F.; Chude, S.; Zenebe, A.; Birhane, E. Climate change impacts on Faidherbia albida (Delile) A. Chev. distribution in dry lands of Ethiopia. Afr. J. Ecol. 2017, 55, 233–243. [Google Scholar]
- Narouei-Khandan, H.A. Ensemble Models to Assess the Risk of Exotic Plant Pathogens in a Changing Climate. Ph.D. Thesis, Lincoln University: Lincoln, New Zealand, 2014. [Google Scholar]
- Huang, J.; Li, G.; Li, J.; Zhang, X.; Yan, M.; Du, S. Projecting the Range Shifts in Climatically Suitable Habitat for Chinese Sea Buckthorn under Climate Change Scenarios. Forests 2017, 9, 9. [Google Scholar] [CrossRef]
- Ray, D.; Behera, M.D.; Jacob, J. Indian Brahmaputra valley offers significant potential for cultivation of rubber trees under changed climate. Curr. Sci. 2014, 107, 461–469. [Google Scholar]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (H. riparia Lour) in Yunnan, China. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhao, Z.; Wei, H.; Gao, B.; Wei, G. Prediction of the potential geographic distribution of the ectomycorrhizal mushroom Tricholoma matsutake under multiple climate change scenarios. Sci. Rep. 2017, 7, 46221. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Luo, C.; Zhu, Y.; Wu, W.; Zhang, Q.; Du, G. Planting regionalization and ecological adaptability of walnut in Sichuan province. J. West China For. Sci. 2012, 3, 2, (in Chinese with English abstract). [Google Scholar]
- Xi, S. Gene resources of Juglans and genetic improvement of Juglans regia in China. Sci. Silvae Sin. 1987, 23, 342–350, (in Chinese with English abstract). [Google Scholar]
- Arellano-Valle, R.B.; Contreras-Reyes, J.E.; Genton, M.G. Shannon entropy and mutual information for multivariate skew-elliptical distributions. Scand. J. Stat. 2013, 40, 42–62. [Google Scholar] [CrossRef]
- Tuck, G.; Glendining, M.J.; Smith, P.; House, J.I.; Wattenbach, M. The potential distribution of bioenergy crops in Europe under present and future climate. Biomass Bioenergy 2006, 30, 183–197. [Google Scholar] [CrossRef]
- Evans, J.M.; Fletcher, R.J.; Alavalapati, J. Using species distribution models to identify suitable areas for biofuel feedstock production. GCB Bioenergy 2010, 2, 63–78. [Google Scholar] [CrossRef]
- Trabucco, A.; Achten, W.M.J.; Bowe, C.; Aerts, R.; Orshoven, J.; Van Norgrove, L.; Muys, B. Global mapping of Jatropha curcas yield based on response of fitness to present and future climate. GCB Bioenergy 2010, 2, 139–151. [Google Scholar] [CrossRef]
| Category | Variable | Description | Unit | Contribution (%) |
|---|---|---|---|---|
| Climate | Bio1 | Annual Mean Temperature | °C | |
| Bio2 | Mean Diurnal Range (Mean of monthly (max temp − min temp)) | °C | ||
| Bio4 | Temperature Seasonality (standard deviation × 100) | °C | ||
| Bio5 | Max Temperature of Warmest Month | °C | 2.0 | |
| Bio6 | Min Temperature of Coldest Month | °C | 63.9 | |
| Bio7 | Temperature Annual Range (Bio5 − Bio6) | °C | 7.1 | |
| Bio8 | Mean Temperature of Wettest Quarter | °C | ||
| Bio9 | Mean Temperature of Driest Quarter | °C | ||
| Bio10 | Mean Temperature of Warmest Quarter | °C | ||
| Bio11 | Mean Temperature of Coldest Quarter | °C | ||
| Bio12 | Annual Precipitation | mm | 12.6 | |
| Bio13 | Precipitation of Wettest Month | mm | ||
| Bio16 | Precipitation of Wettest Quarter | mm | ||
| Bio17 | Precipitation of Driest Quarter | mm | ||
| Bio18 | Precipitation of Warmest Quarter | mm | ||
| Bio19 | Precipitation of Coldest Quarter | mm | ||
| Topography | Elevation | m | 5.1 | |
| Slope | ° | 2.8 | ||
| Aspect | rad | 0.8 | ||
| Soil | t_bulk_den | Topsoil Bulk Density | kg/dm3 | 0.9 |
| s_bulk_den | Subsoil Bulk Density | kg/dm3 | 0.5 | |
| t_clay | Topsoil Clay Fraction | % | 1.7 | |
| s_clay | Subsoil Clay Fraction | % | ||
| t_gravel | Topsoil Gravel Content | % | 0.3 | |
| s_gravel | Subsoil Gravel Content | % | 0.0 | |
| t_ph_h20 | Topsoil pH (H2O) | −log(H+) | 0.3 | |
| s_ph_h20 | Subsoil pH (H2O) | −log(H+) | ||
| t_esp | Topsoil Sodicity (ESP, exchangeable sodium percentage) | % | 0.4 | |
| s_esp | Subsoil Sodicity (ESP, exchangeable sodium percentage) | % | 0.2 | |
| t_sand | Topsoil Sand Fraction | % | 0.2 | |
| s_sand | Subsoil Sand Fraction | % | 0.1 | |
| t_silt | Topsoil Silt Fraction | % | 0.5 | |
| s_silt | Subsoil Silt Fraction | % | 0.6 |
| Future Climate Change Scenario | Unchanged | Loss | Gain | Total change (= Gain − Loss) 1 | |
|---|---|---|---|---|---|
| Period of 2041–2060 | RCP 2.6 | 96.7 | 3.3 | 16.8 | 13.4 |
| RCP 4.5 | 92.3 | 7.7 | 25.7 | 18.0 | |
| RCP 6.0 | 93.3 | 6.7 | 20.5 | 13.7 | |
| RCP 8.5 | 95.3 | 4.7 | 40.6 | 35.8 | |
| Period of 2061–2080 | RCP 2.6 | 90.6 | 9.4 | 19.8 | 10.4 |
| RCP 4.5 | 91.6 | 8.4 | 19.2 | 10.7 | |
| RCP 6.0 | 92.5 | 7.5 | 43.7 | 36.2 | |
| RCP 8.5 | 91.1 | 8.9 | 82.5 | 73.6 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Zhang, H.; Yue, J.; Xie, T.; Xu, Y.; Tian, Y. Predicting Shifts in the Suitable Climatic Distribution of Walnut (Juglans regia L.) in China: Maximum Entropy Model Paves the Way to Forest Management. Forests 2018, 9, 103. https://doi.org/10.3390/f9030103
Xu X, Zhang H, Yue J, Xie T, Xu Y, Tian Y. Predicting Shifts in the Suitable Climatic Distribution of Walnut (Juglans regia L.) in China: Maximum Entropy Model Paves the Way to Forest Management. Forests. 2018; 9(3):103. https://doi.org/10.3390/f9030103
Chicago/Turabian StyleXu, Xiang, Huayong Zhang, Junjie Yue, Ting Xie, Yao Xu, and Yonglan Tian. 2018. "Predicting Shifts in the Suitable Climatic Distribution of Walnut (Juglans regia L.) in China: Maximum Entropy Model Paves the Way to Forest Management" Forests 9, no. 3: 103. https://doi.org/10.3390/f9030103