Building Resistance and Resilience: Regeneration Should Not be Left to Chance

1
Wildland Resources and Ecology Center, Utah State University, Logan, UT 84322, USA
2
Department of Forest Resources, University of Minnesota, St. Paul, MN 55108, USA
3
USDA Forest Service, Forest Inventory and Analysis, Rocky Mountain Research Station, Ogden, UT 84401, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(5), 270; https://doi.org/10.3390/f9050270
Submission received: 4 April 2018
/
Revised: 1 May 2018
/
Accepted: 14 May 2018
/
Published: 16 May 2018
(This article belongs to the Special Issue The Forest Renewal Issue: New Problems and Findings in the Establishment of Tree Regeneration)
Abstract
:Contemporary forest planning has tasked managers with developing goals associated with resistance and resilience. In practice, silviculturists use forest structure and tree species composition to characterize goals and desired future conditions, write prescriptions, and monitor outcomes associated with resistance and resilience. Although rarely discussed in the exploding literature relating to forest resistance and resilience, silvicultural regeneration methods are important and underutilized tools to meet these goals. We propose alternative silvicultural systems for building resistance and resilience to two common large-scale bark beetle disturbance agents in the Intermountain West, United States: mountain pine beetle (Dendroctonus ponderosae Hopkins) and spruce beetle (Dendroctonus rufipennis Kirby). Shelterwood, and shelterwood-with-reserves, silvicultural systems provide the desirable facilitative characteristics of a mature overstory on maintaining advance reproduction and the establishment of new cohorts of desirable tree species. These also allow the timely regeneration of large treatment areas necessary to rapidly promote desired future conditions in the face of inevitable disturbance. When implemented proactively, regeneration treatments allow silviculturists to take advantage of currently existing vegetation for the creation of age class and tree species diversity. In general, these examples illustrate the need for proactive planning for regeneration in response to any disturbance where desired future conditions include particular species. Furthermore, we argue that timely silvicultural interventions that focus on regenerating trees may be a key factor in achieving goals relating to resilience to specific disturbance types. Waiting until after the disturbance has occurred could result in the lost opportunity to establish desired species composition or stand structure—and may well result in a considerable restoration challenge.
1. Introduction
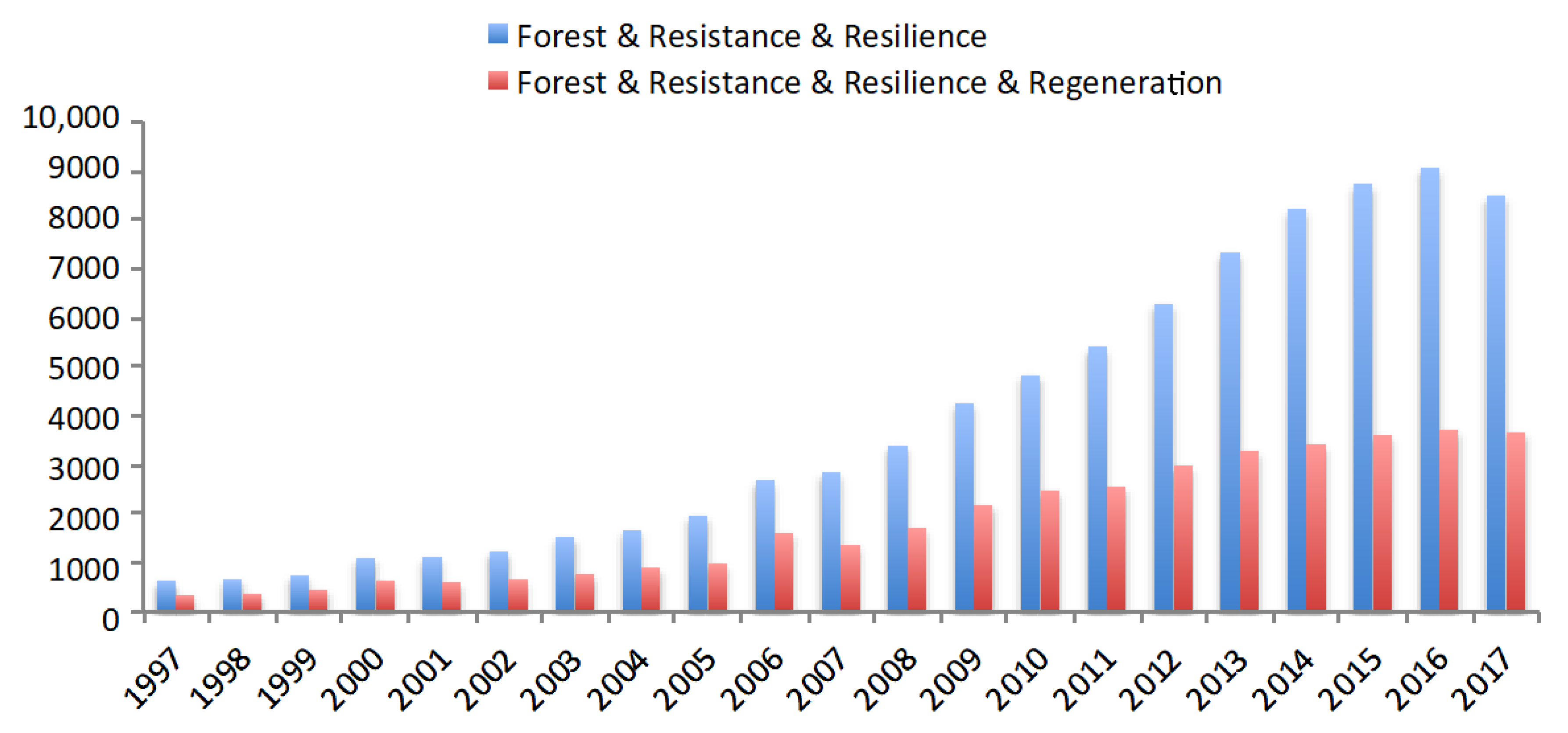
An emerging theme across ecological, policy, and management boundaries is the goal to build resistance and/or resilience to current and future disturbances [1]. Indeed, the number of papers referencing resistance and resilience in forest systems has increased more than 10-fold over the last two decades (Figure 1, Table 1). In forest management, this increased attention has been motivated, in part, by changes in historical disturbance regimes, but also in part by the desire to be proactive and take advantage of conditions before disturbance events. For example, in dry forests in the Western United States that have long been subject to fire exclusion, frequent low-severity fire regimes are being replaced by infrequent high-severity fires [2]. Similarly, there are widespread changes in the frequency, extent, and severity of bark beetle outbreaks [3,4]. Changes in disturbance regimes across Western North America have been attributed to combinations of interactions in land management and climate [5], and this is having cascading effects on the societal perception of forest disturbances, especially fire [6], on forest policy, and on forestland management. As a result, multiple descriptions of resistance and resilience abound in the literature (Table 1). The development of multiple “definitions” or sub-definitions of resistance and resilience increases confusion surrounding the terms, especially when authors do not explicitly identify the definitions they are using. In addition, many of the definitions suffer from translation difficulties when attempting to incorporate them into management action, such as silvicultural treatments/systems focused on regeneration. While forest thinning is a common prescription for building resistance [2,5], building resilience to some specific disturbances requires implementation of appropriate regeneration methods. In the short-term, thinning may also increase risk of windthrow [7].
