How Forest Gap and Elevation Shaped Abies faxoniana Rehd. et Wils. Regeneration in a Subalpine Coniferous Forest, Southwestern China

1
State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
2
School of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Forests 2018, 9(5), 271; https://doi.org/10.3390/f9050271
Submission received: 14 March 2018
/
Revised: 8 May 2018
/
Accepted: 14 May 2018
/
Published: 16 May 2018
(This article belongs to the Section Forest Ecology and Management)
Abstract
:Focusing on the underlying ecological mechanisms of dominant species regeneration in forest gaps at a landscape scale can provide detailed understanding for gap-based forest management. The individual effects of forest gaps or elevation on the regeneration of Abies faxoniana Rehd. et Wils. are well known, although elucidating how gap characteristics and elevation concurrently influence regeneration remains an important challenge. In this paper, we present an explorative study using structural equation models (SEMs) to assess the direct and indirect effects of forest gaps and elevation on Abies faxoniana Rehd. et Wils. regeneration. Four of the predicted SEMs showed the following results: (1) Temperature, photosynthetic photon flux density (PPFD), soil total carbon, gap openness, shrub layer cover, herb layer cover, and moss layer thickness in forest gaps were associated with Abies faxoniana regeneration along an elevation gradient in subalpine coniferous forest. (2) Elevation had a generally negative and indirect effect on Abies faxoniana regeneration. Forest gaps positively affected regeneration when compared with non-gap plots and gap size was positively related to small tree regeneration density and the ratio of height to diameter at breast height (HD ratio) of the tallest Abies faxoniana small trees but was negatively related to Abies faxoniana sapling regeneration density. (3) In forest gaps, the Abies faxoniana sapling density and HD ratio of the tallest Abies faxoniana small trees were mainly indirectly influenced by elevation, and Abies faxoniana small tree regeneration density was directly associated with the dominance of the sapling regeneration density. In summary, Abies faxoniana regeneration was negatively and largely affected by elevation (total effect), although forest gaps enhanced Abies faxoniana regeneration by multiple pathways (direct and indirect effects).
1. Introduction
Forest gaps create important spaces for forest regeneration by modifying microclimate and resource availability [1,2]. In many forests, these gaps not only play the role of resource “hot spots” during the forest growth cycle but also help maintain the late succession of forests [3]. In recent decades, to meet sustainable forest ecosystem functions (e.g., diversity and resilience) and silvan economic benefits (e.g., timber production) [4,5], forest ecologists and managers have increasingly emphasized new silvicultural concepts that imitate natural disturbances for forest management [6,7]. Gap-based silviculture is an intuitive and effective approach for promoting plant regeneration in the majority of mature forests and has been extensively documented in proposals for forest management [8,9]. In this way, forest gaps are thought to play an important role in the natural regeneration and sustainable development of forest ecosystems.
Many studies have focused on gap characteristics and their relationships to regeneration [3,10]. As an important factor of gap characteristics that determines species composition, understanding the effect of gap size on natural regeneration has attracted the attention of many researchers [11,12,13]. In general, gap size is considered an important and direct influence on resource sequestration, gap microclimate and vegetation growth in a forest gap [14,15,16,17]. Additionally, as an important geographical factor, elevational gradients have predictable changes in numerous environmental factors (e.g., temperature and solar radiation) altitudinally on a single mountain, but also have remarkable influence on plant regeneration along with these environmental drives [14]. However, despite numerous studies conducted to describe the relationship between gap characteristics and regeneration in various forest ecosystems [18,19], most of the studies on forest gaps have either focused on gaps created at the same altitude or ignored the altitude factor [20]. Although gaps formed at high elevations are more difficult to measure, they are also important for the regeneration and release of plants [21]. In particular, forest management has a real and urgent need for studying the influence of elevation on regeneration because many gap phase forests in temperate forests throughout the world occur in mountainous regions [14,20].
Our study area provided a unique opportunity to study forest gaps and their effects on natural regeneration over a typical range of elevations and gap size classes because of the terrain conditions [22]. In the subalpine region of southwestern China, widespread coniferous forests have provided an important economic resource (i.e., ecological tourism) and have acted as important ecological refuges along the upper reaches of the Min River [23]. Simultaneously, coniferous forests have also experienced different degrees of disturbance from natural events [22,23]. In this respect, research on forest gaps could be regarded as necessary for the sustainable development of forest ecosystems in our study area.
The study of the dominant species regeneration across different life stages from seedlings to small tree within and among gaps is important for determining the influence of gaps on long-term regeneration succession [24,25]. Abies faxoniana Rehd. et Wils. is one of the dominant tree species observed along the elevation gradient (2800–3800 masl) of subalpine mature coniferous forests in southwestern China. Regeneration is restricted by many factors (e.g., temperature and soil) in forests [22,23]. Therefore, Abies faxoniana saplings and small trees are typically rare in mature subalpine coniferous forests [22,25], and comprehensive research is needed to understand in detail the mechanisms of affecting their natural regeneration. Several studies have shown the effects of gaps on regeneration during different life stages [2,26,27]. However, few reports have focused on the subject of subalpine coniferous forest gaps along an elevation gradient and considered the distinction in resource demand (e.g., light) on a species’ architectural development [27].
Moreover, although many studies have evaluated the effects of elevation or forest gaps on plant regeneration [16,23,28], the methodology is still a considerable limitation for exploring the internal mechanisms in current forest gap studies. For example, the influence of abiotic factors on regeneration is simultaneously induced by elevation and forest gaps. Separating the effects of elevation on regeneration from the effects of forest gaps is difficult [29]. Multiple regression analyses do not allow for the investigation of the direct and indirect effects of elevation, forest gaps, biotic factors and abiotic factors on regeneration; however, a structural equation model (SEM), which is also known as a path analysis, alleviates this problem. SEM is also an extension of the general linear model (GLM), which enables evaluating a network of dependence relationships through the analysis of covariance [29,30]. The general methodology of SEM enables the testing of a hypothesis that infers causation with observational data [31]. Direct pathways represent the path model consistent with the data while indirect pathways between variables are those involving intermediary variables [30,31]. More introductory discussion and practical issues of SEM were described by Kubota et al. [29], Gough et al. [31] and Grace et al. [30]. In conclusion, the use of SEM enables us to determine how Abies faxoniana regeneration is affected by forest gaps and elevation in a subalpine coniferous forest. Therefore, the main purpose of using SEM is to elucidate the complicated ecological processes of forest gaps and elevation effects on Abies faxoniana regeneration. Accordingly, we tested the effects of gap/non-gap plots, gap size and elevation on Abies faxoniana regeneration in a subalpine coniferous forest in southwestern China. Our hypothesis was that the variables of habitat characteristics and elevation cause microenvironment heterogeneity and thus affect Abies faxoniana regeneration at a landscape scale [3,15,28,29,32]. In short, this paper proposes a simple and reliable quantitative method for assessing Abies faxoniana regeneration based on 34 plots distributed along an elevation gradient in a subalpine coniferous forest under a “direct effects and indirect effects” analytical framework.
Specifically, we aimed to answer the following questions: (1) Which endogenous variables are affected by habitat characteristics and/or elevation in our study? (2) How do habitat characteristic and elevation relate to biotic factors (e.g., ground shrub cover, herb layer cover, and moss layer), abiotic factors (excluding exogenous variables (forest gap and elevation), e.g., mean annual temperature, photosynthetic photon flux density (PPFD), and soil properties), and Abies faxoniana regeneration? (3) What is the key factor (largest total effect on Abies faxoniana regeneration in SEM) during the succession regeneration of Abies faxoniana in forest gap along the elevation in our study? Based on our results, several implications related to forest management and providing the best and sustainable development conditions for the regeneration of Abies faxoniana are also discussed.
2. Materials and Methods
2.1. Study Site
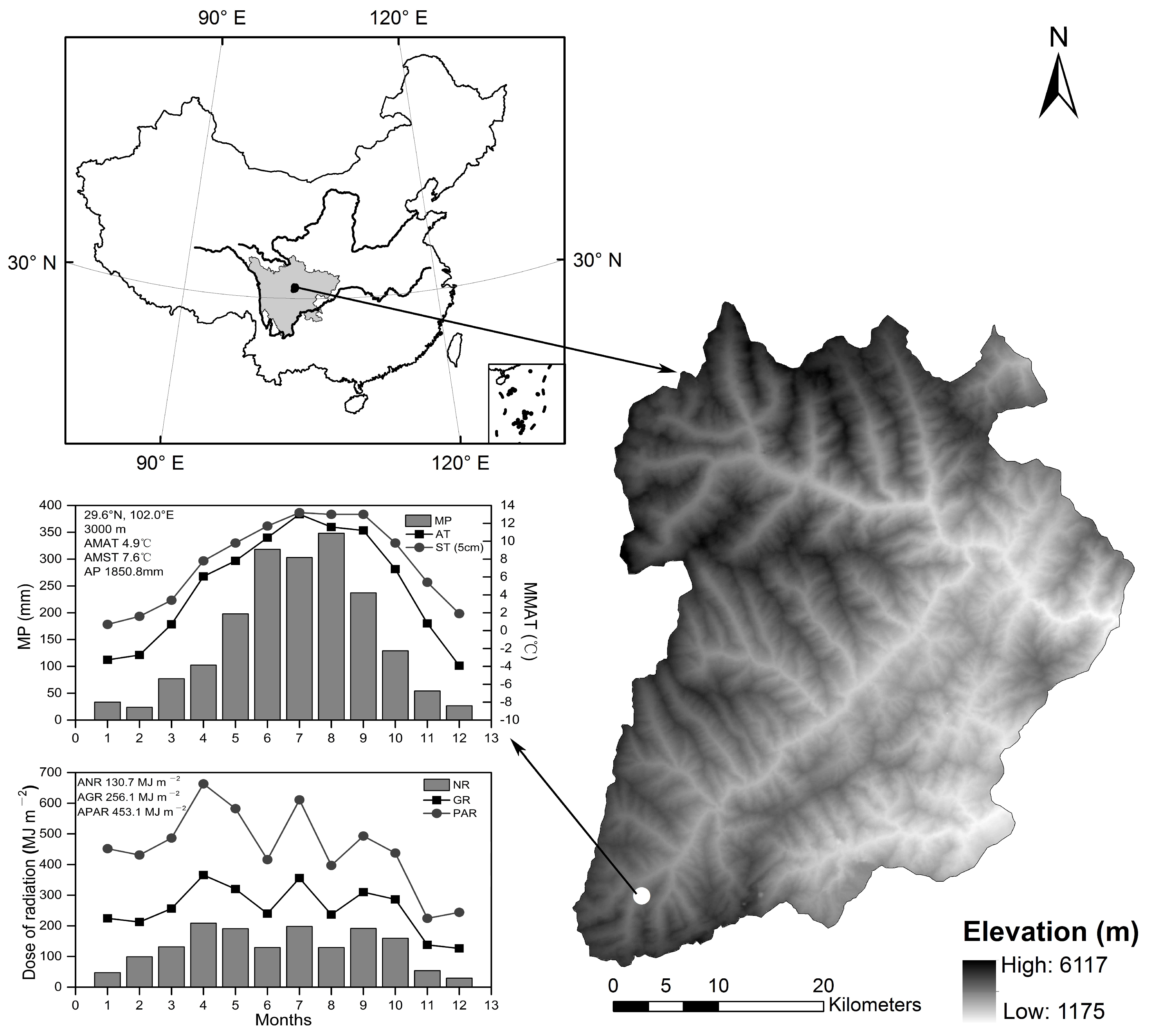
The study was conducted within the Wolong National Nature Reserve in a subalpine mature coniferous forest in southwestern China (Figure 1). Forest gaps were selected in a 4.8 ha study area that ranged from 30°49′ N to 30°50′ N in latitude and from 102°56′ E to 102°57′ E in longitude and had an elevation span of 3000–3700 m. Alpine valleys are the main geomorphic feature, and the climate is subtropical inland and mountainous with a mean annual temperature of 8.4 °C and a mean annual rainfall of 861.8 mm. The parent rock in brown coniferous forest soil is weathered killas and has a pH value that ranges from 3.9 to 5.0. The study area is in an old-growth forest (i.e., a history of natural disturbance, old-growth structures and little or no anthropogenic influence). In this old-growth forest, self-thinning or other small-scale natural disturbances (e.g., wind and snowfall) are common causes of gap formation [22]. Abies faxoniana is the dominant species along the elevation (3000–3700 masl) in this mature subalpine coniferous forest in southwestern China. At middle and low altitudes, dominant companion tree species are Betula utilis D. Don and Acer maximowiczii Pax. With a decline in the elevation gradient, shrubs are mainly Rhododendron faberisp. Prattii (Franch.) D. F. Cha mb. (1978), Rhododendron phaeochrysum Balf f. et W. W. Smith var. agglutinatum, and Fargesia nitida (Mitford ex Stapf) Keng f. ex T. P. Yi (GRIN). The herb layer is rich in species and composed of species that include Circaea alpina L., Parasenecio deltophyllus (Maxim.) Y. L. Chen, and Oxalis acetosella L.
