Development of an Integrated DBH Estimation Model Based on Stand and Climatic Conditions


 ,
,
Abstract
:1. Introduction
2. Data and Methods
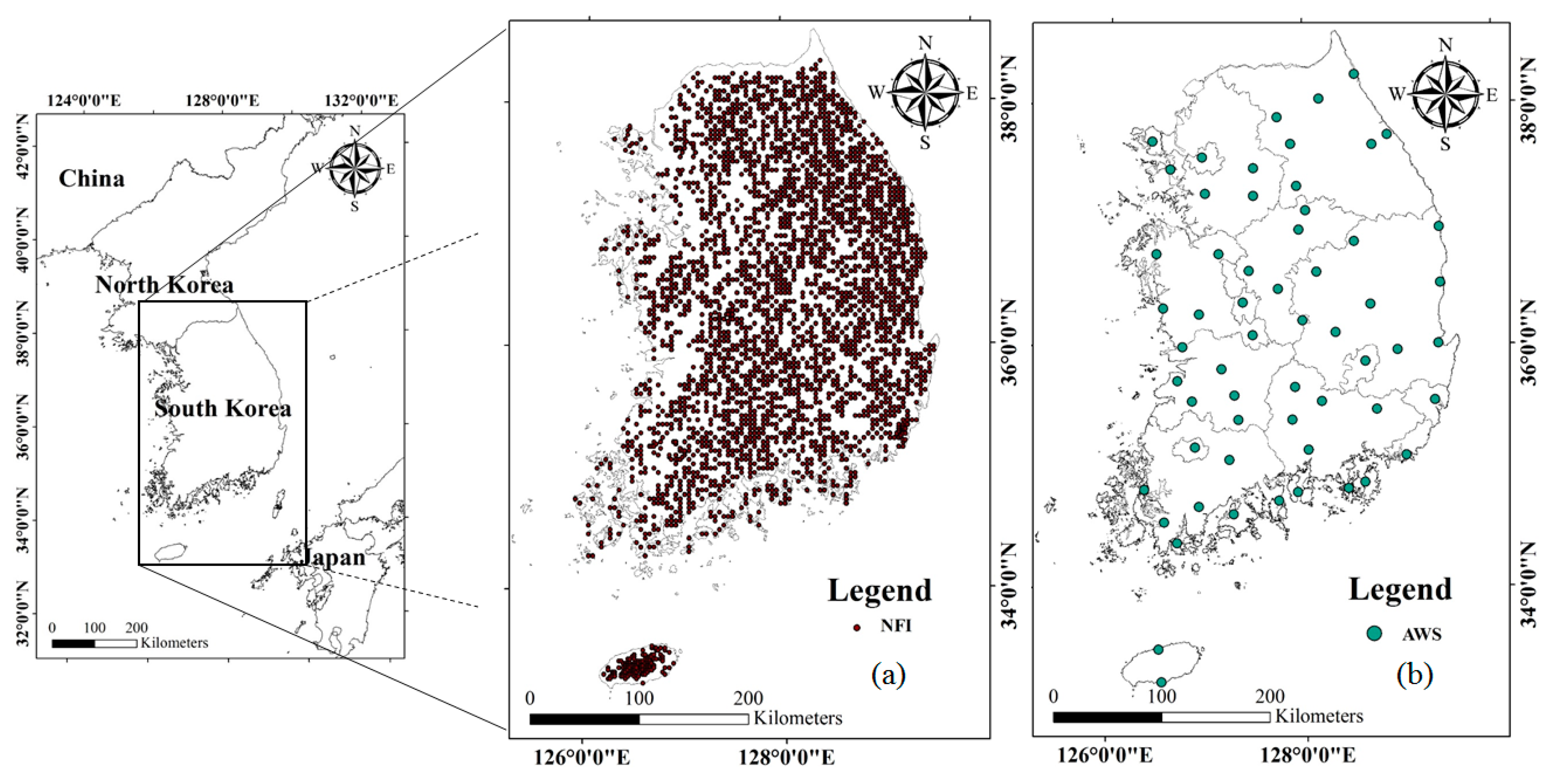
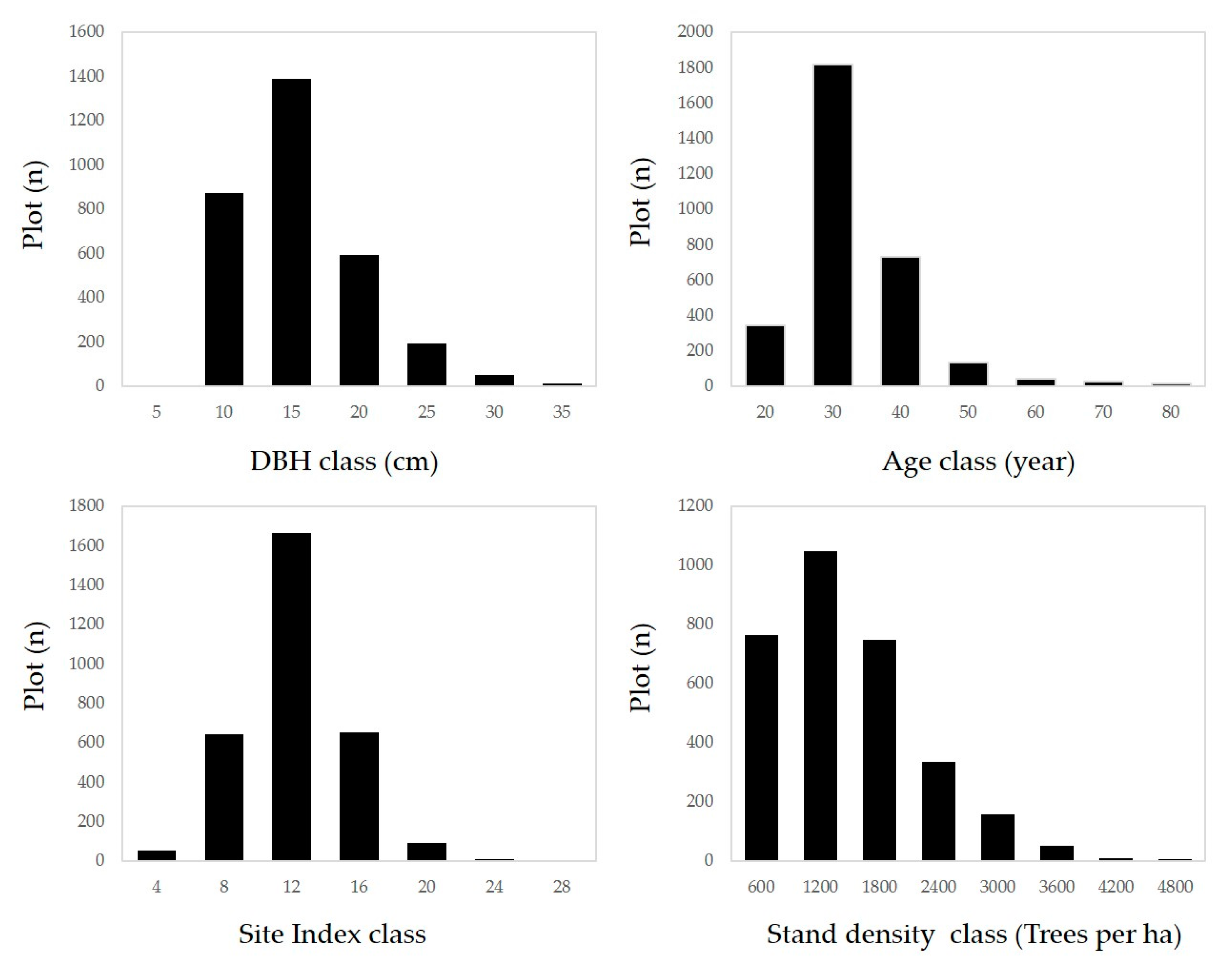
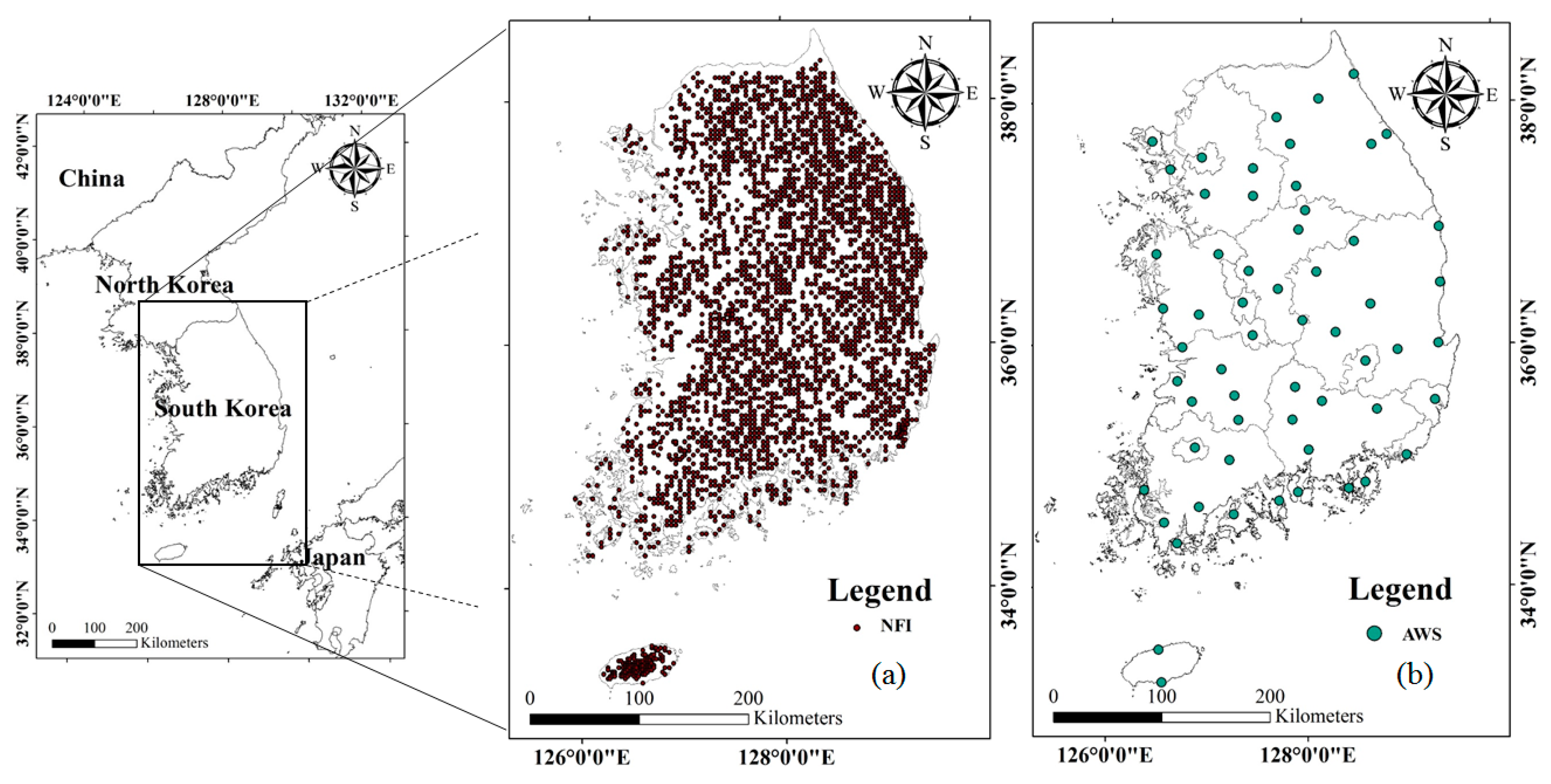
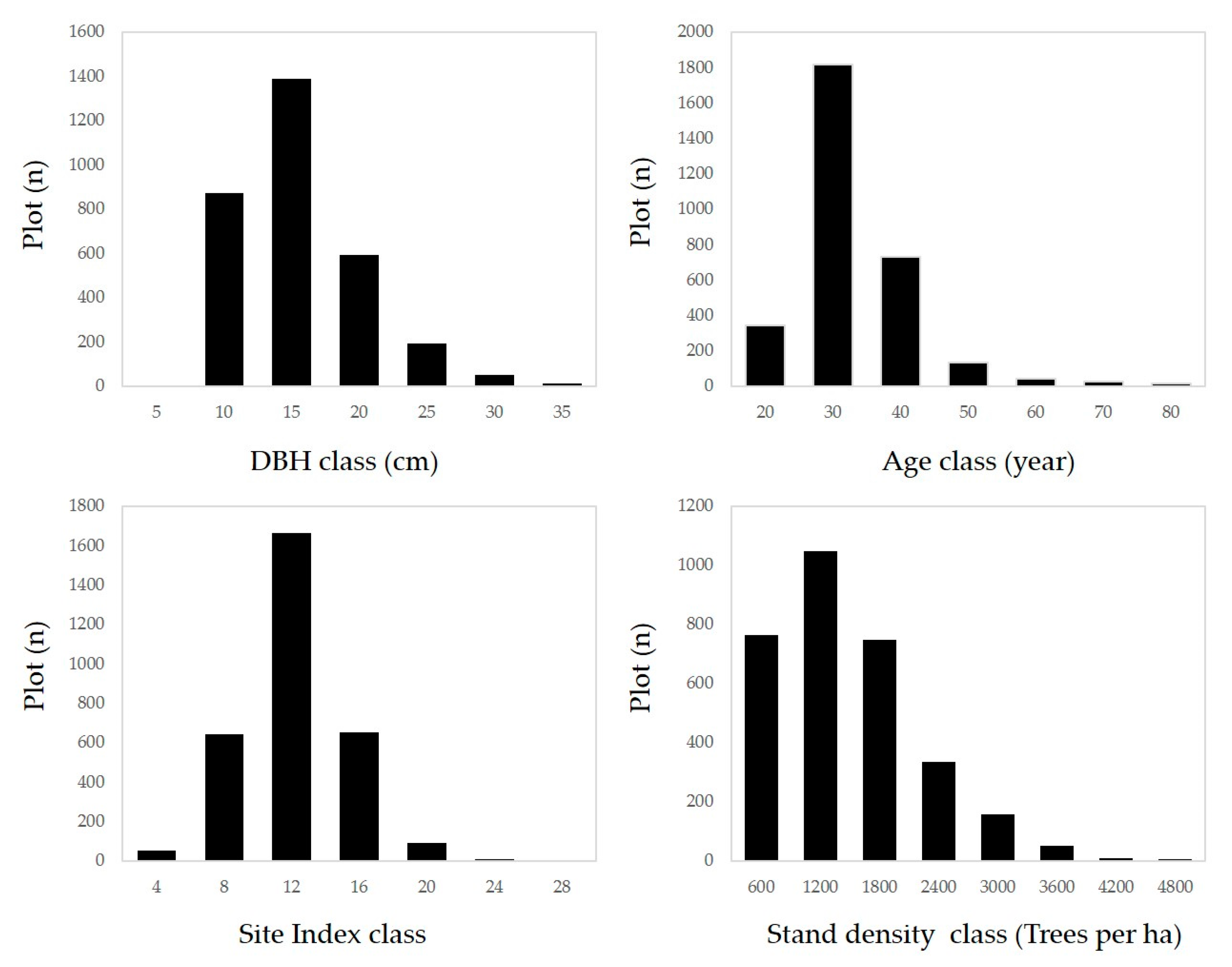
2.1. Study Area and Materials
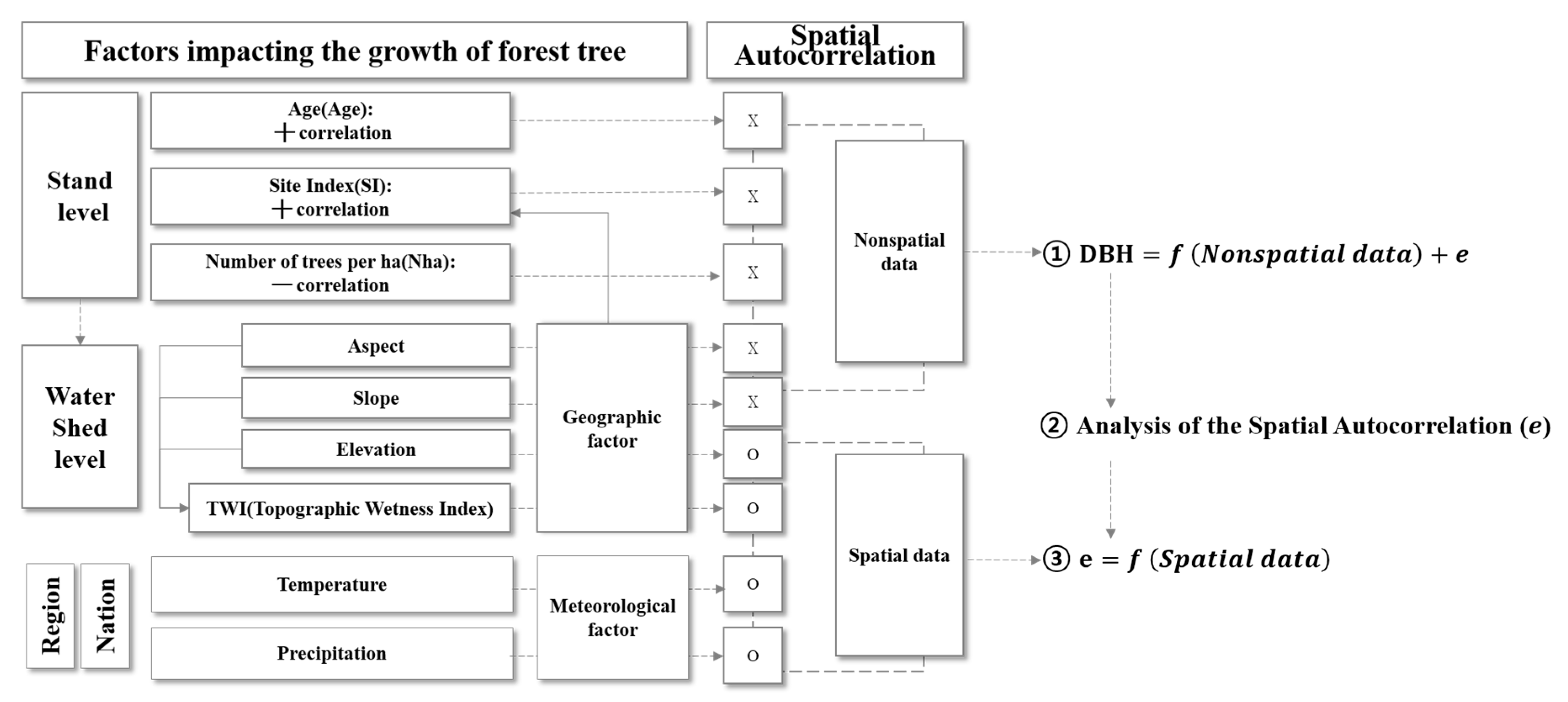
2.2. Method
2.2.1. DBH Estimation Model with Stand Age, SI and Nha
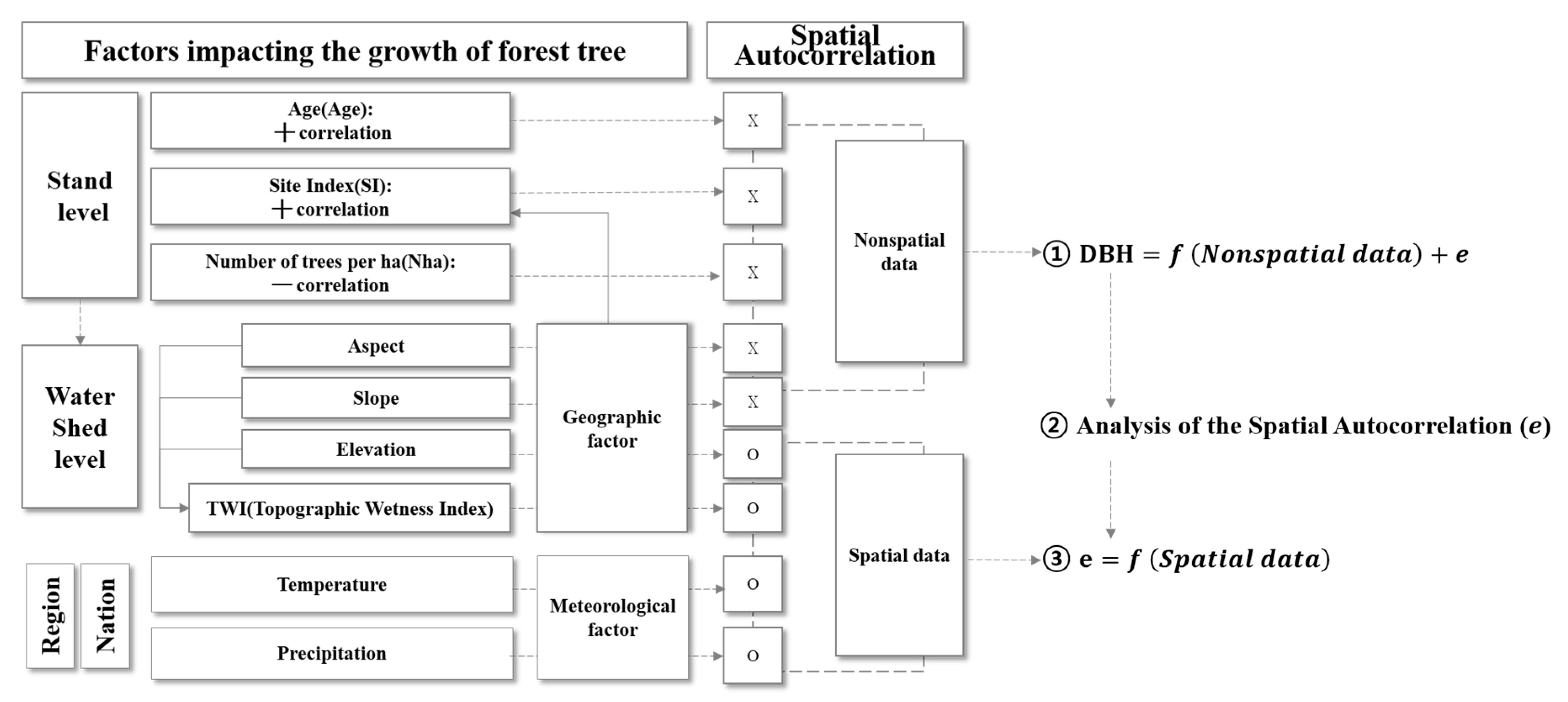
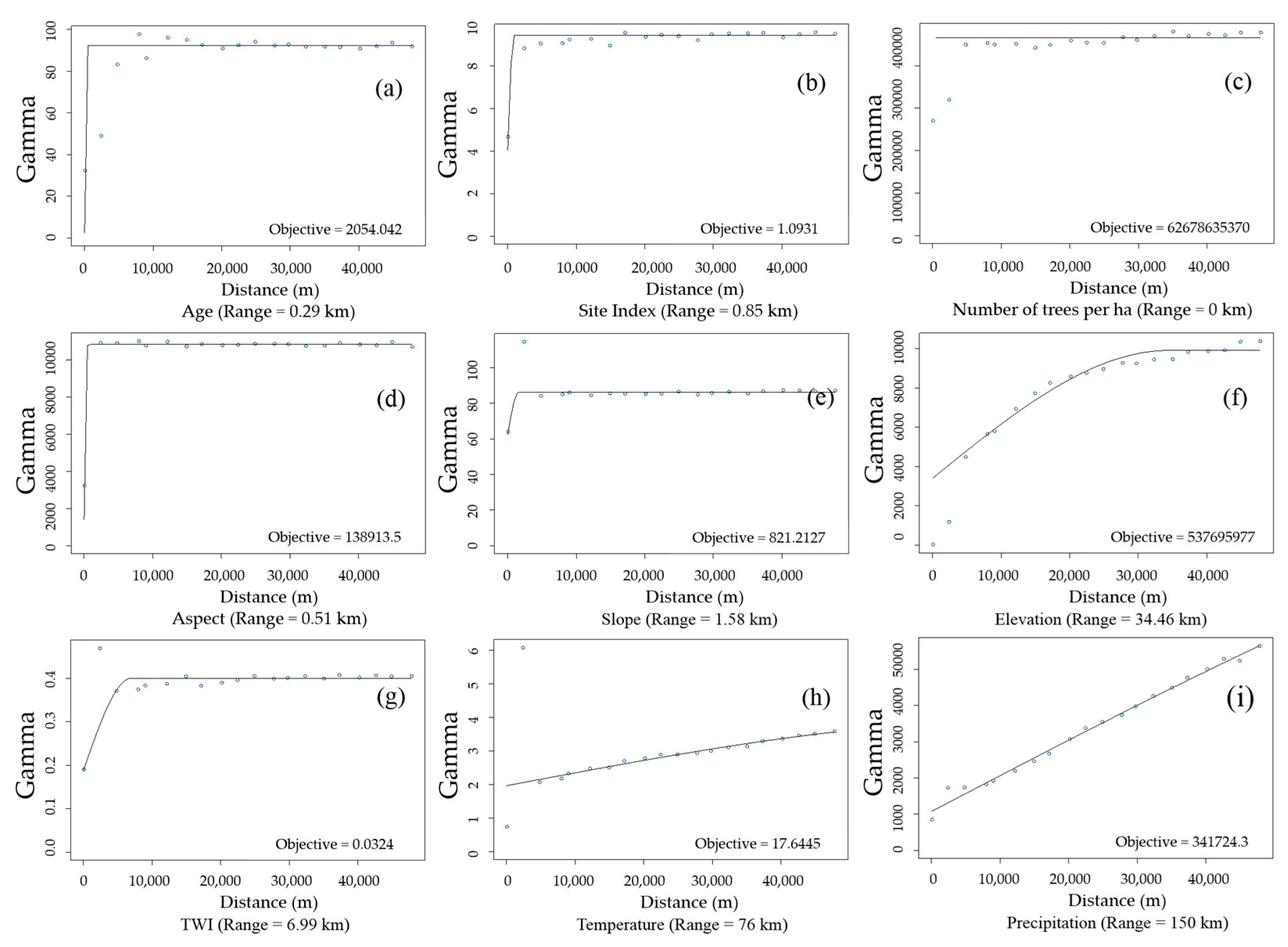
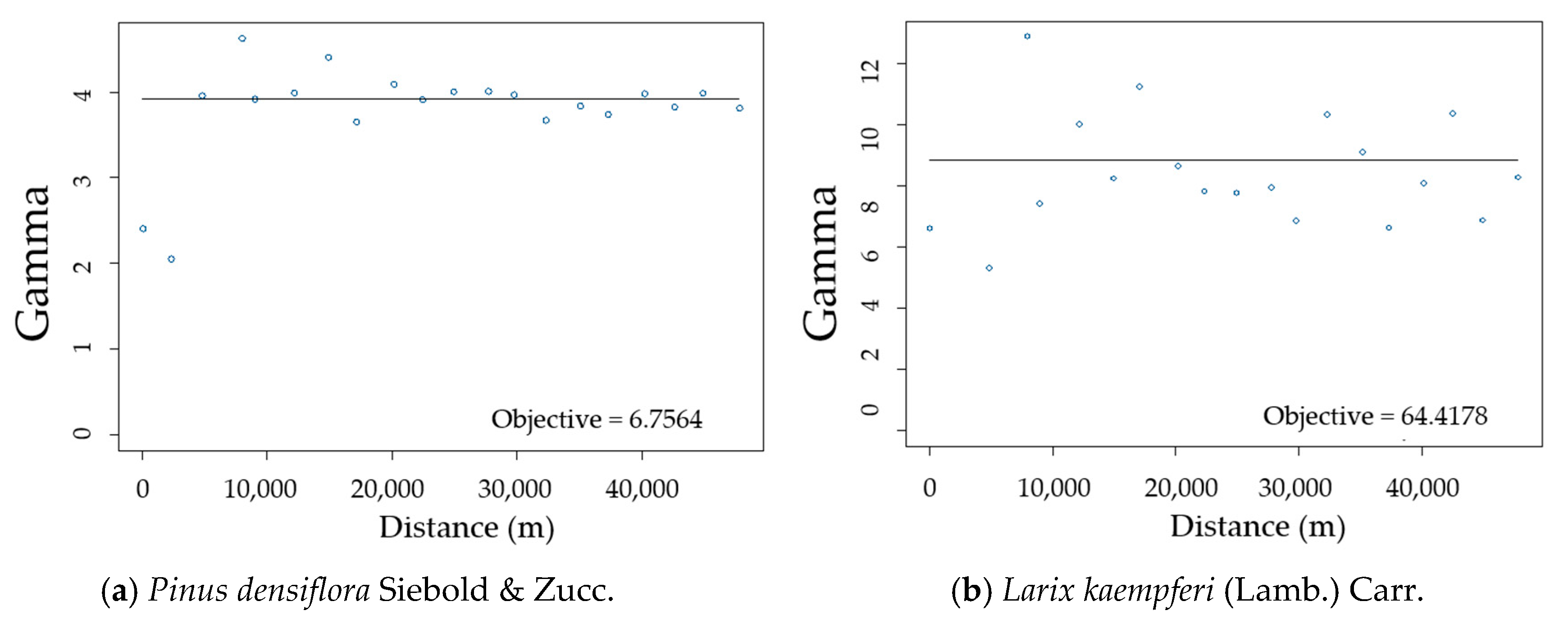
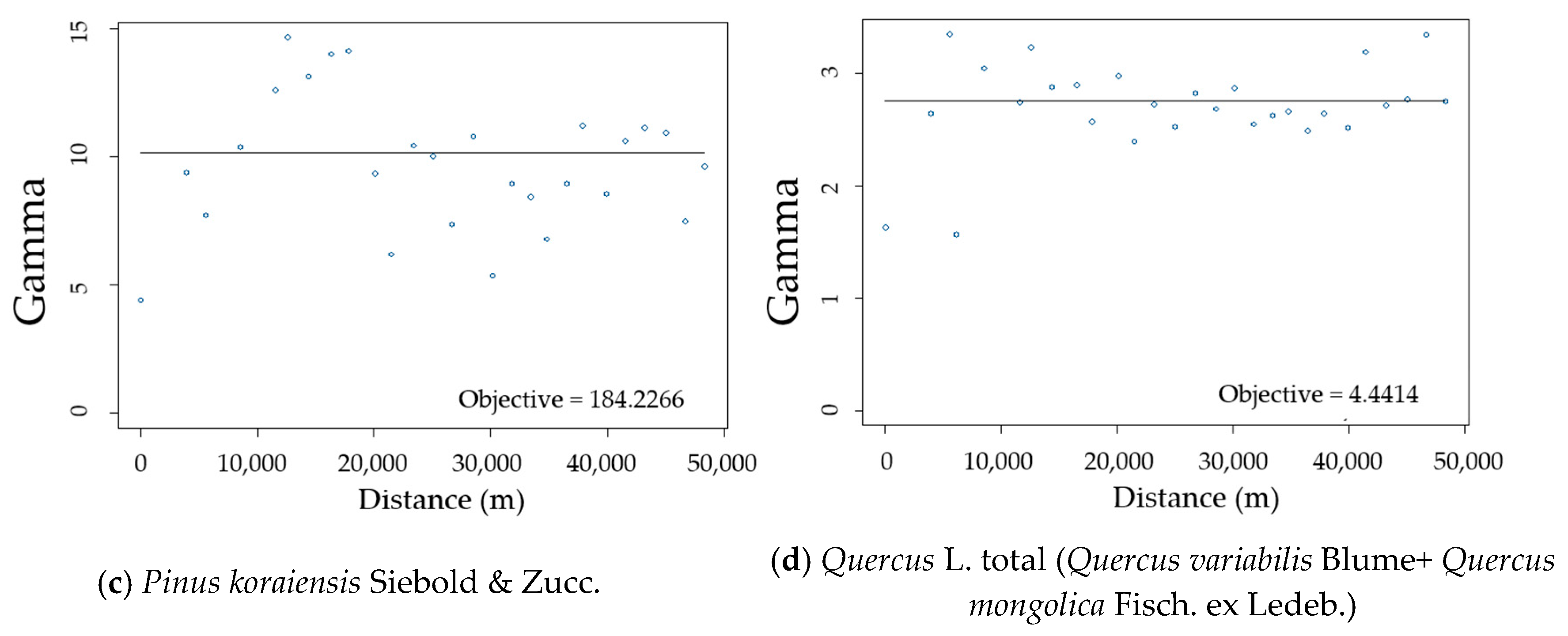
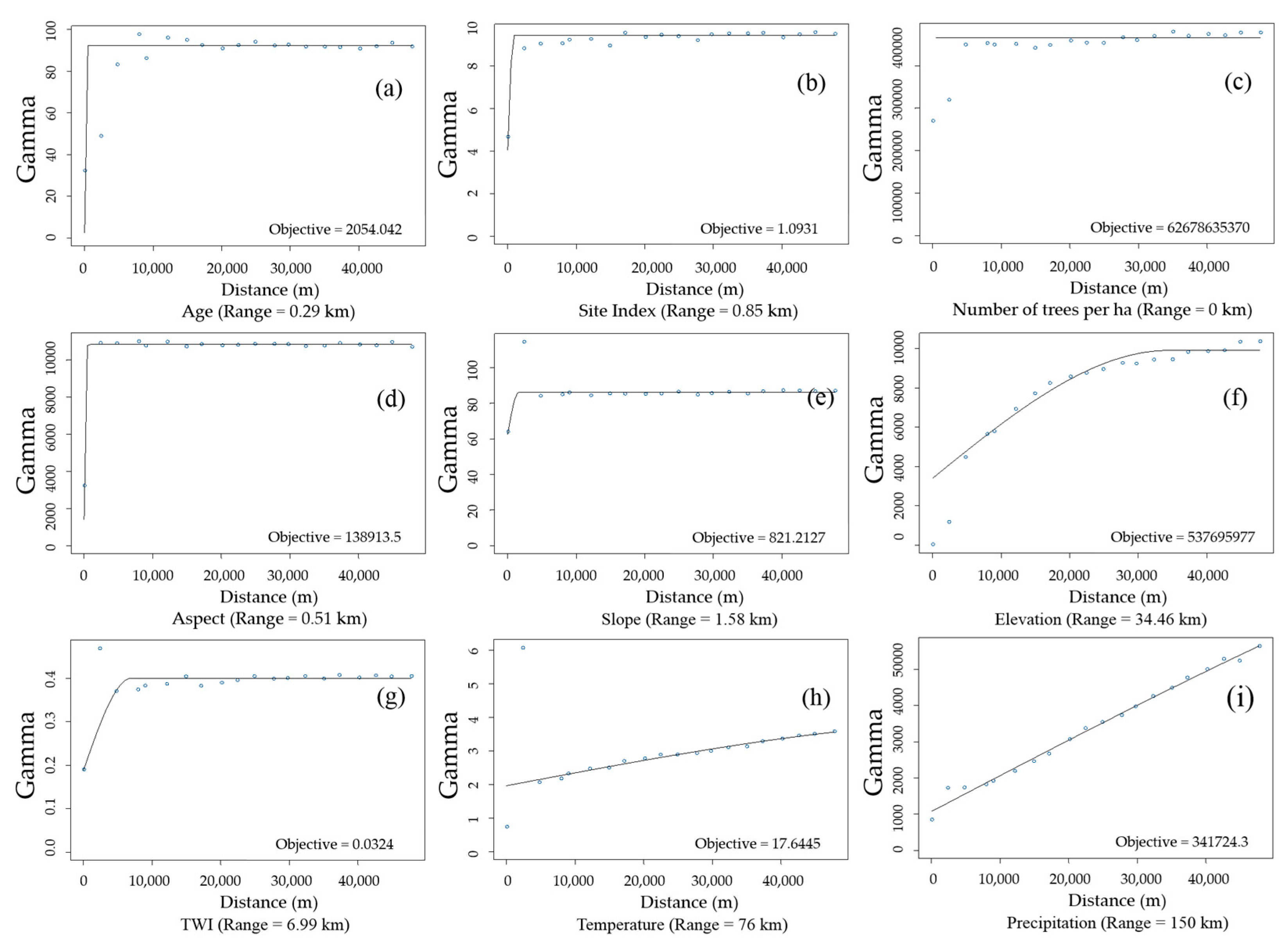
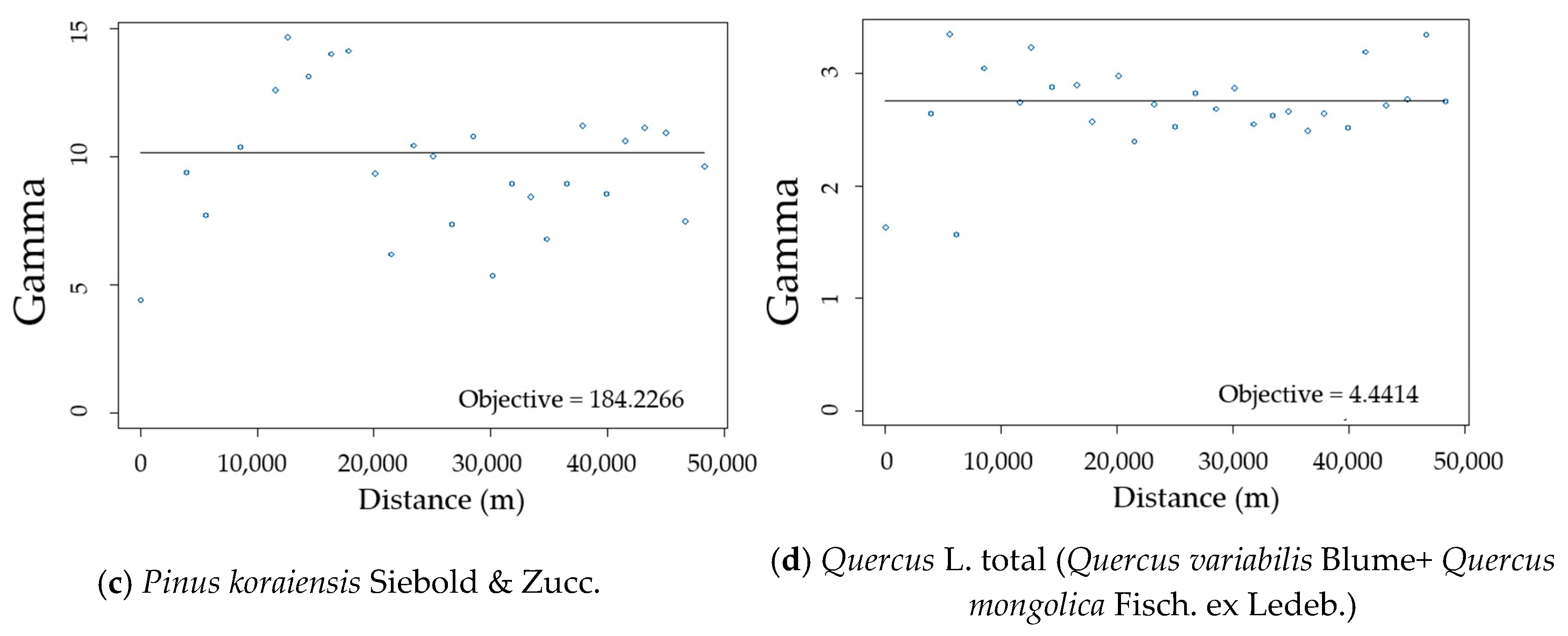
2.2.2. Semi-Variogram Analysis for Residuals of the DBH Estimation Model
2.2.3. Residual Model with Climate Variables
2.2.4. Integrated DBH Estimation Model with Stand-Level and Climate Variables
3. Results and Discussion
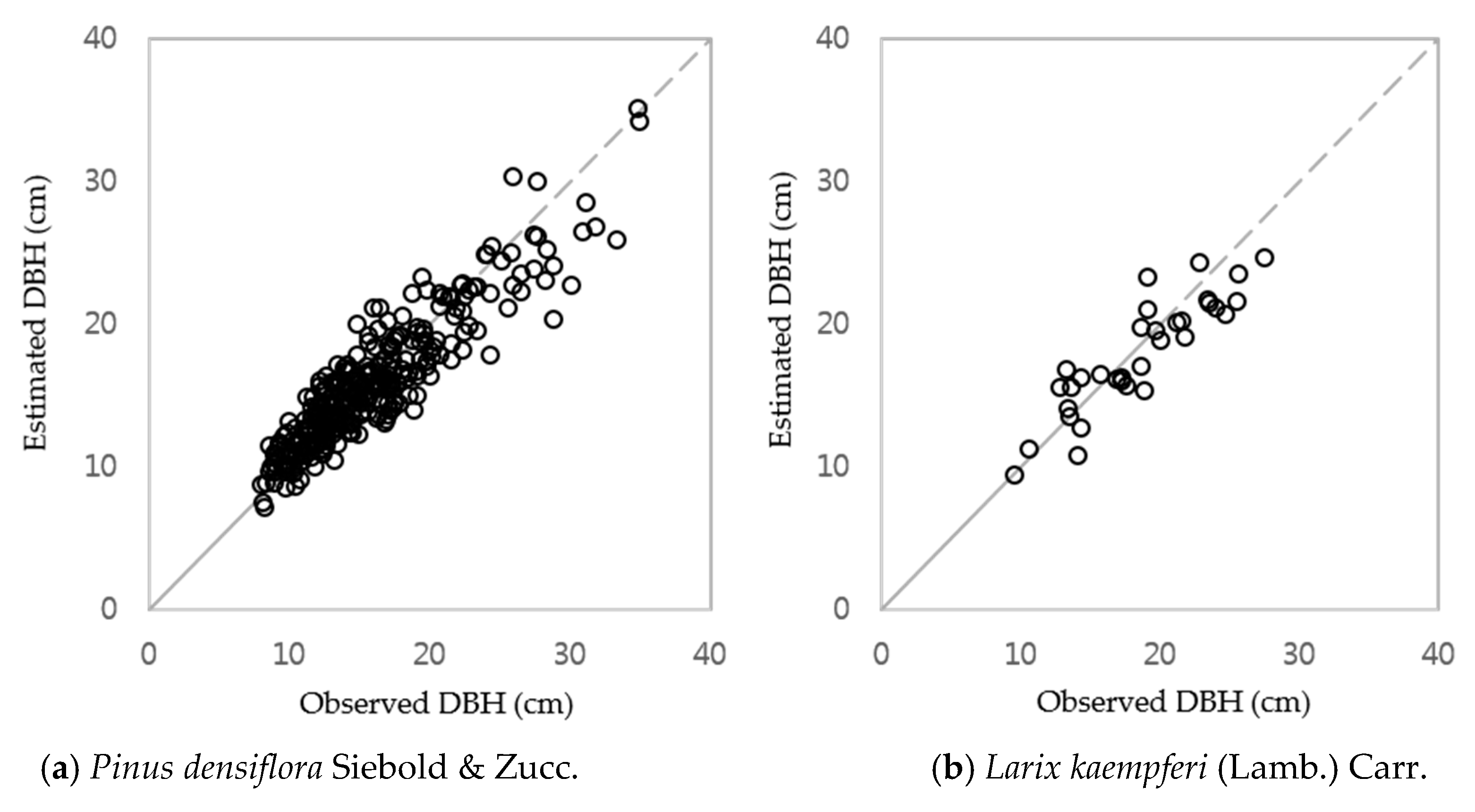
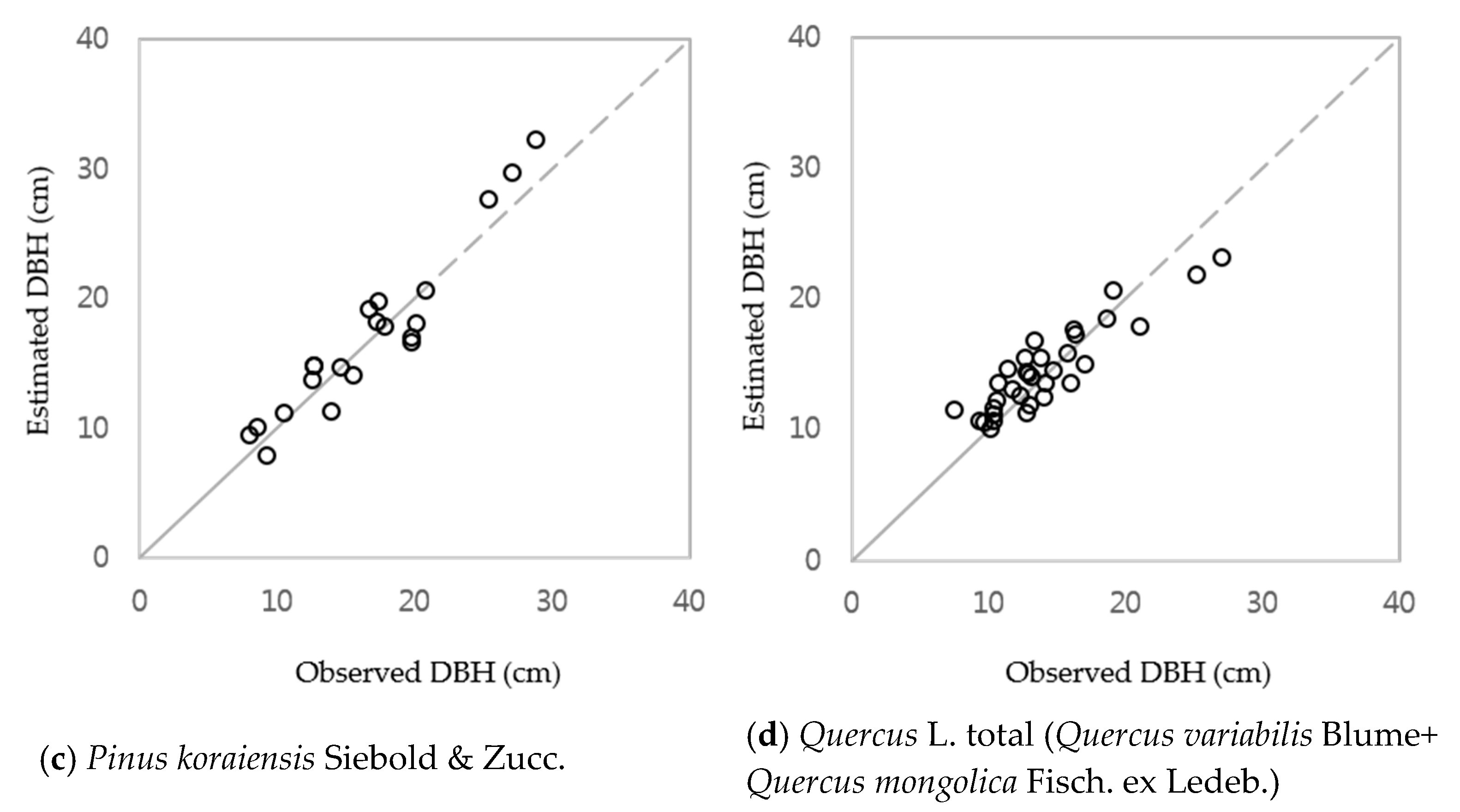
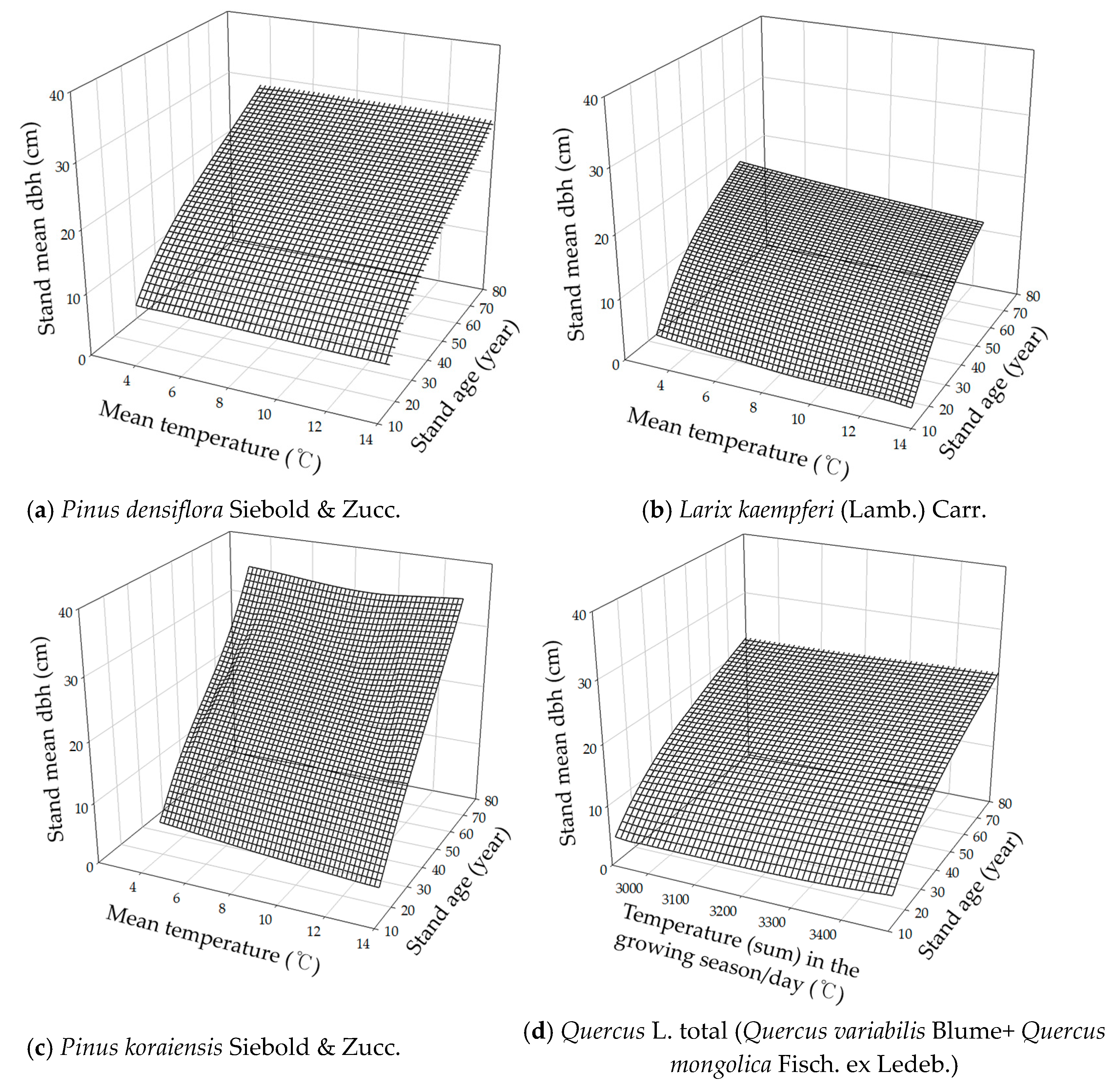
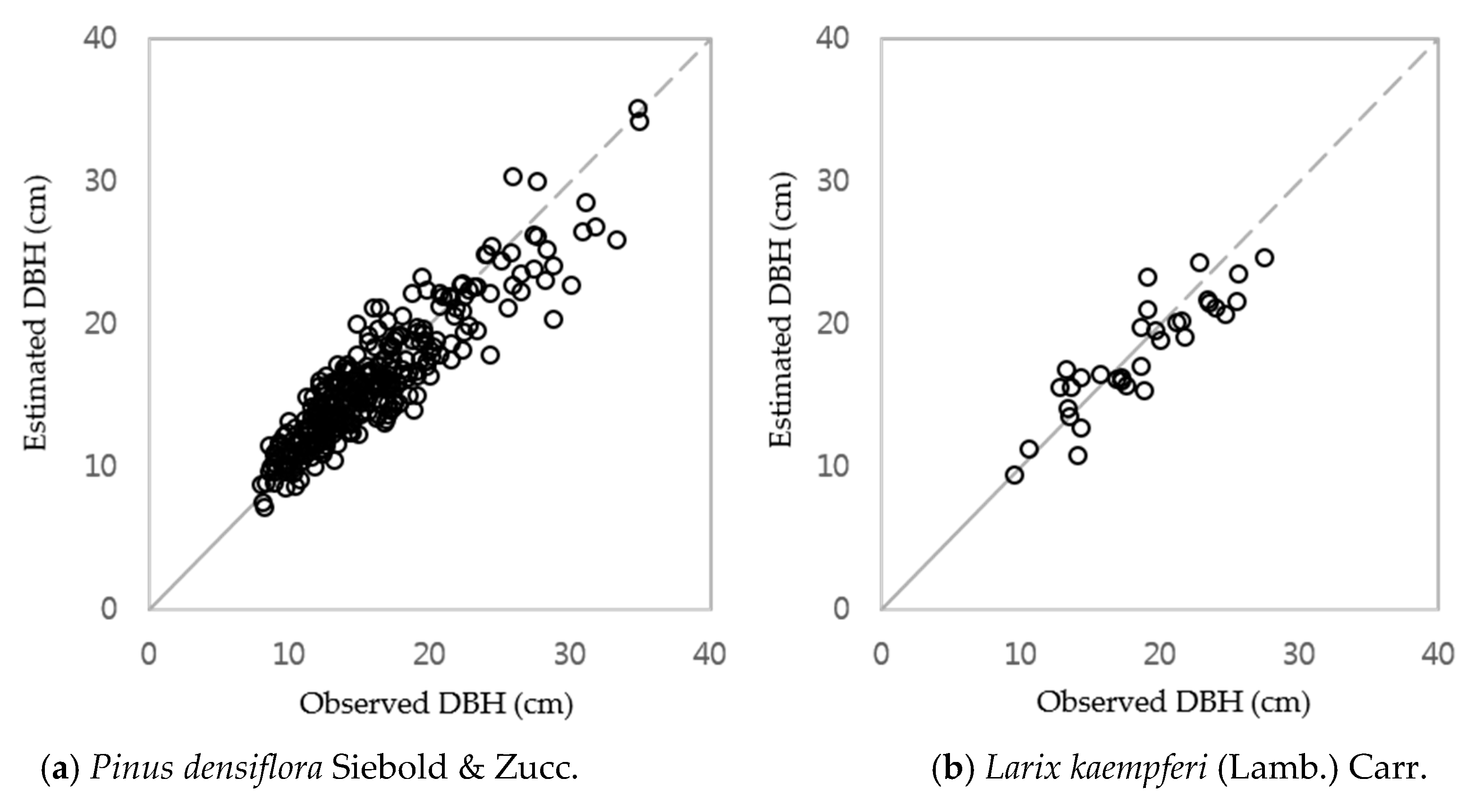
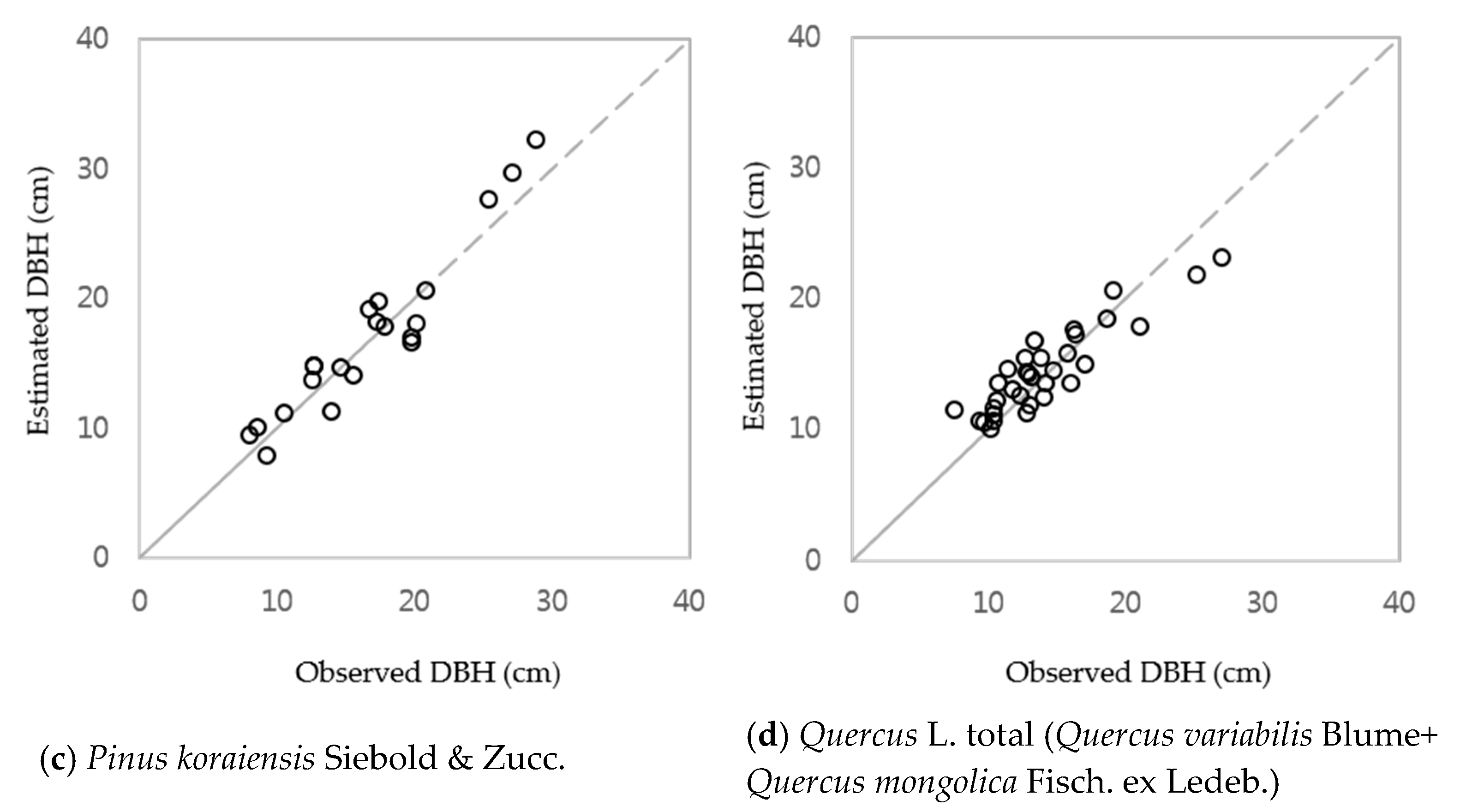
3.1. DBH Estimation with Stand Age, SI and Nha
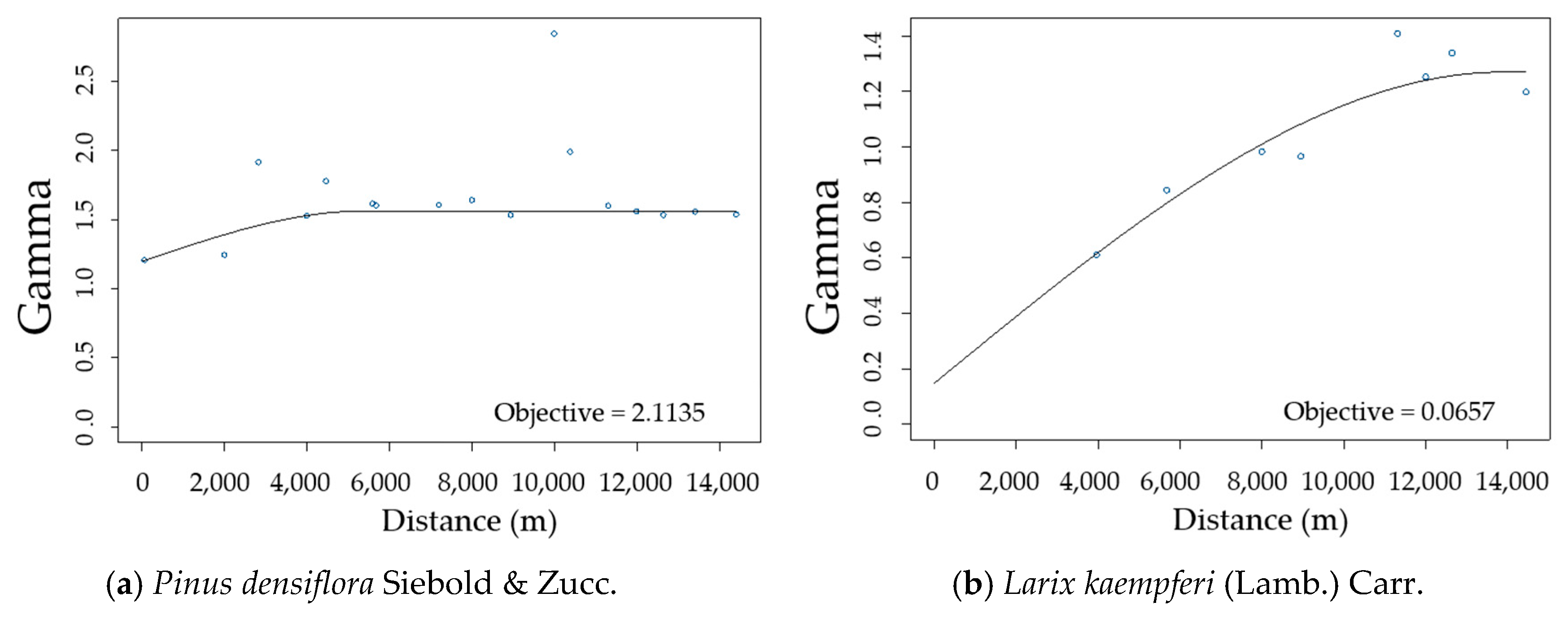
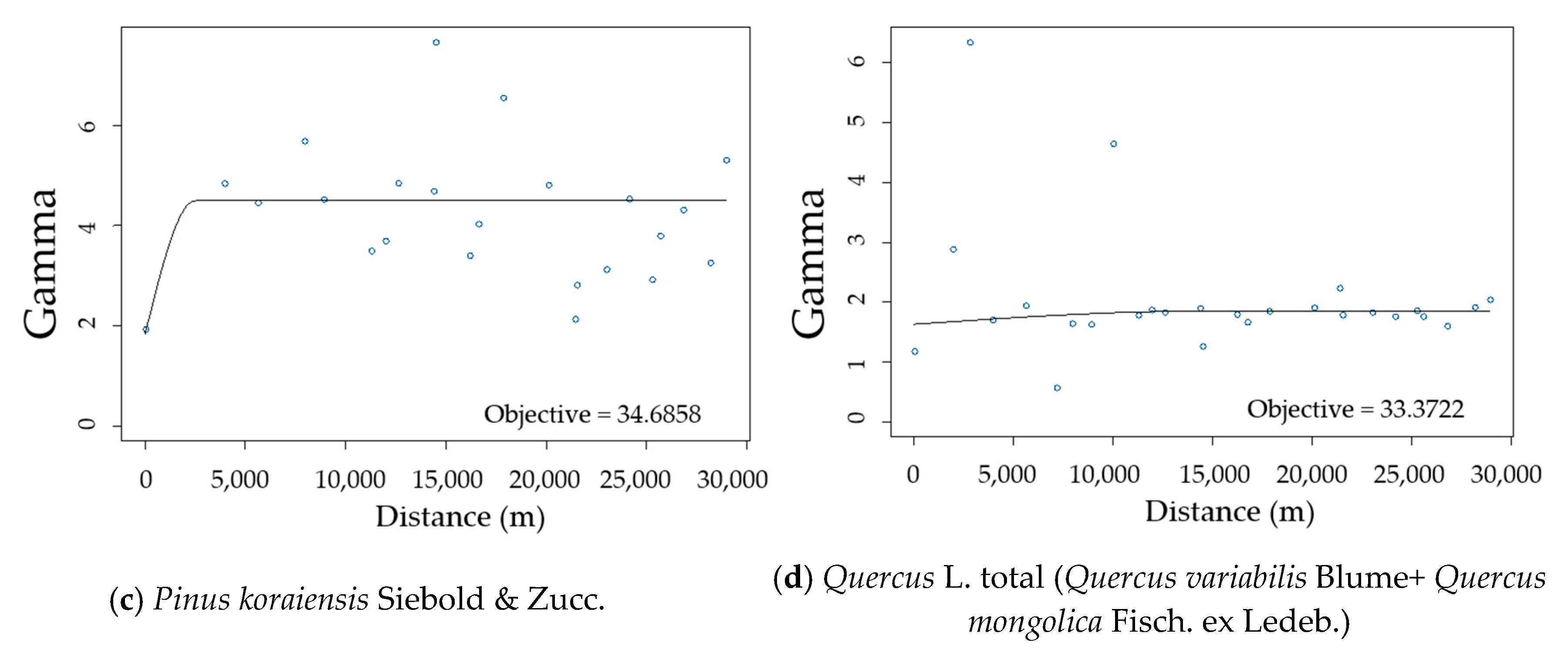
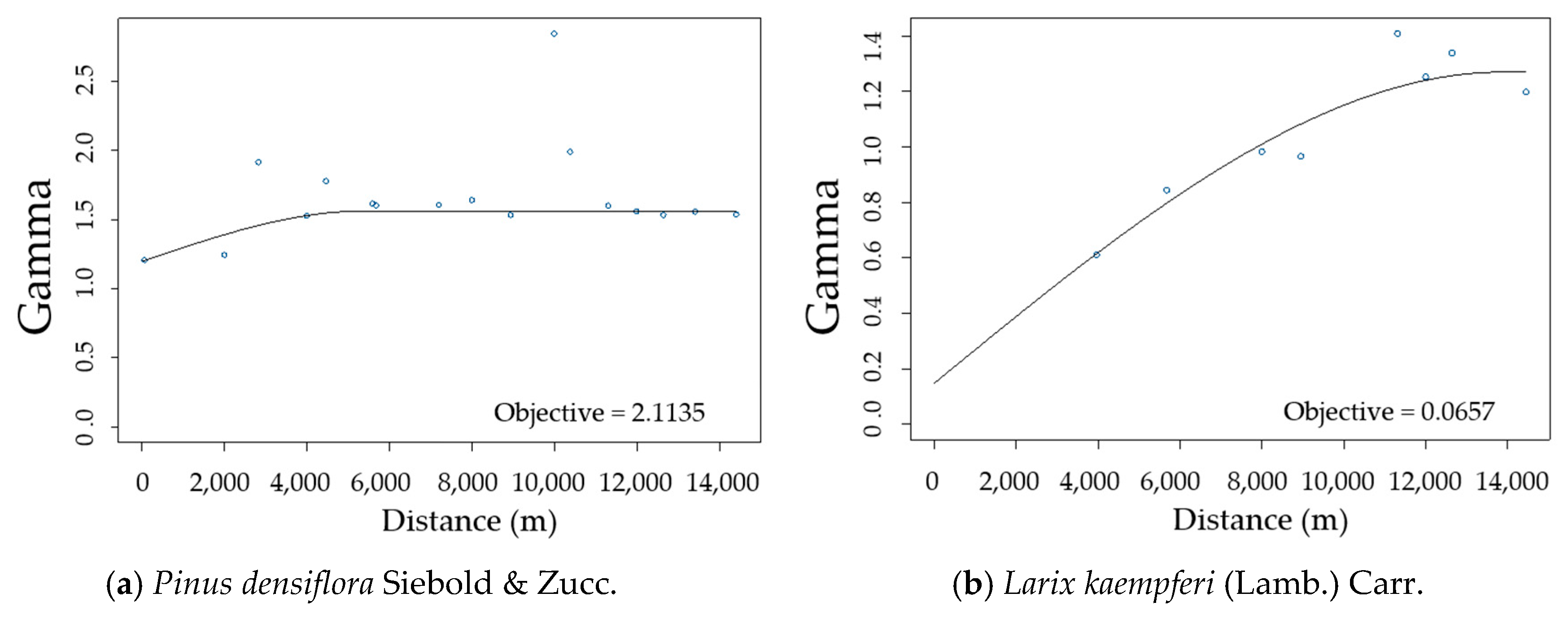
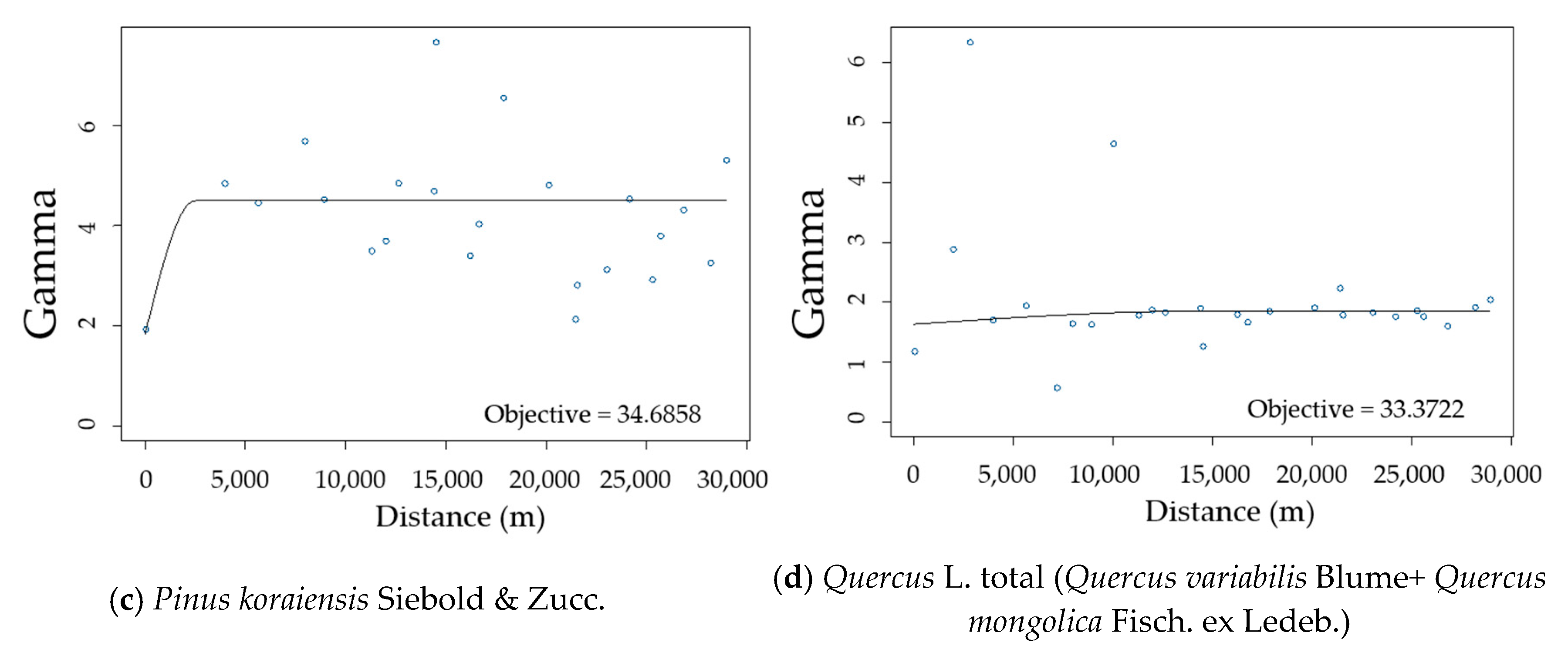
3.2. Spatial Autocorrelation of Residuals
3.3. Residual Model with Climate Factors
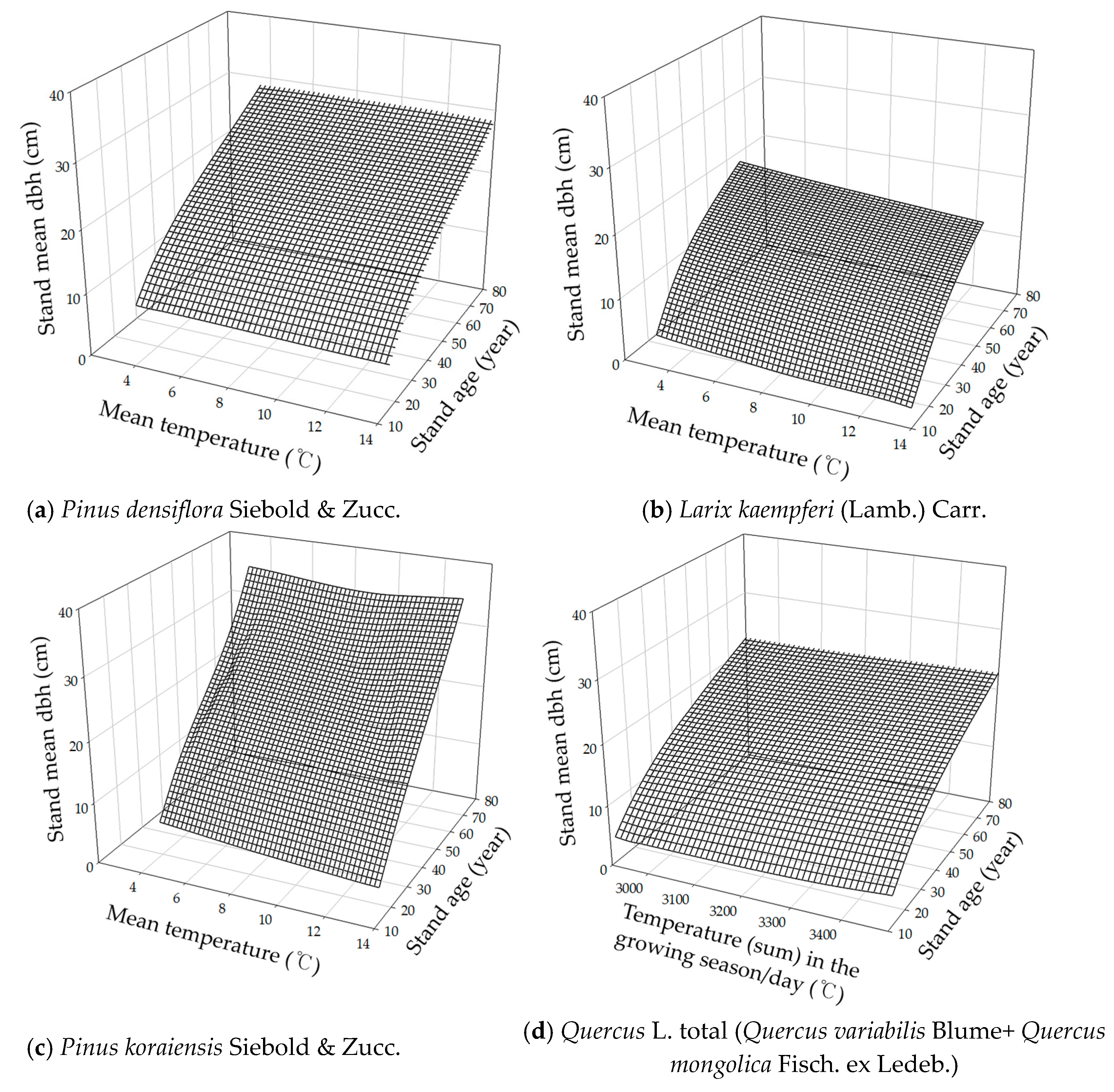
3.4. Integrated DBH Estimation Model with Stand and Climate Variables
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age (Year) | Number of Trees per ha (Nha) | |||
|---|---|---|---|---|
| Pinus densiflora (Site Index: 12) | Larix kaempferi (Site Index: 18) | Pinus koraiensis (Site Index: 14) | Quercus Total (Site Index: 12) | |
| 20 | 1827 | 1316 | 1362 | 2024 |
| 25 | 1681 | 993 | 994 | 1657 |
| 30 | 1418 | 779 | 868 | 1373 |
| 35 | 1186 | 658 | 733 | 1153 |
| 40 | 1006 | 584 | 644 | 981 |
| 45 | 961 | 538 | 583 | 936 |
| 50 | 859 | 508 | 541 | 828 |
| 55 | 781 | 490 | 511 | 741 |
| 60 | 720 | 478 | 489 | 669 |
| 65 | 673 | 471 | 473 | 609 |
| 70 | 636 | 468 | 462 | 560 |
| 75 | 607 | 467 | 454 | 518 |
| 80 | 583 | 467 | 448 | 481 |
References
- Clutter, J.L. Compatible growth and yield models for loblolly pine. For. Sci. 1963, 9, 354–371. [Google Scholar]
- Ritchie, M.W.; Hann, D.W. Development of a tree height growth model for Douglas-fir. For. Ecol. Manag. 1986, 15, 135–145. [Google Scholar] [CrossRef]
