Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion
by
Anthony W. D’Amato
1,*, 
Brian J. Palik
2,
Robert A. Slesak
3,
Greg Edge
4,
Colleen Matula
4 and
Dustin R. Bronson
4 1
Rubenstein School of Environment and Natural Resources, University of Vermont, 204e Aiken Center, 81 Carrigan Drive, Burlington, VT 05405, USA
2
USDA Forest Service Northern Research Station, 1831 Hwy 169 East, Grand Rapids, MN 55744, USA
3
Minnesota Forest Resources Council, 1530 Cleveland Ave. North, St. Paul, MN 55108, USA
4
Wisconsin Department of Natural Resources, 101 S. Webster St., Madison, WI 53703, USA
*
Author to whom correspondence should be addressed.
Forests 2018, 9(6), 348; https://doi.org/10.3390/f9060348
Submission received: 18 April 2018
/
Revised: 9 June 2018
/
Accepted: 12 June 2018
/
Published: 13 June 2018
(This article belongs to the Special Issue Understanding and Managing Emerald Ash Borer Impacts on Ash Forests)
Abstract
:The arrival and spread of emerald ash borer (EAB) across the western Great Lakes region has shifted considerable focus towards developing silvicultural strategies that minimize the impacts of this invasive insect on the structure and functioning of black ash (Fraxinus nigra) wetlands. Early experience with clearcutting in these forests highlighted the risks of losing ash to EAB from these ecosystems, with stands often retrogressing to marsh-like conditions with limited tree cover. Given these experiences and an urgency for increasing resilience to EAB, research efforts began in north-central Minnesota in 2009 followed by additional studies and trials in Michigan and Wisconsin to evaluate the potential for using regeneration harvests in conjunction with planting of replacement species to sustain forested wetland habitats after EAB infestations. Along with these more formal experiments, a number of field trials and demonstrations have been employed by managers across the region to determine effective ways for reducing the vulnerability of black ash forest types to EAB. This paper reviews the results from these recent experiences with managing black ash for resilience to EAB and describes the insights gained on the ecological functioning of these forests and the unique, foundational role played by black ash.
1. Introduction
Novel stressors, such as non-native insects and diseases, present a significant challenge to the long-term sustainable management of forest ecosystems around the globe [1]. For example, recent estimates for the United States indicate that almost two thirds of all forestlands are susceptible to damage from non-native insects and diseases already present in the country [2]. In many cases, these organisms are host-specific with the ability to functionally eliminate a given tree species from an ecosystem [3]. As such, there is a significant need for the development of adaptive management strategies that increase forest resilience to the loss of a constituent tree species as a way to maintain key ecosystem functions after infestation by non-native species [4]. This need becomes urgent when the host species is considered foundational to ecosystem functioning, such that its loss cannot readily be absorbed by other species in the ecosystem [5].
Forest management guidelines developed in response to non-native insects and diseases have generally focused on altering host species structure and abundance to minimize spread and impacts of a given organism. These approaches may include removing all potential host species from forested areas within a set distance of known infestations [6,7] or selective removal of larger host trees to reduce availability of brood trees [8,9]. Although these strategies are warranted in areas proximal to introduction points and ongoing infestations, their application in areas not currently threatened may generate greater ecological and economic impacts than the non-native forest pest itself (e.g., [10]). To date, less attention has been devoted to developing appropriate silvicultural strategies for increasing the resilience of these unimpacted areas to maintain long-term management options and ecological functions even after a pest’s establishment.
The introduced emerald ash borer (Agrilis planipennis Fairmaire [EAB]) is one of the more significant non-native insects threatening North American forests given the importance of ash species across numerous forest types and regions [11], and the ability of this insect to cause widespread mortality within 2–6 years of invasion [12]. Most ash species exist as minor components of mixed species forests [11] and loss of canopy ash to EAB will eliminate the unique ecological functions of ash, but forested conditions are expected to persist in these affected ecosystems. One exception are black ash (Fraxinus nigra Marsh.)-dominated lowland forests in the western Great Lakes region, where black ash often forms almost pure stands, with up to 95% of stems composed of this species [5]. As black ash is a foundational species in these forests, expectations regarding EAB impacts include not only a loss of canopy black ash, but also a concomitant shift in ecosystem structure and function to marsh-like conditions with little to no tree component [13,14]. In particular, historic experience with clearcutting and recent field simulations of EAB impacts indicate that the loss of canopy black ash may cause the water table to rise and increase the duration of water at or near the soil surface [13,14]. These changes represent not only a significant threat to organisms and processes associated with current black ash ecosystems [5], but also a significant challenge to managers tasked with sustaining these ecosystems in the face of the threat of EAB invasion.
As with other forest types threatened by non-native insects and diseases, the management guidance for black ash forests was developed based on research and management experience developed prior to the introduction of EAB. This historical guidance largely focused on silvicultural strategies that encouraged the dominance of black ash over time [13], a condition in contemporary contexts that is highly vulnerable to EAB impacts. Recent management recommendations for addressing EAB in these and other ash forest types have primarily focused on minimizing establishment and spread of the insect through reducing host abundance [8,9]. Although these approaches, when applied in conjunction with insecticides, may reduce spread of EAB in localized, urban settings, they represent a reactive strategy to a current outbreak as opposed to a proactive approach that aims to increase resilience to impacts prior to invasion. Moreover, black ash can dominate extensive areas (1–40 ha) presenting significant logistical challenges for widespread application of insecticides to protect all black ash forests. Given the potential for significant ecosystem phase shifts following EAB mortality or pre-emptive liquidation harvests in lowland black ash forests (cf. [13]), there is a great need for silvicultural strategies that reduce vulnerability of these ecosystems to EAB prior to its arrival. In addition, given the continued spread of this insect and the infeasibility of treating all black ash-dominated areas, there may also be a growing need for rehabilitation approaches to reintroduce forest cover to places where the insect or associated reactive management have resulted in a shift to non-forest conditions.
The overall objective of this paper is to review recent research and experience managing black ash ecosystems to provide general guidance for the adaptive management of these ecosystems in the face of EAB invasion. This includes describing the general ecological setting of these forests as a foundation for adaptive silviculture prescriptions, summarizing insights from long-term research trials evaluating the effectiveness of different silvicultural treatments at increasing the non-ash components in these forests, and discussing potential rehabilitation approaches for returning other species to these areas after EAB has eliminated black ash. To date, EAB has spread to 32 states in the US and three Canadian provinces since its discovery in Michigan in 2002, and has killed nearly 100% of ash stems >2.5 cm diameter at breast height (DBH; 1.3 m) in areas proximal to where it was first introduced [15]. In most cases, the majority of ash trees are killed within 2–6 years of invasion; however, individual trees and stump sprouts have persisted for over a decade in some green ash (Fraxinus pennsylvanica Marsh.)-dominated wetlands in southern Michigan, indicating areas may not always experience complete ash mortality following invasion [16]. As such, the management recommendations reviewed in this work are not definitive given the great uncertainty around long-term EAB impacts and silvicultural outcomes, nor is it expected that they can be applied across all black ash-dominated areas and ownerships. Instead, they are meant provide a framework for approaching this threat to increase resilience and sustain a forested condition over the long term, particularly given the significant losses of ash observed in all ecosystem types impacted by EAB (>62–100% of basal area; [15,16]). At the time of writing, EAB had not been detected in the areas serving as the focus for this review (northern MI, MN, and WI) and the experiments and recommendations largely assumed EAB arrival within the next 10–15 years based on location of known infestations (southern MN and WI, eastern Upper Peninsula of Michigan) relative to expected rates of spread [17].