The process of regeneration, whether focused on promoting or maintaining advance reproduction (trees currently existing in the understory), or the establishment of a new cohort of seedlings (regenerated trees), is a key but sometimes overlooked component of a silvicultural system. It is at this time that the silviculturist can fundamentally shape a stand’s future. A desired future condition (DFC) allows broad stand and landscape level goals and objectives to be translated into quantifiable metrics of structure and composition. These DFCs can then be used to measure the success of silvicultural treatments against the goals.
When the DFCs include resistance and/or resilience, managers can use a conceptual framework (Table 1 in [19]) developed to specifically characterize these goals in terms of structure and composition. Instead of unnecessarily redefining the terms resistance and resilience (Table 1), the conceptual model provides a practical framework for the evaluation and manipulation intended to accomplish objectives that may relate to resistance and resilience at both the stand- and landscape-level. In the DeRose and Long [19] framework, stand resistance is characterized as the influence of structure and composition on the nature or behavior of a particular disturbance (e.g., severity of a bark beetle outbreak). Stand resilience, on the other hand, is characterized as the influence of a specific disturbance on subsequent, post-disturbance, vegetation structure and composition. In this framework, landscape resistance and resilience are similarly characterized in terms of the influence of structure and composition on disturbance (resistance) and the influence of a disturbance of subsequent structure and composition (resilience). These characterizations provide a way to frame resistance/resilience goals and objectives in practical silvicultural terms. For example, resilience must be characterized in terms of desired post-disturbance structure and composition. This will often put specific focus on regeneration and the regeneration environment of the future stand or landscape.
In order to explore the development of silviculturally meaningful objectives relating to resistance and resilience, we examine how traditional and alternative silvicultural systems influence overstory and regeneration dynamics in the context of two different bark beetle species in the Intermountain West, United States. The first example involves spruce beetle (Dendroctonus rufipennis Kirby) in spruce-fir (Picea engelmannii Perry ex Engelm. and Abies lasiocarpa (Hook.) Nutt.) stands and landscapes. The second focuses on mountain pine beetle (Dendroctonus ponderosae Hopkins) in lodgepole pine (Pinus contorta var. latifolia Engelm.) stands and landscapes. For both of these bark beetle-forest systems, we examine the influence of structure and composition on the likelihood of successful bark beetle attacks; i.e., resistance (sensu DeRose and Long [19]). We also examine likely post-outbreak structure and composition in light of alternative DFCs; i.e., resilience (sensu DeRose and Long [19]). If the DFCs are to maintain either Engelmann spruce or lodgepole pine, timely silvicultural treatments focused on regenerating the desired species will be key to building resilience to spruce beetle and mountain pine beetle outbreaks.
2. Methods
Using existing models, we assess resistance of Engelmann spruce and lodgepole pine stands to spruce beetle and mountain pine beetle, respectively. Previous work has identified important elements of stand tree species composition and structure associated with susceptibility to bark beetle attack [22,23]. In both systems (Engelmann spruce and lodgepole pine), susceptibility to bark beetle attack increases with increasing tree size, density, and percentage of the host species.
In the context of a conceptual framework [19], we examine alternative ways to build resilience to bark beetle attack. Stand-specific objectives of resilience are characterized in terms of composition and structure. We than examine how conventional and alternative silvicultural systems might accomplish resilience-based objectives.