- McCullagh, A.; Black, K.; Nieuwenhuis, M. Evaluation of tree and stand-level growth models using national forest inventory data. Eur. J. For. Res. 2017, 136, 251–258. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings Basics and Applications of Dendrochronology; Kluwer Academic Publishing: Dordrecht, The Netherland, 1988; p. 276. [Google Scholar]
- Lee, W.K. A dynamic regional forest management model for the sustainability of forest practice-Considering forest growth and economical conditions. Korean J. For. Econ. 1995, 3, 71–98. [Google Scholar]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Garcia-Gonzalo, J.; Seidel, R.; Delzon, S.; Corona, P.; Kolström, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Pothier, D. Twenty-year results of precommercial thinning in a balsam fir stand. For. Ecol. Manag. 2002, 168, 177–186. [Google Scholar] [CrossRef]
- Nishizono, T. Effects of thinning level and site productivity on age-related changes in stand volume growth can be explained by a single rescaled growth curve. For. Ecol. Manag. 2010, 259, 2276–2291. [Google Scholar] [CrossRef]
- Ji, B.; Tao, J.; Zhang, G.; Xu, D.; Wang, W.; Wu, W. Construction of Nonlinear mixed Model of Stand Growth Rate. J. Southwest For. Univ. 2017, 37, 149–158. [Google Scholar]
- Takahashi, K.; Azuma, H.; Yasue, K. Effects of climate on the radial growth of tree species in the upper and lower distribution limits of an altitudinal ecotone on Mount Norikura, central Japan. Ecol. Res. 2003, 18, 549–558. [Google Scholar] [CrossRef]
- Sørensen, R.; Zinko, U.; Seibert, J. On the calculation of the topographic wetness index: Evaluation of different methods based on field observations. Hydrol. Earth Syst. Sci. Discuss. 2006, 10, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Byun, J.G.; Lee, W.K.; Kim, M.; Kwak, D.A.; Kwak, H.; Park, T.; Byun, W.H.; Son, Y.; Choi, J.K.; Lee, Y.J.; et al. Radial growth response of Pinus densiflora and Quercus spp. to topographic and climatic factors in South Korea. J. Plant Ecol. 2013, 6, 380–392. [Google Scholar] [CrossRef]
- Tian, H.; Melillo, J.M.; Kicklighter, D.W.; McGuire, A.D.; Helfrich, I.J.; Moore Iii, B.; Vörösmarty, C.J. Climatic and biotic controls on annual carbon storage in Amazonian ecosystems. Glob. Ecol. Biogeogr. 2001, 9, 315–335. [Google Scholar] [CrossRef]