2. Ecological Context of Black Ash Wetlands
A central challenge to developing adaptive management approaches for addressing the threat of EAB is the limited amount of information on the ecology and management of black ash forests relative to other forest ecosystems in the Great Lakes region. Black ash has long been a significant component of the cultures and livelihoods of Native Americans and First Nations people across the range of this species [15]. However, its lower historic value to the timber industry in the Great Lakes region resulted in greater historical emphasis of research on more productive, upland forest types, as well as lowland conifers. The introduction of EAB to the region in the early 2000s motivated considerable effort towards generating a greater understanding of the ecology and dynamics of these forests as a foundation for developing appropriate management responses. The following section summarizes our current understanding of the variety in black ash forest habitats as well as several key findings from recent investigations into the hydrology and regeneration dynamics following episodic dieback and their relevance to adaptive management in the context to EAB.
2.1. Black Ash Forest Habitat Types
Black ash wetland forests are complex and exist across a wide range of biophysical settings covering over 1 million ha in Minnesota, Michigan, and Wisconsin [18]. Early work describing the vegetation and silviculture of these forests recognized this complexity and demonstrated the importance of water table depth and nutrient conditions in affecting regeneration dynamics and forest composition [13,19,20]. Nevertheless, historical management guidance and forest classification systems often lumped all black ash-dominated forests into a single “black ash swamp” or “lowland hardwood” category with little consideration for the important variations in soil, nutrient, and vegetation dynamics that occur in these ecosystems [21,22,23]. The advent of forest habitat type classification systems (FHTCS) for the Lake States region during the 1980s and 1990s resulted in a greater appreciation for the range in black ash-dominated forests that existed across the landscape. In particular, they generated a framework that has been critical for recognizing variations in soil, moisture, and vegetation conditions that exist across sites when developing management strategies for addressing EAB [20,24,25].
The first FHTCS to describe the variation in black ash communities were developed in Minnesota, owing in part to the comparatively high abundance of black ash forests in this state relative to Wisconsin and Michigan [20]. Two main black ash forest types, Wet Ash Forests (WFn55) and Very Wet Ash Forests (WFn64), were described by this FHTCS and both are dominated by >75% black ash in the canopy and understory layers [25]. The variation between these two communities is driven by the influence of flooding duration and organic soil depth in affecting nutrient dynamics. The primary distinguishing features between WFn64 and WFn55 communities are peaty versus mineral soils and a greater duration of standing water during the growing season in WFn64 communities. These differences have important management implications, as even the creation of larger group selection openings (0.2 ha) has led to a rise in local water tables and loss of tree cover in areas classified as WFn64 communities (Peter Bundy, personal communication). The importance of overstory tree cover in regulating water table dynamics during recruitment events has also been reinforced by examinations of the age structure of old-growth examples for these two habitat types, which indicate these systems are strongly multi-aged with historic development influenced primarily by gap-scale disturbances (Figure 1; D’Amato et al. in prep). A similar multi-aged population structure was also documented across numerous old-growth black ash riparian forests in the Lake Duparquet region of Quebec [26].
In contrast to the FHTCS developed for Minnesota, early classification systems developed for Wisconsin and Michigan focused primarily on more productive upland forest types leaving key knowledge gaps regarding lowland forest, including black ash ecosystems [24]. The arrival of EAB to the region motivated renewed focus on these ecosystems and led to the development of the Wisconsin Wetland Forest Habitat Type guide using methods and interpretation similar to the forest habitat type guide for upland forests [27]. This guide includes management interpretations provided by the Wisconsin Department of Natural Resources (WDNR; [27]) and has been an important tool for foresters across the state in interpreting complex, wetland forest systems.
The wetland habitat typing system developed for Wisconsin covers five biophysical regions in the northern portion of the state and recognizes 12 lowland Fraxinus (black ash) types that vary in understory vegetation, nutrient availability, soils, and hydrology [27]. Consistent with the silvics of the species, black ash-dominated habitat types are generally characterized by medium rich nutrient regimes (e.g., Figure 2). The Black Ash–Balsam Fir–Red Maple/Sensitive Fern (FnAbArOn) type is the most widespread black ash habitat type in Wisconsin and is characterized by organic soils that can vary from 30 to 90 cm in depth overlaying mineral soils [27]. Habitat type field monitoring of these areas identified organic soil depth as a key driving factor of nutrient availability and duration of flooding, with deeper organic soils having greater potential for hydrological changes and rutting associated with timber harvest. As with the Minnesota habitat types, the less rich habitat types tend to regenerate predominately to black ash following harvesting and have become areas where artificial regeneration is being considered to enhance the non-ash components [28].
Ash-dominance is a common feature across abovementioned habitat types developed in Minnesota and Wisconsin; however, the abundance of non-ash species in the overstory can be quite variable within and across habitat types reflecting variations in local site conditions and land-use history. For example, non-ash species, including Quercus macrocarpa, Ulmus americana, and Abies balsamea, made up 16–30% of the basal area across old-growth WFn64 communities in Minnesota [29]. Similarly, black ash abundance ranged from 25% to 85% of basal area in isolated, depressional wetlands in the Upper Peninsula of Michigan, with Betula alleghaniensis and Acer rubrum often constituting the other primary overstory species [30]. The variation in these community types, particularly in relation to the amount of non-ash trees present in the overstory and regeneration layers, is an important consideration when determining vulnerability to EAB impacts and developing appropriate management responses [31]. Much of the experience with managing black ash forests for resilience to EAB has been in the context of the most vulnerable conditions and habitat types in MN and WI (i.e., >70% black ash) and application of these approaches to other systems and regions will require adaptation to local site conditions and levels of non-ash in the canopy.
2.2. Hydrologic Setting of Black Ash Forests
Black ash wetlands occur in a number of hydrogeomorphic settings that span a range of scales including large expansive complexes [14], floodplains [32], areas along streams and rivers [33], and in topographic low depressions [34,35]. Topography and its influence on drainage is a primary factor influencing wetland hydrology, but soil parent material can also play a large role. In the large expansive wetlands of northern Minnesota, presence of a dense layer of thick clay results in a perched water table that is the primary driver of hydrology. Soils range from deep organic peats (especially in depressional wetlands) to mineral soils overlain by shallow layers of muck, but all are poorly to very-poorly drained. Water source is primarily from precipitation, either as direct rainfall or snow inputs [35] or indirect from localized drainage [34].
Depending on the dominant hydrological process(es), hydroperiod length in black ash wetlands can be quite variable, but most sites are characterized as having water tables at or above the soil surface during a portion of the growing season. Wetlands fed by consistent groundwater sources, whether from deep regional flows or areas occurring at topographic lows in the local landscape, may have very little water level variability throughout the year with prolonged surface water presence [34]. In large expansive wetland complexes, water level variability is typically intermediate, where water tables are generally at or above the surface in spring, followed by a large drawdown in early summer until a stable minimum is reached later in the season (Figure 3).
The dominance of black ash (typically greater than 80% of the canopy; [29]) in wetland settings reflects its unique physiology that allows for transpiration during periods of inundation [36,37]. Because of this, black ash strongly regulates the hydrologic regime with water table drawdown occurring rapidly following black ash leaf expansion in the spring (Figure 3). Evidence of this biological influence is clearly evident in the diurnal water table signature, with pronounced drawdown during the day when trees transpire followed by slight recharge or stability during the night (Figure 3 inset). Black ash transpiration (and its influence on overall evapotranspiration [ET]) is thought to be greatest when water tables are below the surface and at intermediate depths (~50 cm, [38]).