3. Results and Discussion
3.1. Resistance to the Bark Beetles
Spruce beetle and mountain pine beetle are native bark beetles (Dendroctonus spp.) that under endemic conditions play an important role in the stand dynamics of spruce-fir and lodgepole pine stands, respectively. However, as populations of both bark beetle species have recently and dramatically transitioned from endemic to epidemic levels there has been increased interest in the impacts of epidemic beetle populations [3,5]. There have been numerous recent studies that have explored the variable impacts of epidemic beetle populations on subsequent forest conditions in both spruce-fir [24] and lodgepole pine stands [25,26,27] (among others). An observation following beetle outbreaks is that maintenance of the pre-disturbance forest type is not guaranteed.
Active management to build resistance or resilience to anticipated bark beetle disturbances requires practical assessment of hazard or susceptibility. At the stand-level, metrics of structure and composition that characterize stand hazard are fairly well-established for spruce beetle (Table 2). In this context, stand hazard represents susceptibility to spruce beetles and is, essentially the inverse of resistance. In contrast, ‘risk’ is relative hazard in the face of beetle pressure. A stand with high hazard (limited resistance) may have limited risk when beetle population numbers are low; however, it is in these high hazard stands that beetle populations are likely to build. Historically, downed trees (e.g., windthrow, logging debris) have been implicated as catalysts for building spruce beetle populations in susceptible stands [22], and recent literature suggests warming temperatures may facilitate building beetle populations [28].
At the stand-level, metrics for structure and composition are not as well-established for mountain pine beetle (e.g., [29]). This may be due, in part, to the fact that mountain pine beetle has a number of host pine species in addition to lodgepole pine (e.g., ponderosa pine (P. ponderosa Lawson & C. Lawson), limber pine (P. flexilis James), whitebark pine (P. albicaulis Engelm.), sugar pine (P. lambertiana Douglas), and western white pine (P. monticola Douglas ex D. Don). However, for lodgepole pine stands, there is general consensus that resistance is inversely related to the abundance of the host, host tree diameter, and total stand relative density (Table 3; [30,31]).
Resistance, however, has its limits. There is ample evidence for successfully building stand and landscape resistance to different types of disturbances, for example wind and fire (e.g., [32,33]). While manipulating stand structure and composition so as to build resistance to bark beetle attack is certainly feasible [31], there is considerable evidence that it will ultimately fail in the face of overwhelming beetle numbers [19,31,34,35,36]. Indeed, building stand resistance to bark beetle attacks may not be a good use of management effort and limited resources unless explicitly used to further a resilience goal. Short-term resistance can, however, buy time in which desirable regeneration can be established, and this may be critical to building post-outbreak resilience [19,34].
3.2. Resilience of Spruce-Fir to Spruce Beetle Outbreaks
If it is to be a practical, achievable forest management goal, resilience must be characterized, and evaluated, in terms of the desired (or at least acceptable) post-outbreak composition and structure. The mature Engelmann spruce stand illustrated in Figure 2 clearly lacks resistance to spruce beetle attacks—it has high relative density and consists almost entirely of large diameter host trees. Even more importantly, this stand also lacks resilience to spruce beetle attack—there are no mature non-host tree species and advance reproduction of any tree species is extremely limited. If the desired future condition includes anything besides non-forest, then this stand ought to be a candidate for treatment. Any plan to build resilience requires explicit identification of the priorities for post-outbreak composition and structure. If mature living trees are a priority, then a pre-outbreak stand must have overstory species diversity, including, one or more of the several non-host tree species commonly associated with Engelmann spruce [37]. Absent of such overstory species diversity (e.g., Figure 2), an objective of resilience represented by mature trees is unrealistic in the short-term. Indeed, building overstory species diversity would take many decades and can be an important long-term goal, but will be unachievable in the short-term.
In many cases, building resilience to spruce beetle attack will depend on pre-disturbance regeneration. In some stands, this might be represented by advance reproduction established well ahead of the anticipated spruce beetle outbreak. For stands where advance reproduction is absent (e.g., Figure 2) or does not include the desired species (e.g., Engelmann spruce), building resilience requires timely management. It is at this stage where the silviculturist can effectively plan for the future by promoting advance reproduction of spruce or other, non-host, tree species. If increased tree species diversity is a future goal, selecting a diverse planting mix could also aid in reaching that future goal.
In the absence of structure and composition indicative of resilience (i.e., overstory species diversity or advance reproduction), passively waiting for what may be an inevitable spruce beetle outbreak represents an important lost opportunity. There are extensive examples of the cost and uncertainty of restoration in diverse systems including oak-savannas across the Great Plains [38], and floodplain forests along the Mississippi River [39]. Having to start “from scratch”, post-outbreak, (i.e., reacting to the outbreak) presents a restoration challenge that squanders the considerable opportunity that extant forests represent for the management of desired future structure and composition.
Ideally, resilience will be a legacy of a long history of management. In the Intermountain West, the conventional silvicultural system for Engelmann spruce is small group selection with a 20–30 year cutting cycle (Table 4 [40]). This system has been favored because it is thought to mimic gap-phase dynamics and typically results in successful natural regeneration of spruce. After several cutting cycles, the resulting structural and compositional diversity offers considerable resilience to a spruce beetle outbreak, represented by multiple age-classes and tree species diversity [34]. However, for stands in which it is currently lacking, the timely building of resilience with the small group selection system is constrained by the length of the cutting cycle and the small percentage of total area regenerated per entry.