- Enquist, B.J.; West, G.B.; Charnov, E.L.; Brown, J.H. Allometric scaling of production and life-history variation in vascular plants. Nature 1999, 401, 907–911. [Google Scholar] [CrossRef]
- Enquist, B.J. Universal scaling in tree and vascular plant allometry: Toward a general quantitative theory linking plant form and function from cells to ecosystem. Tree Physiol. 2002, 22, 1045–1064. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Lee, W.K.; Park, T.; Kwak, H.; Byun, J.; Nam, K.; Lee, K.H.; Son, Y.M.; Won, H.K.; Lee, S.M. Developing dynamic DBH growth prediction model by thinning intensity and cycle-based on yield table data. J. Korean For. Soc. 2012, 101, 266–278. [Google Scholar]
- Moore, I.D.; Gessler, P.E.; Nielsen, G.A.; Peterson, G.A. Soil attribute prediction using terrain analysis. Soil Sci. Soc. Am. J. 1993, 57, 443–452. [Google Scholar] [CrossRef]
- Zirlewagen, D.; Raben, G.; Weise, M. Zoning of forest health conditions based on a set of soil, topographic and vegetation parameters. For. Ecol. Manag. 2007, 248, 43–55. [Google Scholar] [CrossRef]
- Nirmal, S.; Mahadev, S. Climate-diameter growth relationships of black spruce and jack pine trees in boreal Ontario, Canada. Glob. Chang. Biol. 2013, 19, 505–516. [Google Scholar]
- Gao, L.; Wang, X.; Zhao, X. Growth response of two coexisting species to climate change in broadleaved Korean pine forests in Changbai Mountain, Northeastern China. J. Beijing For. Univ. 2013, 35, 24–31. [Google Scholar]
- Ryan, M.G.; Binkley, D.; Fownes, J.H. Age-related decline in forest productivity: Pattern and process. Adv. Ecol. Res. 2008, 27, 213–262. [Google Scholar]
- Choi, K.; Kim, M.; Lee, W.K.; Gang, H.U.; Chung, D.J.; Ko, E.J.; Yun, B.H.; Kim, C.H. Estimating radial growth response of major tree species using climatic and topographic condition in South Korea. Korea J. Clim. Chang. Res. 2014, 5, 127–137. [Google Scholar] [CrossRef]
- Anselin, L.; Griffith, D.A. Do spatial effects really matter in regression analysis? Pap. Reg. Sci. 1988, 65, 11–34. [Google Scholar] [CrossRef]
- Overmars, K.D.; de Koning, G.H.; Veldkamp, A. Spatial autocorrelation in multi-scale land use models. Ecol. Model. 2003, 164, 257–270. [Google Scholar] [CrossRef]
- Korea Forest Service. Research and Application Based on National Forest Resources Survey; Korea Forest Service: Daejeon, Korea, 2015; p. 130. (In Korean)
- Korea Forest Service. The 5th National Forest Inventory; Korea Forest Service: Daejeon, Korea, 2013; p. 142. (In Korean)