Given the role of ET in site hydrology, a reduction in black ash transpiration is likely to alter water table dynamics, causing the water table to rise and increase the duration of water at or near the soil surface [13,14]. Because of this, black ash wetlands are commonly believed to be highly susceptible to hydrologic alteration following EAB mortality [14,34]. Slesak et al. [14] used experimental girdling to mimic EAB mortality, which caused the water table to be elevated and near the soil surface for longer periods of time relative to control stands. Using a similar approach but in a different hydrogeomorphic setting, Van Grinsven et al. [34] found the effect of black ash loss was more muted, largely because the hydrologic regime was influenced by continuous subsurface inflow associated with topographic position in the landscape. Although there are clear effects of ash loss on water table dynamics [13,14], the magnitude and duration of these effects is not clear. Recent work by Diamond et al. [39] showed that clearcutting caused a distinct change in hydrologic regime that persists for at least five years. Although hydrologic alteration at the site scale is likely following black ash loss, it is not clear how such changes will alter hydrology at larger scales such as the watershed.
2.3. Black Ash Decline and Natural Regeneration Potential for Non-Ash Species
Finding effective adaptive management approaches for black ash ecosystems in the face of EAB is complicated by a long-standing episodic dieback/decline problem that occurs throughout much of the range of black ash, including the Lake States and northeastern United States [40,41,42]. Most dieback episodes are not associated with a specific disease or insect pest, but include areas of crown dieback and tree mortality, particularly on more hydric sites [42]. In fact, the cause is still in doubt, with speculation that it relates to spring drought [41,43] or, conversely, overly wet conditions [42,44]. Regardless, dieback episodes can affect large areas of black ash with over 9000 ha affected in Minnesota in 2009 alone [41].
Regardless of cause, examining successional potential in declining stands can provide insight into the future of EAB impacted black ash wetlands. Unfortunately, this future may not be bright. While black ash regenerates prolifically from stump-sprouts [13], and research has documented high numbers of (likely) sprout-origin black ash seedlings after harvesting [45,46,47], densities may decline significantly as trees grow into larger sizes [22,48], potentially limiting self-replacement. Moreover, this regeneration presumably will suffer the same fate as canopy black ash, i.e., death from EAB after a susceptible size is reached (approximately 2.5 cm diameter).
Natural successional replacement of black ash after EAB mortality may be limited as well. In Minnesota wetlands, for instance, canopy species other than black ash occur in very low densities, generally much less than 20% of relative density [23,48,49]. Consequently, there is not a high potential for replacement of EAB killed black ash from the current sapling or tree layers. Moreover, the regeneration layer in these forests is generally dominated by speckled alder (Alnus incana [L.] Moench) and beaked hazel (Corylus cornuta Marsh.), with only limited abundance of canopy potential species [48]. This regeneration bottleneck may be exacerbated with loss of black ash from EAB if, as discussed above, this results in an upward fluctuating water table, further increasing the potential for regeneration failures.
3. Management Experience with Strategies for Increasing Resilience of Black Ash Wetlands to EAB
3.1. Research Trials Examining Artificial Regeneration Options to Increase Non-Ash Components
The importance of black ash in many lowland habitat types and the hydrological responses observed following the loss of this species to harvesting or decline episodes, led to the establishment of several adaptive silvicultural studies focused on strategies for increasing ecosystem resilience in advance of EAB. A primary component of this work was the evaluation of artificial regeneration of non-ash species as a strategy to maintain tree cover following the loss of overstory ash. Although there is no true cultural or ecological replacement for black ash, the emphasis of these studies was to evaluate approaches for replacing the core ecological functions provided by black ash (e.g., hydrologic regulation, mature tree habitat, live-tree carbon storage) so as to maintain forested wetland conditions following infestation by EAB. The first of these studies was established in north-central Minnesota in a region containing the greatest concentration of lowland black ash forests in the US [14]. This work included canopy treatments intended to emulate EAB (girdling of all ash in 1.4 ha areas), potential management responses to the threat of EAB (group selection and clearcut harvests), and untreated controls, all in conjunction with operational plantings of 12 different tree species [49]. A later, companion study was established based off this work in smaller, depressional black ash wetlands in the Upper Peninsula of Michigan with a specific focus on the effect of planting microsite on seedling survival [50]. Finally, planting trials of non-ash species in riparian black ash systems in Wisconsin were developed to evaluate the effectiveness of underplanting several non-ash species to increase resilience of these forests [50].
The wide range of species evaluated as potential replacements for black ash largely reflects the great uncertainty and limited prior experience with planting trees in these lowland forests. The majority of these species commonly co-occur at low abundance in black ash wetlands, including American elm (Ulmus americana), red maple (Acer rubrum), northern white cedar (Thuja occidentalis), bur oak (Quercus macrocarpa), yellow birch (Betula alleghaniensis), balsam poplar (Populus balsamifera), basswood (Tilia americana), white spruce (Picea glauca), tamarack (Larix laricina [Du Roi] K. Koch), balsam fir (Abies balsamea), and trembling aspen (Populus tremuloides Michx; Table 1). Other species found in wetland forests in the study regions of interest have also been included in these trials, including black spruce (Picea mariana (Mill.) Britton, Sterns & Poggenb.), cottonwood (Populus deltoides W. Bartram ex Marshall), and American sycamore (Platanus occidentalis L.). Wetland species currently found in the climate zone immediately south of these areas, including swamp white oak (Quercus bicolor Willd.), hackberry (Celtis occidentalis L.), and silver maple (Acer saccharinum L.) have also been evaluated in an attempt to anticipate both EAB and climate change impacts [49,50]. Finally, Manchurian ash (Fraxinus mandschurica Rupr.) has been included in Minnesota trials due to its resistance to EAB and potential to serve as a replacement for traditional Native American basket making materials historically provided by black ash (G. Swanson, personal communication).
Table 1 summarizes general recommendations for artificial regeneration of non-black ash species based on early patterns in seedling survival and growth. Survival has been generally quite low for most species examined (e.g., <40%; [49]); however, swamp white oak has consistently performed well in a variety of black ash habitat types (e.g., Figure 4). This greater survival (>75%; [49]) is consistent with findings from planting trials in simulated floodplain conditions that indicate high potential for this species for reforestation efforts in seasonally flooded environments [51]. Swamp white oak early height growth rates have also been similar to those documented in other planting trials and have ranged from 5 to 12 cm year−1 [51]. Within species, seedling survival has largely been greatest under an undisturbed or partial black ash canopy, reflecting the importance of ash in regulating water table dynamics during the establishment phase and reinforcing historic recruitment dynamics in these forests (e.g., Figure 1; [49]). In addition, overstory ash may also limit levels of competition experienced by planted seedlings, as loss of ash often leads to an increased abundance of understory species, including lake sedge and speckled alder (Looney et al., 2017). Planting location has also influenced seedling success with planting on hummocks significantly increasing the survival of more mesic species, such as American basswood, given the shorter duration of seedling exposure to flooding on these microsites [50].
Given that few of the species selected for the adaptive approaches being tested are widely planted in operational settings, there has been limited opportunity to evaluate how stock type and size affects species performance. General patterns suggest larger, bare-root or containerized stock have generally had greater survival and growth when planted in black ash wetlands (Table 1). This includes large, containerized Dutch elm disease (Ophiostoma novo-ulmi Brasier)-tolerant American elm, which has also shown promise in other adaptive plantings to address EAB threats to riparian green ash (Fraxinus pennsylvanica) forests in Ohio [52]. Black ash wetlands often have dense understory layers of shrubs and herbaceous species [47] and larger stock sizes may provide a competitive advantage and increase survival on these sites. This dynamic was reflected in a black ash silviculture field trial in Wisconsin in which competition was controlled at the time of planting. In this context, there was no appreciable difference in survival between containerized and bare-root tamarack and black spruce seedlings (>60% survival after three years), whereas both species have fared poorly when planted as containerized stock in other trials lacking competition control (<20% survival; [43]).
