Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers?


1
Fiducie de Recherche sur la Forêt des Cantons-de-l’Est/Eastern Townships Forest Research Trust, 1 rue Principale, Saint-Benoît-du-Lac, QC J0B 2M0, Canada
2
Department of Biology, University of Regina, 3737 Wascana Parkway, Regina, SK S4S 0A2, Canada
*
Author to whom correspondence should be addressed.
Forests 2018, 9(5), 258; https://doi.org/10.3390/f9050258
Submission received: 10 April 2018
/
Revised: 1 May 2018
/
Accepted: 7 May 2018
/
Published: 10 May 2018
(This article belongs to the Special Issue Hardwood Reforestation and Restoration)
Abstract
:This study was conducted in a riparian buffer bordering a 1 km segment of a headwater stream crossing a pasture site located in southern Québec (Canada). Three species were planted (black walnut (Juglans nigra L.), bur oak (Quercus macrocarpa Michx.), and eastern white pine (Pinus strobus L.)) with three vegetation treatments (control, herbicide (one application/year for 3 years), and black plastic mulch)). The main objective was to determine to which extent herbicide and plastic mulch, used with species having different ecological characteristics, affect tree growth and soil nutrient status in riparian buffers. Survival was high (>93%) for all species in all treatments. In the control (no vegetation treatment), growth was similar among species. Black walnut had the strongest growth response to herbicide and plastic mulch, and white pine had the weakest. For all species, growth was similar in the herbicide and the plastic mulch treatments. During the fifth growing season, plastic mulch increased soil nitrate and phosphorus compared to the herbicide treatment. In the plastic mulch treatment, higher soil nitrate supply was observed for species that preferentially uptake ammonium (black walnut and white pine). Soil nutrient supplies were similar between the control and herbicide treatments. Despite the more favorable nutritional conditions it provides, permanent black plastic mulching does not provide higher growth benefits after 5 years than a 3-year herbicide treatment. The high soil nitrate supply observed in mulched black walnut and mulched white pine may indicate a limited capacity for nitrate phytoremediation by these species.
1. Introduction
During the last decades, many countries have developed policies and programs to stimulate the protection and the establishment of forest riparian buffers on farmland [1,2]. Such buffers can reduce the load of many aquatic pollutants (nitrate, phosphorus, pesticides, sediments, pathogens), while providing stream shading, streambank stabilisation, and flood protection [3,4,5]. Tree riparian buffers are also keystone landscape components for terrestrial, amphibian, and aquatic biodiversity because they provide habitat, shelter, food sources, and movement corridors for many species within agroecosystems [6,7,8,9].
Ecosystem services provide by riparian buffers can be improved by planting high-value hardwood species that produce edible nuts or acorns [10,11]. Many of these species have high light, water, and nutrient requirements to achieve optimal growth [12]. Hence, the large amount of deforested riparian land available along pastures and crop fields [13,14] provides tremendous opportunity to reintroduce nut producing hardwoods. The inclusion of conifers in riparian buffers can also improve their multi-functionality [15], as conifers provide a year-round shelter for bird and mammal species [16].
Although the selection of tree species has important implications for the provision of ecosystem services in riparian buffers, little information is available on the establishment success of different species in farmland riparian habitats. Most often, one or more sylvicultural treatments are needed to improve tree survival and growth in riparian zones [11,17,18]. Vegetation management (weed control) is especially needed when the site is dominated by a dense cover of grasses, which compete strongly with trees for water and nutrients [19]. On sites dominated by a tall herbaceous vegetation cover (i.e., forbs) or shrubs, vegetation management can also be used to increase light availability [20]. Among vegetation management strategies, herbicide application and black plastic (polyethylene) mulching have been widely used in planted buffers [10,15,21,22]. Herbicide and black plastic mulch are both known to enhance angiosperm and gymnosperm growth by locally creating a weed-free plantation environment where soil temperature, humidity, nitrogen (N) mineralisation, and nitrate (NO3) availability are increased [23,24,25,26,27]. Because of its absorptive and transmittance properties, black plastic mulch also increases air temperature locally [28], which may positively affect photosynthetic capacity in seedlings of temperate species [29].
Despite the benefits they provide, it is unclear which of plastic mulching or herbicide is the best treatment to enhance tree growth and survival in riparian buffers and other types of agroforestry systems [30]. Sometimes, growth response to one or the other treatment differed between species, as seen in abandoned farmland and riparian afforestation trials [11,23,31,32]. Change in the magnitude of the growth response to a given vegetation management treatment is also expected when species with contrasting ecologies are compared [11]. For example, white pine (Pinus strobus L.), which has low nutrient requirements [16], was less responsive to plastic mulch than deciduous species [15]. Species-specific feedbacks on riparian soil NO3 can also occur depending on the species used in combination with black plastic mulch [15]. A much higher NO3 supply was found in the soil underneath mulched white pine and red oak (Quercus rubra L.) [15], which are known to preferentially uptake soil ammonium (NH4) [33,34,35,36]. Such species feedback on soil NO3 observed in mulched trees requires further investigation because many riparian buffers are planted to reduce NO3 pollution, which greatly accelerates stream eutrophication [3,37].
An important distinction between herbicide and black plastic mulch is related to the duration of their effects on the growth environment. Once an herbicide treatment is stopped, the regrowth of herbaceous vegetation is generally spontaneous and rapid [20]. In riparian buffers, such vegetation cover has important functions for NO3 pollution mitigation because ruderal herbaceous species naturally growing in riparian buffers are often nitrophilous and their capacity to immobilize N in plant biomass is relatively high (up to 150 kg N/ha) during the summer [35,38,39,40]. Conversely, plastic mulch generally remains on plantation sites for decades because of the lack of biodegradability of the petroleum-based commercial mulches [41]. Thus, the effects of plastic mulch on the growth environment and on vegetation cover are longer-lasting compared to an herbicide treatment that would be stopped once trees were properly established. Consequently, the use of a vegetation treatment that allows the regrowth of herbaceous vegetation underneath trees (e.g., herbicide) is expected to have a long-term effect on reducing soil NO3 supply, thus reducing its potential leaching into streams.
This farm-scale study was conducted along a 1 km segment of a headwater stream crossing a high fertility pasture site located in southern Québec (southeastern Canada). Two nut producing hardwoods (black walnut (Juglans nigra L.) and bur oak (Quercus macrocarpa Michx.)) and eastern white pine were planted in an experimental design with three vegetation treatments (a control treatment, an herbicide treatment (one application per year for 3 years), and a permanent black plastic mulch treatment). The first objective of this study was to evaluate which vegetation treatment is the best to enhance the survival and growth of the three species in a riparian buffer. The second objective was to determine to which extent herbicide and plastic mulch used in combination with species having different nutritional requirements and ecological characteristics affect nutrient status in riparian soil once trees are well-established (fifth growing season).
In this study, four hypotheses are tested: (1) because of its longer-lasting effect on the growing environment, black plastic mulch will be more effective than the herbicide to improve soil NO3 status and the growth of all species; (2) because of its low nutrient requirements, white pine will be less responsive to both vegetation management treatments than black walnut and bur oak; (3) in the plastic mulch treatment, the highest NO3 supply rate will be observed for white pine and black walnut, which preferentially uptake NH4 [33,42]; (4) similar NO3 supply rates will be observed at 5 years in the control and herbicide treatments.
2. Materials and Methods
2.1. Site Description
The study took place at the Carocel farm (45°29′53.61″ N; 71°59′1.36″ W) located in the municipality of Sherbrooke (Bromptonville Borough), which is part of the Estrie administrative region of southern Québec (southeastern Canada). Gentle slopes characterize the site and the regional topography [43]. The study site is within the sugar maple–basswood ecoregion, which is part of the broader northern hardwoods forest ecosystem [43,44]. The regional climate is a continental sub-humid moderate climate, with a growing season of 180–190 days [43], mean annual precipitation of 1145 mm, and a mean annual temperature of 5.6 °C [45]. The soil at the study site has developed on glacial outwash, deposited over lacustrine clay and is generally well-drained [46]. In the 0–60 cm depth range, soil pH ranged 5.7–6.4, soil texture varied within the loam soil class (loam, silty loam, sandy clay loam), and the soil is free of large stones [47].
The riparian buffer was established in spring 2010 along a 1 km segment of a small farm stream draining into the Saint-François River. The stream has been subjected to channel reconfiguration and streambank deforestation. A mean stream width of 1.6–2.0 m and a mean depth of 0.20 m were previously recorded at the study site during summer [48]. The land use adjacent to the riparian buffer is a pasture that is annually fertilized with cattle manure. This pasture supports a cattle density of approximately 0.6 cattle/ha (M. Beauregard, pers. comm.). Prior to the installation of stream fences in 2009, the livestock had full access to the stream and riparian zone, which substantially altered channel morphology and contributed to water quality degradation (B. Truax and J. Fortier, field observations). Mostly warm water fish species, tolerant of water pollution are found in this headwater stream [48]. Prior to the buffer establishment, the vegetation cover was dominated by herbaceous species (mostly pasture grasses and other ruderal species [39]), and was repeatedly grazed by the livestock. Following buffer establishment and fencing, the height of the herbaceous cover ranged 20–50 cm at its maximum development stage during the summer.
2.2. Experimental Design
In May 2010, a randomized block design was established to test the effect of tree species and vegetation treatment on tree growth and the dynamics of soil elements. Three vegetation treatments (control, herbicide, black plastic mulch) and three species (black walnut, bur oak, white pine), replicated in six blocks, were used in this factorial experiment. The experimental design contains 54 plots (3 Species × 3 Vegetation treatments × 6 blocks). Each block contains nine contiguous experimental plots. Each plot contains a single Species/Vegetation treatment combination. Each plot measures 4.5 × 4.5 m (20.5 m2/plot) and contains nine trees planted with a square spacing of 1.5 × 1.5 m (2.25 m2/tree). Each block measured 40.5 m in length (parallel to the stream) by 4.5 m of width (perpendicular to the stream). Topographically, all blocks were placed in areas presenting relatively homogeneous soil and no signs of inadequate drainage. All blocks were located outside of the bankfull stage (above the top of the streambank), but inside the floodplain zone. For the entire experimental design, a total of 486 trees were planted along three rows parallel to the stream.