2.2. Experimental Design
We investigated two main habitat characteristics in our study: gap heterogeneity (forest gaps or non-gap plots, also called control check plots (CK plots)); and gap size (small gaps, medium gaps and large gaps). Gap characteristics include gap area, gap age, and gap openness. All sampling was conducted during August and September of 2014 and 2015. Through a typical sampling investigation, we studied 34 plots (26 gaps (Nlarge = 9; Nmedium = 9; Nsmall = 8) and 8 plots in the understory of a closed forest as CK) in total, which were distributed along an elevation gradient (3000–3700 masl). At each 100 m elevation interval, we selected at least 3 plots (small gaps, medium gaps and large gaps, respectively) randomly, which were established in the study area. Furthermore, at each 100 m elevation interval, we also set a CK plot (10 m × 10 m) at least 10 m away from gaps in the forest. All gaps were defined as canopy openings with areas exceeding 4 m2 that had a gap maker tree [33], and where small regeneration trees in the gaps had not yet reached the base of the canopy layer at gap borders [34]. The longest axis and the perpendicular axis of each gap were measured for calculating the area [2]. The species, quantity, basal diameter, and formation mode were recorded for the gap maker trees. The estimates were also used to determine each gap’s probable age with the highest level of gap maker decomposition [3]. The characteristics of the gap border trees were also recorded with the same method as the gap makers for each gap. The regeneration plants, which were shorter than 2/3 of the forest canopy layer, were investigated in the gaps and CK plots through a census. We recorded plant parameters including the species, quantity, height, diameter at breast height (DBH), and age. Other gap fillers in the shrub layer and herb layer were designated by their species, quantity, height, and cover degree. The shrub cover was estimated in a quadrat (5 m × 5 m), while the herb cover and the moss layer thickness were estimated in three randomly selected subplots (1 m × 1 m) in each plot (gaps and CK plots) [22].
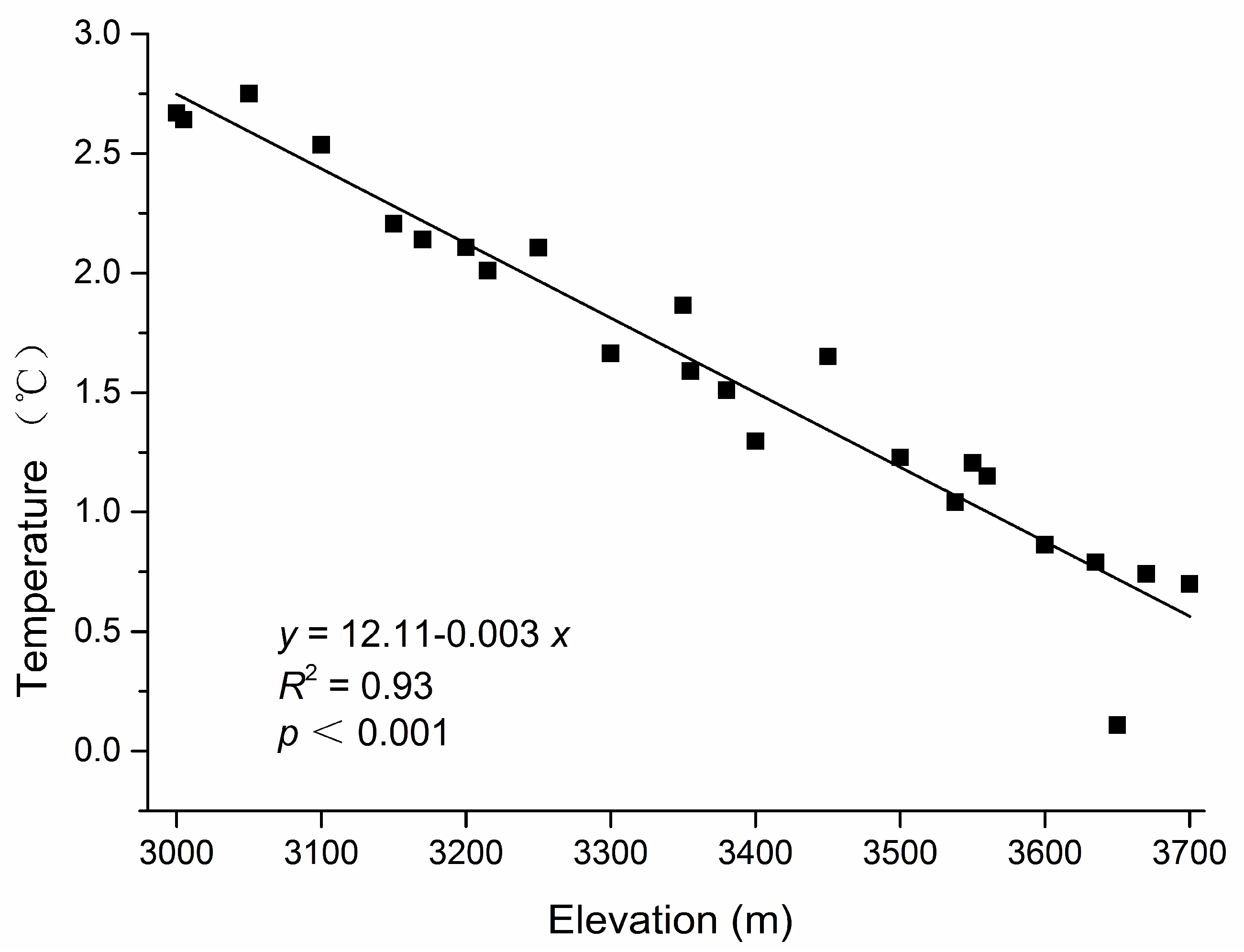
The air temperature was measured every 2 h in the center of each plot using an iButton recorder (iButton DS1923-F5, Maxim/Dallas Semiconductor, Sunnyvale, CA, USA) from 20 August 2014 to 2 September 2015. Mean annual temperature data were based on a combination of field monitoring data and estimations from the regression equation of elevation–temperature plots (slope = −0.003, R2 = 0.93) (Appendix A, Figure A1) [35]. The data for the instantaneous above-canopy photosynthetic photon flux density (PPFD) (LI-190SB, LI-COR, Inc., Lincoln, NE, USA), which was measured under completely overcast sky conditions, were stable and exhibited a good correlation with the growing season PPFD [36,37]. Next, the PPFD was measured at 1 m above the forest floor under completely overcast sky conditions in the middle of the samples sites in our study, and any vegetation shading of the sensor was avoided before the reading was recorded. We collected three random soil samples beneath the litter layer at a depth of 0–10 cm in each plot, and the samples were stored in clean airtight Ziploc bags. In total, 103 soil samples were collected, and at least 250 g was obtained at each sampling point. Soil samples were air-dried for three weeks at room temperature. The soil properties (i.e., soil water content, pH, total organic carbon (C), and total nitrogen (N)) were measured in each sampling quadrat. Plant debris, roots and stones were removed from soil, and the water content was determined with the oven drying method. The soil pH was measured in a 1:5 mixture of soil to deionized water. The analyses of soil total C and total N were measured using a Vario EL III analyzer (Elementar Analysensysteme GmbH, Hanau, Germany).
2.3. Data Processing and Analysis
We divided the elevation range from 3000 masl to 3700 masl into 7 elevation intervals (100 m per elevation gradient). Additionally, we divided the four habitats into small gap (4–100 m2), medium gap (101–200 m2), large gap (201–410 m2) and CK plots (100 m2). Following Runkle’s [38] proposal to calculate gap area when the gap shape was an ellipse, we used the formula [38]:
where A represents the area of an ellipse, L represents the average values of the long axes, and W represents the average values of the short axes.
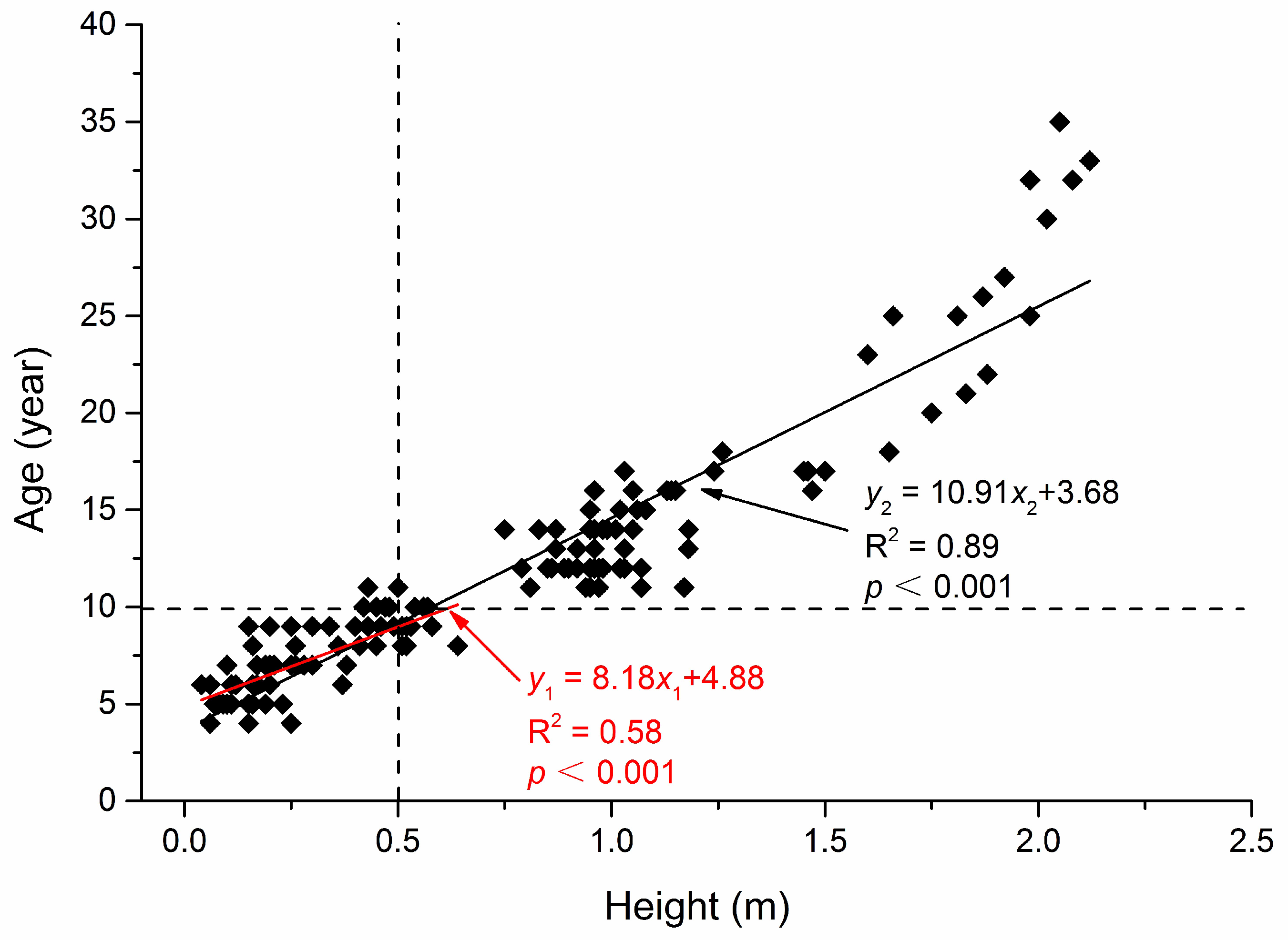
The formula for the area of a circle was used when the difference between the length of longest axis and the length of the perpendicular axes in a gap was less than 1 m, and the gap area was calculated with the mean value of the axes as the radius. For the gap openness, a measurement was performed using the ratio between the average gap diameter and the height of the tallest gap border tree [15]. To assess the recruitment density based on the competition for light capture in different age groups [27], the regeneration height was classified into two groups—saplings (height: 0.05 m < h ≤ 0.5 m), and small trees (height: 0.5 m < h ≤ 5.2 m)—based on linear regression analysis (Appendix A, Figure A2).
The following parameters were assigned as exogenous variables: habitat characteristics and elevation gradient. The endogenous variables were biotic factors (e.g., cover of ground shrub, herb layer, and moss layer), abiotic factors (excluding the exogenous variables, e.g., mean annual temperature, PPFD, and soil properties), and the regeneration of Abies faxoniana (e.g., HD ratio of the tallest small trees, and density of saplings and small trees). To calculate each endogenous variable, we used descriptive statistics (Table 1). A multivariate analysis of variance (MANOVA) analysis was used to test the effects of endogenous variables on exogenous variables. A Duncan test was used to identify the multiple comparisons (after testing for a normal distribution and homogeneity of variance). Pearson correlations were used to test the relationship between biotic/abiotic factors and Abies faxoniana regeneration. Significance level was set at p < 0.05. Bonferroni-adjusted was applied to compensate for multiple testing and the descriptive statistics in this article are described using the mean ± SE (standard error).