3.2. Encouraging Natural Regeneration of Non-Ash Species
There is great uncertainty regarding how effective and operationally feasible planting non-ash species is for increasing the resilience across the large expanse of black ash forests in the Great Lakes region. As a result, there is also considerable interest in the potential for using different regeneration methods to encourage the natural regeneration and recruitment of non-ash species to diversify overall composition and structure. A key source of information on the response of black ash wetlands to different silviculture practices has been the Swamp Hardwood Silviculture Trials established by the Wisconsin DNR beginning in 1977 (Figure 5; [53]). These trials were established prior to the introduction of EAB and correspondingly were focused on developing the most effective strategies for managing and maintaining black ash-dominated forests [28]. Since the introduction of EAB to the region, these trials have been revisited to evaluate the relative effectiveness of different strategies at increasing the non-ash component in these forests, while also maintaining hydrological function.
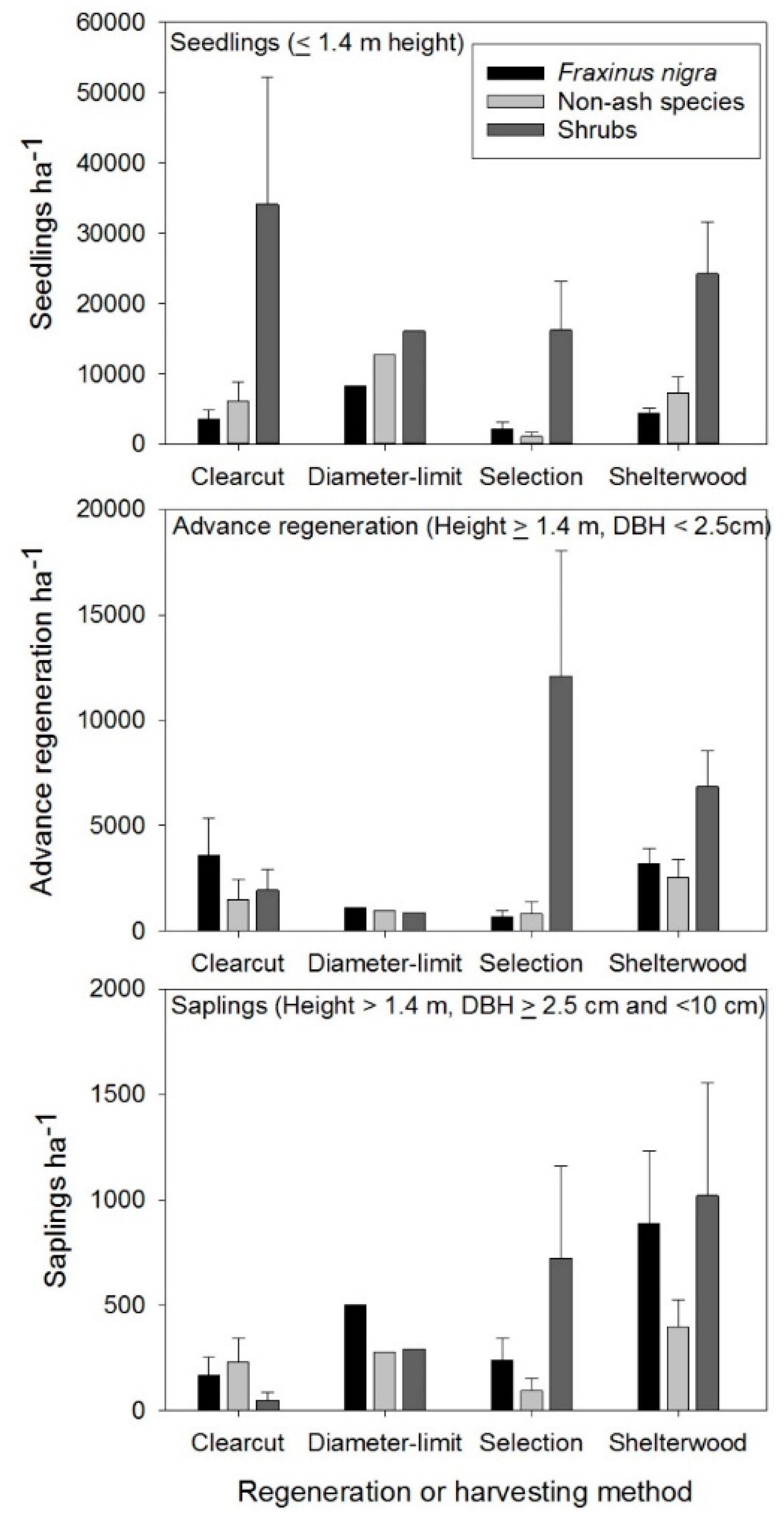
The silviculture methods implemented in the Swamp Hardwood Silviculture Trials included clearcut, coppice with standards, diameter limit, strip shelterwood, and single-tree selection with the majority of trials established in FnAbArOn habitat types given the greater abundance of this community type relative others in the state [53]. Within this habitat type, strip shelterwood harvests consisting of 9 m harvested strips alternating with 18 m unharvested strips generally resulted in the highest proportion of non-ash regeneration, particularly when non-ash seed trees were present in unharvested strips (Figure 6). Non-ash species commonly regenerating following these harvests included red maple, yellow birch, and American elm; however, black ash remained the most abundant species in the regeneration layer in this and all other regeneration methods (Figure 6). In addition, all treatments served to increase the abundance of shrub species in these forests indicating follow-up release work may be necessary to maintain non-ash components in these stands (Figure 6). Single-tree selection was the least effective regeneration method at stimulating non-ash regeneration given the smaller canopy openings in this approach largely served to release black ash advance regeneration and shrub species, such as speckled alder. Beyond positive regeneration responses, there was little evidence of elevated water tables in strip shelterwood harvests, whereas swamping was noted for clearcut and diameter-limit methods, although sample size was limited for the latter method. Strip shelterwoods had the lowest amount of harvesting machine traffic and disturbance across sites given equipment is restricted to harvested strips versus across the entire stand in other methods.
3.3. Decision Support Tools for Guiding Adaptive Management in Black Ash: The Wisconsin DNR Checklist for Evaluating Lowland Ash Stands
Despite the recent focus on managing black ash forests and other ash-dominated lowlands in the context of EAB, current silvicultural guidelines remain tentative and incomplete, due in large part to our limited knowledge and experience managing these ecosystems for species other than ash. Foresters and other natural resources managers have struggled with this knowledge gap while at the same time attempting to develop silvicultural prescriptions that maintain forest productivity and improve resilience to EAB. The WDNR Checklist for Evaluating Lowland Ash Stands was developed in an attempt to create a decision support tool that synthesizes our current state of knowledge, drawing upon lessons learned from the research studies and field trials described above [54]. This tool is complemented by other general guidance provided in the region by state agencies and University Extension (e.g., [55]).
The first component of this checklist is a stand assessment designed to track and evaluate site factors recognized as affecting silvicultural options and outcomes in black ash wetland forests based on research discussed above. These factors include an evaluation of site quality (e.g., Wetland Forest Habitat Type Classification System), timber sale operability, potential EAB impact to stand condition, regeneration potential, and hydrological risk. Potential EAB impact is a key determining factor in how managers may treat a given site and is based on the amount of non-ash acceptable growing stock (AGS) that would remain following the loss of black ash. Stands containing ≥40 non-ash AGS per acre or >45% relative density of non-ash AGS are considered less vulnerable to forest conversion following EAB and can be managed according to other forest cover type guidance. Similarly, stand regeneration potential is determined by evaluating non-ash advance regeneration, alternate seed sources, interfering vegetation, and browse pressure. Hydrologic function and the general risk of “swamping” (i.e., water table rise following timber harvesting) is evaluated using a set of observable site factors, such as seasonal inundation period, depth to water table, soil drainage class, depth to mineral soil, and the presence of impeded drainage.