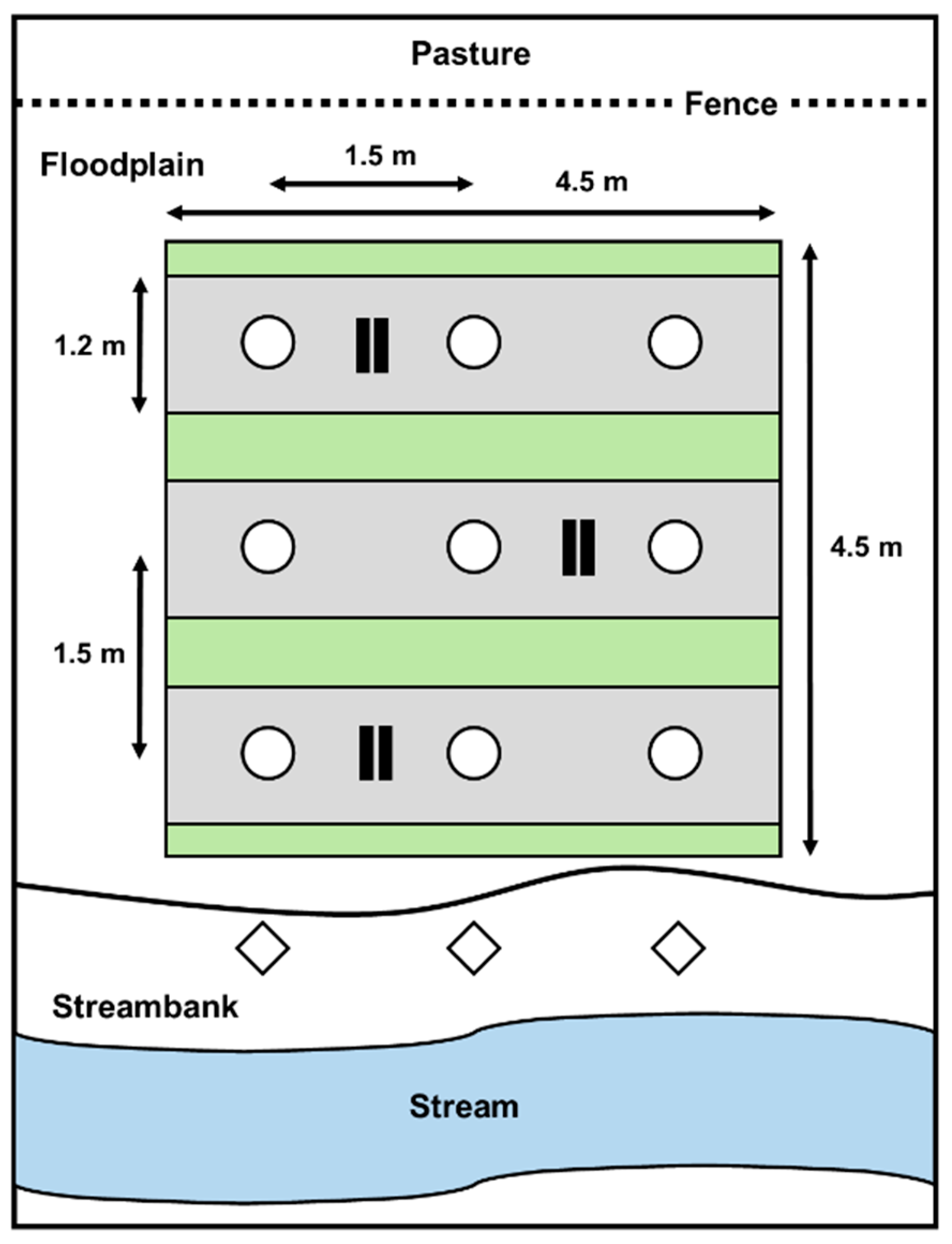
Blocks were spatially separated along a 1 km stream segment in order to maximize environmental variability at the site level. Between blocks, buffer zones were planted to create a continuous linear tree structure in the riparian zone, thereby reducing edge effects on trees located at the ends of blocks. These buffer zones contained the same tree species that were positioned in the first and last plots in a block and they were planted with the same spacing used in the experimental plots. Along all blocks, one row of hybrid willows (Salix × spp.) was planted outside the experimental design, between the top and the toe of the streambank (in the active stream channel zone). These willows were planted to enhance streambank stabilization and to eventually create an overhanging canopy structure above the stream, which provides stream shading and allochthonous inputs of organic matter and terrestrial insect preys [9,49,50]. Livestock was also excluded from the riparian buffer over the entire study duration with electrical fence wires that were installed in summer 2009. At the pasture edge of the experimental design, the fence was located at about 1.5–2.0 m from the first row of trees. A schematic representation of an experimental plot, where a vegetation treatment was applied, is shown in Figure 1.
All trees were planted manually with a shovel, directly through the herbaceous vegetation. No site preparation was done prior to planting. The herbicide (glyphosate) and the plastic mulch treatment were applied in 1.2 m wide strips on each of the three tree rows. The plastic mulch was installed shortly after tree planting (in mid-May 2010). Black plastic mulch strips (0.06 mm thick) were installed manually and pinned down with large wooden pegs. Rocks found nearby were used to maintain the mulch close to soil surface. Black plastic mulch rolls were purchased from Dubois Agrinovation Inc. (Napierville, QC, Canada). The plastic mulch strips remained on site for the entire duration of the study (2010–2014). The herbicide application was done once in mid-June of the first three growing seasons (2010, 2011, 2012). The herbicide was manually applied with a backpack sprayer and a cardboard tube was used to protect trees against drift.
The tree species used in this study were selected for their contrasted ecological characteristics. Black walnut is a naturalized hardwood species (native to southern Ontario, Canada) that mainly grows on well-drained bottomland sites [51]. Bur oak is a native hardwood species in the study area that mainly grows in bottomlands and on riparian sites experiencing short flood events [51]. White pine is a ubiquitous native species in the study area [51]. Characteristics of planted species are presented in Table 1. One-year-old bare-root seedlings were used for black walnut (1-0), two-year-old bare-root seedlings were used for bur oak (2-0), and two-year-old container seedlings were used for white pine (2-0). Height of seedlings at planting was 40 cm for black walnut, 44 cm for bur oak, and 21 cm for white pine. Seedlings were provided by the Berthier nursery (Sainte-Geneviève-de-Berthier, QC, Canada) of the Ministère des Forêts, de la Faune et des Parcs (MFFP) of Québec.
2.3. Measurement of Soil Nutrient Dynamics
In each experimental plot (n = 54), the dynamics (supply rate) of soil nutrients (NO3, NH4, P, calcium (Ca), magnesium (Mg), and sulfur (S)) was determined using Plant Root Simulator (PRSTM-Probes) technology from Western Ag Innovations Inc. (Saskatoon, SK, Canada). The PRS-probes are a type of ion exchange membrane encapsulated in thin plastic probes. The membrane’s surface exhibits sorption characteristics similar to those of a plant root. Nutrient supply rates measured with this method are strongly correlated with conventional soil extraction methods over a wide range of soil types [55], including the agricultural buffer soils of the study area [56]. This technology was also useful in understanding the effect of different tree species/vegetation treatment combinations on NO3 dynamics in a riparian buffer located on another stream 30 km south [15].
On 21 June of the fifth growing season (2014), three pairs of probes (an anion and a cation probe in each pair) were buried in each experimental plot for a 30-day period. The PRS-probes were inserted vertically in the shallow soil (0–10 cm, A horizon) with little disturbance of soil structure. Within a plot, each pair of probes was placed on each of the three tree rows, equidistant between two trees of the same row (Figure 1). On 21 July, probes were removed from the soil, washed in the field with distilled water, and returned to Western Ag Labs for analysis. Plot-level composite samples were made by combining the three pairs of probes collected in each plot. Probe supply rates are reported as µg of nutrient per 10 cm2 per 30 days.
2.4. Growth and Survival Measurements
At the end of the second, third, fourth, and fifth growing seasons (late October 2011–2014), total tree height, basal diameter, and, when possible, diameter at breast height (DBH at 1.3 m) were measured for each tree of the experimental design. A digital caliper was used to make diameter measurements (mean of two diameter measurements taken perpendicularly). Stem volume outside the bark was then calculated for two tree categories, those having no DBH and those having a DBH value. For trees with no DBH, the simple cone volume formula was used [57]:
where V is the stem volume (cm3), DB is the basal diameter (cm), and H is the tree height (cm). For trees with a DBH value, the stem volume was measured by summing the volume of two stem sections: (1) from basal diameter to DBH and (2) from DBH to tree tip. For stem Section 2, Equation (1) was used, but DB was replaced by a DBH value and H was replaced by the height of the stem section from DBH to the tree tip. For stem Section 1, the following formula was used [58]:
where, V is the volume (cm3) of a stem section, D1 is the base diameter (cm) of the stem section, D2 is the diameter (cm) at the top of the stem section, and L is the length (or height) of the stem section. Thus, the volume of stem section 1 was measured by replacing D1 by a basal diameter value, D2 by a DBH value, and L by 130 cm in Equation (2).
V = π DB2H/12
V = π/12(D12 + D22 + D1D2) L
2.5. Statistical Analyses
Main effects (Tree species and Vegetation treatment) and interaction effects (Tree species × Vegetation treatment) on measured variables were analyzed using analysis of variance (ANOVA) in a fixed factorial design [59]. Degrees of freedom were the following: Total, 53; Tree species, 2; Vegetation treatment, 2; Tree species × Vegetation treatment, 4; Block, 5; Error, 40. All of the ANOVAs were run with the complete set of data (3 species × 3 vegetation treatments × 6 blocks = 54 experimental plots). Being proportions, survival data were logit transformed prior to ANOVA [60], but survival rate results are reported in percent values. Main effects or interaction effects were declared statistically significant for four levels of significance (p < 0.1, p < 0.05, p < 0.01, and p < 0.001). The standard error of the difference (SED) was used to separate means [59]. Based on the number of degrees of freedom in this study, this mean separation method is equivalent to the least significant difference test (LSD) at an alpha level of 0.05 [59]. A priori contrasts were further used to test specific hypotheses between particular sets of means [59,61,62]. Contrasts are more powerful than pairwise a posteriori tests and can be used whether or not the F-test of the ANOVA is significant [59]. Using stem volume data collected after two, three, four, and five growing seasons, a multivariate analysis of variance (MANOVA) was used to test the Time factor and its interaction with other effects tested in this study [62]. The Pillai’s trace test-statistic was used to declare significant effects (Time; Time × Tree species; Time × Vegetation treatment; Time × Tree species × Vegetation treatment). Finally, pairwise correlations were used to identify significant correlations between soil nutrient supply rate (NO3, NH4, P, Ca, K, and Mg) and stem volume growth after 5 years. All statistical analyses were done using JMP 11 from SAS Institute (Cary, NC, USA).
