Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest



1
Swiss Federal Institute for Forest, Snow and Landscape Research WSL, Swiss Forest Protection, Zürcherstrasse 111, 8903 Birmensdorf, Switzerland
2
ETH Zurich, Institute of Integrative Biology, Forest Pathology and Dendrology, Universitätstrasse 16, 8092 Zurich, Switzerland
3
ETH Zurich, Institute of Terrestrial Ecosystems, Forest Ecology, Universitätstrasse 16, 8092 Zurich, Switzerland
*
Author to whom correspondence should be addressed.
Forests 2018, 9(3), 120; https://doi.org/10.3390/f9030120
Submission received: 8 February 2018
/
Revised: 26 February 2018
/
Accepted: 2 March 2018
/
Published: 5 March 2018
(This article belongs to the Special Issue The Forest Renewal Issue: New Problems and Findings in the Establishment of Tree Regeneration)
Abstract
:Regeneration of Picea abies in high-elevation mountain forests often depends on the presence of coarse woody debris (CWD), as logs provide sites with more favorable conditions for spruce regeneration compared to the forest floor. However, there is little quantitative knowledge on the factors that are conducive to or hindering spruce establishment on CWD. We examined spruce regeneration on CWD by sampling 303 plots (50 cm × 50 cm each) on 56 downed logs in a virgin forest in the Swiss Alps. Variables describing microsite conditions were measured, and fungi were isolated from wood samples. To investigate the relationship between the ecological factors and establishment success, two models were fitted with seedling and sapling density as response variables, respectively. Besides log diameter, the models identified different ecological factors as significant for seedling and sapling establishment, i.e., regeneration depends on different factors in different development stages. Seedling density depended on the type of rot, log inclination, and decay stage. Sapling density depended mainly on light availability, cover by bark and moss, the time of tree fall, and the distance between the log surface and the forest floor. A total of 22 polypore fungi were isolated from the wood samples, four of them being threatened species. White- and brown-rot fungi were found in all decay stages. The visual assessment of the type of rot in the field corresponded in only 15% of cases to the type of rot caused by the isolated fungi; hence caution is needed when making field assessments of rot types.
1. Introduction
The successful establishment of sufficient amounts of tree regeneration is an important element of forest dynamics. Many European high-elevation Norway spruce forests (“subalpine” forests, usually at elevations >1600 m a.s.l.) harbor only little regeneration today [1], due to (1) forest overexploitation in the past followed by the re-growth of even-aged, dense, dark forest stands; (2) misconceptions of forest ‘cleanliness’ that have led to the elimination of nearly all coarse woody debris (CWD); and (3) harsh abiotic conditions for trees at the subalpine level [2,3]. Insufficient tree regeneration can be harmful in forests that protect people and infrastructure against natural hazards [4].
Too little or too much solar radiation, dry air, heat deficits, long-lasting and high snow cover, winter desiccation, competition by dense herbaceous vegetation, and ungulate browsing render the regeneration of Norway spruce quite difficult at high elevations [3,5]. Even small differences in aspect, inclination, or landform can have a considerable impact on these factors, thus limiting regeneration to a few suitable microsites [6]. Moreover, ‘mast years’ with ample seed production occur approximately every five years only, and even then successful regeneration is not guaranteed [3]. As a response to these harsh conditions, trees at high elevations have evolved strategies for successful regeneration, one of them being the ability to regenerate on CWD [7,8]. CWD has several characteristics that render it advantageous for spruce regeneration, including protection against snow movements, a reduced risk of damage by snow molds due to the shorter duration of snow cover, the reduction of competition with the herbaceous layer, and reduced accessibility for some ungulate species [3,5,9].
Many studies in boreal and temperate forests have confirmed the important role of CWD in tree population dynamics, e.g., for high-elevation forests in Switzerland [10], the Carpathian Mountains in Poland [11,12], the Czech Republic [7], Scandinavia [13], the Colorado Rocky Mountains [14], Canadian forests [15], and the Japanese Taisetu Mountains [8,16]. Most studies have compared regeneration on CWD with regeneration on other microsites (e.g., mineral soil), evaluating predictor variables such as log diameter [8] and length [17], area of log surface [18], or log age/time after tree death and/or decay stage [11,17,19] to evaluate the substrate preference of tree regeneration. Others have evaluated the importance of environmental factors like canopy openness [20] and altitude [21], but only few attempts have been made to assess both log characteristics and the environmental conditions that are decisive for regeneration on the nurse logs themselves.
Evidently, CWD serve as “safe sites” for tree regeneration under specific circumstances only. Kathke and Bruelheide [22] found the highest sapling densities on logs, compared to other microsites such as moss-covered rocks or the forest floor. Bače et al. [23] demonstrated that the degree of ground contact, presence of white-rot-causing fungi (e.g., Armillaria spp. and Phellinus nigrolimitatus), and log diameter were positively related to both seedling and sapling density, and sapling densities were higher on windthrown trees than on trees killed by bark beetles. Conversely, the presence of brown-rot-causing Fomitopsis pinicola and tree death after bark beetle attack were negatively related to regeneration density. Seedling density peaked on medium-decayed logs, whereas sapling density increased monotonically as decay progressed. Thus, decay fungi appear to be of great importance for the establishment of regeneration on CWD.
With our study, we aimed to investigate the interaction and impacts of a wide range of ecological and environmental factors on seedling and sapling density in one of the few virgin forests of Switzerland, the forest reserve Scatlè. Our objectives were (a) to identify the ecological factors that influence the establishment of recently germinated seedlings and saplings on decaying Norway spruce logs, (b) to test how these factors influence the abundance of regeneration, and (c) to investigate the diversity of polypores responsible for the decay of CWD.
2. Materials and Methods
2.1. Study Site
The forest reserve Scatlè is located in the municipality Breil/Brigels (Grisons, Switzerland; 46°47′23.8″ N, 9°02′50.5″ E), at 1580–2015 m a.s.l. on the eastern slope of Piz Dado. The forest reserve is facing ENE, with a high inclination of 70–100%. The core area of 9.13 ha (total area: 24 ha) was placed under strict protection in 1910. Pollen analyses revealed no human use of the forest since at least the 13th century, most likely attributable to its difficult accessibility [24]. A postglacial rockfall of scree and huge boulders has created a system of subterranean crevices through which cold airflow leads to locally very low temperatures, creating a unique microclimate. Therefore, forest texture and structure are rather heterogeneous. The dominant tree species in Scatlè is Norway spruce (Picea abies (L.) Karst.), which is susceptible to windthrow and bark beetle attacks that are interacting with hazards such as avalanches and snow break.
In the reserve, systematic inventories took place in 1965, 1977, 1989, and 2006, where each individual stem with a DBH > 4 cm was individually tagged (numbered) and measured in the core area, such that the fate of each tree can be tracked over the past 50 years. Notably, it was differentiated whether a tree was living; standing dead; or downed.
2.2. Experimental Design and Measured Variables
Regeneration on CWD was examined on 56 Norway spruce logs, selected to represent different decay stages, ranging from newly fallen to extremely strongly decayed. On the oftentimes-heterogeneous logs, observational units were defined as plots of 50 cm × 50 cm at an interval of 3 m along the stem. For plots on logs with a diameter <50 cm, the section within the plot boundaries was investigated and scaled to an area of 2500 cm2.
On each of a total of 303 plots, the numbers of seedlings (defined here as recently germinated plants with the characteristic four to nine cotyledons) and saplings (plants ≥ 10 cm in height) were counted to determine their density (number of plants per m2), and the following variables were measured: height of the tallest individual; log diameter at the plot center; distance between top of the log and forest floor (average of the vertical distances between forest floor and top of plot on each side of the log); decay stage of the log according to Renvall [28]; fractional cover of bark and mosses; mean height of mosses (mean of five measurements: four corners and plot center); fractional cover by herbaceous vegetation (>100% possible if several layers); substrate underneath the log at the plot center (categorical: log without ground contact; CWD; rock; soil); percentage of ground contact of the entire log; plot inclination (measured with an inclinometer); visual assessment of the presence/absence (binary) of white, brown, or both types of rot within the plot boundaries along with the presence of polypore fruiting bodies (visual assessment and classification). Increment cores and wood chips were extracted for the isolation of basidiomycetes in the lab. Fisheye photographs were taken to determine the light regime for each plot. All data were collected in September 2012. Lastly, the time of tree fall was approximated based on tree status in the regular inventories (i.e., before 1965, between 1965–1977, 1977–1989, 1989–2006, and after 2006).
2.3. Data Analysis
All statistical analyses refer to the 50 cm × 50 cm plots as the basic unit. Significance classes for the statistical analyses were based on Stahel [29]. Two models were fitted for determining the importance of different ecological factors (independent variables) for the establishment of Norway spruce regeneration, with seedling and sapling density as the response variables. Both response variables were square root transformed [29]. Since the plots within one piece of CWD are not statistically independent, generalized linear mixed models (GLMM) fitted by the Laplace approximation were used with the assessed variables as fixed effects and the tree individual as a random effect term for the intercept [30]. The independent variables were tested for correlation; for variables with a Pearson correlation >0.5, only one was chosen for model fitting [31]. Model selection was made using the corrected Akaike Information Criterion (AICc) in a backward elimination procedure. The Pearson correlation of observed vs. fitted regeneration density was calculated for the models as an indicator of model performance, and model robustness was tested with a 10-fold cross-validation (100 repetitions; [29,32]).