Based on the stand assessment, users are guided to a decision tool to evaluate management options most appropriate for a specific set of stand conditions. Regeneration methods are recommended in vulnerable stands to naturally and artificially establish non-ash species. For example, partial harvest regeneration methods such as strip shelterwood and group selection are recommended for stands with high hydrological risk factors to limit the potential for swamping. The evaluation of stand regeneration potential is central to determining the need for artificial regeneration, as well as if site preparation and browse protection should be applied. It is important to emphasize that the decision tool is not a “cookbook” for every situation, but meant simply to serve as an aid in the prescription writing process when addressing the threat of EAB. To this end, the tool includes additional narratives that synthesizes lessons learned from the Lake States’ research studies and field trials, in addition to other research and field experience relevant to managing wetland forests.
3.4. Restoring Forest Cover to Areas Impacted by EAB
Much of the research evaluating adaptive silvicultural strategies for EAB has been developed for areas where EAB is not currently present or is not causing significant impact. Given the challenges with treating all areas prior to its arrival, new research in Wisconsin is now being established to assist foresters with restoring forest cover to black ash wetland sites post-EAB. As with other adaptive strategies, the primary goal is to ultimately maintain forested cover in these wetland systems. Work examining vegetation development following black ash mortality has demonstrated the rapidity with which shrubs, native sedges, and invasive species like reed canary grass (Phalaris arundinacea L.) increase on these wetland sites (e.g., [47]) creating a significant challenge to reforestation efforts. There is limited experience with site preparation techniques suitable for addressing these competition conditions on wetland sites and the WDNR has established a new study to better understand the cost and management efforts needed to establish new tree species following complete ash mortality. This study includes three replicates of clearcut harvests of black ash stands >25 hectares to purposely increase flooding duration and allow shrubs and sedges to increase. Study treatments will include brush mowing, herbicide application, and their combination on nested treatments of varying planted tree species that have shown promise as adaptive plantings in previous work (Table 1). In particular, evaluation of these treatments in an earlier silvicultural trial indicate this approach may be affective for establishing planted tamarack and black spruce on these sites [53]. Over the next ten years the WDNR hopes this experiment will generate needed recommendations to foresters on cost-suitable measures for site prep and tree seedling establishment following the loss of black ash to EAB.
4. Conclusions
The invasive EAB has motivated considerable interest and research in understanding the ecological functioning of black ash forests, as well as suitable adaptive silviculture strategies for maintaining these ecological functions even after the arrival of EAB. This work has reinforced the complexity of ecological conditions characterizing black ash-dominated wetlands and the unique, foundational role black ash plays in affecting hydrology, patterns in biodiversity, and vegetation dynamics. Given its unique role, there is truly no replacement for black ash in these forests. Despite this, recent experience with adaptive planting approaches in conjunction with regeneration harvests that maintain canopy cover during the establishment phase (i.e., group selection and shelterwood approaches) suggest there may be opportunities to increase non-ash components to maintain some of the forested wetland functions historically provided by black ash. Results from these studies are still quite short-term; however, they provide a framework for addressing the threat of an introduced pest by focusing on building resilience as opposed to solely focusing on eliminating host tree populations.
Despite the progress made over the past two decades in understanding black ash forests and in developing strategies for increasing their resilience to EAB, there are several key knowledge gaps that remain in relation to our ability to sustain these areas as forested wetlands into the future. One key knowledge gap is the long-term suitability of potential replacement species that are already being evaluated, and a related gap surrounds the suitability of other potential replacements species in these forests, as well as how differences in stock type might affect within species performance. Moreover, general availability of many of the species that have proven successful in early trials is quite limited and a better understanding of nursery capacity for providing planting material necessary for reforesting these areas is crucial. Similarly, there has been little evaluation of other reforestation techniques, particularly aerial seeding of replacement species, which may be more cost-effective and logistically feasible relative to hand planting black ash swamps. The impacts and effectiveness of site preparation treatments in establishing non-ash species and controlling invasive species common in these ecosystems, particularly reed canary grass, also have not been evaluated. Finally, the new field trials in Wisconsin looking at restoration treatments after EAB invasion and tree mortality will provide critical information on approaches for restoring tree cover; however, there is a need for like evaluations across the great variety of habitat types on which black ash currently dominates and associated feedbacks with local site hydrology. Regardless of treatment or site evaluated, all adaptive management efforts should include retention of mature, seed-bearing black ash to maintain its unique ecological functions prior to EAB arrival and provide opportunities for natural resistance and reestablishment after invasion [54].
Author Contributions
A.W.D., B.J.P., and R.A.S. conceived and designed the Minnesota experiments and conceptualized the paper; A.W.D., G.E., C.M., and D.R.B. summarized and integrated results from Wisconsin Lowland Hardwood Field Trials; and all authors contributed to writing the paper.
Acknowledgments
Special thanks to Laura Reuling, Kyle Gill, and Justin Pszwaro for leading field data collections and summaries from the Wisconsin DNR Swamp Hardwood Trials. Christopher Looney and Mitch Slater led field data collections and summaries for adaptive silviculture studies in black ash communities in north-central Minnesota. Nicholas Bolton provided insights on performance and survival of planted seedlings in Michigan and Wisconsin ash forests. Funding was provided by the Minnesota Environment and Natural Resources Trust Fund, the Upper Midwest and Great Lakes Landscape Conservation Cooperative, Department of Interior Northeast Climate Adaptation Science Center, USDA Forest Service Northern Research Station, USDA Forest Service Northeastern Forest Health Protection Program, MN Forest Resources Council, and Wisconsin DNR. Gary Swanson of the Chippewa National forest provided the initial inspiration for the Minnesota study. Comments from three anonymous reviewers and Dr. Marla Emery helped in improving an earlier version of this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ayres, M.P.; Lombardero, M.J. Forest pests and their management in the Anthropocene. Can. J. For. Res. 2017, 48, 292–301. [Google Scholar]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecol. Appl. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and Dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar]
- Waring, K.M.; O’Hara, K.L. Silvicultural strategies in forest ecosystems affected by introduced pests. For. Ecol. Manag. 2005, 209, 27–41. [Google Scholar] [CrossRef]
- Youngquist, M.B.; Eggert, S.L.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A. Potential effects of foundation species loss on Wetland Communities: A case study of black ash wetlands threatened by emerald ash borer. Wetlands 2017, 37, 787–799. [Google Scholar]
- Valachovic, Y.; Lee, C.; Marshall, J.; Scanlon, H. Wildland management of Phytophthora ramorum in northern California forests. In Proceedings of the Sudden Oak Death Science Symposium, Santa Rosa, CA, USA, 5–9 March 2007; Forest Service: Albany, CA, USA, 2008; pp. 305–312. [Google Scholar]
- Smith, M.T.; Turgeon, J.J.; de Groot, P.; Gasman, B. Asian longhorned beetle Anoplophora glabripennis (Motschulsky): Lessons learned and opportunities to improve the process of eradication and management. Am. Entomol. 2009, 55, 21–25. [Google Scholar] [CrossRef]
- McCullough, D.G.; Siegert, N.W. Estimating potential emerald ash borer (Coleoptera: Buprestidae) populations using ash inventory data. J. Econ. Entomol. 2007, 100, 1577–1586. [Google Scholar] [PubMed]
- McCullough, D.G.; Siegert, N.W.; Bedford, J. Slowing ash mortality: A potential strategy to slam emerald ash borer in outlier sites. In Proceedings of the 20th U.S. Department of Agriculture Interagency Research Forum on Invasive Species, Annapolis, MD, USA, 13–16 January 2009; McManus, K.A., Gottschalk, K.W., Eds.; General Technical Report NRS-P-51. U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2009; pp. 44–46. [Google Scholar]
- Kizlinski, M.L.; Orwig, D.A.; Cobb, R.C.; Foster, D.R. Direct and indirect ecosystem consequences of an invasive pest on forests dominated by eastern hemlock. J. Biogeogr. 2002, 29, 1489–1503. [Google Scholar] [CrossRef]
- MacFarlane, D.W.; Meyer, S.P. Characteristics and distribution of potential ash tree hosts for emerald ash borer. For. Ecol. Manag. 2005, 213, 15–24. [Google Scholar]
- Knight, K.S.; Brown, J.P.; Long, R.P. Factors affecting the survival of ash (Fraxinus spp.) trees infested by emerald ash borer (Agrilus planipennis). Biol. Invasions 2013, 15, 371–383. [Google Scholar]
- Erdmann, G.G.; Crow, T.R.; Peterson, R.M., Jr.; Wilson, C.D. Managing Black Ash in the Lake States; General Technical Report, NC-115; USDA Forest Service: Washington, DC, USA, 1987; 12p.