Silvicultural systems incorporating larger openings, such as large group selection and clearcutting, are inappropriate for the regeneration of Engelmann spruce in the Intermountain West because of the need for facilitation during establishment [40]. An alternative silvicultural system combining the required faciliatation and the potential for timely establishment of regeneration, is a shelterwood or shelterwood-with-reserves (Table 4 [37]). In Northern Utah, natural regeneration of Engelmann spruce beneath a uniform shelterwood may still be protracted over decades because of irregular cone crops and undependable summer precipitation [40]. Underplanting beneath the shelterwood is a way to quickly establish Engelmann spruce regeneration. The establishment of an essentially even-aged cohort of Engelmann spruce will, of course, not ensure resilience to spruce beetle in the long-term; as these trees mature they will become susceptible to beetle attack [35]. In addition to Engelmann spruce, the planting prescription could also include additional non-host species such as Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco), limber pine, white fir (Abies concolor (Gord. & Glend.) Lindl. ex Hildebr) and aspen (Populus tremuloides Michx.) These species are adapted to warmer, drier conditions than Engelmann spruce and their inclusion in the planting mix may provide a hedge against climate change and further enhance future resistance and resilience to spruce beetle attack by increasing species diversity.
3.3. Resilience of Lodgepole Pine to Mountain Pine Beetle Outbreaks
Given their typical stand dynamics, lodgepole pine stands have limited resilience to mountain pine beetle outbreaks. Resilience will normally be restricted to either very young or very old stands. An immature stand will have both resistance and resilience if it consists of trees so small (e.g., dbh < 15 cm) that even epidemic population levels of mountain pine beetle will not attack. In contrast, a typical old stand will have an overstory dominated by large lodgepole pine and a well-developed understory of non-host species; e.g., subalpine fir. Such a stand will have very limited resistance to mountain pine beetle but can still be considered resilient if the composition represented by the advance reproduction is acceptable; i.e., no lodgepole pine, only subalpine fir. Building resilience that will include a lodgepole pine component requires planning ahead—waiting until a mountain pine beetle epidemic is underway is too late.
A clearcutting system has conventionally been used for the management of lodgepole pine in the Rocky Mountains (Table 4 [40]). Clearfelling of the overstory would typically be followed by site preparation, either by mechanical or prescribed fire. Natural regeneration of lodgepole pine is often abundant given an adequate seed source represented by either serotinous or nonserotinous cones. Because of the over-abundance of natural regeneration, this silvicultural system will usually include precommercial thinning and sometimes a commercial thinning [40]. However due to policies regarding threatened and endangered species in some areas precommercial thinning is prohibited [42]. Clearcutting and, often intense, site preparation effectively eliminates any advance reproduction and the result is nearly pure even-aged stands of lodgepole pine. Thus, except for a limited window very early in the rotation, when the regenerating pine is still too small to be susceptible to mountain pine beetle attack, this silvicultural system offers essentially no resilience to a mountain pine beetle outbreak. Additionally, when considering landscape-level resistance and resilience, large homogenous lodgepole forests in similar structural conditions can cause mountain pine beetle populations to rapidly increase and spill over into “more resistant” stands (Figure 3).
Alternative silvicultural systems have the potential for building long-term resilience to mountain pine beetle outbreaks. Here we describe the implementation of, and early results from, a shelterwood-with-reserves system in lodgepole pine (Table 4). The seed cutting, a combination of strip and uniform shelterwood, was conducted in 1991. The lodgepole pine overstory was about 140-years-old; there was some aspen in the overstory and the understory consisted of abundant advance reproduction of subalpine fir. Except for scarification associated with harvesting, there was no site preparation. The objectives of the treatment were to capture a new cohort of lodgepole pine and to retain advance reproduction of subalpine fir, a non-host of mountain pine beetle. An additional cohort of non-host trees was added with an enrichment planting of Engelmann spruce [42]. The enrichment planting was necessary because there was no proximate seed source for Engelmann spruce. The role of the retained overwood (approximately 170 large lodgepole pine per hectare) was to limit the abundance of natural regeneration of lodgepole pine to densities that would ideally eliminate the need for a future precommercial thinning and to protect the shade tolerant subalpine fir and Engelmann spruce. Some of the overwood was retained beyond the regeneration period for structural diversity and, therefore, the system can best be characterized as a shelterwood-with-reserves.
Approximately 25 years post-treatment, the structure and composition of the stand reflects considerable resilience to a mountain pine beetle outbreak (Figure 4). If an outbreak were to occur, high beetle numbers entering the stand from outside would almost certainly successfully attack the large lodgepole pine reserve trees. Because of pre-outbreak intervention, however, this stand has young, currently non-susceptible lodgepole pine and an abundance of non-host trees, including aspen, subalpine fir, and Engelmann spruce. The lodgepole pine saplings will eventually become susceptible to mountain pine beetle, but the considerable tree species diversity will continue to provide resilience if this structure and composition is consistent with broad management objectives. Depending on the management context, a particular characterization of resilience to a mountain pine beetle outbreak might be focused on a desired ecosystem process or function. For example, research in Colorado suggests considerable nitrate retention in mountain pine beetle-infested watersheds can result from residual vegetation components unaffected by beetles [43].
This type of silvicultural system represents a shift in thinking regarding lodgepole pine management intended to build resilience to mountain pine beetle outbreaks. It is not appropriate everywhere but can create increased heterogeneity at the stand- and landscape-levels when combined with more traditional silvicultural systems.