- Korea Forest Research Institute. The 5th National Forest Inventory Report; Korea Forest Research Institute: Seoul, Korea, 2011; p. 166. (In Korean)
- Lee, S.C.; Choi, S.H.; Lee, W.K.; Yoo, S.J.; Byun, J.G. The effect of climate data applying temperature lapse rate on prediction of potential forest distribution. Korean Soc. Geospat. Inf. Sci. 2011, 19, 19–27. [Google Scholar]
- Jeong, H.M.; Kim, H.R.; You, Y.H. Growth difference among saplings of Quercus acutissima, Q. variabilis and Q. mongolica under the environmental gradients treatment. Korean J. Environ. Biol. 2009, 27, 82–87. [Google Scholar]
- Reineke, L.H. Perfecting a Stand-Density Index for Even-Aged Forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Sterba, H. Estimating potential density from thinning experiments and inventory data. For. Sci. 1987, 33, 1022–1034. [Google Scholar]
- Lee, W.K.; Seo, J.H.; Bae, S.W. Maximum stem number and mortality model for even-aged Pinus densiflora stand in Kangwon-Province, Korea. J. Korean For. Soc. 2000, 89, 634–644. [Google Scholar]
- Kim, M.; Lee, W.K.; Choi, G.M.; Song, C.; Lim, C.H.; Moon, J.; Piao, D.; Kraxner, F.; Shividenko, A.; Forsell, N. Modeling stand-level mortality based on maximum stem number and seasonal temperature. For. Ecol. Manag. 2017, 386, 37–50. [Google Scholar] [CrossRef]
- Dobbertin, M. Tree growth as indicator of tree vitality and of tree reaction to environmental stress: A review. Eur. J. For. Res. 2005, 124, 319–333. [Google Scholar] [CrossRef]
- Moore, J.A.; Zhang, L.; Stuck, D. Height-diameter equations for ten tree species in the Inland Northwest. West. J. Appl. For. 1996, 11, 132–137. [Google Scholar]
- Martínez Pastur, G.J.; Cellini, J.M.; Lencinas, M.V.; Peri, P.L. Stand growth model using volume increment/basal. J. For. Sci. 2008, 54, 102–108. [Google Scholar] [CrossRef]
- Li, C.M. Simulating basal area growth of fir plantations using a nonlinear mixed modeling approach. J. Beijing For. Univ. 2009, 31, 44–49. [Google Scholar]
- Lim, C.H.; Moon, J.Y.; Lim, Y.J.; Kim, S.J.; Lee, W.K. Estimating spatio-temporal distribution of climate factors in Andong Dam basin. J. Korean Soc. Geospat. Inf. Syst. 2015, 23, 57–65. [Google Scholar] [CrossRef]
- Kirilenko, A.P.; Solomon, A.M. Modeling dynamic vegetation response to rapid climate change using bioclimatic classification. Clim. Chang. 1998, 38, 15–49. [Google Scholar] [CrossRef]