- Slesak, R.A.; Lenhart, C.F.; Brooks, K.N.; D’Amato, A.W.; Palik, B.J. Water table response to harvesting and simulated emerald ash borer mortality in black ash wetlands in Minnesota, USA. Can. J. For. Res. 2014, 44, 961–968. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer Invasion of North America: History, Biology, Ecology, Impacts, and Management. Ann. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Kashian, D.M. Sprouting and seed production may promote persistence of Green ash in the presence of the emerald ash borer. Ecosphere 2016, 7, e01332. [Google Scholar] [CrossRef]
- Iverson, L.; Knight, K.S.; Prasad, A.; Herms, D.A.; Matthews, S.; Peters, M.; Smith, A.; Hartzler, D.M.; Long, R.; Almendinger, J. Potential Species Replacements for Black Ash (Fraxinus nigra) at the Confluence of Two Threats: Emerald Ash Borer and a Changing Climate. Ecosystems 2016, 19, 248–270. [Google Scholar] [CrossRef]
- USDA Forest Service Forest Inventory and Analysis (USDA FIA). Forest Inventory and Analysis Database; U.S. Department of Agriculture, Forest Service, Northern Research Station: St. Paul, MN, USA, 2018. Available online: http://apps.fs.fed.us/fiadb-downloads/datamart.html (accessed on 6 April 2018).
- Weber, M. Factors Affecting Natural Tree Reproduction in Black Ash Communities in Northern Minnesota; MS Plan B Paper; College of Forestry, University of Minnesota: St. Paul, MN, USA, 1985; 58p. [Google Scholar]
- Kurmis, V.; Kim, J.H. Black Ash Stand Composition and Structure in Carlton County, Minnesota; Staff Paper Series Number 69; University of Minnesota Agricultural Experiment Station: St. Paul, MN, USA, 1989; 25p. [Google Scholar]
- Sterrett, W.D. The Ashes: Their Characteristics and Management; USDA Bulletin Number 299; US Department of Agriculture: Washington, DC, USA, 1915; 88p.
- Eyre, F.H. Forest Cover Types of the United States and Canada; Society of American Foresters: Washington, DC, USA, 1980; 148p. [Google Scholar]
- Michigan Department of Natural Resources (MIDNR). Forest Management Guide for Lowland Hardwoods Cover Type; Department of Natural Resources, Forest Management Division: Lansing, MI, USA, 1993; 4p.
- Kotar, J.; Kovach, J.A.; Burger, T.L. A Guide to Forest Communities and Habitat Types of Northern Wisconsin; Department of Forest Ecology and Management, University of Wisconsin-Madison: Madison, WI, USA, 2002. [Google Scholar]
- Aaseng, N.E. Field Guide to the Native Plant Communities of Minnesota: The Laurentian Mixed Forest Province; Minnesota Department of Natural Resources: St. Paul, MN, USA, 2003.
- Tardif, J.; Bergeron, Y. Population dynamics of Fraxinus nigra in response to flood-level variations, in Northwestern Quebec. Ecol. Monogr. 1999, 69, 107–125. [Google Scholar] [CrossRef]
- Kotar, J.; Burger, T.L. Wetland Forest Habitat Type Classification System for Northern Wisconsin; PUB-FR-627; Wisconsin Department of Natural Resources: Madison, WI, USA, 2017; 251p.
- Wisconsin Department of Natural Resources (WDNR). Silviculture Handbook: Chapter 46-Swamp Hardwood Cover Type; State of Wisconsin, Department of Natural Resources: Madison, WI, USA, 2013.
- Looney, C.E.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Reinikainen, M.R. Examining the influences of tree-to-tree competition and climate on size-growth relationships in hydric, multi-aged Fraxinus nigra stands. For. Ecol. Manag. 2016, 375, 238–248. [Google Scholar] [CrossRef]
- Davis, J.C.; Shannon, J.P.; Bolton, N.W.; Kolka, R.K.; Pypker, T.G. Vegetation responses to simulated emerald ash borer infestation in Fraxinus nigra dominated wetlands of Upper Michigan, USA. Can. J. For. Res. 2016, 47, 319–330. [Google Scholar] [CrossRef]
- Bowen, A.K.M.; Stevens, M.H.H. Predicting the effects of emerald ash borer (Agrilus planipennis, Buprestidae) on hardwood swamp forest structure and composition in southern Michigan. J. Torrey Bot. Soc. 2018, 145, 41–54. [Google Scholar] [CrossRef]
- Wright, J.W.; Rauscher, H.M. Black ash. In Silvics of North America, Volume 2, Hardwoods; Burns, R.M., Honkala, B.G., Eds.; Agricultural Handbook 654; United States Department of Agriculture (USDA): Washington, DC, USA, 1990; pp. 344–347. [Google Scholar]
- Palik, B.J.; Batzer, D.P.; Kern, C. Upland forest linkages to seasonal wetlands: Litter flux, processing, and food quality. Ecosystems 2006, 9, 142–151. [Google Scholar] [CrossRef]
- Van Grinsven, M.J.; Shannon, J.P.; Davis, J.C.; Bolton, N.W.; Wagenbrenner, J.W.; Kolka, R.K.; Pypker, T.G. Source water contributions and hydrologic responses to simulated emerald ash borer infestations in depressional black ash wetlands. Ecohydrology 2017, 10, e1862. [Google Scholar] [CrossRef]
- Lenhart, C.; Brooks, K.; Davidson, M.; Slesak, R.; D’Amato, A. Hydrologic source characterization of black ash wetlands: Implications for EAB response. In Proceedings of the American Water Resources Association Summer Specialty Conference Riparian Ecosystems IV: Advancing Science, Economics and Policy, Denver, CO, USA, 27–29 June 2012. [Google Scholar]
- Telander, A.C.; Slesak, R.A.; D’Amato, A.W.; Palik, B.J.; Brooks, K.N.; Lenhart, C.F. Sap flow of black ash in wetland forests of northern Minnesota, USA: Hydrologic implications of tree mortality due to emerald ash borer. Agric. For. Meteorol. 2015, 206, 4–11. [Google Scholar] [CrossRef]
- Shannon, J.; van Grinsven, M.; Davis, J.; Bolton, N.; Noh, N.; Pypker, T.; Kolka, R. Water Level Controls on Sap Flux of Canopy Species in Black Ash Wetlands. Forests 2018, 9, 147. [Google Scholar] [CrossRef]
- Kolka, R.; D’Amato, A.W.; Wagenbrenner, J.; Slesak, R.; Pypker, T.; Youngquist, M.; Grinde, A.; Palik, B. Review of ecosystem level impacts of emerald ash borer on black ash wetlands: What does the future hold? Forests 2018, 9, 179. [Google Scholar] [CrossRef]
- Diamond, J.S.; McLaughlin, D.; Slesak, R.A.; D’Amato, A.W.; Palik, B.J. Forested vs. Herbaceous Wetlands: Can management mitigate ecohydrologic regime shifts from invasive EAB? J. Environ. Manag. 2018, in press. [Google Scholar]
- Croxton, R.J. Detection and Classification of Ash Dieback on Large-Scale Color Aerial Photographs; Research Paper PSW-RP-35; USDA Forest Service: Washington, DC, USA, 1966.