For examining the different fungal species in their relation to the decay stage, non-metric multidimensional scaling (NMDS) using the Bray-Curtis dissimilarity was performed using the community ecology package vegan (V 2.0–5) in R (R for Windows v2.14.0, R Foundation for statistical computing, Vienna, Austria).
Statistical analyses were conducted using the software R (R for Windows v2.14.0, R Foundation for statistical computing, Vienna, Austria). Model fitting was performed using the package lme4 (Version 0.999999-0) for linear mixed-effect models. Graphs were plotted using the package ggplot2 (Version 0.9.1).
The fisheye photographs were analyzed using the Software SideLook (Version 1.1.01) and Gap Light Analyzer (Version 2.0) [33] to determine sunshine duration in June (min/day), canopy openness, and the percentage of the total amount of solar radiation (direct and diffuse, taking the topographic mask into account).
2.4. Detection and Identification of Wood-Decomposing Basidiomycetes
For the isolation of wood-decomposing basidiomycetes, the increment cores and wood chips were placed on a selective malt agar medium (20 g/L malt extract, 16 g/L agar, 230 mg/L thiabendazole; [34]). After incubation, fungal mycelium was collected for molecular identification of the species by sequencing the internal transcribed spacer (ITS) region of the nuclear rDNA [35]. DNA sequences were obtained as described by Queloz et al. [36] and compared to sequences deposited in GenBank/NCBI using BLAST [37].
3. Results
The values of the continuous variables varied strongly among plots (Table 1). In terms of performance and robustness, the sapling model was found to be superior for prediction compared to the seedling model (Table 2). The sapling model also had a smaller standard deviation (170.9) than the seedling model (275.1), indicating that seedling density is much more variable than sapling density (Table 2).
3.1. Seedling Density
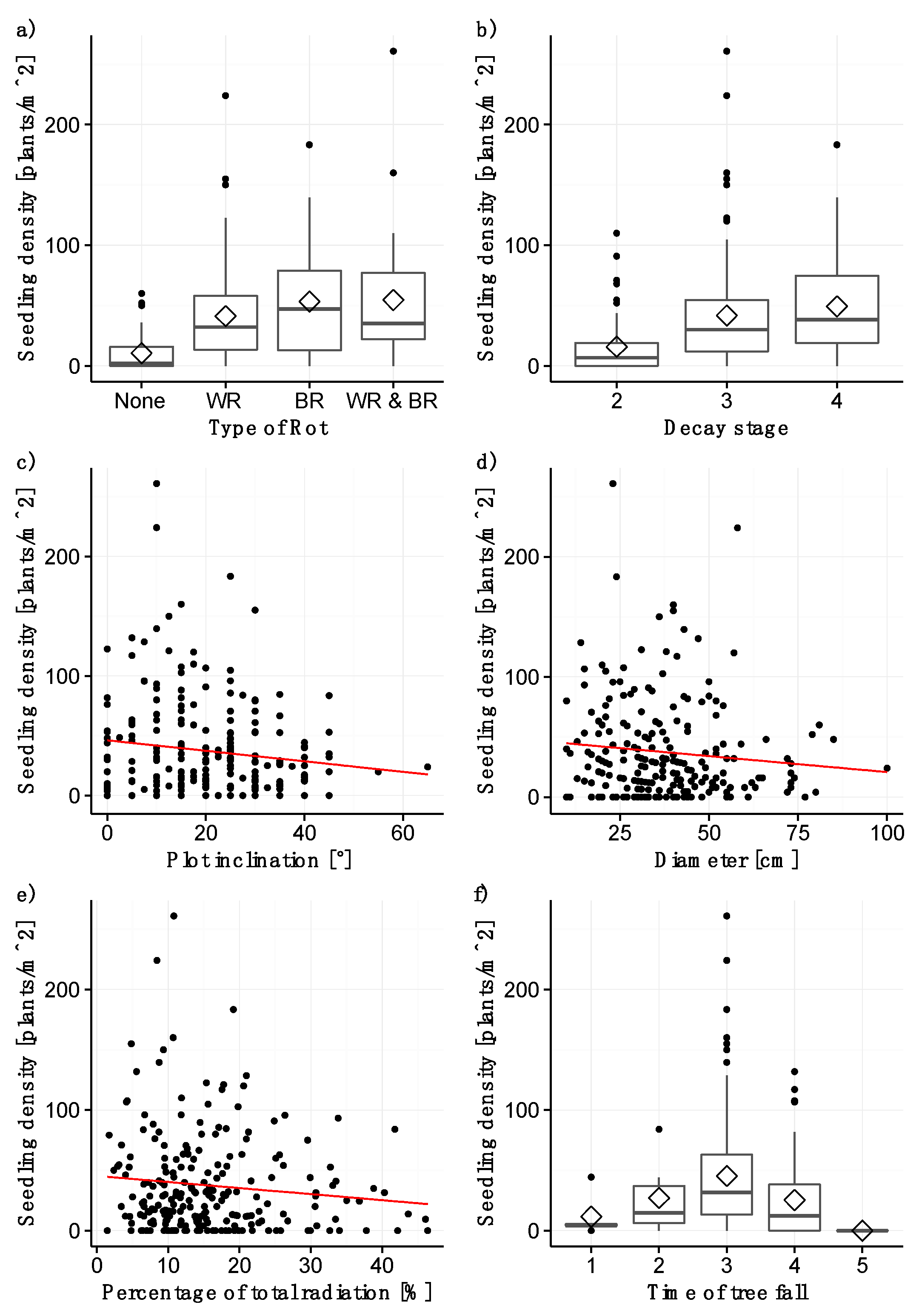
Log diameter and the presence of wood rot were found to be highly significant factors with positive estimates (p < 0.001; Table 2; Figure 1a,d). Most seedlings occur on plots with log diameters between 15 and 70 cm. The model indicates a significant positive slope of the regression line for the diameter, whereas the position of the regression line is uncertain in the diagram shown in Figure 1d. The reason for this situation is that “log” was entered as a random variable in the model, and, thus, the plots on the same log were considered to be dependent, whereas in the diagram, they were treated as being independent. Plots without detectable rot had significantly less regeneration than plots showing signs of fungal decay (pairwise t-test, p < 0.01). However, no differences in plot regeneration densities were detectable between white rot, brown rot, and plots with both types of rot (pairwise t-test, p > 0.05).
Plot inclination influenced seedling density very strongly (p < 0.001; Table 2; Figure 1c), suggesting that high inclination hinders regeneration. Decay stages 3 and 4 (positive estimates) were also important for seedling establishment (p < 0.05 and p < 0.01, respectively; Table 2; Figure 1b). Plots in decay stages 3 and 4 harbor significantly more seedlings than plots of decay stage 2 (pairwise t-test, p < 0.001).
The model with the lowest AICc included the time of tree fall and the percentage of total radiation, even though these variables were not significant predictors of seedling density on the plots (Figure 1e,f).