4. Conclusions
Resistance and resilience are increasingly important concepts in developing forest stand and landscape management goals and objectives. To have management utility, objectives related to resistance and resilience must be framed in terms of structure and composition specific to a given disturbance [19]. When defining the objectives, it is imperative to focus not only on the current stand conditions but also the DFCs and the silvicultural systems, including regeneration methods, that will be required to meet the DFCs. For example, elements of stand structure and composition associated with resistance to bark beetle (e.g., Dendroctonus spp.) attacks are fairly well understood and can be manipulated silviculturally. However, treatments developed for individual stands to build resistance (whether to spruce beetle or mountain pine beetle attacks) focused on the manipulation of overstory structure and composition—are likely to fall short of meeting this goal once beetles reach epidemic levels [35]. However, treatments for resistance can also create an appropriate regeneration environment of the desired species to provide longer-term resilience.
No forest can be resilient to all disturbances and an ambiguous goal of ‘resilience’ is of little help in developing a practical plan for actually achieving it. For some important disturbances, such as spruce beetle and mountain pine beetle epidemics, building resilience will typically require the release of already established advanced reproduction or, in its absence, the regeneration of a new cohort. For other disturbances, e.g., low-severity fire regimes, the focus on establishing new age classes, or tree species diversity via regeneration treatments may be less desirable. Regardless, if the accomplishment of a resilience objective requires establishment of a new cohort of desirable species, it is much better to be proactive. Waiting until after a high-severity disturbance may well result in a considerable restoration challenge.
The focus on measurable post-disturbance metrics highlights the importance of regeneration. Regeneration methods may be overlooked in planning for resistance and resilience but can be a key component in developing desired post-disturbance conditions, especially where particular species are desired. There should be considerable thought given to regeneration in any silviculture prescription, but especially in seasonally dry regions such as the Intermountain West because regeneration survival is not guaranteed. Extended summer drought can cause high mortality in both artificially and naturally regenerating seedlings. Managers should focus on how treatments will influence the regeneration environment to ensure DFCs that may include goals relating to resistance and resilience are met. Furthermore, being proactive, instead of reactive, is of the utmost importance—this will allow silviculturists to take advantage of currently existing vegetation to facilitate the establishment of desirable regeneration in new age classes and, where appropriate, tree species diversity. Finally, the explicit inclusion of targeted species diversity into the regeneration plan could represent bet-hedging with respect to uncertainty relating to future climates. These bark beetle examples illustrate the need to plan for regeneration. There are numerous other disturbance regimes (insect, disease, wind, drought, etc.) where proactive planning for timely regeneration (particularly in previously unmanaged stands) will be central to meeting goals related to resistance and/or resilience in different forest systems.
Author Contributions
J.N.L. conceived the study; J.N.L., R.J.D. and M.W.C. analyzed the data and wrote the paper.
Funding
This research was supported in part by the Utah Agricultural Experiment Station, Utah State University, and approved as journal paper number 9018. This work was also supported by Minnesota Agriculture Experimentation Station (Project MIN-42-100).
Acknowledgments
This paper was prepared in part by an employee of the US Forest Service as part of official duties and is therefore in the public domain.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.T.; McCaffrey, S.; Jain, T.B. Science Basis for Changing Forest Structure to Modify Wildfire Behavior and Severity; General Technical Report RMRS-GTR-120; USDA Forest Service: Ogden, UT, USA, 2004. [Google Scholar]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Jenkins, M.J.; Runyon, J.B.; Fettig, C.J.; Page, W.G.; Bentz, B.J. Interactions among the Mountain Pine Beetle, Fires, and Fuels. For. Sci. 2014, 60, 489–501. [Google Scholar] [CrossRef]
- Fettig, C.J.; Reid, M.L.; Bentz, B.J.; Sevanto, S.; Spittlehouse, D.L.; Wang, T. Changing climates, changing forests: A Western North American perspective. J. For. 2013, 111, 214–228. [Google Scholar] [CrossRef]
- Morris, J.L.; Cottrell, S.; Fettig, C.J.; Hansen, W.D.; Sherriff, R.L.; Carter, V.A.; Clear, J.L.; Clement, J.; DeRose, R.J.; Hicke, J.A.; et al. Managing bark beetle impacts on ecosystems and society: Priority questions to motivate future research. J. Appl. Ecol. 2017, 54, 750–760. [Google Scholar] [CrossRef]
- Yücesan, Z.; Özçelik, S.; Oktan, E. Effects of thinning on stand structure and tree stability in an afforested oriental beech (Fagus orientalis Lipsky) stand in northeast Turkey. J. For. Res. 2015, 26, 123–129. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Shulze, P.C. Engineering within Ecological Constraints; National Academy Press: Washington, DC, USA, 1996; p. 224. [Google Scholar]
- Peterson, G.; Allen, C.R.; Holling, C.S. Ecological resilience, biodiversity, and scale. Ecosystems 1998, 1, 6–18. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological resilience—in theory and application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Carpenter, S.; Walker, B.; Anderies, J.M.; Abel, N. From metaphor to measurement: Resilience of what to what? Ecosystems 2001, 4, 765–781. [Google Scholar] [CrossRef]
- Holling, C.S.; Gunderson, L.H.; Perterson, G.D. Resilience and adaptive cycles. In Panarchy: Understanding Transformations in Human and Natural Systems; Holling, C.S., Gunderson, L.H., Eds.; Island Press: Washington, DC, USA, 2002; pp. 25–62. [Google Scholar]
- Walker, B.; Carpenter, S.; Anderies, J.; Abel, N.; Cumming, G.S.; Janssen, M.; Lebel, L.; Nordberg, J.; Peterson, G.D.; Pritchard, R. Resilience management in social-ecological systems: A working hypothesis for a participatory approach. Conervation Ecol. 2002, 6, 14. [Google Scholar] [CrossRef]
- Gallopín, G.C. Linkages between vulnerability, resilience, and adaptive capacity. Glob. Environ. Change 2006, 16, 293–303. [Google Scholar] [CrossRef]
- Benson, M.H.; Garmestani, A.S. Can We Manage for Resilience? The Integration of Resilience Thinking into Natural Resource Management in the United States. Environ. Manage. 2011, 48, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Peterson, D.L.; Millar, C.I.; Joyce, L.A.; Furniss, M.J.; Halofsky, J.E.; Neilson, R.P.; Morelli, T.L. Responding to Climate Change in National Forests: A Guidebook for Developing Adaptation Options; General Technical Report PNW-GTR-855; United States Department of Agriculture: Portland, OR, USA, 2011.