3. Results
3.1. Soil Nutrient Dynamics
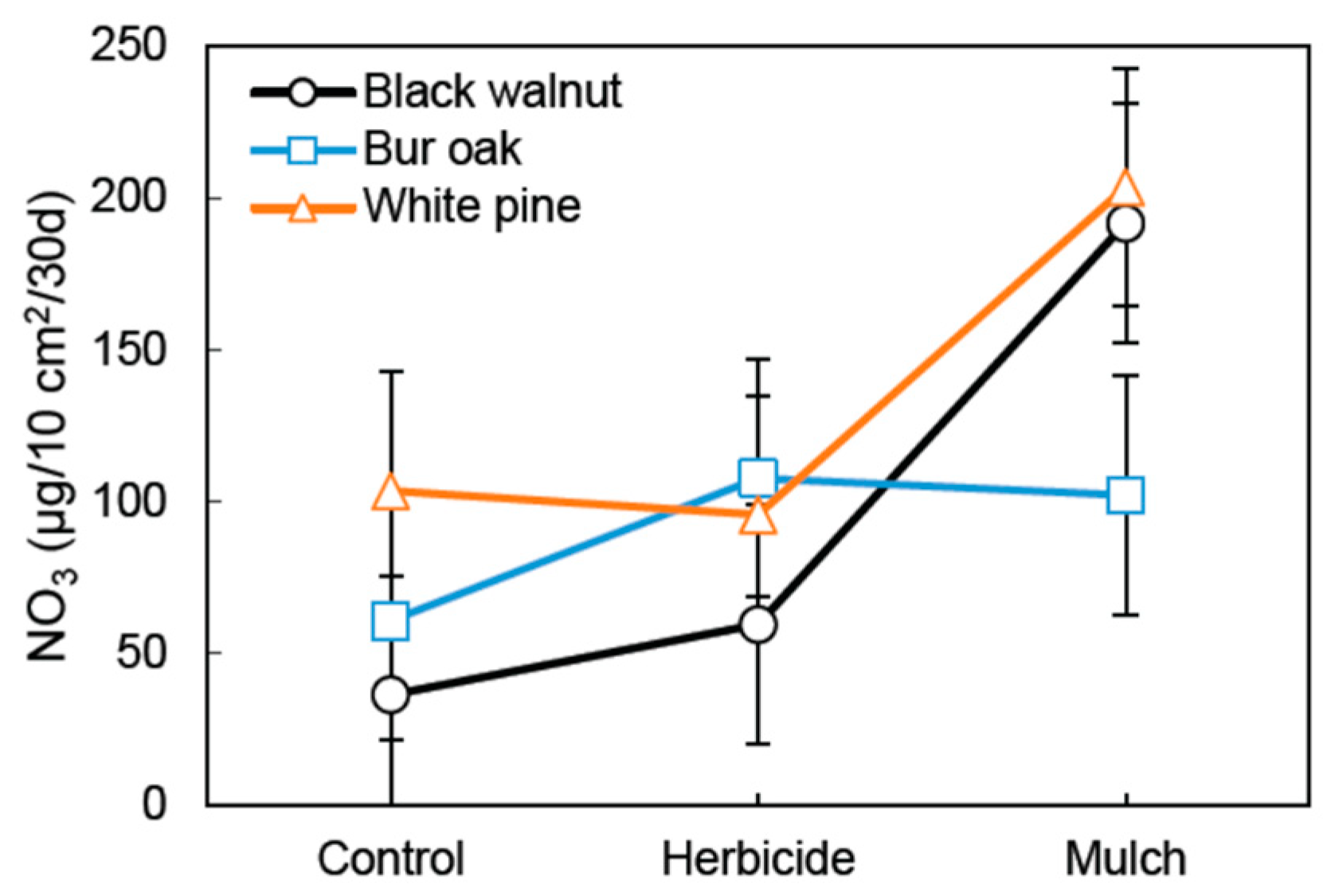
A nearly significant Tree species × Vegetation treatment interaction (p = 0.11) was observed on NO3 supply rate, with the highest values observed for black walnut and white pine in the mulch treatment (Figure 2). Contrast analysis further suggests that in the plastic mulch treatment, species that preferentially uptake NH4 (black walnut and white pine) were associated with a significantly higher NO3 supply rate than bur oak (p < 0.01) (Table 2). The Vegetation treatment effect was significant on the supply rate of several soil elements (Table 3). The strongest effect was observed for soil NO3 (p < 0.001), with the plastic mulch treatment having significantly higher soil NO3 compared to the herbicide treatment (p < 0.01) (Table 2). Despite that NO3 supply rate was found to be slightly higher in the herbicide vs. the control treatment, this difference was not significant (p = 0.37) (Table 2). The P supply rate was 64.3% higher in the mulch treatment compared to the herbicide treatment, but this difference was nearly significant (p = 0.11) (Table 3). A significant Vegetation treatment effect was observed for soil supply of S (p < 0.01) and Ca (p < 0.05). For those nutrients, the observed values in the plastic mulch treatment were significantly higher than in the control treatment (Table 3). However, when the mulch treatment was compared to the herbicide treatment for those elements, only the S supply rate was found to be significantly higher in the mulch treatment. Overall, few differences in soil nutrient dynamics were observed between the herbicide and the control treatment, while highest nutrient supply values were mainly found in the plastic mulch treatment.
3.2. Tree Growth and Survival
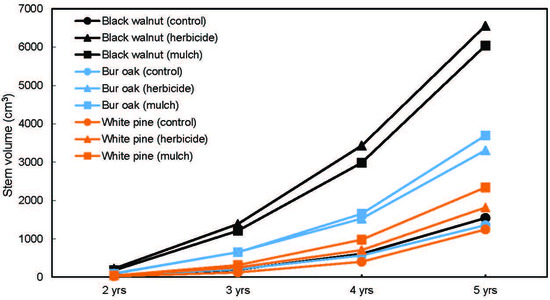
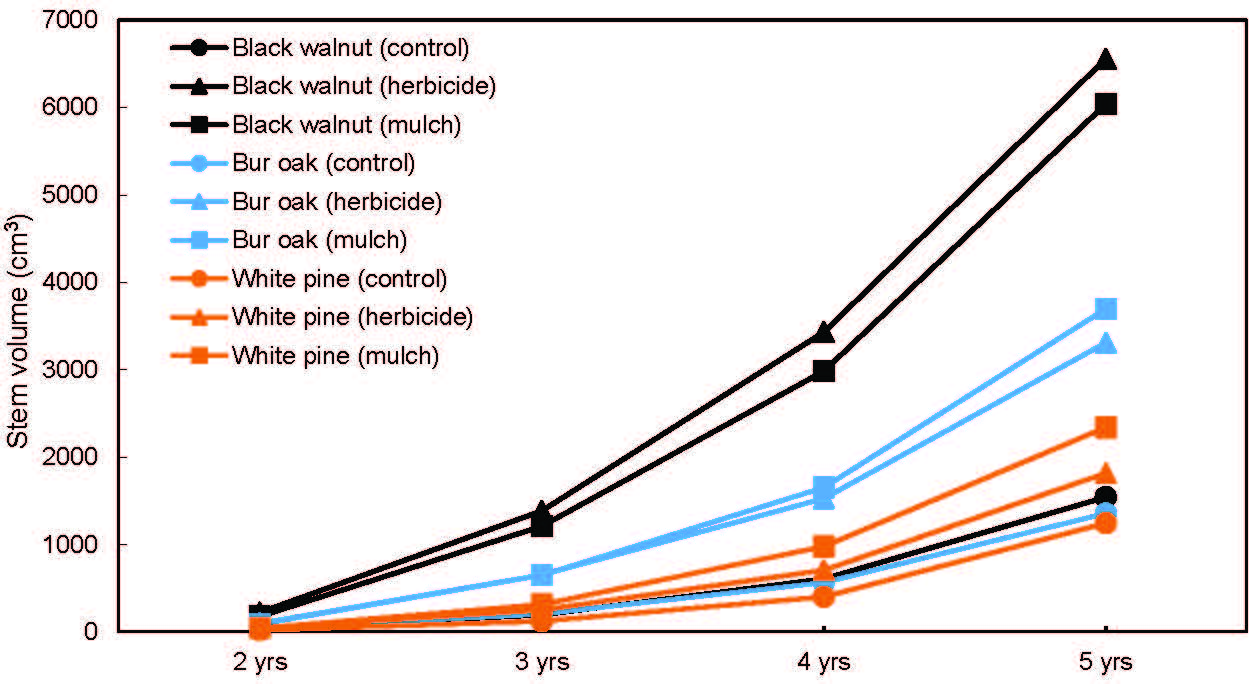
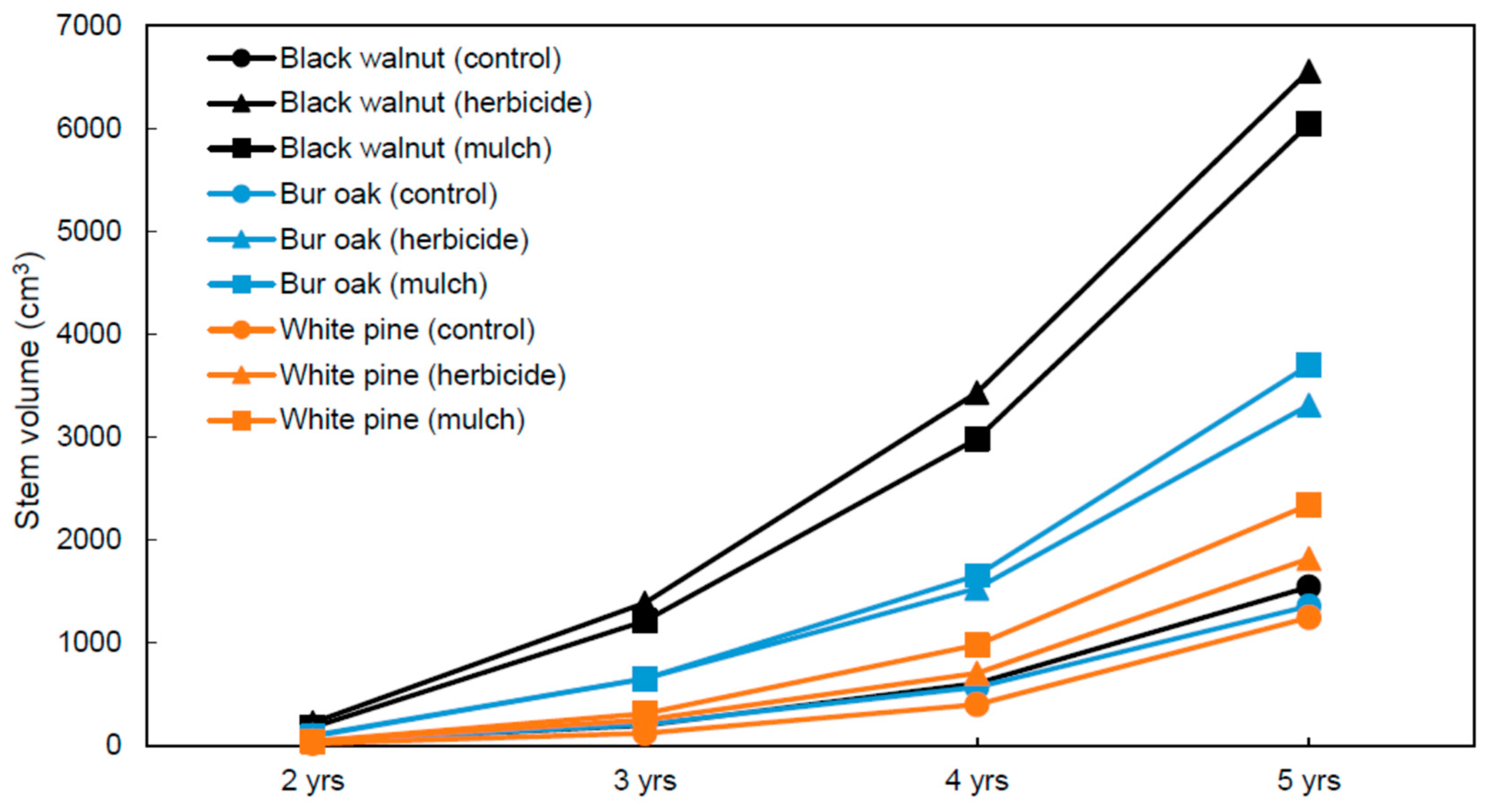
After five growing seasons, survival rate was above 93% for all species across all treatments (Figure 3). A significant Tree species effect was observed (p = 0.05), with bur oak having a significantly higher survival rate than white pine. The MANOVA done on stem volume measured at the end of each of the last four growing seasons detected a significant Tree species × Vegetation treatment × Time interaction (p < 0.05) (Figure 4). In the control treatment, stem volume between the three species remained within a relatively narrow range of values at the end of each growing season (21–39 cm3 after 2 years, 121–195 cm3 after 3 years, 400–605 cm3 after 4 years, and 1244–1541 cm3 after 5 years), with the lowest volume always observed for white pine. However, in the herbicide and plastic mulch treatments, a much wider range of stem volume values was observed between the three species across the last 4 years. In the herbicide treatment, stem volume ranged 45–223 cm3 after 2 years, 251–1386 cm3 after 3 years, 705–3432 cm3 after 4 years, and 1871–6557 cm3 after 5 years, while in the plastic mulch treatment, stem volume ranged 43–178 cm3 after 2 years, 312–1211 cm3 after 3 years, 979–2982 cm3 after 4 years, and 2340–6040 cm3 after 5 years. In the herbicide and mulch treatments, species ranking for stem volume was always the same at the end of each growing season: black walnut > bur oak > white pine. Also, for a given species, the variation in stem volume between the herbicide and plastic mulch treatment was marginal from year 2 to year 5 (Figure 4).