- Liu, Q.; Xia, J.; Xie, W. Application of semi-variogram and Moran’s I to spatial distribution of trace elements in soil: A case study in Shouguang County. Geomat. Inf. Sci. Wuhan Univ. 2011, 36, 1129–1133. [Google Scholar]
- Byun, J.G.; Lee, W.K.; Nor, D.K.; Kim, S.H.; Choi, J.K.; Lee, Y.J. The relationship between tree radial growth and topographic and climatic factors in red pine and oak in central regions of Korea. J. Korean For. Soc. 2010, 99, 908–913. [Google Scholar]
- Cameron, A.C.; Windmeijer, F.A.G. An R-squared measure of goodness of fit for some common nonlinear regression models. J. Econ. 1997, 77, 329–342. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodal Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Kim, T.M.; Lee, W.K.; Jung, S.E.; Kwak, H.B. Analysis of relationship between spatial distribution of Pinus densiflora and topographical factors. J. Korea Spat. Inf. Soc. 2008, 16, 359–372. [Google Scholar]
- Nam, K.; Lee, W.K.; Kim, M.; Kwak, D.A.; Byun, W.H.; Yu, H.; Kwak, H.; Kwon, T.; Sung, J.; Chung, D.J.; et al. Spatio-temporal change in forest cover and carbon storage considering actual and potential forest cover in South Korea. Sci. China Life Sci. 2015, 58, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Lee, W.K.; Kwak, D.A.; Lee, S.; Son, Y.; Lim, J.H.; Saborowski, J. Predicting forest cover changes in future climate using hydrological and thermal indices in South Korea. Clim. Res. 2011, 49, 229–245. [Google Scholar] [CrossRef]
- Korea Forest Service. Table of Tree Volume/Mass and Yield Table; Korea Forest Service: Daejeon, Korea, 2009; p. 261. (In Korean)
- Jang, D.H.; Lee, S. Analysis of forest type changes for vulnerability assessment of climate changes in the mountainous regions of Gangwon-do, Korea. Korea J. Clim. Res. 2013, 8, 169–183. [Google Scholar] [CrossRef]
- Korea Forest Service: Table of Tree Volume/Mass and Yield Table; Korea Forest Service: Daejeon, Korea, 2009. (In Korean)
- Innes, J.L. Forest Health: Its Assessment and Status; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Shigo, A.L. Die neue Baumbiologie; Bernhard Thalacker Verlag: Braunschweig, Germany, 1990. [Google Scholar]
- Gehrig, M. Methoden zur Vitalitätsbeurteilung von Bäumen; Swiss Federal Institute of Technology: Zürich, Germany, 2004; p. 254. [Google Scholar]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Santiago-García, W.; Pérez-López, E.; Quiñonez-Barraza, G.; Rodríguez-Ortiz, G.; Santiago-García, E.; Ruiz-Aquino, F.; Tamarit-Urias, J.C. A dynamic system of growth and yield equations for Pinus patula. Forests 2017, 8, 465. [Google Scholar] [CrossRef]