2.4. Structural Equation Modeling
Structural equation modelling (SEM) is an extension of the general linear model (GLM) and ideal for studying the correlation and causation among variables of complex system [29]. We used SEM to analyze the direct and indirect predictors of Abies faxoniana regeneration [39]. Initially, we developed an a priori hypothetical model based on theoretical and empirical evidence. In the model (Figure 2), we hypothesized that the variations in Abies faxoniana regeneration could be explained by the direct and indirect effects of exogenous variables (habitat characteristics and elevation), and biotic and abiotic conditions, such as the ground herbs cover (%) and mean annual temperature (°C) [15,28]. Second, all conditional independence claims present between variables (gap characteristics, elevation, environmental factors, and Abies faxoniana regeneration) were identified and tested to determine whether the hypothesized pathways could reflect variation present in the data by using directional separation tests. Subsequently, the hypothesized candidate models with significance pathways were compared [29]. Based on the analysis, we identified the fit of working models by: (1) a non-significant χ2 test (p > 0.05); (2) GFI (Goodness Fit Index) and CFI (Comparative Fit Index) > 0.9; and (3) a lower 90% confidence intervals of root mean square error of approximation (RMSEA) < 0.05 [39,40]. Next, we selected a more suitable model from working models using the lowest Akaike’s information criterion (AIC) value. A smaller AIC value indicate that the number of parameters is more parsimonious in SEMs, and it is defined as follows:
where q represents the number of parameters in the fitted model [29]. Lastly, the standard pathway coefficient (β) and p value were used to assess the significance of each independent variables in the final model. Modification indices were used to improve the adequacy of model. The R2 values show the variance of each dependent variable due to the effect of the other variables [29]. The total effects, direct effects and indirect effects of gap characteristics, elevation, biotic factors and abiotic factors on Abies faxoniana regeneration were evaluated. All SEM analyses were performed in Amos 22.0 (SPSS Inc., Chicago, IL, USA).
In our best models, the dependent variables were the regeneration sapling and small tree densities (N m−2) of Abies faxoniana and the ratio of height to DBH of the tallest small trees (HD ratio). The independent variable of gap heterogeneity was coded as gap = 1 and CK = 0, and the other independent variables were gap area (m2) and elevation (m). The biotic and abiotic factors acted as both dependent variables and independent variables, for instance, the PPFD (dependent variable) varied with the elevation (independent variable), and PPFD (independent variable) also influenced Abies faxoniana regeneration (dependent variable).
3. Results
3.1. Gap Heterogeneity, Elevation Gradient and Independent Variables
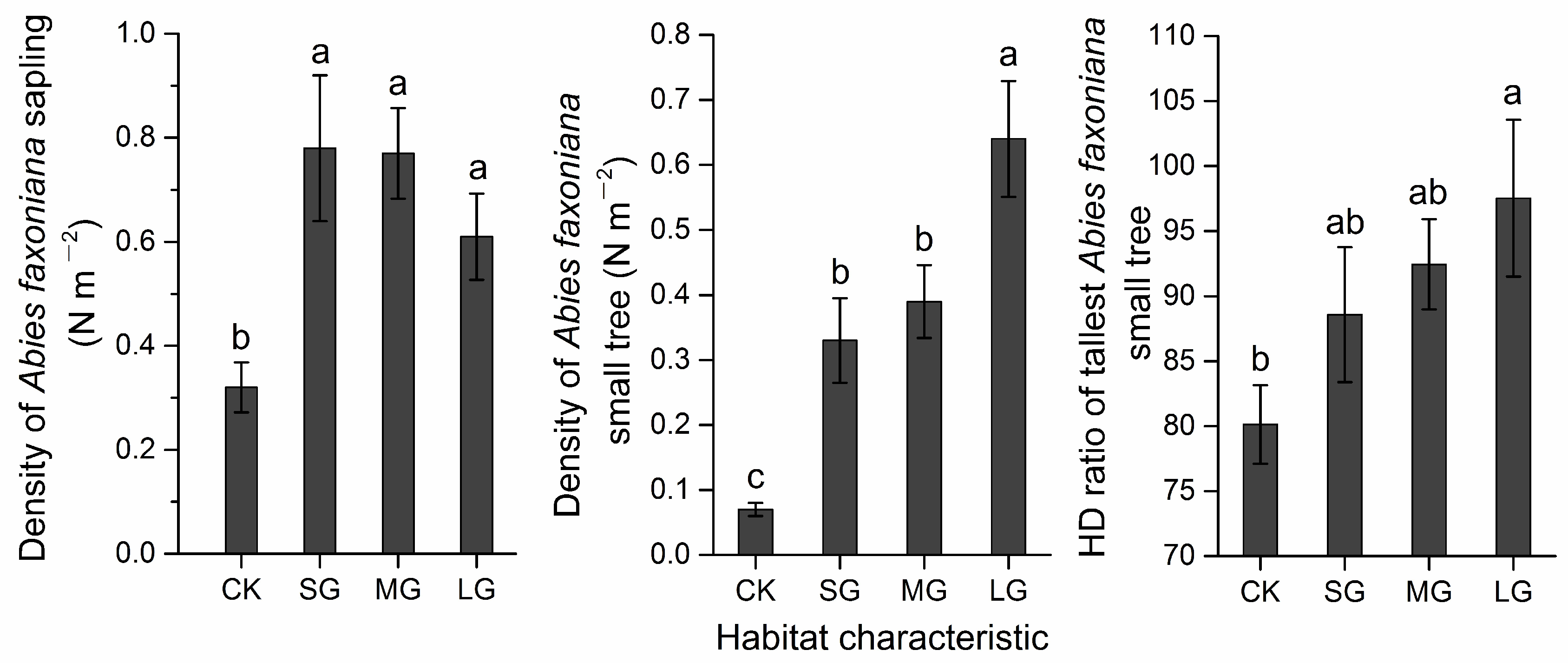
The results for the MANOVA (Table 2) showed that the abiotic variable of PPFD in the gaps was significantly higher than that in the plots from the forest understory (F = 406.434, p < 0.001). Although an increasing trend was observed from the forest understory (1.50 ± 0.23 °C) to the gaps (1.62 ± 0.13 °C), the mean annual temperature was not significantly different between forest gaps and non-gap plots (F = 0.754, p = 0.570). The soil properties, moss thickness, shrub cover and herb cover in the forest understory were similar to those in the gaps (Table 2). The variables for the regeneration density of Abies faxoniana saplings and regeneration density of Abies faxoniana small trees in gaps were significantly higher than those in the forest understory (Table 2 and Figure 3 and Figure 4A). Moreover, significant correlations were observed between the regeneration densities of Abies faxoniana small trees and saplings (R2 = 0.719, p < 0.001), among the HD ratios of the tallest Abies faxoniana small trees (R2 = 0.735, p < 0.001), and between the HD ratio of the tallest Abies faxoniana small trees and shrub coverage (R2 = 0.655, p < 0.001).
The soil properties, such as the soil total N (F = 0.273, p = 0.927) and soil total C (F = 1.185, p = 0.436), were also similar in the plots along the altitude. Compared with the mean annual temperature (F = 140.654, p < 0.001), although a clear gradient was observed along the elevation gradient, the PPFD (F = 2.43, p = 0.174) was not significantly different among the elevation gradients. All studied biotic variables presented significant differences with elevation in our study (Figure 4B).
3.2. Gap Size, Elevation Gradient and Independent Variables
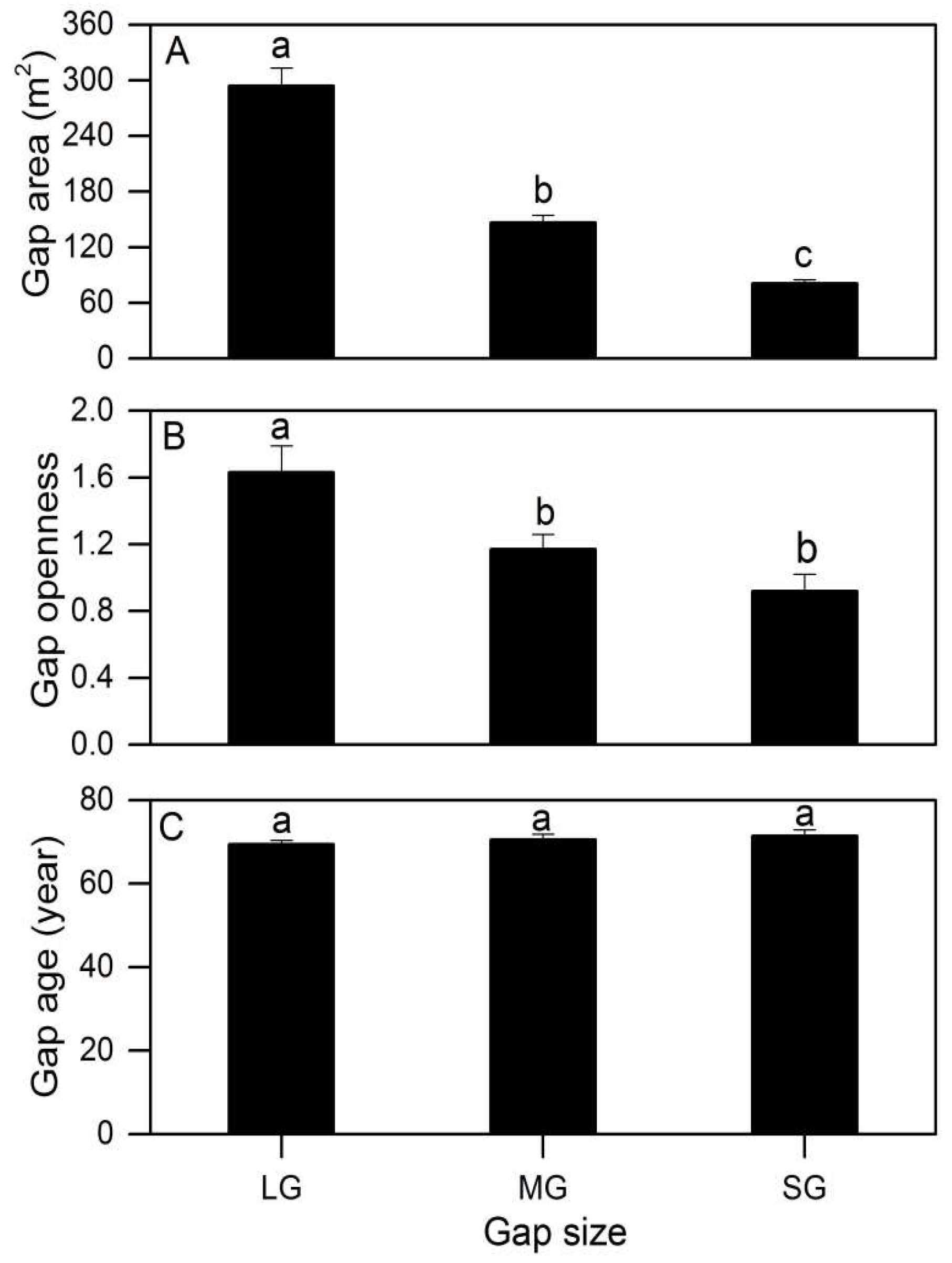
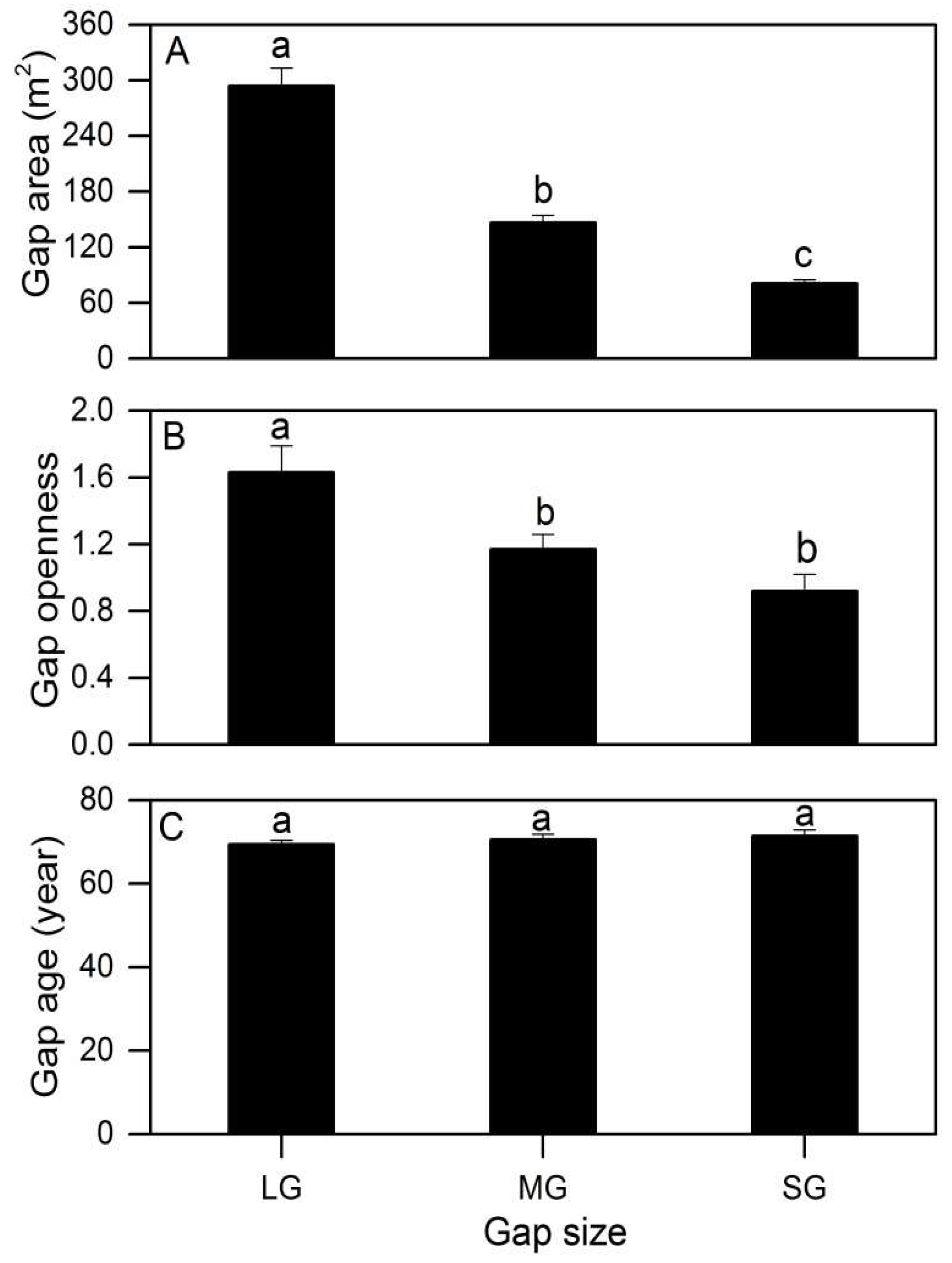
Gaps presented significantly higher openness increments (F = 5.985, p = 0.044), and varied from 0.97 ± 0.10 (small gap) to 1.63 ± 0.16 (large gap) (Appendix A, Figure A3B). The gap size affected the regeneration density of Abies faxoniana small trees in the gaps (F = 38.494, p = 0.001), whereas the gap age, Abies faxoniana sapling density, HD ratio of the tallest Abies faxoniana small trees, PPFD, mean annual temperature, soil total C, soil total N, shrub cover, herb cover and moss thickness did not present statistically significant differences in different size gaps (Table 2 and Figure 4C and Figure A3C). In forest gaps, a significant correlation was observed between the Abies faxoniana sapling density and herb coverage (R2 = 0.785, p < 0.001).