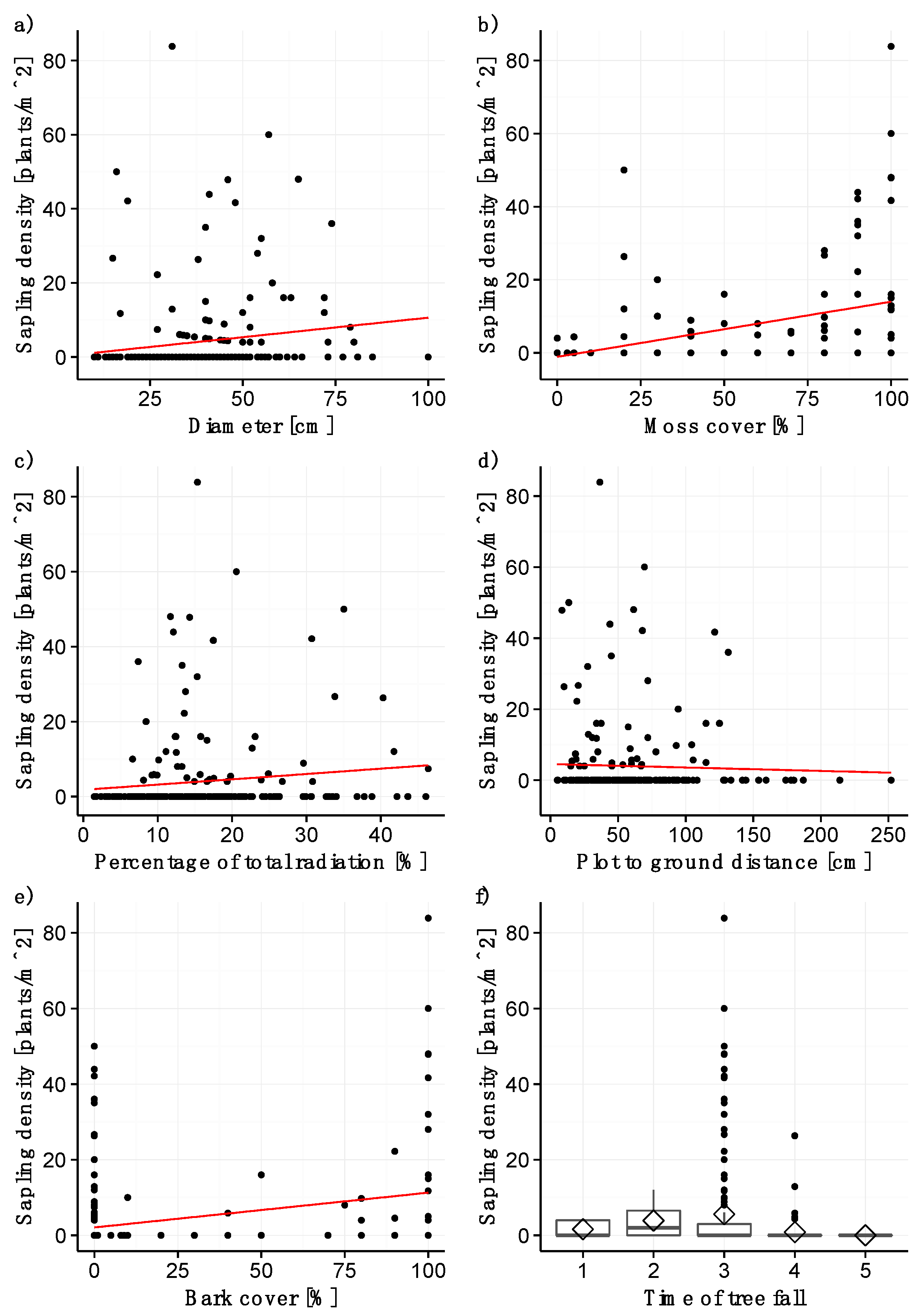
3.2. Sapling Density
Similar to the seedling model, log diameter was highly significant for modeling sapling density (p < 0.001; Table 2; Figure 2a). A log diameter of at least 15 cm is required for the establishment of saplings, and maximum density was observed for log diameters between 30 and 60 cm. Likewise, moss cover (Figure 2b) and the percentage of total radiation (Figure 2c) have a strongly significant (p < 0.001, Table 2) and positive effect on sapling establishment. Plots with a moss cover of 80–100% harbor the greatest sapling density. The distance from the plot surface to the forest floor (Figure 2d) and the time of tree fall (p > 0.01; Table 2; Figure 2f) turned out to be strongly significant, with stems that had fallen between 1977 and 1989 being colonized by saplings most densely. Sapling density was significantly higher on stems that had fallen between 1977 and 1989 than on stems that fell between 1989 and 2006 (pairwise t-test, p < 0.05). Sapling density decreased with increasing distance of the plot from the forest floor, with plots further than 140 cm from the forest floor harboring no saplings. Bark cover was included in the model as a weakly significant variable with a positive estimate (p < 0.05; Table 2), even though sapling density was high on some completely decorticated stems (Figure 2e). No other statistically significant differences were detectable.
3.3. Polypore Diversity and Decay Stage
In total, 37 polypore fruiting bodies representing seven species were macroscopically identified on the 274 plots assessed for fungal decay. The most common fungi present as sporocarps were the red-banded polypore (Fomitopsis pinicola) and the pink hoof-polypore (Fomitopsis rosea), which were found 14 and eight times, respectively. Amylostereum aerolatum and Trichaptum abietinum were only found as sporocarps (Table 3). By visual assessment, 144 (52.5%) plots showed typical white rot symptoms, but only 30 (11%) plots solely those of a typical brown rot. Another 37 (13.5%) plots showed both white and brown rot, whereas 63 plots (23%) did not show any sign of white or brown rot.
From a total of 274 plots, wood was sampled to isolate wood decaying basidiomycetes in the laboratory. Wood samples and increment cores of 80 plots (29%) resulted in a successful isolation of fungal mycelia in the lab. On these 80 plots, 99 basidiomycete cultures representing 21 species were identified by ITS sequencing (Table 3). Again, Fomitopsis pinicola was found to be the most abundant species (33 isolates), followed by Antrodia serialis (16 isolates) and Fomitopsis rosea (11 isolates)—all of them being brown-rot fungi [38]. Fifty-six of the 80 plots hosted brown rot fungi, 13 white rot fungi, three both brown and white rot fungi, and eight plots hosted fungi not known to be involved in the decomposition of wood. The visual assessment of the type of rot in the field corresponded to only 15% with the type of rot caused by the fungi that were isolated in the lab.
Four of the fungal species found in Scatlè are threatened according to IUCN criteria and are listed as threatened macrofungi of Switzerland (‘Red List’): Amylostereum aerolatum, Fomitopsis rosea, Phellopilus (Phellinus) nigrolimitatus, and Veluticeps abietina.
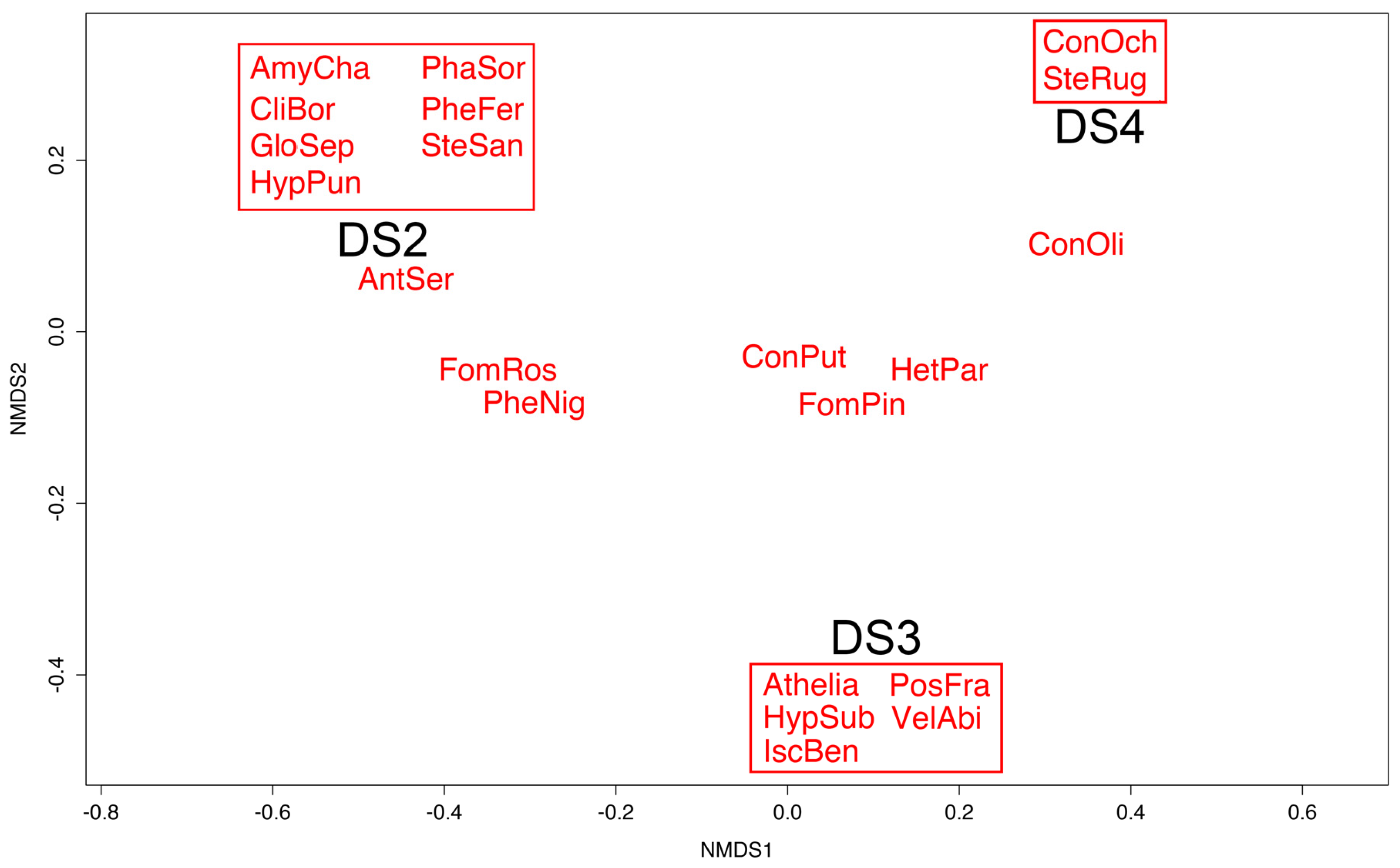
Non-metric multidimensional scaling showed that Amylostereum chailletii, Climacocystis borealis, Gloeophyllum sepiarium, Hypochnicium punctulatum, Phanerochaete sordida, Phellinus ferrugineovelutinus, and Stereum sanguinolentum were only found on plots exhibiting decay stage 2. Similarly, Athelia sp., Hypochnicium subrigescens, Ischnoderma benzoinum, Postia fragilis, and Veluticeps abietina were solely associated with plots of decay stage 3, whereas Coniophora ochraceum and Stereum rugosum were exclusively found on decay stage 4 plots. Fomitopsis pinicola occurred almost equally often on decay stage 2 and 3 logs. The other fungi were not associated with a particular decay stage (Figure 3).