- Churchill, D.J.; Larson, A.J.; Dahlgreen, M.C.; Franklin, J.F.; Hessburg, P.F.; Lutz, J.A. Restoring forest resilience: From reference spatial patterns to silvicultural prescriptions and monitoring. For. Ecol. Manag. 2013, 291, 442–457. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Resistance and Resilience: A Conceptual Framework for Silviculture. For. Sci. 2014, 60, 1205–1212. [Google Scholar] [CrossRef]
- Messier, C.; Puettmann, K.; Chazdon, R.; Andersson, K.P.; Angers, V.A.; Brotons, L.; Filotas, E.; Tittler, R.; Parrott, L.; Levin, S.A. From Management to Stewardship: Viewing Forests as Complex Adaptive Systems in An Uncertain World. Conserv. Lett. 2014, 8, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Nagel, L.M.; Palik, B.J.; Battaglia, M.A.; D’Amato, A.W.; Guldin, J.M.; Swanston, C.W.; Janowiak, M.K.; Powers, M.P.; Joyce, L.A.; Millar, C.I.; et al. Adaptive Silviculture for Climate Change: A National Experiment in Manager-Scientist Partnerships to Apply an Adaptation Framework. J. For. 2017, 115, 167–178. [Google Scholar] [CrossRef]
- Schmid, J.M.; Frye, R.H. Stand Ratings for Spruce Beetles; United States Department of Agriculture: Fort Collins, CO, USA, 1976.
- Randall, C.; Steed, B.; Bush, R. Revised R1 Forest Insect Hazard Rating System User Guide for Use with Inventory Data Stored in Fsveg and/or Analyzed with the Forest Vegetation Simulator; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Hart, S.J.; Veblen, T.T.; Mietkiewicz, N.; Kulakowski, D. Negative Feedbacks on Bark Beetle Outbreaks: Widespread and Severe Spruce Beetle Infestation Restricts Subsequent Infestation. PLoS ONE 2015, 10, e0127975. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.J.; Rhoades, C.C.; Hubbard, R.M.; Battaglia, M.A. Tree regeneration and future stand development after bark beetle infestation and harvesting in Colorado lodgepole pine stands. For. Ecol. Manag. 2011, 261, 2168–2175. [Google Scholar] [CrossRef]
- Kayes, L.J.; Tinker, D.B. Forest structure and regeneration following a mountain pine beetle epidemic in southeastern Wyoming. For. Ecol. Manag. 2012, 263, 57–66. [Google Scholar] [CrossRef]
- Hubbard, R.M.; Rhoades, C.C.; Elder, K.; Negron, J. Changes in transpiration and foliage growth in lodgepole pine trees following mountain pine beetle attack and mechanical girdling. For. Ecol. Manag. 2013, 289, 312–317. [Google Scholar] [CrossRef]
- DeRose, R.J.; Bentz, B.J.; Long, J.N.; Shaw, J.D. Effect of increasing temperatures on the distribution of spruce beetle in Engelmann spruce forests of the Interior West, USA. For. Ecol. Manag. 2013, 308, 198–206. [Google Scholar] [CrossRef]
- Bentz, B.J.; Amman, G.D.; Logan, J.A. A critical assessment of risk classification systems for the mountain pine beetle. For. Ecol. Manag. 1993, 61, 349–366. [Google Scholar] [CrossRef]
- Anhold, J.A.; Jenkins, M.J.; Long, J.N. Management of lodgepole pine stand density to reduce susceptibility to mountain pine beetle attack. West. J. Appl. For. 1996, 11, 50–53. [Google Scholar]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, A.S.; Nebeker, T.E.; Negron, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 25–53. [Google Scholar] [CrossRef]
- Drever, C.R.; Peterson, G.; Messier, C.; Bergeron, Y.; Flannigan, M. Can forest management based on natural disturbances maintain ecological resilience? Can. J. For. Res. 2006, 36, 2285–2299. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.J. The windthrow triangle: A relative windthrow hazard assessment procedure for forest managers. For. Chron. 1995, 71, 446–450. [Google Scholar] [CrossRef]
- Windmuller-Campione, M.A.; Page, D.H.; Long, J.N. Does the Practice of Silviculture Build Resilience to the Spruce Beetle? A Case Study of Treated and Untreated Spruce-Fir Stands in Northern Utah. J. For. 2017, 115, 559–567. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Factors influencing the spatial and temporal dynamics of Engelmann spruce mortality during a spruce beetle outbreak. For. Sci. 2012, 58, 1–14. [Google Scholar] [CrossRef]
- Temperli, C.; Hart, S.J.; Veblen, T.T.; Kulakowski, D.; Hicks, J.J.; Andrus, R. Are density reduction treatments effective at managing for resistance or resilience to spruce beetle disturbance in the southern Rocky Mountains. For. Ecol. Manag. 2014, 334, 53–63. [Google Scholar] [CrossRef]
- Windmuller-Campione, A.M.; Long, N.J. If Long-Term Resistance to a Spruce Beetle Epidemic is Futile, Can Silvicultural Treatments Increase Resilience in Spruce-Fir Forests in the Central Rocky Mountains. Forests 2015, 6. [Google Scholar] [CrossRef]
- Frelich, L.E.; Reich, P.B.; Peterson, D.W. Fire in Upper Midwestern Oak Forest Ecosystems: An Oak Forest Restoration and Management Handbook; General Technical Report PNW-GTR-914; United States Department of Agriculture: Portland, OR, USA, 2015.