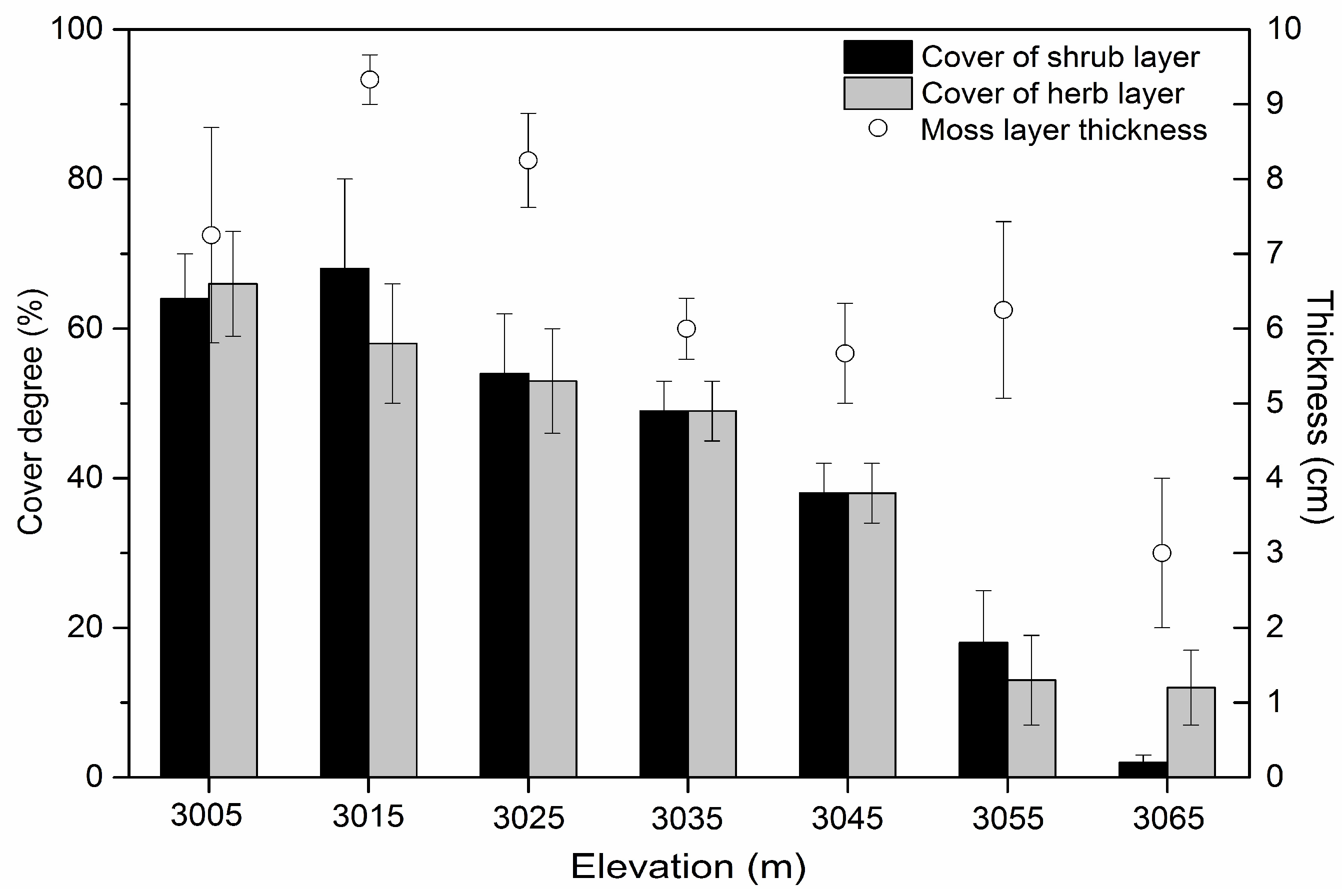
For gaps, the results of the MANOVA (Table 2) showed that the mean annual temperature (F = 157.861, p < 0.001), shrub cover (F = 20.784, p = 0.002), herb cover (F = 21.057, p = 0.002) and moss thickness (F = 5.536, p = 0.028) significantly decreased along the elevation gradient (Figure 4D). High elevation significantly decreased the regeneration density of saplings (F = 25.887, p = 0.001), small trees (F = 27.657, p = 0.001) and the HD ratio of the tallest Abies faxoniana small trees (F = 10.443, p = 0.010). Neither the gap characteristics nor the soil properties of the gaps were significantly affected by elevation in our study (Figure 4D).
3.3. Relationship between Indicators and Abies faxoniana Regeneration in SEM
The reasonably good fit of many models indicated that our conceptual model adequately describes the complicated relationships among the variables. The optimal mode was composed of gap heterogeneity, plot area (including gap area and the plots in the forest understory), elevation, PPFD, mean annual temperature, soil total C, herb cover, shrub cover, moss thickness, sapling and small tree density, and the HD ratio of the tallest Abies faxoniana small trees (Table 3). In the study, many variables, such as the regeneration density of Abies faxoniana saplings (0.72 ± 0.06 N m−2), density of Abies faxoniana small trees (0.46 ± 0.05 N m−2), PPFD (266.19 ± 3.65 μmol m−2 s−1), and shrub cover (0.40 ± 0.05%) in forest gaps, were elevated relative to the CK plots. Gap openness and the regeneration density of small trees increased with the increase in gap size (Table 1 and Figure 5 and Figure A3B). The mean annual temperatures, shrub cover, herb cover, and moss thickness decreased along the elevation gradient (Figure 5).
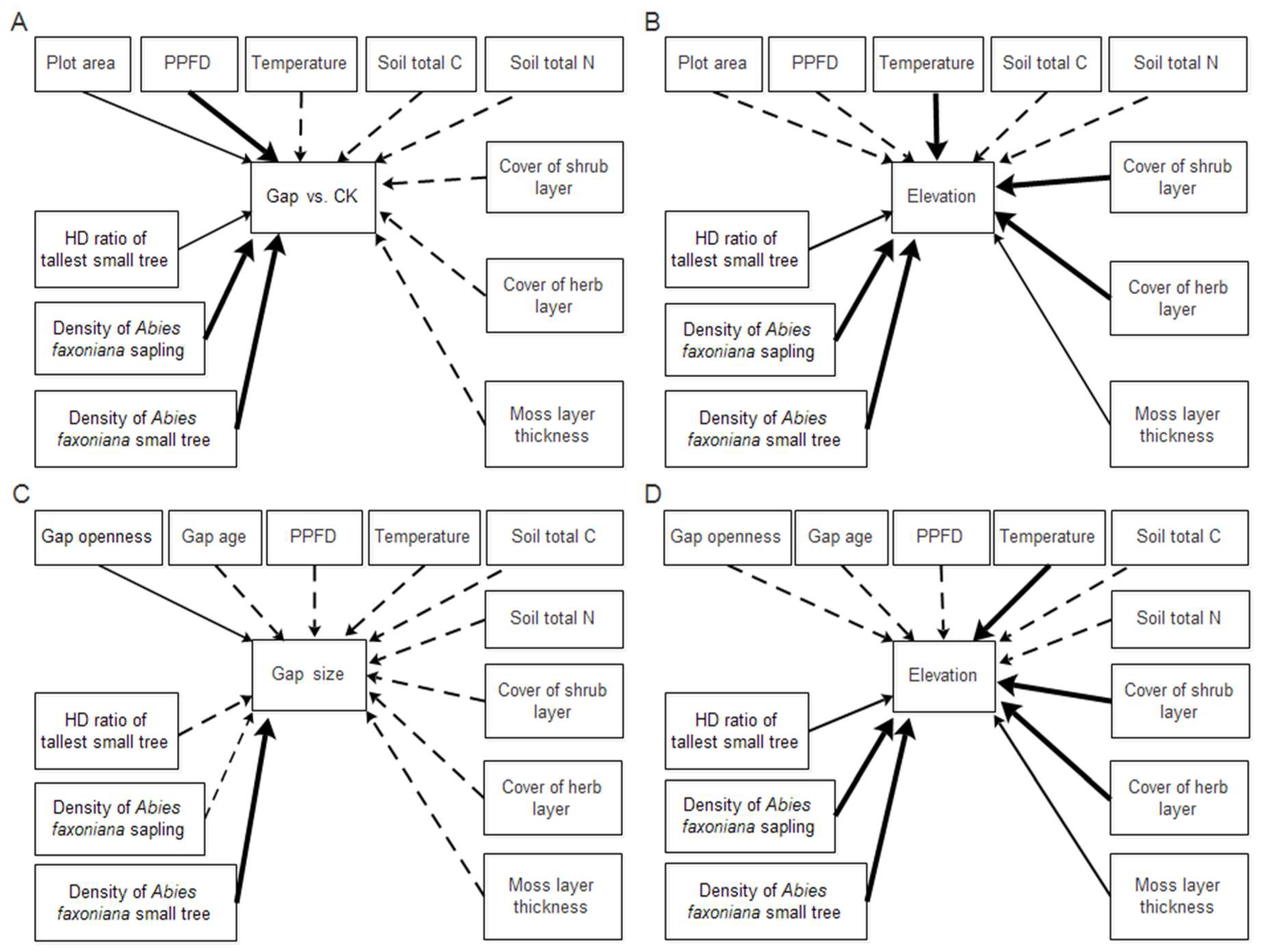
Of the pathways present in the best models (Figure 6), all were consistent with our conceptual model. Overall, the explanation degree of the HD ratio of the tallest Abies faxoniana small trees and the regeneration densities of saplings and small trees were no less than 40%, which was explained by other variables in the best models. The strong correlation between variables indicates that forest gaps and elevation had an influence on the abiotic factors, biotic factors, and regeneration.
The total impact of the direct and indirect effects of HD ratio attributes of the tallest small trees and the regeneration densities of saplings and small trees was used to compare the relative importance of causal factors (Table 4). For example, the total effect of gap size on the density of small trees can be derived from paths leading directly from the gap area to the density of small tree, as well as from the gap openness or the density of saplings (indirect effects). The regeneration density of saplings amounted to the largest total effect (0.715) on the regeneration density of small trees, although the gap area (0.653) was also significant. The regeneration density of small trees among gaps was primarily influenced by the density of saplings (total effect = 0.715) followed by the gap area (0.653), elevation (−0.585), mean annual temperature (0.44), and shrub cover (0.194).
4. Discussion
Our findings revealed the relationships among habitat characteristics, biotic factors, abiotic factors, and Abies faxoniana regeneration in forest gaps along the elevation. First, according to the initial results almost all of the biotic factors exerted a direct and positive effect on Abies faxoniana regeneration in our study. Second, compared with the biotic factors, the effect of abiotic factors on Abies faxoniana regeneration showed a complex ecological process that involved more additional factors and indirect effects. Third, the SEM explicitly predicted attributes by providing circumstantial direct/indirect effects analysis.