4. Discussion
While the density of Norway spruce seedlings and saplings on nurse logs in Scatlè depends on log diameter, incident radiation, and the time of tree fall, other ecological factors differ in their relevance for the two life stages, as discussed.
4.1. Factors with a Significant Effect on Both Seedling and Sapling Density
Total α-diversity of wood colonizing vascular plants, lichen, and bryophytes has been found to increase with increasing diameter of the decaying log [39]. Log diameter was highly significant, with positive coefficients indicating that the larger the log, the higher the regeneration density. This is consistent with observations by Takahashi [8] and Bače et al. [23]. In our study, the highest regeneration densities occurred on logs with a diameter between 20 and 70 cm. Similarly, Iijima et al. [40] observed low numbers of seedlings and saplings on logs <20 cm. Zielonka and Piątek [41] suggested that the larger surface of the thicker logs enhances seed germination probability. Also, smaller logs tend to dry out faster [9]. Eichrodt [2] concluded that the diameter has to be >25 cm for the successful establishment of spruce regeneration, and our results confirm this.
The fraction of solar radiation reaching the plot had a negative effect on seedling establishment, which may be counter-intuitive. However, high-altitude air contains little water vapor and dust, thus reducing the reflection and scattering of radiation, leading to high direct radiation. Since solar radiation is coupled with temperature, objects warm up fast even if the air temperature is relatively low [3], and since seedlings have a small rooting system, they are particularly prone to drought [5]. Christy and Mack [42] found the death of <5-year-old regeneration of Tsuga heterophylla to be common during the summer months due to desiccation. Norway spruce is a moderately shade-tolerant species, and seedlings can survive light intensities of less than 5% of full sunlight for one to two years [43,44]; hence shaded conditions are more conducive for seedlings than high sunlight.
However, light availability was strongly significant and positive for saplings. Insufficient light availability is one of the main factors limiting the growth of taller Norway spruce regeneration [11,45]. According to Kimmins [43] (p.182), between 15–24% of direct sunlight is necessary if Norway spruce regeneration is to develop well at 10–15 years of age. Most saplings in Scatlè were found on plots with 10–30% of full sunlight. Other studies frequently use the average sunshine duration in June to account for the presence/absence of regeneration [46]. Ott et al. [3] suggested a minimum of 120–180 min of daily direct sunlight in June for the good growth of spruce regeneration in the subalpine zone of the Swiss Alps, whereas Wasser and Frehner [47] suggested 60–120 min/day. The latter is consistent with our measurements in Scatlè, where most saplings experience between 50–120 min/day in June. Thus, low light intensities appear to be less limiting for seedlings than desiccation caused by high incident radiation. In contrast to seedlings, the root system of saplings is better developed, thus reducing the danger of desiccation and making higher light intensities advantageous for thriving.
The time of tree fall (i.e., between 1977 and 1989) was found to be highly significant for sapling establishment, in contrast to the other periods. It was also used in the seedling model, but solely for improving model fit. Thus, logs seem to be most suitable for sapling establishment 23–47 years after tree fall, i.e., 25+ years after tree death. This finding is consistent with observations by Zielonka [11], who performed cross-dating of fallen conifer logs in order to account for the time of tree fall and discovered that decaying logs become most suitable for regeneration 30–60 years after tree death. According to Zielonka and Piątek [41], even about 50 years have to elapse before a tree becomes suitable for harboring vascular plants and Norway spruce regeneration.
Due to the heterogeneity of microsites in Scatlè, only a weak and insignificant correlation between the decay stage of the stems and the time of tree fall was detectable (r = −0.282), as decomposition is a highly complex process [48]. Thus, it is impossible to predict the decay state of a log in the absence of detailed data on its microsite (shading, distance to the forest floor, etc.).
4.2. Factors with a Significant Effect on Seedling Density
The importance of the type of rot was also studied by Fukasawa [49] in Japan, where the seedling density of Clethra barbinervis was significantly higher on logs that had been classified visually as ‘brown-rotted’, although the nutrient content of brown- vs. white-rotted wood is not thought to differ significantly [48]. However, no preference of seedlings for wood that was decayed by either white or brown rot was detectable in our study. In contrast, seedling density was higher on white-rotted wood than on brown-rotted wood in the study by Bače et al. [23]. Hence the current state of knowledge does not indicate a clear pattern.
It has been suggested that decaying wood harbors higher amounts of active mycorrhizal fungi during dry summer months than soil [50]. However, none of the basidiomycetes that we isolated are known to be involved in the formation of mycorrhiza in Norway spruce [51]. Nonetheless, wood decaying fungi are closely linked to the presence of mycorrhizal fungi, which in turn are important for seedling establishment on CWD [50].
Plot inclination had a highly significant and negative influence on seedlings, but not on saplings. This is likely attributable to seed retention, i.e., higher chances of seeds being trapped on surfaces of lower inclination. Seeds falling on strongly inclined logs are likely to slip off the surface, or they may later be washed off by rain. Moreover, the slower runoff of water on horizontal or less inclined logs leads to a higher infiltration of moisture into the logs, with positive effects on seed germination, seedling survival, and establishment. In addition, surface texture has been shown to influence seed retention, i.e., crevices in the log surface are highly important structures [11].
We found recently germinated seedlings on plots of all decay stages, starting with stage 2 since no stems of decay stage 1 were present at the time of sampling. Červenka et al. [52] found a tendency of regeneration with heights of 0–5 cm to establish on little decayed and less occupied logs over a three-year observation period. Takahashi et al. [53], distinguishing between five decay stages, also reported the presence of Picea jezoensis and Picea glehnii regeneration on logs of decay stage 2. These microsites appear to provide all requirements for germination and early establishment. Similarly, seedling density was highest on logs of medium-decayed Norway spruce logs in the Bohemian Forest [23] and in the Polish Karkonosze Mountains [18]. In our study, seedlings were found preferentially on plots of decay stages 3 and 4, but both stages were found to be equally suitable for seedlings. The same relationship was observed by Narukawa et al. [17] in the mountains of Japan and by Szewczyk and Szwagrzyk [12] and Zielonka and Piątek [19] in Poland. However, these studies revealed a continuous increase of regeneration density with increasing wood decay. This can be explained by the fact that as CWD decays, its density decreases, and as soon as the density has decreased to about 50%, water-holding capacity and nutrient content increase, making the wood more and more suitable for seedling establishment [9,54,55]. Moss cover, however, was not a significant factor in the seedling model. This is consistent with Iijima et al. [40] who found that the presence of moss does not affect the survival of one-year old regeneration of Picea jezoensis on fallen logs. Nevertheless, the colonization of downed logs also depends on the microsite, as other studies found intermediate decay stages to be most suitable for seedling colonization [18].
4.3. Factors with a Significant Effect on Sapling Density
We found that the presence of saplings strongly depends on the presence of moss, congruent with observations by Sugita and Nagaike [56] in Japan. Chmura et al. [57] found that the presence of a bryophyte carpet made the colonization of vascular plants more probable on Norway spruce logs than on beech logs. Nakamura [58] also suggested a positive effect of bryophytes on the survival of conifer regeneration. This relationship is probably due to the ability of mosses to retain water and therefore to provide a moist environment, which is particularly important for the drought-prone Norway spruce [17,44]. Typically, a nutrient-rich humus layer consisting of dead moss and decayed wood develops underneath the moss, increasing in depth as decomposition progresses [58,59]. In Canada, Simard et al. [15] found taller saplings (>10 cm) to be associated mostly with moss because of higher moisture availability and lower competition with the herbaceous layer, thus also agreeing with our observations.
Bark cover was found to be weakly significant and positive for sapling density. Two observations made in Scatlè explain this positive relationship. Firstly, moss has been found to grow preferably on bark, and it has the ability to rapidly colonize bark [46]. In Scatlè, we observed that a thick moss cover was often found on very soft and partly decayed bark layers. Nevertheless, bark and moss covers were not strongly correlated (r = 0.33). According to Holeksa et al. [60], broken and uprooted lying stems retain their bark for a longer time than snags, even when the wood is becoming softer, thus allowing for the penetration by the roots of small trees. Bark is more nutritious than wood, and thus the combination of (1) moisture-retaining moss; (2) soft, partly decayed, relatively nutrient-rich bark [53] underneath the moss cover; and (3) partly decayed wood in which tree roots can grow to stabilize the young plant most likely has a positive influence on sapling density in Scatlè. Secondly, we observed relatively well-developed regeneration in cracks of the bark of recently fallen stems, which—in combination with litter accumulations in the cracks—can provide sites for tree germination [9]. However, the longevity of these individuals is questionable, since the bark often falls off before the roots are able to penetrate the bark and the wood for establishment. This is probably one of the reasons for the bimodal distribution of sapling density relative to the percentage of the stem covered by bark (Figure 2e). The completely decorticated stems were mostly in advanced stages of decay, which are conducive to tree establishment, even though the decay-density relationship was not significant for our saplings.