| Variables | Use | Pinus densiflora Siebold & Zucc. | Larix kaempferi (Lamb.) Carr. | Pinus koraiensis Siebold & Zucc. | Quercus Total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | S.D. | Mean | S.D. | Mean | S.D. | Mean | S.D. | ||
| No. of plots | Model | 2796 | 301 | 185 | 1481 | ||||
| Verify | 311 | 33 | 20 | 165 | |||||
| Stand age (year)* | Model | 33.6 | 8.4 | 33.9 | 7.9 | 28.8 | 8.5 | 37.1 | 5.7 |
| Verify | 33.8 | 8.4 | 31.5 | 7.1 | 35.6 | 9.7 | 38.1 | 7.0 | |
| Stand mean DBH (cm) | Model | 15.5 | 4.5 | 17.9 | 4.3 | 17.8 | 6.4 | 15.1 | 4.1 |
| Verify | 15.8 | 4.5 | 18.5 | 4.6 | 24.3 | 7.5 | 15.0 | 4.0 | |
| Site index* | Model | 12.1 | 2.9 | 17.2 | 3.2 | 14.1 | 2.8 | 12.6 | 1.2 |
| Verify | 12.1 | 3.1 | 16.8 | 3.0 | 15.1 | 2.5 | 12.3 | 1.7 | |
| Tree height (m) | Model | 10.45 | 2.75 | 15.38 | 3.85 | 11.80 | 3.81 | 11.19 | 11.47 |
| Verify | 10.20 | 2.72 | 15.01 | 3.82 | 9.71 | 3.20 | 10.50 | 2.15 | |
| Stand density (n/ha) | Model | 1467.0 | 729.1 | 875.9 | 383.4 | 952.5 | 562.2 | 1410.4 | 284.4 |
| Verify | 1463.9 | 724.1 | 890.2 | 126.9 | 403.6 | 537.8 | 1359.7 | 223.6 | |
| Mean temperature (°C) | Model | 11.0 | 2.1 | 8.8 | 2.0 | 9.5 | 1.9 | 8.9 | 1.1 |
| Verify | 10.9 | 0.7 | 8.8 | 0.5 | 1.6 | 1.8 | 8.4 | 1.0 | |
| Temperature (sum) in the growing season* (°C) | Model | 3323.3 | 84.3 | 3282.7 | 76.5 | 3243.9 | 80.1 | 3281.1 | 43.0 |
| Verify | 3324.4 | 80.0 | 3299.8 | 77.5 | 3296.9 | 79.9 | 3265.4 | 42.5 | |
| Precipitation in growing season* (mm) | Model | 980.5 | 184.1 | 921.2 | 59.5 | 918.7 | 50.3 | 941.0 | 84.7 |
| Verify | 981.9 | 154.3 | 926.8 | 47.1 | 47.1 | 43.1 | 951.8 | 92.1 | |
| Tree Species | Equation (1) | Equation (2) | Equation (3) | Optimal Equation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| R2 | AIC | RMSE | R2 | AIC | RMSE | R2 | AIC | RMSE | ||
| Pinus densiflora Siebold & Zucc. | 0.826 | 1826.4 | 2.241 | 0.748 | 2096.0 | 2.798 | 0.835 | 1766.5 | 2.134 | Equation (3) |
| Larix kaempferi (Lamb.) Carr. | 0.787 | 238.5 | 2.446 | 0.741 | 244.0 | 2.497 | 0.790 | 236.2 | 2.424 | Equation (3) |
| Pinus koraiensis Siebold & Zucc. | 0.886 | 179.7 | 2.971 | 0.867 | 192.7 | 3.223 | 0.904 | 178.8 | 2.956 | Equation (3) |
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | 0.574 | 1119.6 | 2.379 | 0.563 | 1188.4 | 2.510 | 0.814 | 1104.1 | 2.351 | Equation (3) |