4.1. Effect of Biotic Factors on Abies faxoniana Regeneration
Many studies have indicated a negative relationship between sapling regeneration density and vegetation cover because of the existing competition [22,41,42]. However, Tirado and Pugnaire [43] indicated that positive plant–plant interactions (facilitation) occurred in constraining environments, and the impacts of facilitation are receiving increasing attention in community ecological research [44,45]. Our findings concurred that the regeneration density of saplings is directly and positively affected by the herb layer cover (Figure 6C). This finding may be closely related to the plant functional type. Zhu et al. [28] stressed that the high regeneration density of shade tolerant species might create a trade-off among different factors in forest gaps that had higher light intensity environments than those in an understory forest. Our results partially support this finding because the sapling of Abies faxoniana, a common shade tolerant species, might benefit from a low light environment with sufficiently high herb and shrub layer coverage for survival. Concurrently, the herb layer and shrub layer could also protect the saplings from harsh weather conditions in gaps [46]. However, the competition among plants cannot be ignored if a niche overlap of plant communities or resources limited within late-successional gaps, which slowly revert to those of closed forest, are present [3].
The classification criteria for different life stages of plants were not consistent [27,47], and the late regeneration of plants is also one of important focuses of gap regeneration research [2,19]. Numerous studies have considered environmental variables as important drivers of plant survival and growth because of the strong correlations between these drivers in forest gap research [48,49]. However, vegetation interactions are also an important factor in regeneration dynamics [50]. In fact, the early regeneration plant densities had a direct effect on the late regeneration plant density. Our results support that the Abies faxoniana small tree density is positively and moderately explained by the sapling density (Figure 6D). Moreover, our SEM also indicated the direct and/or indirect effects on the small tree density through other variables. Thus, the influence of other factors cannot be denied, and the mechanisms influencing gap tree regeneration were not single ecological processes.
Aiba and Kohyama [51] stressed that the maximum tree height could reflect the competitive capacity of light capture along a vertical gradient. Furthermore, the HD ratio of the tallest Abies faxoniana small trees was also applied to indicate plant competition for resources in forest gaps such as light, nutrients and water [10]. Our results showed that the HD ratio was directly affected by the shrub cover and small tree density. The appropriate shrub coverage is conducive to Abies faxoniana regeneration in forest gaps and has been discussed in our study. However, a favorable micro-environment is not only beneficial to an increased regeneration density but also promotes plant competitiveness. Our results concurred that the HD ratio of the tallest Abies faxoniana small trees in forest gaps was positively associated with Abies faxoniana regeneration density. This finding indicated that features of plant regeneration might respond differently to influence factors but might also have internal relations. Overall, in addition to the competition, intraspecific mutualism and interspecific mutualism are important relationships in plants. Accordingly, further studies are needed to better understand the relationships between biotic factors and Abies faxoniana regeneration in forest gaps.
4.2. Effect of Abiotic Factors on Abies faxoniana Regeneration
Excluding gap size because of its classification as an exogenous variable in our study, studies have reported strong correlations between gap characteristics and regeneration in various forests [15,28,52,53]. In Cameroonian forests, Bongjoh and Mama [54] determined that seedling density was highest in the youngest gaps. However, we did not observe a significant difference in gap age among the gaps in our study. As an important gap characteristic that influences endogenous factors in Abies faxoniana regeneration, gap openness showed an indirect and positive effect on Abies faxoniana regeneration density via herb layer coverage. Specifically, an appropriately high herb layer coverage resulted from larger gap openness and could promote Abies faxoniana regeneration density. According to Vajari et al. [7], a gap openness >1.0 is the minimum size for Pinus thunbergii regeneration; however, a low level of Abies faxoniana regeneration was observed in the low openness gaps (gap openness > 1.0) and understory. Considering the difference in effect mechanisms among different forests, we concluded that gap openness should be further studied in future gap studies. Furthermore, enlarging the gap openness might be considered by forest management because it could improve the herb layer cover and then indirectly promote the regeneration density of Abies faxoniana small trees.
Gaps play an important role in maintaining the heterogeneity of micro-environments (e.g., temperature, PPFD, and soil) in a forest. Simultaneously, these variables also influence gap floristic establishment, development, and gap composition. For gap regeneration, many studies have focused on the relationships among gap characteristics, regeneration success and the micro-environment above the ground (e.g., light level, air temperature and precipitation) [28,42,55,56]. Poulson and Platt [57] reported that the amount of light in gaps was related to their topographic position and sky condition. Additionally, direct radiation can also increase the air temperature and precipitation [11]. Our study revealed a higher PPFD in forest gaps than that in the understory under completely overcast sky conditions, and showed a positive influence of PPFD on the HD ratio of the tallest small trees despite the negative relationship between PPFD and Abies faxoniana regeneration density. The air temperature was mainly influenced by elevation and had a direct effect on Abies faxoniana regeneration density. Numerous studies have shown that higher light values were detected in larger gaps [15,26]. However, if we wished to consider the accumulative total PPFD or temperature in all forest gap areas among different sizes of gaps long-term observations would be required. Combined with the results previous studies, small gaps could be applied in the early period of forest management because these conditions would be more suitable for Abies faxoniana sapling regeneration; moreover, gaps might better meet the requirements of Abies faxoniana small trees in subalpine coniferous forests in southwestern China.
Soil properties reflected several different environmental features of belowground environments and influenced plant regeneration [15,57,58]. However, studies on the effects of soil conditions on gap regeneration have not provided clear results. Studies have shown that small gaps had higher C/N ratios (an index for monitoring litter decomposition) than larger gaps [8,59]. According to Denslow et al. [60], litter decomposition was not significantly related to gap size. Our results did not detect a significant relationship among gap size, elevation and soil properties, although soil total C, as an indirect but not a major factor, affected the Abies faxoniana regeneration density in the SEM. In this subalpine forest, variations in habitat characteristics and elevation lead to regeneration heterogeneity, a mutual consequence of multiple factors, although additional evidence supporting the edaphic requirements of Abies faxoniana regeneration in gaps was not obtained. To explore the relationship among Abies faxoniana regeneration density, the HD ratio of the tallest Abies faxoniana small trees and soil conditions, we may need to consider additional indices of soil properties (, , etc.).
4.3. Direct and Indirect Effects of Forest Gap and Elevation on Abies faxoniana Regeneration in the SEM
The importance of the SEM is that it can be used to detect the influence of forest gaps and elevation on Abies faxoniana regeneration. The variance in plant regeneration in forest gaps was explained by different categorical variables effects as documented by Zhu et al. [28], and our SEM further indicated both the direct forest gap and elevation effects on Abies faxoniana regeneration and their indirect effects through impacts on the biotic and abiotic factors in gaps.
In our study, gap heterogeneity directly and positively influenced Abies faxoniana sapling regeneration density. Our findings also support the results indicating that gap size had a dissimilar effect on the Abies faxoniana sapling and small tree regeneration densities [15,61]. More importantly, further studies showed that regeneration is explained adequately by direct effects, but not entirely. An opposite indirect effect could be caused by the same factor to a dependent variable. For example, in our study region, gap size directly and negatively influenced Abies faxoniana sapling regeneration density, and although the total effect was still positive, the indirect effect was also positive. Compared with the direct effect, the indirect effect is more complex. Our results showed that the forest gap exerted multiple indirect effects on the HD ratio of the tallest small trees. For example, gap heterogeneity showed an indirect effect on the HD ratio of the tallest Abies faxoniana small trees via the strengths of the PPFD and shrub layer coverage in forest gaps and was associated with an increase in the HD ratio of the tallest Abies faxoniana small trees. In our study region, the indirect effect of gap size on the HD ratio of the tallest Abies faxoniana small trees was also observed through the Abies faxoniana regeneration density of small trees. Surprisingly, the intermediate factor of an indirect gap size positively affecting the HD ratio of the tallest Abies faxoniana small trees was the active relationship with Abies faxoniana small trees regeneration density in our study; the result may also further confirm that large gap benefited the late regeneration of Abies faxoniana in our study.
According to the SEM results, elevation exerted a purely indirect effect on regeneration. First, as the most important intermediary factor, temperature significantly decreased with high elevation and positively influenced the Abies faxoniana regeneration density and the HD ratio of the tallest Abies faxoniana small trees. Second the indirect elevation effects on the Abies faxoniana small tree regeneration density and the HD ratio of the tallest Abies faxoniana small trees were occurred through biotic factors, although the intermediate factor of temperature did not directly affect them. Additionally, Kramer et al. [42] stressed that soil pH may be a major factor that indirectly and positively influences regeneration density, and regeneration density is surprisingly only slightly influenced by elevation. In short, most studies still showed direct and negative relationships between elevation and regeneration density [42,62], although the influence process is still unclear and further studies are needed to elucidate the complex processes.
The relationship between elevation and habitat characteristics is documented by Gale [20]. Accordingly, the proportion of low gaps was negatively correlated with elevation. Unfortunately, we did not find evidence of the influence of elevation on gap characteristics. Such a non-definitive relationship could be caused by the experimental design. In our study, the extensive gaps were not considered and the field survey method of intersect or strip transects may present a clearer relationship between elevation and gap characteristics [3].
In our study, several different results were presented when analyzing the relationships between exogenous predictors and endogenous predictors using SEM and MANOVA analyses. For example, the non-significant difference result of soil total C in different gap sizes was shown through MANOVA analyses, and the significant relationship between gap areas was still revealed. These differences likely originated from three predominant sources. First, the statistical accuracy was different between the two methods. Second, the data used for analysis in the two methods were not completely equivalent. Third, independent variables need to be grouped in MANOVA analyses; however, the data in SEM are set up without grouping. Additionally, several significant indirect effects can be identified via SEMs but not with MANOVA analyses. For example, the direct effect of gap area on Abies faxoniana sapling regeneration density was non-significant in both the SEM and MANOVA analysis results. However, the SEM result indicated that the regeneration density of Abies faxoniana saplings was indirectly affected by the gap area.
4.4. Implications for Management and Gaps
Forest managers striving to identify a balance between protection and utilization must adopt a realistic approach along elevation gradients. For Abies faxoniana regeneration, our results largely confirm the advantages presented by gaps. The negative effect of gap area on the regeneration density of Abies faxoniana (shade tolerant species) saplings seems to overwhelmingly and notably benefit from low-light conditions [15]. This finding supported the lack of effect of a higher PPFD on the regeneration density of Abies faxoniana saplings (Figure 6C). For the regeneration of Abies faxoniana small trees, the results in our study were contrary to that of saplings, and the large gaps were more suitable for the regeneration of Abies faxoniana small trees. Therefore, we hold the opinion that smaller gaps provided a better habitat for designing silvicultural schemes and conducting initial forest management. However, during the late regeneration of Abies faxoniana, its higher growth and regeneration rates would continue in large gaps [27]; therefore, continuous regeneration or low renewal could be promoted by properly increasing the opening area in later management.
The elevation and temperature play important roles in the different life stages of Abies faxoniana regeneration. The regeneration density of Abies faxoniana saplings was more directly affected compared with that of small trees. Furthermore, our SEM further indicated an indirect effect of elevation and temperature through direct and positive biotic factors (e.g., the positive and direct effect of herb coverage on sapling density) effects on Abies faxoniana regeneration. Forest managers cannot easily change the topography or temperature. In these situations, adequate shrub and herb coverage in forest gaps should be considered in forest management.
5. Conclusions
Despite the limited attributes used for predicting regeneration variables and such complex and involved ecological processes in subalpine coniferous forest gaps along an elevation gradient, our study tentatively demonstrates how forest gaps and elevation affect Abies faxoniana regeneration, both directly and indirectly, through the simplified classification of influencing factors and factors associated with important and typical regeneration indices in SEM. Thus, our study showed that gap heterogeneity (forest gap vs. CK plots), gap size, and elevation favored Abies faxoniana regeneration (sapling density, small tree density, and the HD ratio of the tallest Abies faxoniana small trees) by promoting abiotic factors and positive interactions of biotic factors by imposing different effects on the various factors. Our results showed that forest gaps and elevation seem to simultaneously shape Abies faxoniana regeneration by imposing different on the various factors. Forest gaps appear to affect regeneration more directly, elevation is more strongly correlated with indirect effects. Our results also indicated the importance of elevation and temperature for Abies faxoniana sapling regeneration, illustrating the primary role of Abies faxoniana sapling regeneration density on maintaining small tree regeneration density, and highlighting the different effects among elevation, small tree regeneration density, and herb layer cover on the HD ratio of the tallest Abies faxoniana small trees. Furthermore, the use of an appropriately increased gap size during late regeneration and the avoidance of identically sized gaps when carrying out forest management helps promote low renewal and is useful for maintaining continual regeneration.
Author Contributions
L.C. and G.L. conceived and designed the experiments; L.C. and D.L. led field data collection and data analysis; and L.C. wrote the paper.