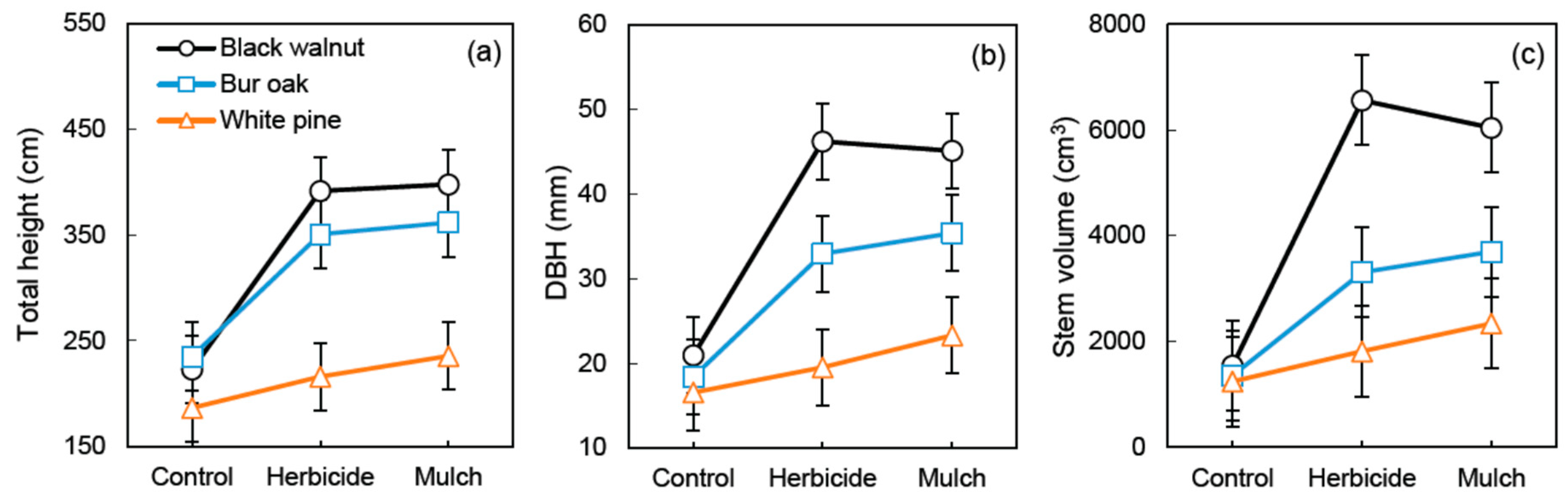
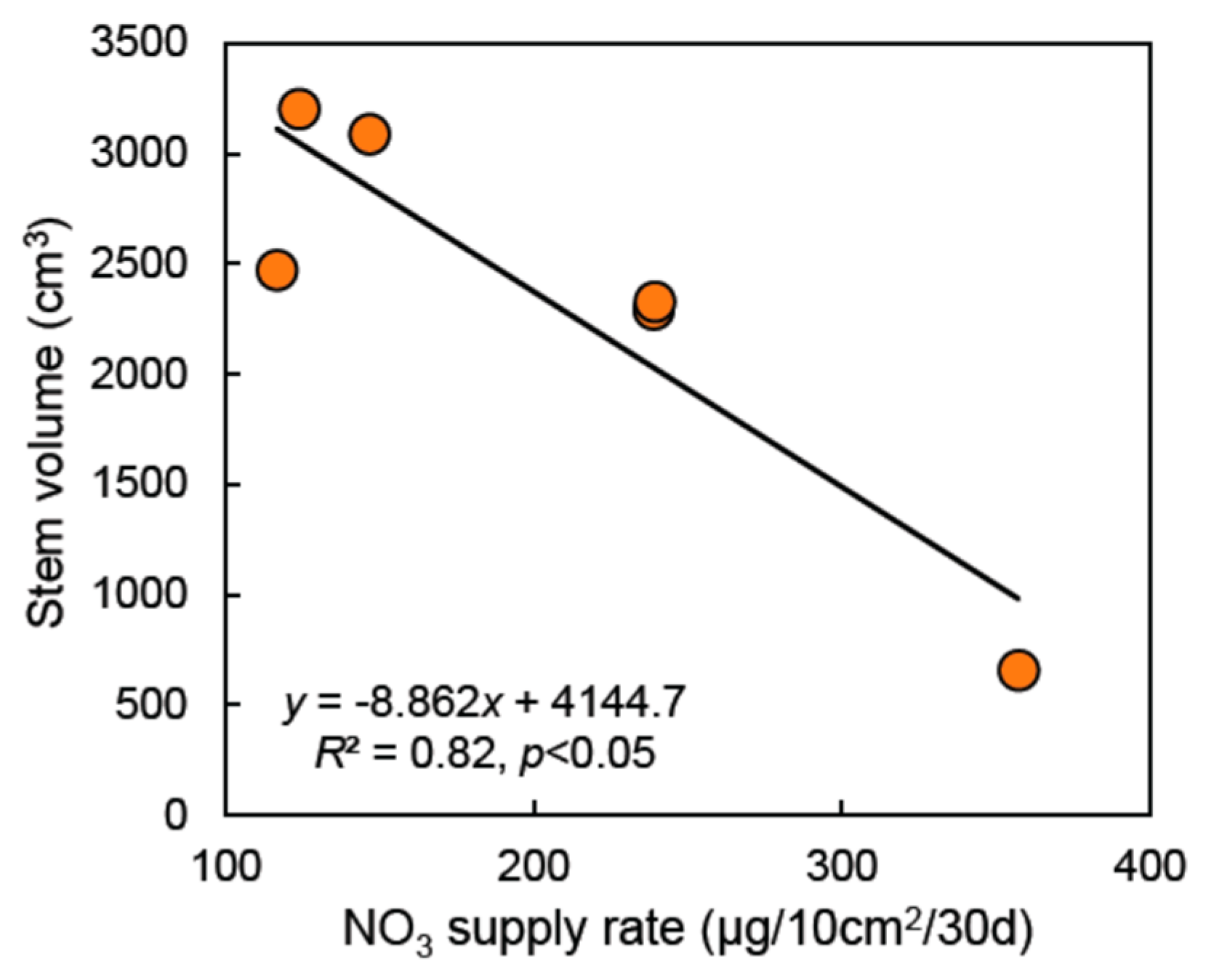
More specifically, after five growing seasons, a significant Tree species × Vegetation treatment interaction was observed on total height (p < 0.05), DBH (p < 0.05), and stem volume growth (p < 0.01) (Figure 5). Furthermore, for all growth variables measured after 5 years (height, DBH, basal diameter, and stem volume), main effects (Tree species and Vegetation treatment) were always highly significant (p < 0.001). All of those growth indicators showed a positive response of each species to both vegetation management treatments. However, the magnitude of the growth response varied between species, with black walnut having the strongest response to vegetation treatments and white pine the weakest (Figure 5, Table 4). Indeed, although stem volume of white pine was increased by 46% in the herbicide treatment and by 88% in the plastic mulch treatment, its volume growth in those treatments was not significantly different from the control treatment (p = 0.25) (Table 4). After 5 years, stem volume of black walnut was 3.9–4.2 times higher in the vegetation treatments than in the control treatment, while for bur oak stem volume was 2.4–2.7 times higher in the vegetation treatments than in the control. Yet, at the species level, there were no statistical differences in any growth indicators when the herbicide and the black plastic mulch treatments where compared after 5 years (Figure 5, Table 4). Finally, in the black plastic mulch treatment, a strong negative relationship (R2 = 0.82, p < 0.05) between soil NO3 supply rate and stem volume growth was observed for white pine during the fifth growing season.
4. Discussion
4.1. Vegetation Treatments and Tree Species Affect Riparian Buffer Growth and Soil Nutrient Dynamics
The selection of the optimal vegetation treatment can greatly accelerate tree establishment and growth in riparian buffers, and thus, reduce the amount of time needed to restore ecosystem services and functions that are linked to the buffer’s structural attributes (habitats and movement corridors for forest species, carbon and nutrient storage in woody biomass, flood and wind protection, pesticide drift interception, stream shading, streambank stabilisation, etc.) [63]. We had hypothesized that superior growth would be observed for all species in the plastic mulch treatment because this treatment generally provides a longer-term increase in soil N availability compared to an herbicide treatment that would be stopped after 3 years [15,64]. As expected, plastic mulch was more effective at maintaining high soil NO3 supply over the long term (Table 2 and Table 3). Furthermore, compared to the herbicide, plastic mulch also increased the supply of P, another growth limiting nutrient [65], and the supply of S, a secondary macronutrient important for the synthesis of some amino-acids and vitamins [66] (Table 3). Although not measured in this study, permanent plastic mulching may also be superior to a 3-year herbicide treatment to increase soil and air temperatures, and soil moisture content over the long term [67]. However, despite the more favorable growth conditions provided by the mulch, the growth of all species was not statistically different between the herbicide and the mulch treatments (Figure 5, Table 4). In the Canadian Prairies, herbicide and plastic mulch also provided similar growth gains for four-year-old agroforestry trees belonging to various genera [30].