| Tree Species | Parameter | Estimate | Std. Error | t-Value | p-Value |
|---|---|---|---|---|---|
| Pinus densiflora Siebold & Zucc. | a | 2.437 | 0.162 | 15.171 | <0.001 |
| b | 0.613 | 0.010 | 60.118 | <0.001 | |
| c | 0.491 | 0.011 | 45.491 | <0.001 | |
| d | −0.213 | 0.005 | −47.333 | <0.001 | |
| Larix kaempferi (Lamb.) Carr. | a | 1.982 | 0.387 | 5.116 | <0.001 |
| b | 0.670 | 0.036 | 18.862 | <0.001 | |
| c | 0.370 | 0.039 | 9.386 | <0.001 | |
| d | −0.172 | 0.014 | −12.134 | <0.001 | |
| Pinus koraiensis Siebold & Zucc. | a | 1.563 | 0.425 | 3.681 | <0.001 |
| b | 0.781 | 0.046 | 17.169 | <0.001 | |
| c | 0.358 | 0.066 | 5.454 | <0.001 | |
| d | −0.172 | 0.019 | −9.182 | <0.001 | |
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | a | 16.564 | 1.560 | 10.61578 | <0.001 |
| b | 0.488 | 0.014 | 35.36957 | <0.001 | |
| c | 0.060 | 0.019 | 3.210811 | <0.001 | |
| d | −0.281 | 0.008 | −36.4036 | <0.001 |
| Tree Species | Parameter | Estimate | Std. Error | t-Value | p-Value |
|---|---|---|---|---|---|
| Pinus densiflora Siebold & Zucc. | a | 43.896 | 1.906 | 23.033 | <0.001 |
| b | −22.959 | 0.381 | −60.259 | <0.001 | |
| c | 0.485 | 0.011 | 45.745 | <0.001 | |
| d | −0.216 | 0.004 | −49.269 | <0.001 | |
| Larix kaempferi Carr. | a | 37.753 | 5.423 | 6.962 | <0.001 |
| b | −18.456 | 1.059 | −17.434 | <0.001 | |
| c | 0.369 | 0.040 | 9.295 | <0.001 | |
| d | −0.177 | 0.014 | −12.385 | <0.001 | |
| Pinus koraiensis | a | 56.486 | 11.450 | 4.933 | <0.001 |
| b | −24.406 | 1.513 | −16.130 | <0.001 | |
| c | 0.314 | 0.066 | 4.793 | <0.001 | |
| d | −0.167 | 0.019 | −8.941 | <0.001 | |
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | a | 177.200 | 12.552 | 14.117 | <0.001 |
| b | −18.896 | 0.545 | −34.672 | <0.001 | |
| c | 0.053 | 0.014 | 3.804 | <0.001 | |
| d | −0.290 | 0.008 | −38.145 | <0.001 |
| Tree Species | Parameter | Estimate | Std. Error | t-Value | p-Value |
|---|---|---|---|---|---|
| Pinus densiflora Siebold & Zucc. | a | 0.69769 | 0.25136 | 2.78 | 0.0055 |
| b | −0.03962 | 0.01804 | −2.2 | 0.0282 | |
| c | −0.00038 | 0.000204 | −1.88 | 0.0608 | |
| Larix kaempferi (Lamb.) Carr. | a | −1.36713 | 1.90875 | −0.72 | 0.4746 |
| b | −0.26735 | 0.06112 | −4.37 | <0.0001 | |
| c | 0.00516 | 0.00196 | 2.64 | 0.0089 | |
| Pinus koraiensis Siebold & Zucc. | a | 13.39043 | 3.47305 | 3.86 | 0.002 |
| b | −0.25759 | 0.09842 | −2.62 | 0.01 | |
| c | −0.0072 | 0.0035 | −2.08 | 0.0386 | |