Funding
This work was supported by the National Key Research and Development Program of China (2016YFC0502102).
Acknowledgments
We are grateful to Soling Shi for his assistance in fieldwork. We thank Huifeng Liu, Ting Li and Xing Wu helping us learns SEM and put forward some advices of modification.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Figure A1.
Relationship between annual mean temperature and elevation in a subalpine coniferous forest. Partial data were used because some thermometers were lost or did not operate correctly in the non-gap plots in our study, and the data for estimations from the regressions equation of elevation–temperature plots were monitored in the non-gap plots from 2013 to 2015 in our study area.
Figure A1.
Relationship between annual mean temperature and elevation in a subalpine coniferous forest. Partial data were used because some thermometers were lost or did not operate correctly in the non-gap plots in our study, and the data for estimations from the regressions equation of elevation–temperature plots were monitored in the non-gap plots from 2013 to 2015 in our study area.
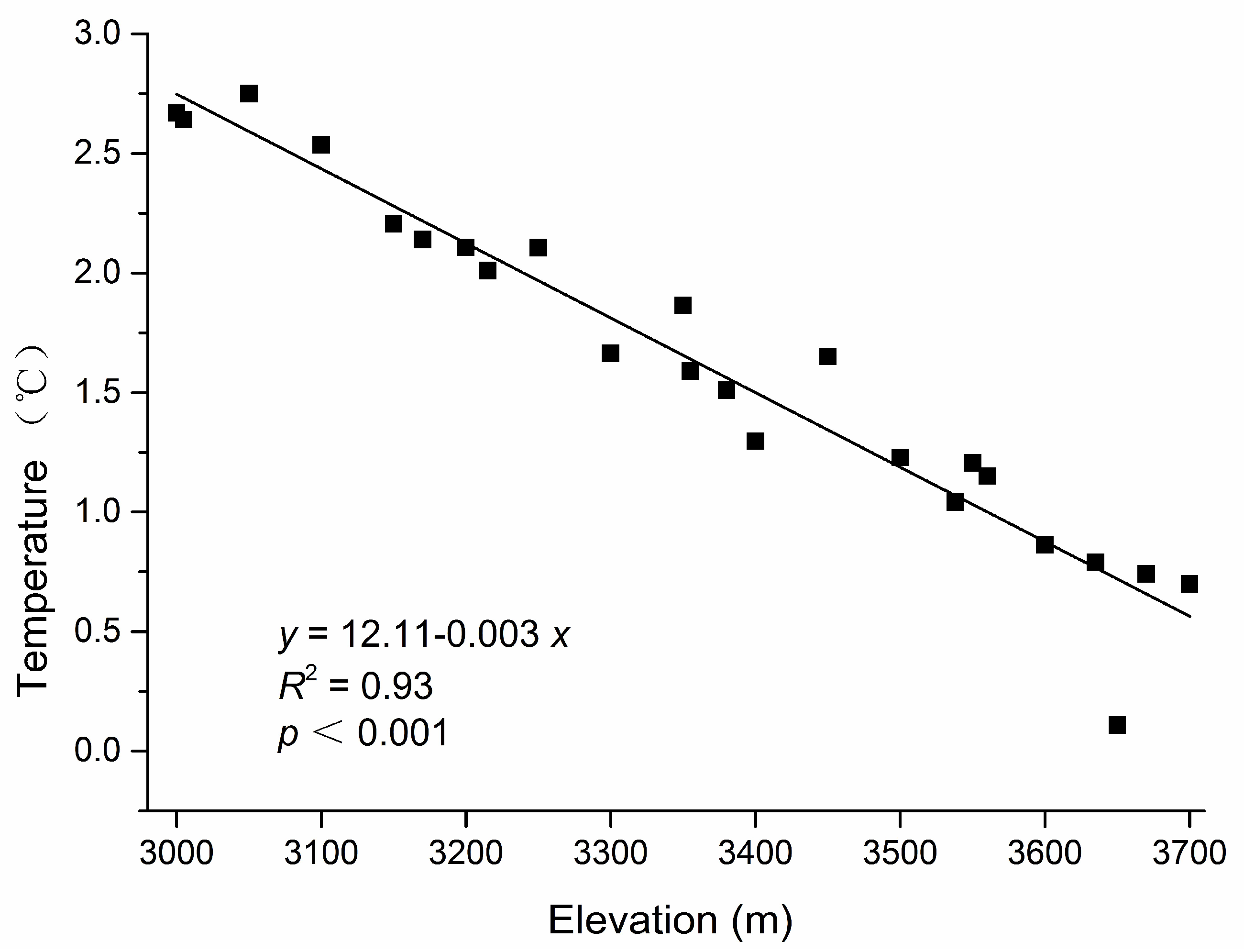
Figure A2.
Relationship between the age and height of Abies faxoniana in forest gaps in a subalpine coniferous forest.
Figure A2.
Relationship between the age and height of Abies faxoniana in forest gaps in a subalpine coniferous forest.

Figure A3.
Gap characteristics (A: Gap area; B: Gap openness; C: Gap age) between the different gap sizes in a subalpine coniferous forest. Error bars represent standard error. Different letters represent significant differences (Tukey’s HSD test) (Bonferroni-adjusted p-values < 0.02). LG, MG, and SG represent a large gap, medium gap, and small gap, respectively.
Figure A3.
Gap characteristics (A: Gap area; B: Gap openness; C: Gap age) between the different gap sizes in a subalpine coniferous forest. Error bars represent standard error. Different letters represent significant differences (Tukey’s HSD test) (Bonferroni-adjusted p-values < 0.02). LG, MG, and SG represent a large gap, medium gap, and small gap, respectively.
References
- Denslow, J.S. Gap partitioning among tropical rainforest trees. Biotropica 1980, 12, 47–55. [Google Scholar] [CrossRef]
- Zhang, C.; Zou, C.J.; Peltola, H.; Wang, K.Y.; Xu, W.D. The effects of gap size and age on natural regeneration of Picea mongolica in the semi-arid region of Northern China. New. For. 2013, 44, 297–310. [Google Scholar] [CrossRef]
- Schliemann, S.A.; Bockheim, J.G. Methods for studying treefall gaps: A review. For. Ecol. Manag. 2011, 261, 1143–1151. [Google Scholar] [CrossRef]
- Nakamura, A.; Morimoto, Y.; Mizutani, Y. Adaptive management approach to increasing the diversity of a 30-year-old planted forest in an urban area of Japan. Landsc. Urban Plan. 2005, 70, 291–300. [Google Scholar] [CrossRef]
- Adamic, M.; Diaci, J.; Rozman, A.; Hladnik, D. Long-term use of uneven-aged silviculture in mixed mountain Dinaric forests: A comparison of old-growth and managed stands. Forestry 2017, 90, 279–291. [Google Scholar] [CrossRef]
- Christensen, N.L.; Bartuska, A.M.; Brown, J.H.; Carpenter, S.; D’Antonio, C.; Francis, R.; Franklin, J.F.; MacMahon, J.A.; Noss, R.F.; Parsons, D.J.; et al. The report of the ecological society of America committee on the scientific basis for ecosystem management. Ecol. Appl. 1996, 6, 665–691. [Google Scholar] [CrossRef]
- Vajari, A.K.; Jalilvand, H.; Pourmajidian, M.R.; Espahbodi, K.; Moshki, A. Effect of canopy gap size and ecological factors on species diversity and beech seedlings in managed beech stands in Hyrcanian forests. J. For. Res. 2012, 23, 217–222. [Google Scholar] [CrossRef]
- Muscolo, A.; Mallamaci, C.; Sidari, M.; Mercurio, R. Effects of gap size and soil chemical properties on the natural regeneration in black pine (Pinus nigra Arn.) Stands. Tree For. Sci. Biotechnol. 2011, 5, 65–71. [Google Scholar]
- Buettel, J.C.; Ondei, S.; Brook, B.W. Look down to see what’s up: A systematic overview of treefall dynamics in forests. Forests 2017, 8, 123. [Google Scholar] [CrossRef]
- Sapkota, I.P.; Oden, P.C. Gap characteristics and their effects on regeneration, dominance and early growth of woody species. J. Plant Ecol. 2009, 2, 21–29. [Google Scholar] [CrossRef]
- Malcom, D.C.; Mason, W.L.; Clarke, G.C. The transformation of conifer forests in Britain- regeneration, gap size and silvicultural systems. For. Ecol. Manag. 2001, 151, 7–23. [Google Scholar] [CrossRef]
- Naaf, T.; Wulf, M. Effects of gap size, light and herbivory on the herb layer vegetation in European beech forest gaps. For. Ecol. Manag. 2007, 244, 141–149. [Google Scholar] [CrossRef]
- Wangchuk, K.; Darabant, A.; Rai, P.B.; Wurzinger, M.; Zollitsch, W.; Gratzer, G. Species richness, diversity and density of vegetation along disturbance gradients in the Himalayan conifer forest. J. Mt. Sci. 2014, 11, 1182–1191. [Google Scholar] [CrossRef]
- Lima, R.A.F.D.; Moura, L.C. Gap disturbance regime and composition in the Atlantic Montane Rain Forest: The influence of topography. Plant Ecol. 2008, 197, 239–253. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- De Montigny, L.E.; Smith, N.J. The effects of gap size in a group selection silvicultural system on the growth response of young, planted Douglas-fir: A sector plot analysis. Forestry 2017, 90, 426–435. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.Z.; Yang, W.Q.; Xu, L.Y.; Ni, X.Y.; He, J.; Tan, B.; Hu, Y. Effects of forest gaps on litter lignin and cellulose dynamics vary seasonally in an alpine forest. Forests 2016, 7, 27. [Google Scholar] [CrossRef]
- Sapkota, I.P.; Tigabu, M.; Oden, P.C. Species diversity and regeneration of old-growth seasonally dry Shorea robusta forests following gap formation. J. For. Res. 2009, 20, 7–14. [Google Scholar] [CrossRef]
- Kang, W.; Tian, C.; Kang, D.W.; Wang, M.J.; Li, Y.X.; Wang, X.R.; Li, J.Q. Effects of gap size, gap age, and bamboo Fargesia denudata on Abies faxoniana recruitment in South-western China. For. Syst. 2015, 24. [Google Scholar] [CrossRef]
- Gale, N. The Relationship between canopy gaps and topography in a western Ecuadorian rain forest. Biotropica 2000, 32, 653–661. [Google Scholar] [CrossRef]
- Runkle, J.R.; Yetter, T.C. Treefalls revisited—Gap dynamics in the southern Appalachians. Ecology 1987, 68, 417–424. [Google Scholar] [CrossRef]
- Taylor, A.H.; Jiang, S.W.; Zhu, L.J.; Liang, C.P.; Miao, C.J.; Huang, J.Y. Regeneration patterns and tree species coexistence in old-growth Abies–Picea forests in southwestern China. For. Ecol. Manag. 2006, 223, 303–317. [Google Scholar] [CrossRef]
- Bossard, C.C.; Cao, Y.; Wang, J.; Rose, A.; Tang, Y. New patterns of establishment and growth of Picea, Abies and Betula tree species in subalpine forest gaps of Jiuzhaigou National Nature Reserve, Sichuan, southwestern China in a changing environment. For. Ecol. Manag. 2015, 356, 84–92. [Google Scholar] [CrossRef]
- Dee, J.R.; Menges, E.S. Gap ecology in the Florida scrubby flatwoods: Effects of time-since-fire, gap area, gap aggregation and microhabitat on gap species diversity. J. Veg. Sci. 2014, 25, 1235–1246. [Google Scholar] [CrossRef]
- Klopcic, M.; Simoncic, T.; Boncina, A. Comparison of regeneration and recruitment of shade-tolerant and light-demanding tree species in mixed uneven-aged forests: Experiences from the Dinaric region. Forestry 2015, 88, 552–563. [Google Scholar] [CrossRef]
- Zhu, J.J.; Matsuzaki, T.; Lee, F.Q.; Gonda, Y. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. For. Ecol. Manag. 2003, 182, 339–354. [Google Scholar] [CrossRef]
- Dekker, M.; van Breugel, M.; Sterck, F.J. Effective height development of four co-occurring species in the gap-phase regeneration of Douglas fir monocultures under nature-oriented conversion. For. Ecol. Manag. 2007, 238, 189–198. [Google Scholar] [CrossRef]
- Zhu, J.J.; Lu, D.L.; Zhang, W.D. Effects of gaps on regeneration of woody plants: A meta-analysis. J. For. Res. 2014, 25, 501–510. [Google Scholar] [CrossRef]
- Kubota, Y.; Murata, H.; Kikuzawa, K. Effects of topographic heterogeneity on tree species richness and stand dynamics in a subtropical forest in Okinawa Island, southern Japan. J. Ecol. 2004, 92, 230–240. [Google Scholar] [CrossRef]
- Grace, J.B.; Pugesek, B.H. A structural equation model of plant species richness and its application to a coastal wetland. Am. Nat. 1997, 149, 436–460. [Google Scholar] [CrossRef]
- Gough, L.; Grace, J.B. Effects of Environmental Change on Plant Species Density: Comparing Predictions with Experiments. Ecology 1999, 80, 882. [Google Scholar] [CrossRef]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. Forestry 2017, 90, 4–17. [Google Scholar] [CrossRef]
- Runkle, J.R. Gap regeneration in some old-growth forests of the Eastern-United-States. Ecology 1981, 62, 1041–1051. [Google Scholar] [CrossRef]
- Sefidi, K.; Mohadjer, M.R.M.; Mosandl, R.; Copenheaver, C.A. Canopy gaps and regeneration in old-growth Oriental beech (Fagus orientalis Lipsky) stands, northern Iran. For. Ecol. Manag. 2011, 262, 1094–1099. [Google Scholar] [CrossRef]
- Beck, J.; Chey, V.K. Explaining the elevational diversity pattern of geometrid moths from Borneo: A test of five hypotheses. J. Biogeogr. 2008, 35, 1452–1464. [Google Scholar] [CrossRef]
- Parent, S.; Messier, C. A simple and efficient method to estimate microsite light availability under a forest canopy. Can. J. Forest. Res. 1996, 26, 151–154. [Google Scholar] [CrossRef]
- Gendron, F.; Messier, C.; Comeau, P.G. Comparison of various methods for estimating the mean growing season percent photosynthetic photon flux density in forests. Agric. For. Meteorol. 1998, 92, 55–70. [Google Scholar] [CrossRef]
- Runkle, J.R. Patterns of disturbance in some old-growth mesic forests of eastern North America. Ecology 1982, 63, 1533–1546. [Google Scholar] [CrossRef]
- Morante-Filho, J.C.; Arroyo-Rodriguez, V.; Lohbeck, M.; Tscharntke, T.; Faria, D. Tropical forest loss and its multitrophic effects on insect herbivory. Ecology 2016, 97, 3315–3325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kissling, W.D.; He, F.L. Local forest structure, climate and human disturbance determine regional distribution of boreal bird species richness in Alberta, Canada. J. Biogeogr. 2013, 40, 1131–1142. [Google Scholar] [CrossRef]
- Huth, F.; Wagner, S. Gap structure and establishment of Silver birch regeneration (Betula pendula Roth.) in Norway spruce stands (Picea abies L. Karst.). For. Ecol. Manag. 2006, 229, 314–324. [Google Scholar] [CrossRef]