All studied species benefited from both vegetation treatments, but the magnitude of the growth response was species-specific; a trend that became more and more evident over the years (Figure 4). Hardwood species had the strongest growth response to the vegetation treatments and white pine had the weakest (Figure 4 and Figure 5), which supports our second hypothesis and previous observations made in a nearby riparian buffer and in agroforestry systems of the Prairies [15,30]. In fact, stem volume, height, and diameter growth were not statistically different between the three vegetation treatments for white pine (Figure 5). Compared to most hardwood species, white pine has relatively low nutrient requirements to achieve optimal growth [16], but also has a very effective N retention strategy, where N losses through litter fall are minimized and N residence time in biomass is relatively long [68]. Such a N cycling strategy allows white pine to colonise sites dominated by herbaceous species, even though these species are strong competitors for mineral N in shallow soil [68]. Thus, vegetation management is not essential for white pine establishment in riparian buffers.
On the other hand, for the studied hardwoods, the use of a vegetation treatment resulted in major stem volume increases (292–325% for black walnut and 144–173% for bur oak). Hence, although all species had comparable height, DBH, and stem volume growth after 5 years in the control treatment, tree size varied considerably between species in the plastic mulch and in the herbicide treatments (Figure 5). The strong growth response of black walnut to vegetation management is consistent with previous observations within the Juglans genus [23,69,70,71]. Black walnut is especially sensitive to soil conditions as it requires deep, well-drained, nearly neutral pH, moist and fertile soil to achieve optimal growth [52]. Such favorable soil conditions were found at the study site (see Section 2.1). However, without proper management of herbaceous competition, this species remains far from reaching optimal growth conditions in agricultural riparian buffers (Figure 4 and Figure 5). The same can be said about bur oak, although this species is known for its good tolerance to herbaceous competition [15,54]. Bur oak also has a more conservative early-growth pattern than black walnut [52,54] (Table 1), which can explain its intermediate growth response to both vegetation treatments (Figure 4 and Figure 5).
In agreement with our third hypothesis, and with previous findings near the study area [15], results show that the use of black plastic mulch with tree species having a preference for NH4 uptake (white pine and black walnut) [33,42] can highly increase the NO3 supply rate in riparian soils, even on the longer term (Figure 2, Table 2). Thus, particular tree species/vegetation treatment combinations may create undesirable effects on soil N, especially when the goal is to mitigate non-point source NO3 pollution reaching farm streams. Thus, even though black walnut grew a larger stem volume than bur oak in the plastic mulch treatment (Figure 5), the soil NO3 supply rate in mulched walnuts was about two times higher than in mulched bur oaks (Figure 2). Such a result may appear counterintuitive given that black walnut requires high soil fertility to reach its full growth potential [52,72]. Previous studies have also reported reduced stand growth, accumulation of soil NO3, and NO3 leaching loss in a mature Pinus plantation where a high dose of N fertilizer was applied for three consecutive years [73]. Our observations are consistent with such growth decline in pine under elevated soil NO3. Despite better growth of white pine in the plastic mulch treatment (Figure 5), a strong negative correlation between soil NO3 supply and volume growth (R2 = 0.82, p < 0.05) was observed in mulched pines (Figure 6). Thus, it is potentially not the higher soil NO3 supply that enhances white pine growth in the plastic mulch treatment, but the higher soil P supply (Table 3). While N limitation is the main factor controlling photosynthesis in deciduous species, P limitation may interact with N in controlling peak photosynthetic capacity in white pine [74]. Also, under open field conditions, white pine develops many branches close to the ground where air temperature is expected to be maximal in the plastic mulch treatment. Such warmer air conditions would enhance the photosynthetic activity of white pines [29].
As hypothesized, the NO3 supply rate measured during the fifth growing season was not statistically different between the herbicide and control treatments (Figure 2 and Table 2 and Table 3). Thus, although herbicide treatments generally enhance soil NO3 during the year of application [24], they have little residual effects on soil NO3 dynamics in the subsequent years. The regrowth of herbaceous vegetation underneath trees in the herbicide treatment after the last application (third growing season) may explain such a result (Figure 7). Most herbaceous plants found in the agricultural riparian zone of the study site are ruderal plants [39], which are well-known for their high capacity to uptake soil NO3 [35]. In a previous study, soil NO3 supply rate did not differ significantly in the control treatment (no vegetation management) between five different tree species with contrasted growth rates and nutritional requirements [15], a finding corroborated by this study (Figure 2). Recent evidence also suggests that herbaceous plants are more effective than trees at reducing NO3 leaching in agricultural riparian zones [40].
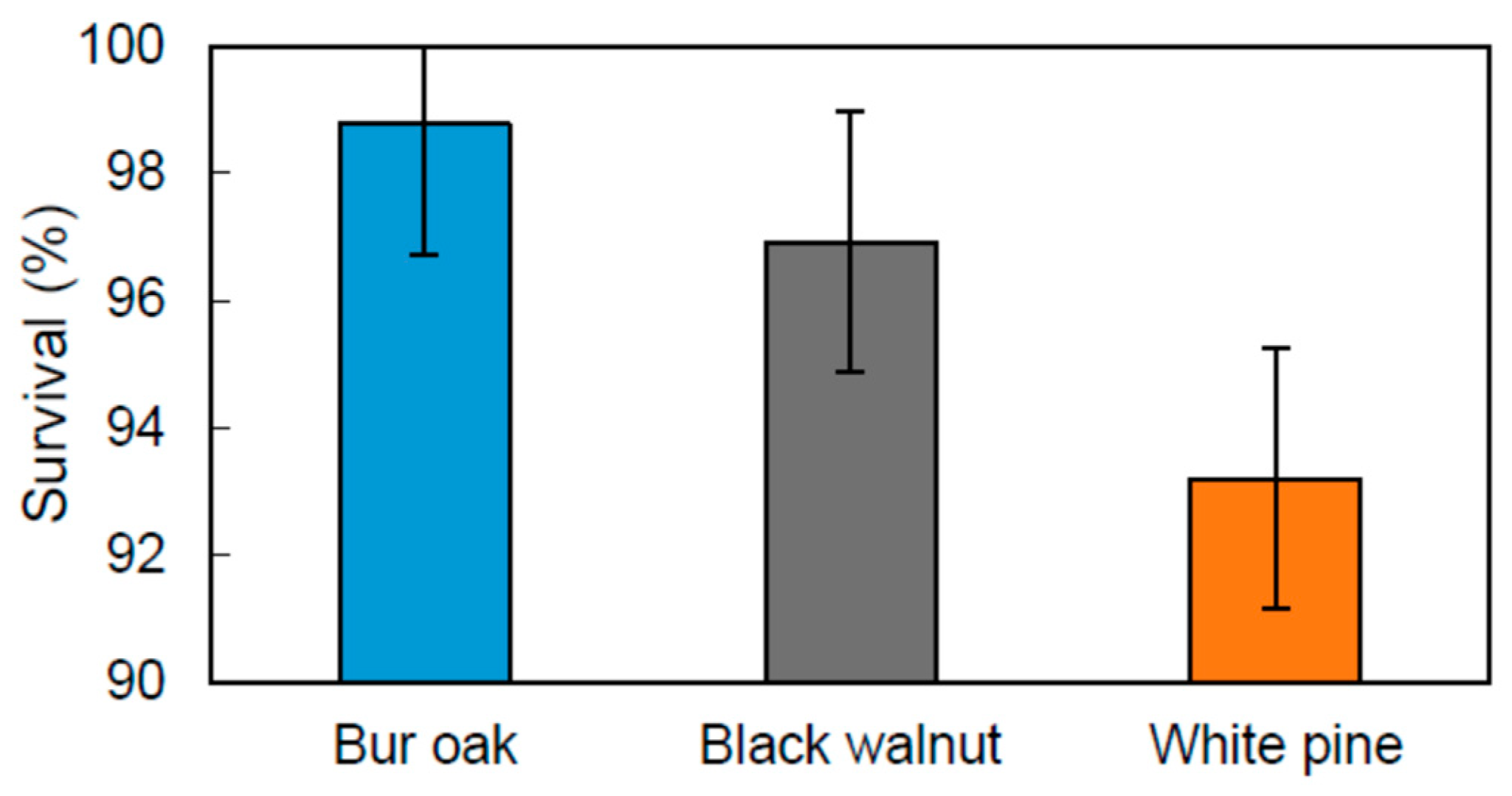
After 5 years, a very high survival rate (>93%) was achieved by all species (Figure 3) and the use of a vegetation treatment was not a significant factor affecting survival (p = 0.18), as equally observed by Sweeney and Czapka [11] in a floodplain ecosystem. Tree species selection is sometimes more important to increase survival rate than the use of vegetation treatments [75]. However, in other riparian afforestation studies, where survival rates were low, competing vegetation management was a determining factor affecting tree survival [18,31]. High survival rates were potentially linked to the high soil fertility and the good soil moisture conditions that characterised the studied riparian zone, which has a good hydrological connectivity with the stream (B. Truax and J. Fortier, field observations). The presence of electric fence wires along the stream corridor may have equally reduced herbivory by the white-tailed deer (Odocoileus virginianus Zimm.), which has become a serious problem for hardwood and white pine regeneration regionally [76]. Also, the species selected for this study may have contributed to obtaining high survival rates, even in the control treatment. White pine has a high capacity to invade sites dominated by herbaceous plants [68], while black walnut and bur oak, two early-successional species of bottomlands, rapidly form a deep tap root, which increases establishment success in environments dominated by shallow-rooted herbaceous plants [52,77]. However, over the years, growth stagnation related to N limitation may occur for black walnut in the control treatment, as observed in field plantations receiving only a few years of vegetation management [70].
4.2. Selecting a Vegetation Treatment for the Establishment of Trees in Riparian Buffers
The wide-scale and intensive use of herbicides in agriculture has raised many concerns in the last decades and is still keenly debated today. A high herbicide use is known to pose serious threats to aquatic biodiversity, water quality, and human health [78,79,80]. Although much less intensive, the use of herbicide to establish tree riparian buffers remains controversial because herbicides are applied in zones that are hydrologically interconnected with freshwater ecosystems. The social acceptability related to the use of herbicides in tree plantations is also relatively low [81].