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | a | −0.48854 | 0.38888 | −1.26 | 0.2092 |
| b | 0.09703 | 0.02479 | 3.91 | <0.0001 | |
| c | −0.00065 | 0.00035 | −1.85 | 0.0652 |
| Tree Species | Parameter | Estimate | Std. Error | t-Value | p-Value |
|---|---|---|---|---|---|
| Pinus densiflora Siebold & Zucc. | a | −0.67988 | 1.46426 | −0.46 | 0.6425 |
| b | 0.000323 | 0.000451 | 0.72 | 0.4738 | |
| c | −0.00052 | 0.000205 | −2.52 | 0.0119 | |
| Larix kaempferi (Lamb.) Carr. | a | 0.63004 | 6.04169 | 0.1 | 0.917 |
| b | −0.00146 | 0.00168 | −0.87 | 0.3878 | |
| c | 0.00568 | 0.00205 | 2.78 | 0.006 | |
| Pinus koraiensis Siebold & Zucc. | a | 21.27081 | 8.31303 | 2.56 | 0.0117 |
| b | −0.00296 | 0.00241 | −1.23 | 0.2204 | |
| c | −0.01268 | 0.00383 | −3.31 | 0.0012 | |
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | a | −7.16166 | 2.62723 | −2.73 | 0.0065 |
| b | 0.00232 | 0.000814 | 2.85 | 0.0044 | |
| c | −0.00076 | 0.000358 | −2.13 | 0.0334 |
| Tree Species | Integrated DBH Estimation Model | |
|---|---|---|
| Non-Spatial Variable | Spatial Variable | |
| Pinus densiflora Siebold & Zucc. | ||
| Larix kaempferi (Lamb.) Carr. | ||
| Pinus koraiensis Siebold & Zucc. | ||
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | ||
| Tree Species | DBH Estimation Model R2 | Integrated DBH Estimation Model R2 |
|---|---|---|
| Pinus densiflora Siebold & Zucc. | 0.835 | 0.836 |
| Larix kaempferi (Lamb.) Carr. | 0.790 | 0.821 |
| Pinus koraiensis Siebold & Zucc. | 0.904 | 0.906 |
| Quercus L. total (Quercus variabilis Blume+ Quercus mongolica Fisch. ex Ledeb.) | 0.6395 | 0.6927 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piao, D.; Kim, M.; Choi, G.-M.; Moon, J.; Yu, H.; Lee, W.-K.; Wang, S.W.; Jeon, S.W.; Son, Y.; Son, Y.-M.; et al. Development of an Integrated DBH Estimation Model Based on Stand and Climatic Conditions. Forests 2018, 9, 155. https://doi.org/10.3390/f9030155
Piao D, Kim M, Choi G-M, Moon J, Yu H, Lee W-K, Wang SW, Jeon SW, Son Y, Son Y-M, et al. Development of an Integrated DBH Estimation Model Based on Stand and Climatic Conditions. Forests. 2018; 9(3):155. https://doi.org/10.3390/f9030155
Chicago/Turabian StylePiao, Dongfan, Moonil Kim, Go-Mee Choi, Jooyeon Moon, Hangnan Yu, Woo-Kyun Lee, Sonam Wangyel Wang, Seong Woo Jeon, Yowhan Son, Yeong-Mo Son, and et al. 2018. "Development of an Integrated DBH Estimation Model Based on Stand and Climatic Conditions" Forests 9, no. 3: 155. https://doi.org/10.3390/f9030155