CWD is an elevated microsite that reduces competition with the herbaceous layer and is not conducive to litter accumulation, which may also hinder the establishment of regeneration [3,42]. Our data suggest that sapling density is highest on plots that are 20–70 cm above ground, regardless of whether the logs showed any ground contact or not. Botting and DeLong [61] suggested that suspended logs have a lower moisture content in summer and therefore are less suitable regeneration sites. Our results are consistent with observations by Zielonka and Piątek [41], according to whom the plant cover of logs close to the ground does not hinder spruce regeneration.
In our study, sapling density depended on neither the decay stage nor the type of rot. When modeling seedling (<15 cm) and sapling (≥15 cm) densities, Bače et al. [23] reached very different conclusions: seedling density was significantly influenced by log diameter, ground contact, surrounding vegetation and dominant rot fungi; and sapling density by log diameter, decay stage, ground contact, and dominant rot fungi. We also found log diameter to be a significant factor for both seedling and sapling density. However, in our study, ground contact and surrounding vegetation were eliminated during the model fit, and decay stage was significant for seedling density only. These differences could be an indicator for strong variability in the importance of the different log characteristics due to environmental factors, e.g., higher precipitation combined with a lower mean annual temperature in Scatlè.
4.4. Polypore Diversity and Decay Stage
The investigation of the sporocarps in the field yielded seven species only. All visually assessed fungi except Amylostereum aerolatum and Trichaptum abietinum were also successfully isolated from the boring cores. According to Allmér et al. [62], solely investigating sporocarp diversity will not reveal the full species richness, since some fungi can be present only as mycelia, as the formation of sporocarps depends strongly on current environmental conditions. In Scatlè, the molecular identification of fungal isolates yielded 21 species, i.e., three times as many as the field assessment. Similarly, Rajala et al. [63] detected 2.6 times more fungal species by PCR amplification and sequencing of extracted DNA from wood compared to field observations of sporocarps. Therefore, the isolation of wood-decaying basidiomycetes led to a more complete picture of fungal species richness in Scatlè.
The type of rot diagnosed by visual assessment corresponded in only 15% of the cases to the type of rot that is caused by the polypore isolated from wood from those plots. This indicates that a visual assessment is not a reliable basis for diagnosing rot type. The large diversity of wood decay fungi, along with the abundant presence of the rare Fomitopsis rosea [38,64,65], is an additional indicator of the primeval character of the Scatlè forest. By comparison, Lindblad [66] and Penttilä et al. [67] found that species richness of wood decay fungi is significantly lower in managed forests.
Jönsson et al. [68] grouped sporocarps of 13 dominant wood decay fungi on Norway spruce wood into pioneer colonizers, early secondary colonizers, and late secondary colonizers. Eight of those 13 species were found in Scatlè as well. The overall tendency of their data is similar to those of our study, with one exception: Jönsson et al. [68] found 201 sporocarps of Phellopilus nigrolimitatus and classified them as belonging to the late secondary colonizers. In Scatlè, no fruiting body of Phellopilus nigrolimitatus was found, but the fungus was isolated from three plots that were in decay stage 2 or 3, which would better correspond to pioneer and early secondary colonizers.
The brown-rot fungus Fomitopsis pinicola was the most abundant fungus and found throughout all decay stages, and Stereum sanguinolentum was confined to less decayed logs, thus confirming the findings by Høiland and Bendiksen [69]. Finally, similar to Rajala et al. [63] we detected both white- and brown-rot fungi across all decay stages, but differences were observed in colonization frequencies of the different decay stages. In contrast to their study, however, we found brown-rot fungi in all decay stages rather than mostly in decay stage 2 and 3. The most common brown-rot fungi Antrodia serialis and Coniophora olivacea identified by Rajala et al. [63] were also isolated in Scatlè. Antrodia serialis was the second most abundant brown-rot agent in Scatlè and Coniophora olivacea was only found twice. Evidently, the successional patterns of wood decay fungi in Norway-spruce logs depend on many factors such as decay stage, micro- and macroclimatic conditions, or the vicinity of inoculum sources on previously colonized logs [63,68].
As logs decay, their physical and chemical properties change [9,70]. Water content, nitrogen, and phosphorous content increase, while wood density decreases with increasing decay stage [70]. The increase in nitrogen content is mainly attributed to the active transport of nutrients from the forest soil to the substrate by wood-decaying fungi [46,71,72], while a smaller role is ascribed to nitrogen fixation [54]. Further, the cellulose content of deadwood was shown to decrease as abundant brown-rot fungi preferably degrade cellulose [70,71,72].
Finally, the cause of tree death, e.g., bark beetle attack or windfall, is probably another factor influencing both the attractivity of logs and snags for primary wood decay fungi and the dynamics of wood-decaying fungal communities.
5. Conclusions
This study provides evidence that regeneration on CWD in the virgin forest Scatlè is a result of a complex interplay of many ecological factors that are differentially influencing the regeneration at different development stages. Recently germinated seedlings and saplings exhibited substantially different requirements with respect to suitable microsites on decaying logs. Recently germinated seedlings were found mainly on plots with an advanced decay and a moderate plot inclination, whereas saplings occurred mostly on slightly elevated logs that had fallen approximately 30 years ago; were covered by bark and moss; and experienced about 10–30% of full light availability.
Only log diameter was identified as a significant factor for both stages of regeneration. The higher standard deviation of model 1 (seedling density) compared to model 2 (sapling density) suggests a more heterogeneous distribution of the seedlings on the sampled plots. Model 2 (saplings) showed better performance and robustness. For drawing inferences on the long-term success of tree establishment on CWD, the factors included in model 2 should therefore be taken into account.
Finally, the molecular identification of mycelial isolates from decayed wood revealed three times more species than the macroscopic identification of fruiting bodies in the field. Also, the rot type as identified by visual assessment of the plots did not correspond well to the type of rot ascribed to the fungi isolated from these plots. The great species diversity and the presence of Fomitopsis rosea and four species classified as threatened according to IUCN criteria underline the primeval character of this forest reserve.
Acknowledgments
We wish to thank the Genetic Diversity Center at ETH Zurich (GDC) for providing the necessary laboratory facilities; Tea Wey for excellent technical assistance; and Corina Pescatore, Christoph Laule, Andrin Gross, as well as Christine and Manuel Vögtli for their help in the field. Furthermore, we would like to thank Valentin Queloz for helping with all molecular analyses, Christof Bigler for his support with the linear mixed models, and two anonymous reviewers for valuable input.
Author Contributions