- Hughes, F.M.R.; Colston, A.; Mountford, J.O. Restoring Riparian Ecosystems: The Challenge of Accommodating Variability and Designing Restoration Trajectories. Ecol. Soc. 2005, 10. [Google Scholar] [CrossRef]
- Long, J.N. The middle and southern Rocky Mountain Region. In Regional Silviculture of the United States; Barrett, J.W., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1994; pp. 335–386. [Google Scholar]
- Roberts, S.D.; Long, J.N. Effects of storage, planting date, and shelter on Engelmann spruce containerized seedlings in the Central Rockies. West. J. Appl. For. 1991, 6, 36–38. [Google Scholar]
- Interagency Lynx Biology Team. Canada lynx conservation assessment and strategy, 3rd ed.; USDA Forest Service, USDI Fish Wildlife Service, USDI Bureau of Land Management, USDI National Park Service, Forest Service Publication: Missoula, MT, USA, 2013.
- Rhoades, C.C.; McCutchan, J.H.; Cooper, L.A.; Clow, D.; Detmer, T.M.; Briggs, J.S.; Stednick, J.D.; Veblen, T.T.; Ertz, R.M.; Likens, G.E.; et al. Biogeochemistry of beetle-killed forests: Explaining a weak nitrate response. Proc. Natl. Acad. Sci. USA 2013, 110, 1756. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Number of publications by year since 1997 for the queries ‘forest AND resistance AND resilience’ and ‘forest AND resistance AND resilience AND regeneration’ using Scholar PlotR (https://csullender.com/scholar/) accessed on 23 February 2018.
Figure 1.
Number of publications by year since 1997 for the queries ‘forest AND resistance AND resilience’ and ‘forest AND resistance AND resilience AND regeneration’ using Scholar PlotR (https://csullender.com/scholar/) accessed on 23 February 2018.

Figure 2.
Pure, mature Engelmann spruce at about 3000 m elevation in the Tushar Mountains, Fishlake National Forest, Central Utah. This stand has no history of management and essentially no resistance nor resilience to spruce beetle attack.
Figure 2.
Pure, mature Engelmann spruce at about 3000 m elevation in the Tushar Mountains, Fishlake National Forest, Central Utah. This stand has no history of management and essentially no resistance nor resilience to spruce beetle attack.

Figure 3.
This lodgepole pine stand on the Uintah National Forest in Utah had a fairly small to average diameter and low relative density. While otherwise resistant to mountain pine beetle attack, it was nevertheless overwhelmed by a landscape-scale outbreak.
Figure 3.
This lodgepole pine stand on the Uintah National Forest in Utah had a fairly small to average diameter and low relative density. While otherwise resistant to mountain pine beetle attack, it was nevertheless overwhelmed by a landscape-scale outbreak.

Figure 4.
Compared to traditional regeneration treatments in lodgepole pine stands, a shelterwood-with-reserves greatly increases structural, age-class, and species diversity. Maintenance of advance reproduction combined with post-treatment regeneration enhances resistance and resilience to mountain pine beetle attacks for this stand on the Daniel Experimental Forest in Northern Utah.
Figure 4.
Compared to traditional regeneration treatments in lodgepole pine stands, a shelterwood-with-reserves greatly increases structural, age-class, and species diversity. Maintenance of advance reproduction combined with post-treatment regeneration enhances resistance and resilience to mountain pine beetle attacks for this stand on the Daniel Experimental Forest in Northern Utah.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
A small subset of the resistance and resilience literature includes multiple definitions of the terms. The theoretical papers serve as an important base of basic understanding but are often too abstract to translate in applied management. There has been an increasing effort to provide frameworks for managers in developing resistance and/or resilience forest ecosystems (see References for full citations).
Table 1.