Because of the many limitations related to herbicide use, black plastic mulch is increasingly used along farm streams and is generally perceived as a more sustainable vegetation treatment by land managers and ecological engineers. Yet, the use of plastic mulches also leads to noticeable impacts (degradation of stable organic matter, micro-plastics and phthalates leaching, modification of soil structure and infiltration properties, impeded root growth, lack of recycling opportunity for plastics contaminated with soil particles) [41,82,83,84]. Moreover, plastic mulches are rarely removed from plantation sites because this operation is time consuming and expensive [41]. As a result, the impacts of plastic mulch on the soil properties will be long-lasting and should be fully considered as riparian buffer pollutant removal efficiency is tightly bound to water infiltration capacity in soil, to soil health, and to the colonization of soil by plant roots [3]. Also, the permanent barrier created by mulches restricts herbaceous vegetation growth underneath trees (Figure 7c), an important consequence because herbaceous plants have a central role in runoff and N leaching reduction [40,85]. In this study, soil NO3 and P supplies after 5 years were the highest in the plastic mulch treatment, although species-specific trends were observed for soil NO3 (Figure 2, Table 3). This situation may be undesirable, since many agricultural riparian buffers are designed to phytoremediate excess soil NO3 and P generated by adjacent agricultural use. The use of plastic mulch will also reduce the area of riparian habitats that can be colonised by herbaceous plants, shrubs, and tree species, which would otherwise contribute toward creating a more diversified, multifunctional, and resilient buffer system. On the other hand, riparian zones can be important reservoirs of agricultural weeds and invasive exotic plants [6,86,87]. Therefore, the reduction in plant habitat size caused by plastic mulches could be seen positively by some farmers, land planners, and ecological engineers.
Herbicide treatments also have noticeable effects on targeted plant communities, with changes in plant species dominance being more frequent than changes in species composition and diversity [20]. In terms of impacts on soil, extensive herbicide use in tree plantations mostly affects C stocks in the organic layer, with only minor effects being observed on mineral C stocks and stability [64,88]. Moreover, extensive use of glyphosate-based herbicides in tree plantations does not represent a significant risk to human health and to terrestrial and aquatic ecosystems [89]. From an economic perspective, herbicide application is by far the more cost-effective compared to plastic mulching [32]. Technically, plastic mulch application is also more restrictive than an herbicide treatment, which can be done with a backpack sprayer, in almost all types of riparian terrains, including steep and stony streambanks. Furthermore, if a plastic mulch layer is used to install mulch strips, soil tillage may be required, while no such site preparation is needed prior to an herbicide treatment for tree buffer establishment. Finally, from the perspective of agricultural land aesthetics, the visual impact of an herbicide treatment is only temporary, while black plastic mulch remains visible for many years.
Both vegetation treatments used in this study had a similar effectiveness for enhancing hardwood growth (Figure 5, Table 4) but had different environmental impacts. Hence, their use in streamside buffers should be made with the goal of reducing their potential negative impacts, both in regard to herbaceous vegetation and non-target organisms. From that perspective, it is recommended to apply herbicide manually, outside of periods with high runoff or flood event probability, and to use herbicide formulations recommended for aquatic use, which are less toxic to stream fauna [90]. Herbicides should be applied in strips or in spots [11,21], and the number of applications kept to a minimum. Additional studies are needed to identify the most efficient extensive herbicide application strategies for different tree species planted across a gradient of riparian sites differing in resource availability and in competing vegetation characteristics. In some riparian environments, tree survival and growth rate may not be significantly affected by the use of a single or a multi-year herbicide treatment [91]. The identification of key site variables responsible for such a lack of response is essential to the future selection of sites where tree buffers could be installed at lower costs (with no vegetation management). If plastic mulch is used to establish riparian buffers, planting tree species that are nitrophilous is recommended in order to minimize the accumulation of soil NO3 underneath mulches. Plastic mulches should also be retrieved from riparian sites once trees are well established. Finally, it should be acknowledged that the faster a riparian buffer grows, the faster it will provide key ecosystem services and functions. Thus, land managers and ecological engineers should fully consider the tradeoffs related to the use of a particular vegetation management treatment in the specific case of tree riparian buffer establishment and growth on agricultural land.
5. Conclusions
This study has shown that plastic mulch and herbicide, applied in 1.2 m wide strips, have a similar efficacy to enhance tree growth after 5 years in a fertile agricultural riparian buffer, despite the fact that plastic mulch provides longer-term improvement of soil nutrient status. Because black walnut and bur oak were highly responsive to plastic mulching and herbicide, the use of these vegetation treatments will greatly shorten the time needed to reach canopy closure with those hardwoods. On the other hand, white pine growth was less affected by both vegetation treatments, which indicates that this species can be successfully planted at low cost and with little impact along headwater streams on farmland.
The high NO3 supply rate observed in the soil underneath plastic mulch where species that preferentially uptake NH4 (black walnut and white pine) grew suggests caution when selecting particular tree species/vegetation treatment combinations, especially if the objective of the riparian buffer is to mitigate NO3 pollution in waterbodies. Non-nitrophilous tree species potentially have a limited capacity to reduce NO3 leaching from intensive cropping systems. The high soil P supply generated by the plastic mulch treatment may also be undesirable over the long term in riparian buffer soils. Conversely, after 5 years, soil nutrient supply was similar between plots receiving no vegetation treatment and plots where a 3-year herbicide treatment was applied, indicating little residual effect of the herbicide on soil nutrient status. In a context where plastic mulch is rarely removed following riparian buffer establishment, further studies are needed to evaluate the long-term impacts of such a vegetation treatment on riparian buffer soils.
Author Contributions
B.T. and D.G. conceived and designed the experiment. B.T. and F.L. were involved in sampling design and field sampling. J.F., B.T., and F.L. analyzed the data. J.F. wrote the first draft of the manuscript. B.T., D.G., and F.L. critically revised the manuscript.
Acknowledgments
We gratefully acknowledge Agriculture and Agri-Food Canada (Agricultural Greenhouse Gas Program), the Ministère des Forêts, de la Faune et des Parcs du Québec, and Tree/Arbres Canada for the funding received. We wish to thank tree planters and field assistants for their help (F. Gendron, A. Laflamme, J. Lemelin, L. Meulien, M.-A. Pétrin, A. Richard, L. Godbout). We also thank M. Beauregard and C. Vincent (Ferme Carocel) who have kindly allowed the use of part of their property for the establishment of the riparian buffer. We acknowledge the Berthier nursery of the Ministère des Forêts, de la Faune et des Parcs of Québec for providing high quality planting stock.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Muscutt, A.D.; Harris, G.L.; Bailey, S.W.; Davies, D.B. Buffer zones to improve water quality: A review of their potential use in UK agriculture. Agric. Ecosyst. Environ. 1993, 45, 59–77. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Blaine, J.G. River conservation, restoration, and preservation: Rewarding private behavior to enhance the commons. Freshw. Sci. 2016, 35, 755–763. [Google Scholar] [CrossRef]
- Dosskey, M.G.; Vidon, P.; Gurwick, N.P.; Allan, C.J.; Duval, T.P.; Lowrance, R. The role of riparian vegetation in protecting and improving chemical water quality in streams. JAWRA 2010, 46, 261–277. [Google Scholar]
- Mander, Ü.; Hayakawa, Y.; Kuusemets, V. Purification processes, ecological functions, planning and design of riparian buffer zones in agricultural watersheds. Ecol. Eng. 2005, 24, 421–432. [Google Scholar] [CrossRef]
- Tabacchi, E.; Lambs, L.; Guilloy, H.; Planty-Tabacchi, A.M.; Muller, E.; Décamps, H. Impacts of riparian vegetation on hydrological processes. Hydrol. Proc. 2000, 14, 2959–2976. [Google Scholar] [CrossRef]
- Boutin, C.; Jobin, B.; Bélanger, L. Importance of riparian habitats to flora conservation in farming landscapes of southern Québec, Canada. Agric. Ecosyst. Environ. 2003, 94, 73–87. [Google Scholar] [CrossRef]
- Jobin, B.; Bélanger, L.; Boutin, C.; Maisonneuve, C. Conservation value of agricultural riparian strips in the Boyer River watershed, Québec (Canada). Agric. Ecosyst. Environ. 2004, 103, 413–423. [Google Scholar] [CrossRef]
- Lovell, S.T.; Sullivan, W.C. Environmental benefits of conservation buffers in the United States: Evidence, promise, and open questions. Agric. Ecosyst. Environ. 2006, 112, 249–260. [Google Scholar] [CrossRef]
- Baxter, C.V.; Fausch, K.D.; Saunders, W.C. Tangled webs: Reciprocal flows of invertebrate prey link streams and riparian zones. Freshw. Biol. 2005, 50, 201–220. [Google Scholar] [CrossRef]
- Schultz, R.C.; Isenhart, T.M.; Simpkins, W.W.; Colletti, J.P. Riparian forest buffers in agroecosystems—Lessons learned from the Bear Creek Watershed, central Iowa, USA. Agrofor. Syst. 2004, 61–62, 35–50. [Google Scholar]
- Sweeney, B.W.; Czapka, S.J. Riparian forest restoration: Why each site needs an ecological prescription. For. Ecol. Manag. 2004, 192, 361–373. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Silvics of North America; Forest Service Agriculture: Washington, DC, USA, 1990. [Google Scholar]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Potential for hybrid poplar riparian buffers to provide ecosystem services in three watersheds with contrasting agricultural land use. Forests 2016, 7, 37. [Google Scholar] [CrossRef]
- Rheinhardt, R.; Brinson, M.; Meyer, G.; Miller, K. Carbon storage of headwater riparian zones in an agricultural landscape. Carbon Bal. Manag. 2012, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Riparian buffer growth and soil nitrate supply are affected by tree species selection and black plastic mulching. Ecol. Eng. 2017, 106, 82–93. [Google Scholar] [CrossRef]
- Wendel, G.W.; Clay Smith, H. Eastern white pine. In Silvics of North America: 1. Conifers. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Andrews, D.M.; Barton, C.D.; Czapka, S.J.; Kolka, R.K.; Sweeney, B.W. Influence of tree shelters on seedling success in an afforested riparian zone. New For. 2010, 39, 157–167. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Czapka, S.J.; Yerkes, T. Riparian forest restoration: Increasing success by reducing plant competition and herbivory. Rest. Ecol. 2002, 10, 392–400. [Google Scholar] [CrossRef]
- Davies, R.J. The importance of weed control and the use of tree shelters for establishing broadleaved trees on grass-dominated sites in England. Forestry 1985, 58, 167–180. [Google Scholar] [CrossRef]
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2006, 79, 3–27. [Google Scholar] [CrossRef]
- Fortier, J.; Gagnon, D.; Truax, B.; Lambert, F. Biomass and volume yield after 6 years in multiclonal hybrid poplar riparian buffer strips. Biomass Bioenergy 2010, 34, 1028–1040. [Google Scholar] [CrossRef]
- Borin, M.; Vianello, M.; Morari, F.; Zanin, G. Effectiveness of buffer strips in removing pollutants in runoff from a cultivated field in North-East Italy. Agric. Ecosyst. Environ. 2005, 105, 101–114. [Google Scholar] [CrossRef]