H.B., M.W., and S.S. conceived and designed the experiments; S.S. performed the field work; S.S. and T.N.S. performed all laboratory analyses; S.S. analyzed the data, M.W. and T.N.S. contributed analysis tools; S.S. wrote the paper with input from M.W., T.N.S., and H.B.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wohlgemuth, T.; Kull, P.; Wüthrich, H. Disturbance of microsites and early tree regeneration after windthrow in Swiss mountain forests due to the winter storm Vivian 1990. For. Snow Landsc. Res. 2002, 77, 17–47. [Google Scholar]
- Eichrodt, R. Über die Bedeutung von Moderholz für die Natürliche Waldverjüngung im Subalpinen Fichtenwald; Beiheft zu den Zeitschriften des Schweizerischen Forstvereins; Bühler Buchdruck: Zurich, Switzerland, 1970; Volume 45. [Google Scholar]
- Ott, E.; Frehner, M.; Frey, H.-U.; Lüscher, P. Gebirgsnadelwälder; Verlag Paul Haupt Berne: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 1997; ISBN 3-258-05601-3. [Google Scholar]
- Brang, P. Resistance and elasticity: Promising concepts for the management of protection forests in the European Alps. For. Ecol. Manag. 2001, 145, 107–119. [Google Scholar] [CrossRef]
- Landolt, E. Unsere Alpenflora, 7th ed.; SAC-Verlag: Bern, Switzerland, 2003; ISBN 3-85902-218-0. [Google Scholar]
- Leibundgut, H. Die Natürliche Waldverjüngung; Verlag Paul Haupt Berne: Bern, Switzerland; Stuttgart, Germany, 1984; ISBN 3-258-03415-X. [Google Scholar]
- Svoboda, M.; Fraver, S.; Janda, P.; Bače, R.; Zenáhlíková, J. Natural development and regeneration of a Central European montane spruce forest. For. Ecol. Manag. 2010, 260, 707–714. [Google Scholar] [CrossRef]
- Takahashi, K. Effect of size structure, forest floor type and disturbance regime on tree species composition in a coniferous forest in Japan. J. Ecol. 1994, 82, 769–773. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1986; Volume 15, pp. 133–302. ISBN 978-0-12-013915-6. [Google Scholar]
- Kupferschmid, A.D.; Bugmann, H. Effect of microsites, logs and ungulate browsing on Picea abies regeneration in a mountain forest. For. Ecol. Manag. 2005, 205, 251–265. [Google Scholar] [CrossRef]
- Zielonka, T. When does dead wood turn into a substrate for spruce replacement? J. Veg. Sci. 2006, 17, 739–746. [Google Scholar] [CrossRef]
- Szewczyk, J.; Szwagrzyk, J. Tree regeneration on rotten wood and on soil in old-growth stand. Vegetatio 1996, 122, 37–46. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Hofgaard, A. The structure and regeneration of high-altitude Norway spruce forests: A review of Arnborg (1942, 1943). Scand. J. For. Res. 2011, 26, 17–24. [Google Scholar] [CrossRef]
- Anderson, L.J.; Winterton, A.J. Germination as a determinant of seedling distributions among natural substrates in Picea engelmannii (Pinaceae) and Abies lasiocarpa (Pinaceae). Am. J. Bot. 1996, 83, 112–117. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer seedling recruitment in a southeastern Canadian boreal forest: The importance of substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Narukawa, Y.; Yamamoto, S. Development of conifer seedlings roots on soil and fallen logs in boreal and subalpine coniferous forests of Japan. For. Ecol. Manag. 2003, 175, 131–139. [Google Scholar] [CrossRef]
- Narukawa, Y.; Iida, S.; Tanouchi, H.; Abe, S.; Yamamoto, S. State of fallen logs and the occurrence of conifer seedlings and saplings in boreal and subalpine old-growth forests of central Japan. Ecol. Res. 2003, 18, 267–277. [Google Scholar] [CrossRef]
- Staniaszek-Kik, M.; Żarnowiec, J.; Chmura, D. The vascular plant colonization on decaying Picea abies logs in Karkonosze mountain forest belts: The effects of forest community type, cryptogam cover, log decomposition and forest management. Eur. J. For. Res. 2016, 135, 1145–1157. [Google Scholar] [CrossRef]
- Zielonka, T.; Piątek, G. Norway spruce regeneration on decaying logs in subalpine forests in the Tatra National Park. Pol. Bot. J. 2001, 46, 251–260. [Google Scholar]
- Heinemann, K.; Kitzberger, T. Effects of position, understorey vegetation and coarse woody debris on tree regeneration in two environmentally contrasting forests of north-western Patagonia: A manipulative approach. J. Biogeogr. 2006, 33, 1357–1367. [Google Scholar] [CrossRef]
- Chmura, D.; Żarnowiec, J.; Staniaszek-Kik, M. Interactions between plant traits and environmental factors within and among montane forest belts: A study of vascular species colonising decaying logs. For. Ecol. Manag. 2016, 379, 216–225. [Google Scholar] [CrossRef]
- Kathke, S.; Bruelheide, H. Interaction of gap age and microsite type for the regeneration of Picea abies. For. Ecol. Manag. 2010, 259, 1597–1605. [Google Scholar] [CrossRef]
- Bače, R.; Svoboda, M.; Pouska, V.; Janda, P.; Červenka, J. Natural regeneration in Central-European subalpine spruce forests: Which logs are suitable for seedling recruitment? For. Ecol. Manag. 2012, 266, 254–262. [Google Scholar] [CrossRef]
- Hillgarter, F. Waldbauliche und Ertragskundliche Untersuchungen im Subalpinen Fichtenurwald Scatlè/Brigels. Ph.D. Thesis, ETH Zurich, Zurich, Switzerland, 1971. [Google Scholar]
- AWN, G. Naturwaldreservat Scatlè. 2011. Available online: http://www.webcitation.org/6x4wxntEI (accessed on 14 December 2012).
- Bugmann, H. The Forest Reserve “Scatlè”: A Brief Overview; ETH Zurich: Zurich, Switzerland, 2010. [Google Scholar]
- Heiri, C.; Hallenbarter, D. Der Urwald Scatlè. In Waldreservate. 50 Jahre Natürliche Waldentwicklung in der Schweiz; Brang, P., Heiri, C., Bugmann, H., Eds.; Haupt: Bern, Switzerland; Stuttgart, Germany; Wien, Austria, 2011; ISBN 978-3-258-07725-3. [Google Scholar]
- Renvall, P. Community structure and dynamics of wood-rotting Basidiomycetes on decomposing co-nifer trunks in northern Finland. Karstenia 1995, 35, 1–52. [Google Scholar] [CrossRef]
- Stahel, W. Statistische Datenanalyse, 5th ed.; Vieweg + Teubner: Wiesbaden, Germany, 2008; ISBN 978-3-8348-0410-5. [Google Scholar]
- Zuur, A.F.; Savaliev, A.A.; Ieno, E.N. Zero Inflated Models and Generalized Linear Mixed Models with R; Highland Statistics Ltd.: Newburgh, NY, USA, 2012; ISBN 0-9571741-1-X. [Google Scholar]
- Dormann, C.F. Parametrische Statistik. In Verteilungen, Maximum Likelihood and GLM in R; Springer: Berlin, Germany, 2017. [Google Scholar]
- Rohner, B.; Bigler, C.; Wunder, J.; Brang, P.; Bugmann, H. Fifty years of natural succession in Swiss forest reserves: Changes in stand structure and mortality rates of oak and beech. J. Veg. Sci. 2012, 23, 892–905. [Google Scholar] [CrossRef]
- Frazer, G. Gap Light Analyzer (GLA): Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs. Users Manual and Program Documentation, version 2.0; Simon Fraser University: Burnaby, BC, Canada; Institute of Ecosystem Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- Sieber, T.N. Pyrenochaeta ligni-putridi sp. nov., a new coelomycete associated with butt rot of Picea abies in Switzerland. Mycol. Res. 1995, 99, 274–276. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Queloz, V.; Grünig, C.R.; Berndt, R.; Kowalski, T.; Sieber, T.N.; Holdenrieder, O. Cryptic speciation in Hymenoscyphus albidus. For. Pathol. 2011, 41, 133–142. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Breitenbach, J.; Kränzlin, F. Fungi of Switzerland—Non Gilled Fungi; Mycologia: Lucerne, Switzerland, 1986; Volume 2, ISBN 3-85604-220-2. [Google Scholar]
- Dittrich, S.; Jacob, M.; Bade, C.; Leuschner, C.; Hauck, M. The significance of deadwood for total bryophyte, lichen, and vascular plant diversity in an old-growth spruce forest. Plant Ecol. 2014, 215, 1123–1137. [Google Scholar] [CrossRef]
- Iijima, H.; Shibuya, M.; Saito, H. Effects of surface and light conditions of fallen logs on the emergence and survival of coniferous seedlings and saplings. J. For. Res. 2007, 12, 262–269. [Google Scholar] [CrossRef]
- Zielonka, T.; Piątek, G. The herb and dwarf shrubs colonization of decaying logs in subalpine forest in the Polish Tatra Mountains. Plant Ecol. 2004, 172, 63–72. [Google Scholar] [CrossRef]
- Christy, E.J.; Mack, R.N. Variation in demography of juvenile Tsuga heterophylla across the substratum mosaic. J. Ecol. 1984, 72, 75–91. [Google Scholar] [CrossRef]