A small subset of the resistance and resilience literature includes multiple definitions of the terms. The theoretical papers serve as an important base of basic understanding but are often too abstract to translate in applied management. There has been an increasing effort to provide frameworks for managers in developing resistance and/or resilience forest ecosystems (see References for full citations).
| References | Theoretical Definitions of Resistance and/or Resilience: |
| Holling (1973) [8] | Resilience and stability of ecological systems. |
| Holling (1996) [9] | Engineering resilience versus ecological resilience. |
| Peterson et al. (1998) [10] | Ecological resilience, biodiversity, and scale. |
| Gunderson (2000) [11] | Ecological resilience—in theory and application. |
| Carpenter et al. (2001) [12] | From metaphor to measurement: resilience of what to what? |
| Holling & Gunderson (2002) [13] | Resilience and adaptive cycles |
| Cummings et al. (2005) [14] | An exploratory framework for the empirical measurement of resilience. |
| Gallopín (2006) [15] | Linkages between vulnerability, resilience, and adaptive capacity. |
| Benson & Garmestani (2015) [16] | Can we manage for resilience? The integration of resilience thinking into natural resource management in the United States |
| References | Translational Frameworks for Resistance and Resilience Applied Forest Management |
| Millar et al. (2007) [1] | Climate change and forests of the future: managing in the face of uncertainty. |
| Peterson et al. (2011) [17] | Responding to climate change in national forests: a guidebook for developing adaptation options. |
| Churchill et al. (2013) [18] | Restoring forest resilience: from reference spatial patterns to silvicultural prescriptions and monitoring. |
| DeRose and Long (2014) [19] | Resistance and resilience: A conceptual framework for silviculture. |
| Messier et al. (2015) [20] | From management to stewardship: viewing forests as complex adaptive systems in an uncertain world. |
| Nagel et al. (2017) [21] | Adaptive silviculture for climate change: A national experiment in manager-scientist partnerships to apply an adaptation framework. |
Table 2.
Important elements of stand composition and structure associated with susceptibility to spruce beetle attack (after [22]). In general, susceptibility increases with increasing size, density, and percentage of spruce.
Table 2.
Important elements of stand composition and structure associated with susceptibility to spruce beetle attack (after [22]). In general, susceptibility increases with increasing size, density, and percentage of spruce.
| Level of Susceptibility | Physiographic Location/Site Index | Spruce QMD 1 > 25.4 cm dbh | Stand Basal Area (m2∙ha−1) | Proportion of Spruce (%) |
|---|---|---|---|---|
| High | Spruce on well-drained sites in creek bottoms | >40.6 | >34.4 | >65 |
| Medium | Spruce on sites with site index 24.4–36.6 m | 30.5–40.6 | 23.0–34.4 | 50–65 |
| Low | Spruce on sites with site index 12.2–24.4 m | <30.5 | <23.0 | <50 |
1 QMD is quadratic mean diameter. dbh: diameter at breast height.
Table 3.
Important elements of stand composition and structure associated with susceptibility to mountain pine beetle attack in lodgepole pine forests (after [23]). In general, susceptibility increases with increasing size, density, and percentage of lodgepole pine.
Table 3.
Important elements of stand composition and structure associated with susceptibility to mountain pine beetle attack in lodgepole pine forests (after [23]). In general, susceptibility increases with increasing size, density, and percentage of lodgepole pine.
| Level of Susceptibility | Lodgepole Pine QMD 1 > 12.7 cm dbh | Basal Area (BA) of All Species (m2∙ha−1) | Percent of total Basal Area in Lodgepole Pine > 12.7 cm dbh |
|---|---|---|---|
| High | ≥20.3 | 27.6 ≤ BA < 57.4 | ≥50 |
| Moderate | 17.8 ≤ QMD < 20.3 | 18.4 ≤ BA < 27.6 | 25 ≤ BA < 50 |
| Low | <17.8 | <18.4 or ≥57.4 | ≤25 |
1 QMD is quadratic mean diameter.
Table 4.
Comparison of conventional and alternative silvicultural systems for Engelmann spruce and lodgepole pine in the Central Rockies.
Table 4.
Comparison of conventional and alternative silvicultural systems for Engelmann spruce and lodgepole pine in the Central Rockies.
| Engelmann Spruce |
| Conventional: Small group selection system [40] |
| 0.1 ha group size |
| 20–30 year cutting cycle |
| Natural regeneration often combined with enrichment planting |
| <25% of compartment regenerated in a cutting cycle |
| Alternative: Shelterwood-with-reserves [37] |
| Uniform shelterwood with retention of overwood beyond regeneration period |
| Natural regeneration combined with planting |
| 100% of compartment regenerated following initial entry |
| Lodgepole Pine |
| Conventional: Clearcutting system [26,40] |
| 4–15 ha harvest size |
| 80–140 year rotation |
| Mechanical site preparation |
| Natural regeneration |
| Precommercial thinning |
| Alternative: Shelterwood-with-reserves [41] |
| Uniform &/or strip shelterwood with retention of overwood beyond regeneration period |
| 80–140 year rotation |
| No site preparation beyond disturbance during harvesting |
| Retention of advance regeneration, natural regeneration, enrichment planting for tree species diversity |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Long, J.N.; Windmuller-Campione, M.; DeRose, R.J. Building Resistance and Resilience: Regeneration Should Not be Left to Chance. Forests 2018, 9, 270. https://doi.org/10.3390/f9050270
AMA Style
Long JN, Windmuller-Campione M, DeRose RJ. Building Resistance and Resilience: Regeneration Should Not be Left to Chance. Forests. 2018; 9(5):270. https://doi.org/10.3390/f9050270
Chicago/Turabian StyleLong, James N., Marcella Windmuller-Campione, and R. Justin DeRose. 2018. "Building Resistance and Resilience: Regeneration Should Not be Left to Chance" Forests 9, no. 5: 270. https://doi.org/10.3390/f9050270
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.