- Livingston, W.H.; Hager, A.; White, A.S.; Hobbins, D. Drought associated with brown ash dieback in Maine. Phytopathology 1995, 85, 1554–1561. [Google Scholar]
- Palik, B.J.; Ostry, M.E.; Venette, R.C.; Abdela, E. Fraxinus nigra (black ash) dieback in Minnesota: Regional variation and potential contributing factors. For. Ecol. Manag. 2011, 261, 128–135. [Google Scholar] [CrossRef]
- Livingston, W.H.; White, A.S. May drought confirmed as likely cause of brown ash dieback in Maine. Phytopathology 1997, 87, S59. [Google Scholar]
- Trial, H., Jr.; Devine, M.E. Forest Health Monitoring Evaluation: Brown Ash (Fraxinus nigra) in Maine. A Survey of Occurrence and Health; Technical Report No. 33; Insect and Disease Management Division: August, ME, USA, 1994. [Google Scholar]
- Peterson, C.E. Natural Regeneration after Logging of Black Ash Stands in Central Minnesota. Master’s Thesis, College of Natural Resources, University of Minnesota, St. Paul, MN, USA, 1989; 58p. [Google Scholar]
- Kashian, D.M.; Witter, J.A. Assessing the potential for ash canopy tree replacement via current regeneration following emerald ash borer-caused mortality on southeastern Michigan landscapes. For. Ecol. Manag. 2011, 261, 480–488. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A.; Slater, M.A. The response of Fraxinus nigra forest ground-layer vegetation to emulated emerald ash borer mortality and management strategies in Northern Minnesota, USA. For. Ecol. Manag. 2017, 389, 352–363. [Google Scholar] [CrossRef]
- Palik, B.J.; Ostry, M.E.; Venette, R.C.; Abdela, E. Tree regeneration in black ash (Fraxinus nigra) stands exhibiting crown dieback in Minnesota. For. Ecol. Manag. 2012, 269, 26–30. [Google Scholar] [CrossRef]
- Looney, C.E.; D’Amato, A.W.; Palik, B.J.; Slesak, R.A. Overstory treatment and planting season affect survival of replacement tree species in emerald ash borer threatened Fraxinus nigra forests in Minnesota, USA. Can. J. For. Res. 2015, 45, 1728–1738. [Google Scholar] [CrossRef]
- Bolton, N.; Shannon, J.; Davis, J.; Grinsven, M.; Noh, N.; Schooler, S.; Kolka, R.; Pypker, T.; Wagenbrenner, J. Methods to improve survival and growth of planted alternative species seedlings in black ash ecosystems threatened by emerald ash borer. Forests 2018, 9, 146. [Google Scholar] [CrossRef]
- Kabrick, J.M.; Dey, D.C.; van Sambeek, J.W.; Coggeshall, M.V.; Jacobs, D.F. Quantifying flooding effects on hardwood seedling survival and growth for bottomland restoration. New For. 2012, 43, 695–710. [Google Scholar] [CrossRef]
- Knight, K.S.; Slavicek, J.M.; Kappler, R.; Pisarczyk, E.; Wiggin, B.; Menard, K. Using Dutch elm disease-tolerant elm to restore floodplains impacted by emerald ash borer. In Proceedings of the 4th International Workshop on Genetics of Host–Parasite Interactions in Forestry: Disease and Insect Resistance in Forest Trees, Eugene, OR, USA, 31 July–5 August 2011; General Technical Report PSW-GTR-240. USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2012; pp. 317–323. [Google Scholar]
- WDNR. Silviculture Trials Database: Swamp Hardwood; State of Wisconsin, Department of Natural Resources: Madison, WI, USA, 2015. Available online: https://dnr.wi.gov/topic/forestmanagement/silviculturetrials.html (accessed on 18 April 2018).
- WDNR. Checklist for Evaluating Lowland Ash Stands; State of Wisconsin, Department of Natural Resources: Madison, WI, USA, 2017; 8p.
- University of Minnesota (UMN) Extension. Ash Management Guidelines for Private Forest Landowners; University of Minnesota: St. Paul, MN, USA, 2011; p. 76. [Google Scholar]
Figure 1.
Age structure for two old-growth black ash forests in Minnesota. Sample size corresponds to number of cored individuals for which recruitment ages were obtainable, including several downed, dead black ash. The Kupcho Run site is classified as a “Very Wet Ash Forest (WFn64)”, whereas the Lake Williams site is classified as a “Wet Ash Forest (WFn55)” based on Aaseng et al. [25]. Other hardwood species are largely Ulmus americana L., Tilia americana L., Betula alleghaniensis Britt., Acer rubrum L., and Quercus macrocarpa Michx. Conifer species are Abies balsamea (L.) Mill., Picea glauca (Moench) Voss, and Thuja occidentalis L.
Figure 1.
Age structure for two old-growth black ash forests in Minnesota. Sample size corresponds to number of cored individuals for which recruitment ages were obtainable, including several downed, dead black ash. The Kupcho Run site is classified as a “Very Wet Ash Forest (WFn64)”, whereas the Lake Williams site is classified as a “Wet Ash Forest (WFn55)” based on Aaseng et al. [25]. Other hardwood species are largely Ulmus americana L., Tilia americana L., Betula alleghaniensis Britt., Acer rubrum L., and Quercus macrocarpa Michx. Conifer species are Abies balsamea (L.) Mill., Picea glauca (Moench) Voss, and Thuja occidentalis L.