- Kimmins, J. Forest Ecology: A Foundation for Sustainable Forest Management and Environmental Ethics in Forestry, 3rd ed.; Pearson Prentice Hall: London, NY, USA, 2004; ISBN 0-13-066258-5. [Google Scholar]
- Schmidt-Vogt, H. Die Fichte. Band 1: Taxonomie, Verbreitung, Morphologie, Ökologie, Waldgesellschaften; Paul Parey: Hamburg/Berlin, Germany, 1987; ISBN 3-490-08216-8. [Google Scholar]
- Pisek, A.; Winkler, E. Licht- und Temperaturabhängigkeit der CO2-Assimilation von Fichte (Picea excelsa Link), Zirbe (Pinus cembra L.) und Sonnenblume (Helianthus annuus L.). Planta 1959, 53, 532–550. [Google Scholar] [CrossRef]
- Harmon, M.E. Effects of bark fragmentation on plant succession on conifer logs in the Picea-Tsuga forests of Olympic National Park, Washington. Am. Midl. Nat. 1989, 121, 112–124. [Google Scholar] [CrossRef]
- Wasser, B.; Frehner, M. Wegleitung Minimale Pflegemassnahmen für Wälder mit Schutzfunktion; Bundesamt für Umwelt, Wald und Landschaft BUWAL: Bern, Switzerland, 1996. [Google Scholar]
- Ostrofsky, A.; Jellison, J.; Smith, K.T.; Shortle, W.C. Changes in cation concentrations in red spruce wood decayed by brown rot and white rot fungi. Can. J. For. Res. 1997, 27, 567–571. [Google Scholar] [CrossRef]
- Fukasawa, Y. Effects of wood decomposer fungi on tree seedling establishment on coarse woody debris. For. Ecol. Manag. 2012, 266, 232–238. [Google Scholar] [CrossRef]
- Harvey, A.E.; Jurgensen, M.F.; Larsen, M.J. Seasonal distribution of ectomycorrhizae in a mature Douglas-fir/Larch forest soil in western Montana. For. Sci. 1978, 24, 203–208. [Google Scholar]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- Červenka, J.; Bače, R.; Svoboda, M. Stand-replacing disturbance does not directly alter the succession of Norway spruce regeneration on dead wood. J. For. Sci. 2014, 60, 417–424. [Google Scholar] [CrossRef]
- Takahashi, M.; Sakai, Y.; Ootomo, R.; Shiozaki, M. Establishment of tree seedlings and water-soluble nutrients in coarse woody debris in an old-growth Picea abies forest in Hokkaido, northern Japan. Can. J. For. Res. 2000, 30, 1148–1155. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Robert, E.; Brais, S.; Harvey, B.D.; Greene, D. Seedling establishment and survival on decaying logs in boreal mixedwood stands following a mast year. Can. J. For. Res. 2012, 42, 1446–1455. [Google Scholar] [CrossRef]
- Sugita, H.; Nagaike, T. Microsites for seedling establishment of subalpine conifers in a forest with moss-type undergrowth on Mt. Fuji, central Honshu, Japan. Ecol. Res. 2005, 20, 678–685. [Google Scholar] [CrossRef]
- Chmura, D.; Zarnowiec, J.; Staniaszek-Kik, M. Comparison of traits of non-colonized and colonized decaying logs by vascular plant species. IForest-Biogeosci. For. 2018, 11, 11–16. [Google Scholar] [CrossRef]
- Nakamura, T. Effect of bryophytes on survival of conifer seedlings in subalpine forests of central Japan. Ecol. Res. 1992, 7, 155–162. [Google Scholar] [CrossRef]
- Doi, Y.; Mori, A.S.; Takeda, H. Conifer establishment and root architectural responses to forest floor heterogeneity in an old-growth subalpine forest in central Japan. For. Ecol. Manag. 2008, 255, 1472–1478. [Google Scholar] [CrossRef]
- Holeksa, J.; Zielonka, T.; Żywiec, M. Modeling the decay of coarse woody debris in a subalpine Norway spruce forest of the West Carpathians, Poland. Can. J. For. Res. 2008, 38, 415–428. [Google Scholar] [CrossRef]
- Botting, R.S.; DeLong, C. Macrolichen and bryophyte responses to coarse woody debris characteristics in sub-boreal spruce forest. For. Ecol. Manag. 2009, 258, S85–S94. [Google Scholar] [CrossRef]
- Allmér, J.; Vasiliauskas, R.; Ihrmark, K.; Stenlid, J.; Dahlberg, A. Wood-inhabiting fungal communities in woody debris of Norway spruce (Picea abies (L.) Karst.), as reflected by sporocarps, mycelial isolations and T-RFLP identification: Wood-inhabiting fungal communities in woody debris of Norway spruce. FEMS Microbiol. Ecol. 2006, 55, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Rajala, T.; Peltoniemi, M.; Pennanen, T.; Mäkipää, R. Fungal community dynamics in relation to substrate quality of decaying Norway spruce (Picea abies (L.) Karst.) logs in boreal forests. FEMS Microbiol. Ecol. 2012, 81, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.M.; Work, T.T. Linking deadwood-associated beetles and fungi with wood decomposition rates in managed black spruce forests. Can. J. For. Res. 2012, 42, 1477–1490. [Google Scholar] [CrossRef]
- Schmidt, O. Wood and Tree Fungi; Springer: Berlin/Heidelberg, Germany, 2006; ISBN 978-3-540-32138-5. [Google Scholar]
- Lindblad, I. Wood-inhabiting fungi on fallen logs of Norway spruce: Relations to forest management and substrate quality. Nord. J. Bot. 1998, 18, 243–255. [Google Scholar] [CrossRef]
- Penttilä, R.; Siitonen, J.; Kuusinen, M. Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol. Conserv. 2004, 117, 271–283. [Google Scholar] [CrossRef]
- Jönsson, M.T.; Edman, M.; Jonsson, B.G. Colonization and extinction patterns of wood-decaying fungi in a boreal old-growth Picea abies forest: Colonization-extinction dynamics of wood-decaying fungi. J. Ecol. 2008, 96, 1065–1075. [Google Scholar] [CrossRef]
- Høiland, K.; Bendiksen, E. Biodiversity of wood-inhabiting fungi in a boreal coniferous forest in Sør-Trøndelag County, Central Norway. Nord. J. Bot. 1996, 16, 643–659. [Google Scholar] [CrossRef]
- Petrillo, M.; Cherubini, P.; Sartori, G.; Abiven, S.; Ascher, J.; Bertoldi, D.; Camin, F.; Barbero, A.; Larcher, R.; Egli, M. Decomposition of Norway spruce and European larch coarse woody debris (CWD) in relation to different elevation and exposure in an Alpine setting. iForest-Biogeosci. For. 2016, 9, 154–164. [Google Scholar] [CrossRef]
- Lombardi, F.; Cherubini, P.; Tognetti, R.; Cocozza, C.; Lasserre, B.; Marchetti, M. Investigating biochemical processes to assess deadwood decay of beech and silver fir in Mediterranean mountain forests. Ann. For. Sci. 2013, 70, 101–111. [Google Scholar] [CrossRef]
- Bütler, R.; Patty, L.; Le Bayon, R.-C.; Guenat, C.; Schlaepfer, R. Log decay of Picea abies in the Swiss Jura Mountains of central Europe. For. Ecol. Manag. 2007, 242, 791–799. [Google Scholar] [CrossRef]
Figure 1.
Significant variables for the seedling model: (a) type of rot (WR = white rot, BR = brown rot), (b) decay stage, (c) plot inclination, (d) diameter, (e) percentage of total radiation, and (f) time of tree fall (1 = before 1965, 2 = 1965–1977, 3= 1977–1989, 4 = 1989–2006, 5 = after 2006). The straight line represents the model fit, the grey shaded area the confidence interval. The rhombuses in (a,b,f) indicate the mean seedling density.
Figure 1.
Significant variables for the seedling model: (a) type of rot (WR = white rot, BR = brown rot), (b) decay stage, (c) plot inclination, (d) diameter, (e) percentage of total radiation, and (f) time of tree fall (1 = before 1965, 2 = 1965–1977, 3= 1977–1989, 4 = 1989–2006, 5 = after 2006). The straight line represents the model fit, the grey shaded area the confidence interval. The rhombuses in (a,b,f) indicate the mean seedling density.

Figure 2.
Significant variables for the sapling model: (a) diameter of stems, (b) moss cover, (c) percentage of total radiation, (d) distance from plot surface to forest floor, (e) bark cover, and (f) time of tree fall (1 = before 1965, 2 = 1965–1977, 3= 1977–1989, 4 = 1989–2006, 5 = after 2006). The straight line represents the model fit, the grey shaded area the confidence interval. The rhombuses in (f) indicate the mean sapling density.
Figure 2.
Significant variables for the sapling model: (a) diameter of stems, (b) moss cover, (c) percentage of total radiation, (d) distance from plot surface to forest floor, (e) bark cover, and (f) time of tree fall (1 = before 1965, 2 = 1965–1977, 3= 1977–1989, 4 = 1989–2006, 5 = after 2006). The straight line represents the model fit, the grey shaded area the confidence interval. The rhombuses in (f) indicate the mean sapling density.
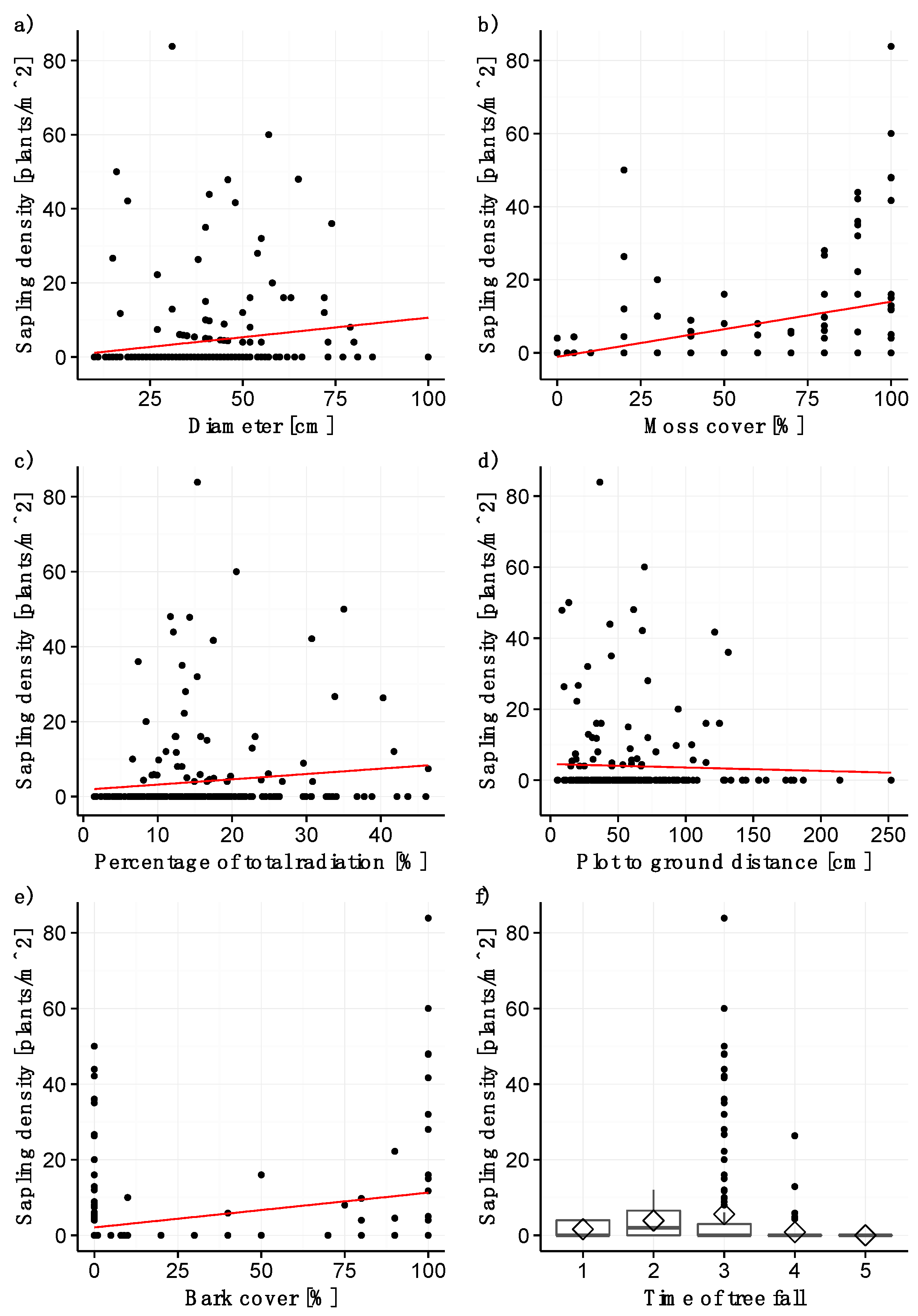
Figure 3.