- Lambert, F.; Truax, B.; Gagnon, D.; Chevrier, N. Growth and N nutrition, monitored by enzyme assays, in a hardwood plantation: Effects of mulching materials and glyphosate application. For. Ecol. Manag. 1994, 70, 231–244. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Chevrier, N. Nitrate reductase activity in relation to growth and soil N-forms in red oak and red ash planted in three different environments: Forest, clear-cut and field. For. Ecol. Manag. 1994, 64, 71–82. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D. Effects of straw and black plastic mulching on the initial growth and nutrition of butternut, white ash and bur oak. For. Ecol. Manag. 1993, 57, 17–27. [Google Scholar] [CrossRef]
- Munson, A.D.; Margolis, H.A.; Brand, D.G. Intensive silvicultural treatment: Impacts on soil fertility and planted conifer response. Soil Sci. Soc. Am. J. 1993, 57, 246–255. [Google Scholar] [CrossRef]
- Black, A.; Greb, B. Nitrate accumulation in soils covered with plastic mulch. Agron. J. 1962, 54, 366. [Google Scholar] [CrossRef]
- Kwabiah, A.B. Growth and yield of sweet corn (Zea mays L.) cultivars in response to planting date and plastic mulch in a short-season environment. Sci. Hortic. 2004, 102, 147–166. [Google Scholar] [CrossRef]
- Dreyer, E.; Le Roux, X.; Montpied, P.; Daudet, F.A.; Masson, F. Temperature response of leaf photosynthetic capacity in seedlings from seven temperate tree species. Tree Physiol. 2001, 21, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, W.R.; Naeem, H. Effect of weed control methods on growth of five temperate agroforestry tree species in Saskatchewan. For. Chron. 2017, 93, 271–281. [Google Scholar] [CrossRef]
- Smaill, S.J.; Ledgard, N.; Langer, E.R.; Henley, D. Establishing native plants in a weedy riparian environment. N. Z. J. Mar. Freshw. Res. 2011, 45, 357–367. [Google Scholar] [CrossRef]
- Von Althen, F.W. Effects of weed control on the survival and growth of planted black walnut, white ash and sugar maple. For. Chron. 1971, 47, 223–226. [Google Scholar] [CrossRef]
- Bauer, G.A.; Berntson, G.M. Ammonium and nitrate acquisition by plants in response to elevated CO2 concentration: The roles of root physiology and architecture. Tree Physiol. 2001, 21, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Beckjord, P.R.; Adams, R.E.; Smith, D.W. Effects of nitrogen fertilization on growth and ectomycorrhizal formation of red oak. For. Sci. 1980, 26, 529–536. [Google Scholar]
- Gebauer, G.; Rehder, H.; Wollenweber, B. Nitrate, nitrate reduction and organic nitrogen in plants from different ecological and taxonomic groups of Central Europe. Oecologia 1988, 75, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Walters, M.B.; Willis, J.L.; Gottschalk, K.W. Seedling growth responses to light and mineral N form are predicted by species ecologies and can help explain tree diversity. Can. J. For. Res. 2014, 44, 1356–1368. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Biomass carbon, nitrogen and phosphorus stocks in hybrid poplar buffers, herbaceous buffers and natural woodlots in the riparian zone on agricultural land. J. Environ. Manag. 2015, 154, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Fortier, J.; Gagnon, D.; Truax, B.; Lambert, F. Understory plant diversity and biomass in hybrid poplar riparian buffer strips in pastures. New For. 2011, 42, 241–265. [Google Scholar] [CrossRef]
- Neilen, A.D.; Chen, C.R.; Parker, B.M.; Faggotter, S.J.; Burford, M.A. Differences in nitrate and phosphorus export between wooded and grassed riparian zones from farmland to receiving waterways under varying rainfall conditions. Sci. Total Environ. 2017, 598, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Kasirajan, S.; Ngouajio, M. Polyethylene and biodegradable mulches for agricultural applications: A review. Agron. Sustain. Dev. 2012, 32, 501–529. [Google Scholar] [CrossRef]
- Nicodemus, M.; Salifu, K.; Jacobs, D. Nitrate reductase activity and nitrogen compounds in xylem exudate of Juglans nigra seedlings: Relation to nitrogen source and supply. Tree Struct. Funct. 2008, 22, 685–695. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.-P. Paysages Régionaux du Québec Méridional; Les Publications du Québec: Ste-Foy, QC, Canada, 1998. [Google Scholar]
- Westveld, M. Natural forest vegetation zones of New England. J. For. 1956, 54, 332–338. [Google Scholar]
- Government of Canada. Station Results—1981–2010 Climate Normals and Averages. Available online: http://climate.weather.gc.ca/climate_normals/station_select_1981_2010_e.html?searchType=stnProv&lstProvince=QC (accessed on 16 February 2017).
- Cann, D.B.; Lajoie, P. Études Des Sols Des Comtés de Stanstead, Richmond, Sherbrooke et Compton Dans la Province de Québec; Ministère de l’Agriculture: Ottawa, ON, Canada, 1943.
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Root biomass and soil carbon distribution in hybrid poplar riparian buffers, herbaceous riparian buffers and natural riparian woodlots on farmland. SpringerPlus 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Simavi, M.A. Effet de Plantations de Bandes Riveraines d’arbres sur l’abondance et la Répartition de la Faune Aquatique dans des Ruisseaux Dégradés de Milieux Agricoles dans Les Cantons-de-l’Est. M.Sc. Thesis, Université du Québec à Montréal, Montréal, QC, Canada, 2012. [Google Scholar]
- Lyons, J.; Thimble, S.W.; Paine, L.K. Grass versus trees: Managing riparian areas to benefit streams of central North America. JAWRA 2000, 36, 919–930. [Google Scholar] [CrossRef]
- Wallace, J.B.; Eggert, S.L.; Meyer, J.L.; Webster, J.R. Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 1997, 277, 102–104. [Google Scholar] [CrossRef]
- Farrar, J.L. Les Arbres du Canada; Fides et le Service Canadien des Forêts, Ressources naturelles Canada: St-Laurent, QC, Canada, 2006. [Google Scholar]
- Williams, R.D. Black walnut. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Kabrick, J.M.; Dey, D.C.; Van Sambeek, J.W.; Coggeshall, M.V.; Jacobs, D.F. Quantifying flooding effects on hardwood seedling survival and growth for bottomland restoration. New For. 2012, 43, 695–710. [Google Scholar] [CrossRef]
- Johnson, P.S. Bur oak. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Qian, P.; Schoenau, J.J.; Huang, W.Z. Use of ion exchange membranes in routine soil testing. Commun. Soil Sci. Plant Anal. 1992, 23, 1791–1804. [Google Scholar] [CrossRef]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Mature hybrid poplar riparian buffers along farm streams produce high yields in response to soil fertility assessed using three methods. Sustainability 2013, 5, 1893–1916. [Google Scholar] [CrossRef]
- West, P. Tree and Forest Measurement; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Perron, J.-Y. Inventaire forestier. In Manuel de Foresterie; Les Presses de l’Université Laval: Ste-Foy, QC, Canada, 1996; pp. 390–473. [Google Scholar]
- Petersen, R.G. Design and Analysis of Experiments; Marcel-Dekker: New York, NY, USA, 1985. [Google Scholar]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Day, R.W.; Quinn, G.P. Comparisons of treatments after an analysis of variance in ecology. Ecol. Monogr. 1989, 59, 433–463. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Ellison, A.M. A Primer of Ecological Statistics; Sinauer Associated, Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Bentrup, G. Conservation Buffers: Design Guidelines for Buffers, Corridors, and Greenways; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2008. [Google Scholar]
- Périé, C.; Munson, A.D. Ten-year responses of soil quality and conifer growth to silvicultural treatments. Soil Sci. Soc. Am. J. 2000, 64, 1815–1826. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W. Sulfur cycling in forests. Biogeochemie 1984, 1, 29–43. [Google Scholar] [CrossRef]
- Sæbø, A.; Fløistad, I.S.; Netland, J.; Skúlason, B.; Edvardsen, Ø.M. Weed control measures in Christmas tree plantations of Abies nordmanniana and Abies lasiocarpa on agricultural land. New For. 2009, 38, 143–156. [Google Scholar] [CrossRef]
- Laungani, R.; Knops, J.M.H. Species-driven changes in nitrogen cycling can provide a mechanism for plant invasions. Proc. Natl. Acad. Sci. USA 2009, 106, 12400–12405. [Google Scholar] [CrossRef] [PubMed]
- Paris, P.; Cannata, F.; Olimpieri, G. Influence of alfalfa (Medicago sativa L.) intercropping and polyethylene mulching on early growth of walnut (Juglans spp.) in central Italy. Agrofor. Syst. 1995, 31, 169–180. [Google Scholar] [CrossRef]
- Von Althen, F.W. Revitalizing a black walnut plantation through weed control and fertilization. For. Chron. 1985, 61, 71–74. [Google Scholar] [CrossRef]
- Garrett, H.E.; Jones, J.E.; Kurtz, W.B.; Slusher, J.P. Black walnut (Juglans nigra L.) agroforestry—Its design and potential as a land-use alternative. For. Chron. 1991, 67, 213–218. [Google Scholar] [CrossRef]
- Von Althen, F.W. Afforestation of former farmland with high-value hardwoods. For. Chron. 1991, 67, 209–212. [Google Scholar] [CrossRef]
- Aber, J.D.; Magill, A.; Boone, R.; Melillo, J.M.; Steudler, P. Plant and soil responses to chronic nitrogen additions at the Harvard forest, Massachusetts. Ecol. Appl. 1993, 3, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Schoettle, A.W. Role of phosphorus and nitrogen in photosynthetic and whole plant carbon gain and nutrient use efficiency in eastern white pine. Oecologia 1988, 77, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Steele, K.L.; Kabrick, J.M.; Dey, D.C.; Jensen, R.G. Restoring riparian forests in the Missouri Ozarks. North. J. Appl. For. 2013, 30, 109–117. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Multiple-use zoning model for private forest owners in agricultural landscapes: A case study. Forests 2015, 6, 3614–3664. [Google Scholar] [CrossRef]
- Danner, B.T.; Knapp, A.K. Growth dynamics of oak seedlings (Quercus macrocarpa Michx. and Quercus muhlenbergii Engelm.) from gallery forests: Implications for forest expansion into grasslands. Trees 2001, 15, 271–277. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.W.; Gilliom, R.J.; Ryberg, K.R. Pesticides in U.S. streams and rivers: Occurrence and trends during 1992–2011. Environ. Sci. Technol. 2014, 48, 11025–11030. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.P.; Antoniou, M.N.; Blumberg, B.; Carroll, L.; Colborn, T.; Everett, L.G.; Hansen, M.; Landrigan, P.J.; Lanphear, B.P.; Mesnage, R.; et al. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environ. Health 2016, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortier, J.; Messier, C. Are chemical or mechanical treatments more sustainable for forest vegetation management in the context of the TRIAD? For. Chron. 2006, 82, 806–818. [Google Scholar] [CrossRef]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Tröger, J.; Muñoz, K.; Frör, O.; Schaumann, G.E. Plastic mulching in agriculture. Trading short-term agronomic benefits for long-term soil degradation? Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.J.; Liu, W.; Wang, E.; Zhou, T.; Xin, P. Residual plastic mulch fragments effects on soil physical properties and water flow behavior in the Minqin Oasis, Northwestern China. Soil Tillage Res. 2017, 166, 100–107. [Google Scholar] [CrossRef]
- Yan, C.; Liu, E.; Shu, F.; Liu, Q.; Liu, S.; He, W. Review of agricultural plastic mulching and its residual pollution and prevention measures in China. J. Agric. Res. Environ. 2014, 31, 95–102. [Google Scholar]
- Knight, K.W.; Schultz, R.C.; Mabry, C.M.; Isenhart, T.M. Ability of remnant riparian forests, with and without grass filters, to buffer concentrated surface runoff. JAWRA 2010, 46, 311–322. [Google Scholar]
- Hood, W.G.; Naiman, R.J. Vulnerability of riparian zones to invasion by exotic vascular plants. Plant Ecol. 2000, 148, 105–114. [Google Scholar] [CrossRef]
- Pysek, P.; Prach, K. Plant invasions and the role of riparian habitats: A comparison of four species alien to central Europe. J. Biogeogr. 1993, 20, 413–420. [Google Scholar] [CrossRef]
- Maillard, É.; Paré, D.; Munson, A.D. Soil carbon stocks and carbon stability in a twenty-year-old temperate plantation. Soil Sci. Soc. Am. J. 2010, 74, 1775–1785. [Google Scholar] [CrossRef]
- Rolando, A.C.; Baillie, R.B.; Thompson, G.D.; Little, M.K. The risks associated with glyphosate-based herbicide use in planted forests. Forests 2017, 8. [Google Scholar] [CrossRef]
- Environmental Protection Authority (EPA) of South Australia. Safe and Effective Herbicide Use—A Handbook for Near-Water Applications; EPA: Adelaide (SA), Australia, 2017.