- Kramer, K.; Brang, P.; Bachofen, H.; Bugmann, H.; Wohlgemuth, T. Site factors are more important than salvage logging for tree regeneration after wind disturbance in Central European forests. For. Ecol. Manag. 2014, 331, 116–128. [Google Scholar] [CrossRef]
- Tirado, R.; Pugnaire, F.I. Community structure and positive interactions in constraining environments. Oikos 2005, 111, 437–444. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends. Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Maestre, F.T.; Valladares, F.; Reynolds, J.F. Is the change of plant-plant interactions with abiotic stress predictable? A meta-analysis of field results in arid environments. J. Ecol. 2005, 93, 748–757. [Google Scholar] [CrossRef]
- Denslow, J.S. Tropical rainforest gaps and tree species diversity. Annu. Rev. Ecol. Syst. 1987, 18, 431–451. [Google Scholar] [CrossRef]
- Taylor, A.H.; Qin, Z.S. Regeneration patterns in old growth Abies-Betula forests in the Wolong Natural Reserve, Sichuan, China. J. Ecol. 1998, 76, 1204–1218. [Google Scholar] [CrossRef]
- Abd Latif, Z.; Blackburn, G.A. The effects of gap size on some microclimate variables during late summer and autumn in a temperate broadleaved deciduous forest. Int. J. Biometeorol. 2010, 54, 119–129. [Google Scholar] [CrossRef] [PubMed]
- He, Z.S.; Liu, J.F.; Wu, C.T.; Zheng, S.Q.; Hong, W.; Su, S.J.; Wu, C.Z. Effects of forest gaps on some microclimate variables in Castanopsis kawakamii natural forest. J. Mt. Sci. 2012, 9, 706–714. [Google Scholar] [CrossRef]
- Qian, H.; Zhang, Y.J.; Zhang, J.; Wang, X.L. Latitudinal gradients in phylogenetic relatedness of angiosperm trees in North America. Glob. Ecol. Biogeogr. 2013, 22, 1183–1191. [Google Scholar] [CrossRef]
- Aiba, S.I.; Kohyama, T. Tree species stratification in relation to allometry and demography in a warm-temperate rain forest. J. Ecol. 1996, 84, 207–218. [Google Scholar] [CrossRef]
- D’Oliveira, M.V.N.; Ribas, L.A. Forest regeneration in artificial gaps twelve years after canopy opening in Acre State Western Amazon. For. Ecol. Manag. 2011, 261, 1722–1731. [Google Scholar] [CrossRef]
- Kern, C.C.; Montgomery, R.A.; Reich, P.B.; Strong, T.F. Canopy gap size influences niche partitioning of the ground-layer plant community in a northern temperate forest. J. Plant Ecol. 2013, 6, 101–112. [Google Scholar] [CrossRef]
- Bongjoh, C.A.; Mama, N. Early regeneration of commercial timber species in a logged-over forest of southern Cameroon. In Seminar FORAFRI Libreville-Session 2: Knowledge Ecosystem; CIRAD: Montpellier, QC, Canada, 1999; pp. 1–9. [Google Scholar]
- Yin, H.J.; Liu, Q.; Lai, T. Warming effects on growth and physiology in the seedlings of the two conifers Picea asperata and Abies faxoniana under two contrasting light conditions. Ecol. Res. 2008, 23, 459–469. [Google Scholar] [CrossRef]
- Poulson, T.L.; Platt, W.J. Replacement patterns of beech and sugar maple in Warren Woods, Michigan. Ecology 1996, 77, 1234–1253. [Google Scholar] [CrossRef]
- Ritter, E.; Starr, M.; Vesterdal, L. Losses of nitrate from gaps of different sizes in a managed beech (Fagus sylvatica) forest. Can. J. Forest. Res. 2005, 35, 308–319. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Mercurio, R. Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine (Pinus laricio, Poiret) stands. For. Ecol. Manag. 2007, 242, 412–418. [Google Scholar] [CrossRef]
- Taylor, B.R.; Parsons, W.F.J.; Parkinson, D. Decomposition of Populus-Tremuloides Leaf litter accelerated by addition of Alnus-Crispa Litter. Can. J. For. Res. 1989, 19, 674–679. [Google Scholar] [CrossRef]
- Denslow, J.S.; Ellison, A.M.; Sanford, R.E. Treefall gap size effects on above- and below-ground processes in a tropical wet forest. J. Ecol. 1998, 86, 597–609. [Google Scholar] [CrossRef]
- Nagel, T.A.; Svoboda, M.; Rugani, T.; Diaci, J. Gap regeneration and replacement patterns in an old-growth Fagus-Abies forest of Bosnia-Herzegovina. Plant Ecol. 2010, 208, 307–318. [Google Scholar] [CrossRef]
- Van Mantgem, P.J.; Stephenson, N.L.; Keeley, J.E. Forest reproduction along a climatic gradient in the Sierra Nevada, California. For. Ecol. Manag. 2006, 225, 391–399. [Google Scholar] [CrossRef]
Figure 1.
Diagram showing the location, topography, and climate in Wolong National Nature Reserve in southwestern China. The meteorological data were obtained from the neighboring weather station in the Gongga Mountains (29°36′ N, 101°53′ E, 3000 m) in 2014. MP: monthly precipitation (mm); MMAT: monthly mean air temperature (°C); AMAT: annual mean air temperature; AMST: annual mean soil temperature (0–5 cm); AP: annual mean precipitation (mm); AT: air temperature; ST: soil temperature (0–5 cm); NR: net radiation; GR: global radiation; PAR: photosynthetically active radiation; ANR: Annual total net radiation; AGR: Annual total global radiation; APAR: Annual total photosynthetically active radiation.
Figure 1.
Diagram showing the location, topography, and climate in Wolong National Nature Reserve in southwestern China. The meteorological data were obtained from the neighboring weather station in the Gongga Mountains (29°36′ N, 101°53′ E, 3000 m) in 2014. MP: monthly precipitation (mm); MMAT: monthly mean air temperature (°C); AMAT: annual mean air temperature; AMST: annual mean soil temperature (0–5 cm); AP: annual mean precipitation (mm); AT: air temperature; ST: soil temperature (0–5 cm); NR: net radiation; GR: global radiation; PAR: photosynthetically active radiation; ANR: Annual total net radiation; AGR: Annual total global radiation; APAR: Annual total photosynthetically active radiation.

Figure 2.
A priori hypothetical model of SEM. A general model of the connection between the indicators. Habitat characteristics included gap vs. CK and gap area; Abies faxoniana Rehd. et Wils. regeneration included the density of Abies faxoniana saplings, density of Abies faxoniana small trees, and the HD ratio of the tallest Abies faxoniana small trees.
Figure 2.
A priori hypothetical model of SEM. A general model of the connection between the indicators. Habitat characteristics included gap vs. CK and gap area; Abies faxoniana Rehd. et Wils. regeneration included the density of Abies faxoniana saplings, density of Abies faxoniana small trees, and the HD ratio of the tallest Abies faxoniana small trees.

Figure 3.
Data of Abies faxoniana regeneration (density of Abies faxoniana saplings, density of Abies faxoniana small trees, and the HD ratio of the tallest Abies faxoniana small trees) in different habitats (forest and different gap size) for the study area in a subalpine coniferous forest. LG, MG, and SG represent a large gap, medium gap, and small gap, respectively. CK represents non-gap plots. Different letters denote significant difference between treatments (Bonferroni-adjusted p-values < 0.01) by Duncan test.
Figure 3.
Data of Abies faxoniana regeneration (density of Abies faxoniana saplings, density of Abies faxoniana small trees, and the HD ratio of the tallest Abies faxoniana small trees) in different habitats (forest and different gap size) for the study area in a subalpine coniferous forest. LG, MG, and SG represent a large gap, medium gap, and small gap, respectively. CK represents non-gap plots. Different letters denote significant difference between treatments (Bonferroni-adjusted p-values < 0.01) by Duncan test.

Figure 4.
MANOVA analyses results showing endogenous variables difference estimates. Solid lines indicate the endogenous variables were significantly affected by the exogenous variables, and dash lines indicate the non-significant relationship between the endogenous variables and exogenous variables (p ≥ 0.05). Solid line width is weighted by the magnitude of significant coefficients (thick lines represent p < 0.001, and fine lines represent p < 0.05). (A,B) Data include forest gap (N = 26) and CK (N = 8). (C,D) Data only show the difference in forest gaps (N = 26).
Figure 4.
MANOVA analyses results showing endogenous variables difference estimates. Solid lines indicate the endogenous variables were significantly affected by the exogenous variables, and dash lines indicate the non-significant relationship between the endogenous variables and exogenous variables (p ≥ 0.05). Solid line width is weighted by the magnitude of significant coefficients (thick lines represent p < 0.001, and fine lines represent p < 0.05). (A,B) Data include forest gap (N = 26) and CK (N = 8). (C,D) Data only show the difference in forest gaps (N = 26).

Figure 5.
Data of biotic factors (shrub layer cover, herb layer cover, and moss layer thickness) in gaps for the study area in a subalpine coniferous forest along elevation (3000–3700 m). Error bars represent standard error.
Figure 5.
Data of biotic factors (shrub layer cover, herb layer cover, and moss layer thickness) in gaps for the study area in a subalpine coniferous forest along elevation (3000–3700 m). Error bars represent standard error.

Figure 6.
Best structural equation models for predicting Abies faxoniana regeneration. Four models are presented, (A) how gap heterogeneity and elevation shaped density of Abies faxoniana saplings and density of Abies faxoniana small trees; (B) how gap heterogeneity and elevation shaped the HD ratio of the tallest Abies faxoniana small trees; (C) how gap area and elevation shaped density of Abies faxoniana saplings and density of Abies faxoniana small trees; (D) how gap area and elevation shaped the HD ratio of the tallest Abies faxoniana small trees. The variables of Abies faxoniana regeneration in boxes are their variances explained by the model. All path coefficients represent standardized partial regression coefficients. Error variables are not shown in this figure. The exogenous variable of gap heterogeneity is coded as forest gap = 1 and forest = 0. The endogenous variables, as predicted variables, are the plot area (the area of gap and CK) (m2), gap area (m2), gap openness, cover of shrub layer (%), cover of herb layer (%), moss layer thickness (cm), PPFD (μmol m−2 s−1), temperature (°C), soli total C (%), density of Abies faxoniana saplings (N m−2), density of Abies faxoniana small trees (N m−2), and the HD ratio of tallest Abies faxoniana small trees. The shaded arrows represent the negative effect and the black arrows represented the positive effect. The thickness of the arrows indicates the magnitude of correlation between variables and significant coefficients are indicated by asterisk (* p < 0.05, ** p < 0.01, *** p < 0.001).
Figure 6.