Non-metric multidimensional scaling of the different fungal species in relation to the decay stage (DS) of the stems. Species within boxes were solely associated with the corresponding decay stage. Increment-boring-core data only. See Table 3 for abbreviations of fungal species names.
Figure 3.
Non-metric multidimensional scaling of the different fungal species in relation to the decay stage (DS) of the stems. Species within boxes were solely associated with the corresponding decay stage. Increment-boring-core data only. See Table 3 for abbreviations of fungal species names.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Minimum, median, mean, and maximum values of the variables assessed on each plot (not including factorial variables).
Table 1.
Minimum, median, mean, and maximum values of the variables assessed on each plot (not including factorial variables).
| Variable | Unit | Minimum | Median | Mean | Maximum |
|---|---|---|---|---|---|
| Average sunshine duration in June | Min/day | 1.47 | 76.07 | 95.63 | 361.19 |
| Bark cover | % | 0 | 0 | 20.74 | 100 |
| Vegetation coverage | % | 0 | 0 | 9.34 | 130 |
| Canopy openness | % | 1.22 | 9.76 | 9.95 | 23.7 |
| Log diameter | cm | 10 | 26 | 37.23 | 100 |
| Moss cover | % | 0 | 20 | 33.73 | 100 |
| Moss height | cm | 0 | 1.5 | 1.63 | 8 |
| No. of individuals | Count | 0 | 6 | 8.99 | 61 |
| No. of saplings >10 cm | Count | 0 | 0 | 0.79 | 15 |
| No. of seedlings | Count | 0 | 4 | 6.3 | 56 |
| Plot inclination | ° | 0 | 20 | 19.99 | 65 |
| Plot to ground distance | cm | 5 | 50 | 57.87 | 252 |
| Tallest individual | cm | 0 | 2.3 | 7.71 | 135 |
| Total radiation | % | 1.47 | 13.95 | 15.68 | 46.38 |
Table 2.
Deviance, performance, robustness, and significance of the different variables used in the final model for the seedling (recently germinated plants with the characteristic four to nine cotyledons) and sapling (height > 10 cm) density on the plots.
Table 2.
Deviance, performance, robustness, and significance of the different variables used in the final model for the seedling (recently germinated plants with the characteristic four to nine cotyledons) and sapling (height > 10 cm) density on the plots.
| Seedling Density | Sapling Density | |||
|---|---|---|---|---|
| Deviance | 275.1 | 170.9 | ||
| Performance | r = 0.62 | r = 0.72 | ||
| Robustness | r = 0.21 | r = 0.66 | ||
| Variable | p | Estimate | p | Estimate |
| White rot | 8.08 × 10−7 (***) | 7.217 × 10−1 | - | - |
| Brown rot | 1.26 × 10−5 (***) | 7.897 × 10−1 | - | - |
| White & brown rot | 3.13 × 10−7 (***) | 8.995 × 10−1 | - | - |
| Decay stage 3 | 0.0266 (*) | 2.898 × 10−1 | - | - |
| Decay stage 4 | 0.0043 (**) | 3.982 × 10−1 | - | - |
| Plot inclination | 0.0004 (***) | −1.118 × 10−1 | - | - |
| Diameter | 0.0002 (***) | 8.969 × 10−1 | 9.62 × 10−10 (***) | 3.687 × 10−1 |
| Moss cover | - | <2 × 10−16 (***) | 3.230 × 10−1 | |
| % of total radiation | 0.0689 (.) | −8.986 × 10−1 | 1.68 × 10−8 (***) | 7.601 × 10−1 |
| Time of tree fall: 1977–1989 | 0.1275 (n.s.) | 5.988 × 10−1 | 0.0031 (**) | 2.189 |
| Bark cover | - | - | 0.0288 (*) | 5.419 × 10−1 |
| Distance plot to ground | - | - | 0.0030 (**) | −9.195 × 10−1 |
Significance codes: *** p ≤ 0.001; ** 0.001 < p ≤ 0.01; * 0.01 < p ≤ 0.05; . 0.05 < p ≤ 0.1.
Table 3.
Macroscopically identified and sequenced wood decay fungi, GenBank accession number of closest match and own isolates, frequencies and type of rot [38].
Table 3.
Macroscopically identified and sequenced wood decay fungi, GenBank accession number of closest match and own isolates, frequencies and type of rot [38].
| Species | Abbrev. | Identity | GenBank Accession Number of Closest Match | GenBank Accession Number | Isolation Frequency | Fruiting Body Frequency | Type of Rot |
|---|---|---|---|---|---|---|---|
| Amylostereum chailletii | AmyCha | 1 | GQ422470 | KT943921 | 2 | - | unknown |
| Amylostereum aerolatum | AmyAer | - | - | - | - | 1 | white |
| Antrodia serialis | AntSer | 0.99 | FM872469 | KT943942 KT943943 | 16 | - | brown |
| Athelia sp. | - | 0.98 | GU187537 | KT943944 | 1 | - | unknown |
| Climacocystis borealis | CliBor | 0.99 | JN710527 | KT943915 | 3 | - | white |
| Conferticium ochraceum | ConOch | 0.99 | AY781254 | KT943933 | 1 | - | unknown |
| Coniophora olivacea | ConOli | 0.99 | AJ345009 | KT943916 | 2 | - | brown |
| Coniophora puteana | ConPut | 1 | AM946631 | KT943922 KT943923 KT943924 | 7 | - | brown |
| Fomitopsis pinicola | FomPin | 1 | JX501306 | KT943917 KT943918 KT943919 KT943920 | 33 | 14 | brown |
| Fomitopsis rosea | FomRos | 0.99 | DQ491410 | KT943945 KT943946 | 11 | 8 | brown |
| Gloeophyllum sepiarium | GloSep | 1 | JN182924 | KT943931 | 1 | 2 | brown |
| Heterobasidion parviporum | HetPar | 1 | FJ903330 | KT943925 KT943926 | 5 | - | white |
| Hypochnicium punctulatum | HypPun | 1 | AF429412 | KT943936 | 1 | - | unknown |
| Hypochnicium subrigescens | HypSub | 0.99 | AF429427 | KT943939 | 1 | - | unknown |
| Ischnoderma benzoinum | IscBen | 1 | JQ518274 | KT943932 | 1 | - | white |
| Phanerochaete sordida | PhaSor | 1 | FJ228210 | KT943938 KT943940 | 4 | - | unknown |
| Phellinus ferrugineovelutinus | PheFer | 0.98 | AY558618 | KT943948 | 2 | - | unknown |
| Phellopilus nigrolimitatus | PheNig | 0.95 | JQ358813 | KT943927 KT943928 | 3 | - | white |
| Postia fragilis | PosFra | 1 | JF950573 | KT943930 | 1 | - | brown |
| Stereum rugosum | SteRug | 1 | FN539052 | KT943935 | 1 | - | white |
| Stereum sanguinolentum | SteSan | 0.99 | EU673086 | KT943941 | 2 | 1 | white |
| Trichaptum abietinum | TriAbi | - | - | - | - | 2 | white |
| Veluticeps abietina | VelAbi | 0.99 | EU118619 | KT943929 | 1 | - | unknown |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stroheker, S.; Weiss, M.; Sieber, T.N.; Bugmann, H. Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest. Forests 2018, 9, 120. https://doi.org/10.3390/f9030120
AMA Style
Stroheker S, Weiss M, Sieber TN, Bugmann H. Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest. Forests. 2018; 9(3):120. https://doi.org/10.3390/f9030120
Chicago/Turabian StyleStroheker, Sophie, Marc Weiss, Thomas N. Sieber, and Harald Bugmann. 2018. "Ecological Factors Influencing Norway Spruce Regeneration on Nurse Logs in a Subalpine Virgin Forest" Forests 9, no. 3: 120. https://doi.org/10.3390/f9030120
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.