- Sweeney, B.W.; Czapka, S.J.; Petrow, L.C.A. How planting method, weed abatement, and herbivory affect afforestation success. South J. Appl. Ecol. 2007, 31, 85–92. [Google Scholar]
Figure 1.
Schematic representation of an experimental plot for a single tree species (4.5 m × 4.5 m) that received a vegetation treatment (herbicide or plastic mulch). The plot is located in the floodplain zone and it contains nine trees (white circles) and a single vegetation treatment applied on each row on a 1.2 m wide strip (grey rectangles). Spacing between trees is 1.5 m × 1.5 m. Small black rectangles indicate the positioning of each pair of Plant Root Simulator (PRS)-probes ion exchange membranes. Hybrid willows (white diamonds) were planted between the top and toe of the streambank, outside the experimental plot.
Figure 1.
Schematic representation of an experimental plot for a single tree species (4.5 m × 4.5 m) that received a vegetation treatment (herbicide or plastic mulch). The plot is located in the floodplain zone and it contains nine trees (white circles) and a single vegetation treatment applied on each row on a 1.2 m wide strip (grey rectangles). Spacing between trees is 1.5 m × 1.5 m. Small black rectangles indicate the positioning of each pair of Plant Root Simulator (PRS)-probes ion exchange membranes. Hybrid willows (white diamonds) were planted between the top and toe of the streambank, outside the experimental plot.

Figure 2.
Species × Vegetation treatment interaction for soil NO3 supply rate (p = 0.11) measured for 30 days (21 June to 21 July) of the fifth growing season in an agricultural riparian buffer. Vertical bars represent standard error of the difference (SED).
Figure 2.
Species × Vegetation treatment interaction for soil NO3 supply rate (p = 0.11) measured for 30 days (21 June to 21 July) of the fifth growing season in an agricultural riparian buffer. Vertical bars represent standard error of the difference (SED).

Figure 3.
Species effect on survival rate (p = 0.05) in an agricultural riparian buffer. Vertical bars represent standard error of the difference (SED).
Figure 3.
Species effect on survival rate (p = 0.05) in an agricultural riparian buffer. Vertical bars represent standard error of the difference (SED).

Figure 4.
Stem volume per tree measured after 2, 3, 4, and 5 years of growth in an agricultural riparian buffer. The Species × Vegetation treatment × Time interaction is significant at p < 0.05 according to multivariate analysis of variance (MANOVA).
Figure 4.
Stem volume per tree measured after 2, 3, 4, and 5 years of growth in an agricultural riparian buffer. The Species × Vegetation treatment × Time interaction is significant at p < 0.05 according to multivariate analysis of variance (MANOVA).

Figure 5.
Species × Vegetation treatment interaction for growth variables measured at the end of the fifth growing season in an agricultural riparian buffer: (a) total height (p < 0.05), (b) diameter at breast height (DBH) (p < 0.05), and (c) stem volume per tree (p < 0.01). Vertical bars represent standard error of the difference (SED).
Figure 5.
Species × Vegetation treatment interaction for growth variables measured at the end of the fifth growing season in an agricultural riparian buffer: (a) total height (p < 0.05), (b) diameter at breast height (DBH) (p < 0.05), and (c) stem volume per tree (p < 0.01). Vertical bars represent standard error of the difference (SED).

Figure 6.
Negative relationship between soil NO3 supply rate and stem volume growth after five growing seasons for white pine in the black plastic mulch treatment (n = 6). The solid line represents a linear least-square regression.
Figure 6.
Negative relationship between soil NO3 supply rate and stem volume growth after five growing seasons for white pine in the black plastic mulch treatment (n = 6). The solid line represents a linear least-square regression.

Figure 7.
During the fifth growing season (June 2014), a dense herbaceous vegetation cover is observed underneath black walnut in the control treatment (a) and in the herbicide treatment (b). Permanent black plastic mulch treatment (c) restricts herbaceous vegetation growth over the long-term (photo taken after mulch installation around white pines in May 2010).
Figure 7.
During the fifth growing season (June 2014), a dense herbaceous vegetation cover is observed underneath black walnut in the control treatment (a) and in the herbicide treatment (b). Permanent black plastic mulch treatment (c) restricts herbaceous vegetation growth over the long-term (photo taken after mulch installation around white pines in May 2010).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Ecological characteristics of the studied species.
| Common Name | Habitat Range | Site Fertility Class | Early Growth | Shade Tolerance | N-Form Preference | Successional Status | Flood Tolerance |
|---|---|---|---|---|---|---|---|
| Black walnut 1 | Well-drained bottomlands | High | Moderatly fast | Low | NH4 | Early | Low |
| Bur oak 1 | Bottomlands, riparian zones, and dry calcareous sites | High | Slow to moderate | Low to intermediate | NO3:NH4 | Early | Intermediate |
| White pine 1 | All types of sites ranging from rocky hill top to peatland | Low to moderate | Slow to moderate | Intermediate | NH4 | Mid | Low |
Table 2.
A priori contrasts for the analysis of variance (ANOVA) testing the effects of Species and Vegetation treatment on soil NO3 supply rate measured during the fifth growing season.
Table 2.
A priori contrasts for the analysis of variance (ANOVA) testing the effects of Species and Vegetation treatment on soil NO3 supply rate measured during the fifth growing season.
| Contrast | Species | Treatment 2 | Black Walnut | Bur Oak | White Pine | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | ||||
| (1) | NH4 pref. 1 vs. oak | Mulch | 0 | 0 | −1 | 0 | 0 | 2 | 0 | 0 | −1 | <0.01 |
| (2) | All | Mulch vs. Herb. | 0 | 1 | −1 | 0 | 1 | −1 | 0 | 1 | −1 | <0.01 |
| (3) | All | Herb. vs. Cont. | 1 | −1 | 0 | 1 | −1 | 0 | 1 | −1 | 0 | 0.37 |
1 Species having preference for NH4 uptake are white pine and black walnut. 2 Mulch = black plastic mulch; Herb. = Herbicide; Cont. = Control.
Table 3.
Vegetation treatment effect on the supply rate (µg/10 cm2/30 days) of various soil elements measured during the fifth growing season in an agricultural riparian buffer.
Table 3.
Vegetation treatment effect on the supply rate (µg/10 cm2/30 days) of various soil elements measured during the fifth growing season in an agricultural riparian buffer.
| Treatment | NO3 | NH4 | P | Ca | K | Mg | S |
|---|---|---|---|---|---|---|---|
| Control | 66.9 | 3.23 | 9.13 | 2049 | 201.0 | 434.4 | 53.9 |
| Herbicide | 87.6 | 2.67 | 7.72 | 2161 | 149.5 | 380.6 | 52.0 |
| Plastic mulch | 165.8 | 2.58 | 12.69 | 2292 | 175.4 | 375.4 | 83.4 |
| SED 1 | 22.7 | 0.35 | 2.35 | 95 | 60.7 | 36.2 | 10.0 |
| p-value | <0.001 | 0.13 | 0.11 | <0.05 | 0.70 | 0.21 | <0.01 |
1 SED: standard error of the difference.
Table 4.
A priori contrasts for the ANOVA testing the effects of Species and Vegetation treatment on stem volume after 5 years.
Table 4.
A priori contrasts for the ANOVA testing the effects of Species and Vegetation treatment on stem volume after 5 years.
| Contrast | Species | Treatment 1 | Black Walnut | Bur Oak | White Pine | p-Values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | ||||
| (1) | B. walnut | Veg. vs. Cont. | 2 | −1 | −1 | 0 | 0 | 0 | 0 | 0 | 0 | <0.001 |
| (2) | B. walnut | Herb. vs. Mulch | 0 | −1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0.55 |
| (3) | B. oak | Veg. vs. Cont. | 0 | 0 | 0 | 2 | −1 | −1 | 0 | 0 | 0 | <0.01 |
| (4) | B. oak | Herb. vs. Mulch | 0 | 0 | 0 | 0 | −1 | 1 | 0 | 0 | 0 | 0.65 |
| (5) | W. pine | Veg. vs. Cont. | 0 | 0 | 0 | 0 | 0 | 0 | 2 | −1 | −1 | 0.25 |
| (6) | W. pine | Herb. vs. Mulch | 0 | 0 | 0 | 0 | 0 | 0 | 0 | −1 | 1 | 0.54 |
1 Veg. = Vegetation treatments; Herb. = Herbicide; Cont. = Control.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers? Forests 2018, 9, 258. https://doi.org/10.3390/f9050258
AMA Style
Truax B, Fortier J, Gagnon D, Lambert F. Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers? Forests. 2018; 9(5):258. https://doi.org/10.3390/f9050258
Chicago/Turabian StyleTruax, Benoit, Julien Fortier, Daniel Gagnon, and France Lambert. 2018. "Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers?" Forests 9, no. 5: 258. https://doi.org/10.3390/f9050258
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.