Best structural equation models for predicting Abies faxoniana regeneration. Four models are presented, (A) how gap heterogeneity and elevation shaped density of Abies faxoniana saplings and density of Abies faxoniana small trees; (B) how gap heterogeneity and elevation shaped the HD ratio of the tallest Abies faxoniana small trees; (C) how gap area and elevation shaped density of Abies faxoniana saplings and density of Abies faxoniana small trees; (D) how gap area and elevation shaped the HD ratio of the tallest Abies faxoniana small trees. The variables of Abies faxoniana regeneration in boxes are their variances explained by the model. All path coefficients represent standardized partial regression coefficients. Error variables are not shown in this figure. The exogenous variable of gap heterogeneity is coded as forest gap = 1 and forest = 0. The endogenous variables, as predicted variables, are the plot area (the area of gap and CK) (m2), gap area (m2), gap openness, cover of shrub layer (%), cover of herb layer (%), moss layer thickness (cm), PPFD (μmol m−2 s−1), temperature (°C), soli total C (%), density of Abies faxoniana saplings (N m−2), density of Abies faxoniana small trees (N m−2), and the HD ratio of tallest Abies faxoniana small trees. The shaded arrows represent the negative effect and the black arrows represented the positive effect. The thickness of the arrows indicates the magnitude of correlation between variables and significant coefficients are indicated by asterisk (* p < 0.05, ** p < 0.01, *** p < 0.001).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Descriptive statistics for all variables used for the MANOVA analyses and structural equation model (SEM) analyses in our gap study of a subalpine coniferous forest (mean ± SE). The exogenous variables are denoted by * and the endogenous variable are denoted by #. DS, DST, and HDR represent density of Abies faxoniana Rehd. et Wils. saplings, density of Abies faxoniana small trees, and the ratio of height to diameter at breast height (HD ratio) of the tallest Abies faxoniana small trees, respectively; CK represents control check plots (non-gap plots).
Table 1.
Descriptive statistics for all variables used for the MANOVA analyses and structural equation model (SEM) analyses in our gap study of a subalpine coniferous forest (mean ± SE). The exogenous variables are denoted by * and the endogenous variable are denoted by #. DS, DST, and HDR represent density of Abies faxoniana Rehd. et Wils. saplings, density of Abies faxoniana small trees, and the ratio of height to diameter at breast height (HD ratio) of the tallest Abies faxoniana small trees, respectively; CK represents control check plots (non-gap plots).
| Variables | Gap + CK | CK | Gap | ||
|---|---|---|---|---|---|
| Large Gap | Medium Gap | Small Gap | |||
| N | 34 | 8 | 9 | 9 | 8 |
| Elevation * (m) | 3356.0 ± 36.6 | 3383.1 ± 75.7 | 3339.0 ± 80.1 | 3379.0 ± 71.1 | 3322.0 ± 77.0 |
| Habitat characteristic * | |||||
| Area (m2) | 161.20 ± 15.50 | 100.00 ± 0.00 | 294.07 ± 19.13 | 149.64 ± 8.12 | 85.94 ± 5.84 |
| Gap openness | 1.63 ± 0.16 | 1.15 ± 0.09 | 0.97 ± 0.10 | ||
| Gap age (year) | 69.4 ± 1.0 | 71.1 ± 1.4 | 71.3 ± 1.3 | ||
| Biotic factor # | |||||
| Cover of shrub layer (%) | 36.3 ± 4.4 | 25.3 ± 6.5 | 47.3 ± 10.2 | 35.9 ± 8.7 | 35.4 ± 8.8 |
| Cover of herb layer (%) | 42.9 ± 4.6 | 49.6 ± 4.1 | 46.1 ± 7.2 | 39.4 ± 7.3 | 36.5 ± 9.7 |
| Moss layer thickness (cm) | 6.70 ± 0.44 | 6.75 ± 0.31 | 7.22 ± 0.89 | 6.11 ± 1.01 | 6.63 ± 1.20 |
| Abiotic factor # | |||||
| Temperature (°C) | 1.59 ± 0.11 | 1.50 ± 0.23 | 1.64 ± 0.24 | 1.55 ± 0.22 | 1.68 ± 0.23 |
| PPFD (μmol m−2 s−1) | 216.44 ± 15.87 | 54.75 ± 2.80 | 264.89 ± 8.64 | 262.67 ± 1.13 | 271.63 ± 5.54 |
| Soil total C (%) | 7.45 ± 0.52 | 6.74 ± 0.74 | 6.14 ± 0.96 | 8.40 ± 1.37 | 6.62 ± 0.92 |
| Soil total N (%) | 0.45 ± 0.02 | 0.43 ± 0.30 | 0.44 ± 0.06 | 0.52 ± 0.05 | 0.39 ± 0.03 |
| Abies faxoniana regeneration # | |||||
| DS (N m−2) | 0.63 ± 0.05 | 0.32 ± 0.05 | 0.61 ± 0.08 | 0.73 ± 0.09 | 0.82 ± 0.13 |
| DST (N m−2) | 0.37 ± 0.05 | 0.07 ± 0.01 | 0.64 ± 0.09 | 0.37 ± 0.06 | 0.36 ± 0.06 |
| HDR | 90.10 ± 2.44 | 80.13 ± 3.02 | 97.54 ± 6.02 | 92.20 ± 3.87 | 89.344.56 |
Table 2.
MANOVA results comparing effects of gap vs. CK and gap size. F and p values are derived from analysis of variance. PPFD, MAT, DS, DST and HD ratio TSD represent photosynthetic photon flux density, mean annual temperature, density of Abies faxoniana saplings, density of Abies faxoniana small trees and HD ratio of the tallest Abies faxoniana small trees, respectively. Values in bold are different from 0 with a significance level alpha = 0.05.
Table 2.
MANOVA results comparing effects of gap vs. CK and gap size. F and p values are derived from analysis of variance. PPFD, MAT, DS, DST and HD ratio TSD represent photosynthetic photon flux density, mean annual temperature, density of Abies faxoniana saplings, density of Abies faxoniana small trees and HD ratio of the tallest Abies faxoniana small trees, respectively. Values in bold are different from 0 with a significance level alpha = 0.05.
| Variables | Gap vs. CK | Elevation | Gap Size | Elevation | ||||
|---|---|---|---|---|---|---|---|---|
| (Ngap = 26, NCK = 8) | N = 34 | (Nlarge = 9, NMedium = 9, NSmall = 7) | N = 26 | |||||
| F | p | F | p | F | p | F | p | |
| gap openness | 5.985 | 0.044 | 0.874 | 0.570 | ||||
| gap age | 0.540 | 0.613 | 0.984 | 0.518 | ||||
| PPFD | 406.434 | <0.001 | 2.43 | 0.174 | 0.839 | 0.485 | 3.165 | 0.113 |
| MAT | 0.745 | 0.570 | 140.654 | <0.001 | 0.789 | 0.504 | 157.861 | <0.001 |
| soil total N | 0.409 | 0.754 | 0.273 | 0.927 | 0.613 | 0.578 | 0.419 | 0.840 |
| soil total C | 0.253 | 0.857 | 1.185 | 0.436 | 0.377 | 0.704 | 1.295 | 0.397 |
| soil water content | 0.216 | 0.881 | 0.788 | 0.615 | 0.041 | 0.960 | 1.369 | 0.374 |
| moss thickness | 0.593 | 0.646 | 3.049 | 0.050 | 0.879 | 0.471 | 5.536 | 0.028 |
| cover of shrub | 4.772 | 0.063 | 17.961 | 0.003 | 3.888 | 0.096 | 20.784 | 0.002 |
| cover of herb | 3.262 | 0.118 | 15.694 | 0.004 | 3.05 | 0.136 | 21.057 | 0.002 |
| DS | 30.819 | 0.001 | 21.803 | 0.002 | 0.779 | 0.510 | 25.887 | 0.001 |
| DST | 82.595 | <0.001 | 20.281 | 0.002 | 38.494 | 0.001 | 27.657 | 0.001 |
| HD ratio of TSD | 5.962 | 0.042 | 10.154 | 0.011 | 1.999 | 0.23 | 10.443 | 0.010 |
Table 3.
Values of the fit indices of four structural equation models for Abies faxoniana regeneration in a subalpine coniferous forest. For information associated with Models A, B, C, and D see Figure 5. DF, RMSEA, GFI and CFI represent degree of freedom, root mean square error of approximation, goodness fit index and comparative fit index, respectively.
Table 3.
Values of the fit indices of four structural equation models for Abies faxoniana regeneration in a subalpine coniferous forest. For information associated with Models A, B, C, and D see Figure 5. DF, RMSEA, GFI and CFI represent degree of freedom, root mean square error of approximation, goodness fit index and comparative fit index, respectively.
| Models | Model Fit Indices | ||||||
|---|---|---|---|---|---|---|---|
| χ2 | DF | p | RMSEA | GFI | CFI | AIC | |
| A | 4.727 | 10 | 0.909 | <0.001 | 0.959 | 1 | 40.727 |
| B | 3.675 | 6 | 0.721 | <0.001 | 0.963 | 1 | 33.675 |
| C | 8.505 | 11 | 0.668 | <0.001 | 0.915 | 1 | 58.505 |
| D | 3.689 | 8 | 0.869 | <0.001 | 0.948 | 1 | 29.869 |
Table 4.
Standardized total effect, indirect effect and direct effect of predictor variables on Abies faxoniana regeneration for the best SEMs shown in Figure 2. HY, T, PPFD, PA, SC, GA, STC, GO, HC, MLT, DS, DST, and HDR represent gap heterogeneity, temperature, photosynthetic photon flux density, plot area, shrub coverage, gap area, soil total C, gap openness, herb coverage, the moss layer thickness, density of Abies faxoniana saplings, density of Abies faxoniana small trees, and the HD ratio of the tallest Abies faxoniana small trees, respectively.
Table 4.
Standardized total effect, indirect effect and direct effect of predictor variables on Abies faxoniana regeneration for the best SEMs shown in Figure 2. HY, T, PPFD, PA, SC, GA, STC, GO, HC, MLT, DS, DST, and HDR represent gap heterogeneity, temperature, photosynthetic photon flux density, plot area, shrub coverage, gap area, soil total C, gap openness, herb coverage, the moss layer thickness, density of Abies faxoniana saplings, density of Abies faxoniana small trees, and the HD ratio of the tallest Abies faxoniana small trees, respectively.
| Regeneration Indexes | Gap + CK | Gap | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Elevation | HY | T | PPFD | PA | SC | DS | Elevation | GA | T | STC | GO | HC | MLT | DS | DST | ||
| Total effect | |||||||||||||||||
| DS | −0.74 | 0.47 | 0.79 | −0.71 | −0.27 | - | 0 | −0.82 | −0.36 | 0.61 | 0.10 | 0.09 | 0.27 | - | 0 | - | |
| DST | −0.53 | 0.60 | 0.56 | −0.51 | 0.46 | - | 0.71 | −0.59 | 0.65 | 0.44 | 0.07 | 0.06 | 0.19 | - | 0.72 | - | |
| HDR ratio | −0.43 | 0.36 | 0.43 | 0.27 | - | 0.53 | - | −0.63 | 0.29 | 0.43 | - | - | - | 0.35 | - | 0.56 | |
| Direct effect | |||||||||||||||||
| DS | 0 | 1.28 | 0.79 | −0.71 | −0.27 | 0 | 0 | −0.43 | 0.61 | 0 | 0 | 0.27 | - | 0 | - | ||
| DST | 0 | 0 | 0 | 0 | 0.65 | 0.71 | 0 | 0.91 | 0 | 0 | 0 | 0 | - | 0.72 | - | ||
| HDR ratio | 0 | 0 | 0 | 0.27 | 0.53 | 0 | 0 | 0 | - | - | - | 0.35 | - | 0.56 | |||
| Indirect effect | |||||||||||||||||
| DS | −0.74 | −0.81 | 0 | 0 | 0 | - | 0 | −0.82 | 0.07 | 0 | 0.10 | 0.09 | 0 | - | - | - | |
| DST | −0.53 | 0.60 | 0.56 | −0.51 | −0.19 | - | 0 | −0.59 | −0.26 | 0.44 | 0.07 | 0.06 | 0.19 | - | 0 | - | |
| HDR | −0.43 | 0.36 | 0.43 | 0 | - | 0 | - | −0.63 | 0.29 | 0.43 | - | - | - | 0 | - | 0 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, L.; Liu, G.; Liu, D. How Forest Gap and Elevation Shaped Abies faxoniana Rehd. et Wils. Regeneration in a Subalpine Coniferous Forest, Southwestern China. Forests 2018, 9, 271. https://doi.org/10.3390/f9050271
AMA Style
Chen L, Liu G, Liu D. How Forest Gap and Elevation Shaped Abies faxoniana Rehd. et Wils. Regeneration in a Subalpine Coniferous Forest, Southwestern China. Forests. 2018; 9(5):271. https://doi.org/10.3390/f9050271
Chicago/Turabian StyleChen, Li, Guohua Liu, and Dan Liu. 2018. "How Forest Gap and Elevation Shaped Abies faxoniana Rehd. et Wils. Regeneration in a Subalpine Coniferous Forest, Southwestern China" Forests 9, no. 5: 271. https://doi.org/10.3390/f9050271
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.