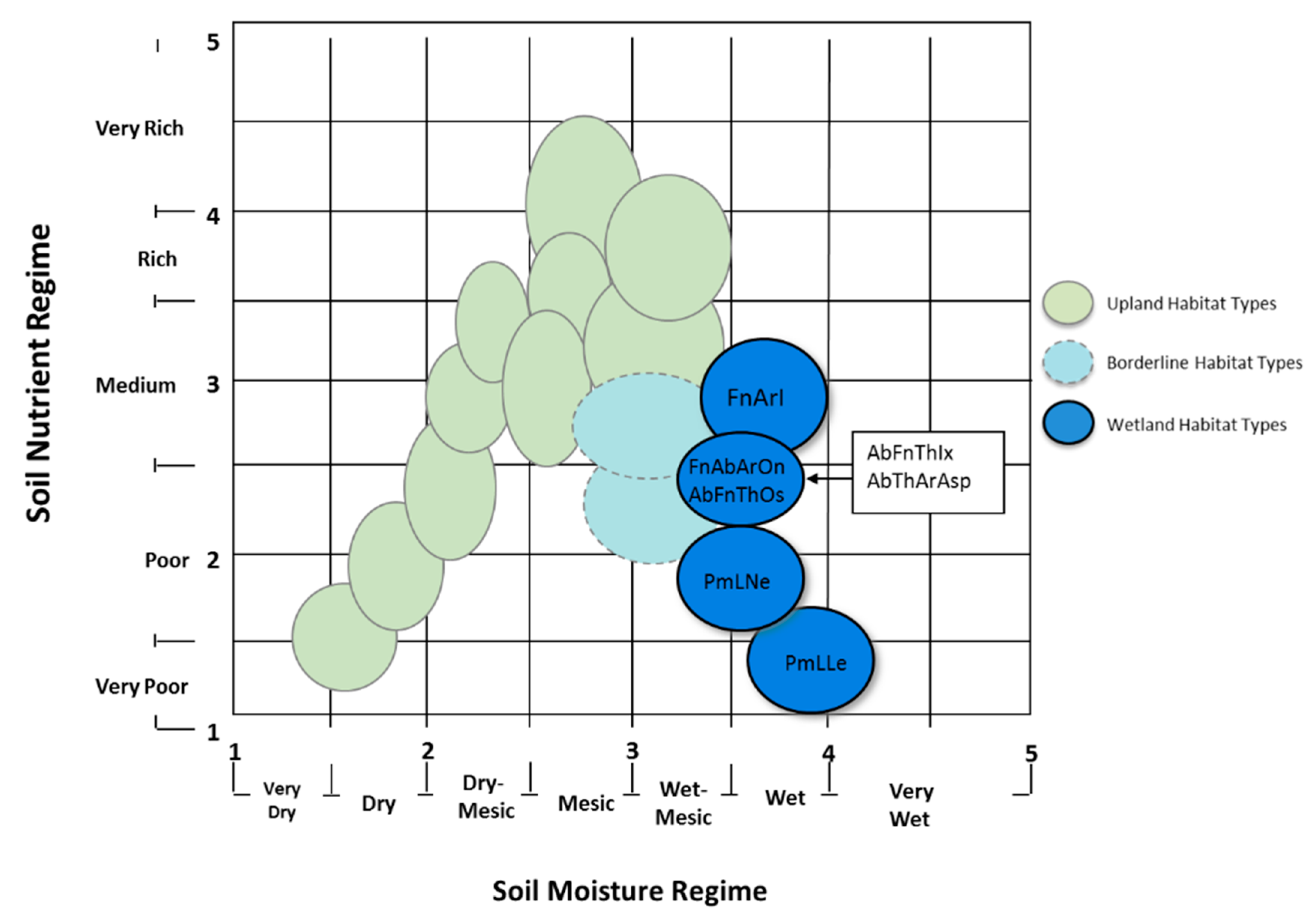
Figure 2.
Synecological coordinates of forest habitat types in north-central Wisconsin (Region 3) based on understory plant species communities, soil moisture, and nutrient regimes. Habitat types in blue are recently added wetland habitat types, including several that are dominated by black ash: Fraxinus-Acer/Impatiens (FnArl), Fraxinus-Abies-Acer/Onoclea (FnAbArOn), Abies-Fraxinus-Thuja/Ilex (AbFnThIx), and Abies-Fraxius-Thuja/Osmunda (AbFnThOs). Black ash-dominated habitat types in general are most mesic in nutrient regimes relative to other forested wetland communities in the region. From Kotar and Burger [27].
Figure 2.
Synecological coordinates of forest habitat types in north-central Wisconsin (Region 3) based on understory plant species communities, soil moisture, and nutrient regimes. Habitat types in blue are recently added wetland habitat types, including several that are dominated by black ash: Fraxinus-Acer/Impatiens (FnArl), Fraxinus-Abies-Acer/Onoclea (FnAbArOn), Abies-Fraxinus-Thuja/Ilex (AbFnThIx), and Abies-Fraxius-Thuja/Osmunda (AbFnThOs). Black ash-dominated habitat types in general are most mesic in nutrient regimes relative to other forested wetland communities in the region. From Kotar and Burger [27].

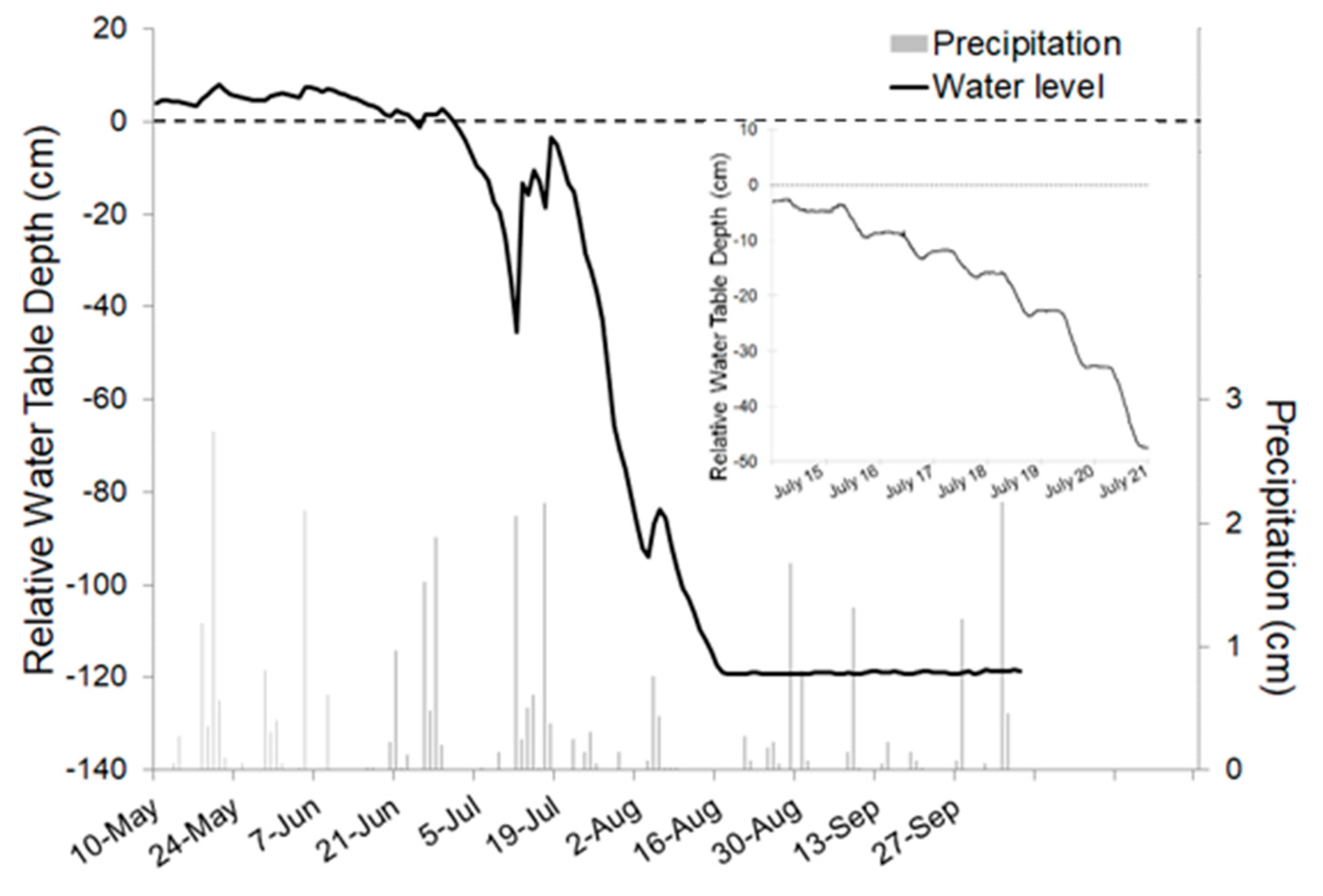
Figure 3.
Water table hydrograph for a black ash wetland in northern Minnesota (classified as WFn64 based on Aaseng et al. [25]). Water tables are above the soil surface following spring snowmelt, and then rapidly drawdown once black ash leaf expansion occurs (usually in mid-June). The influence of black ash transpiration on this pattern is readily apparent in the diurnal water table pattern once water tables are below the soil surface (inset).
Figure 3.
Water table hydrograph for a black ash wetland in northern Minnesota (classified as WFn64 based on Aaseng et al. [25]). Water tables are above the soil surface following spring snowmelt, and then rapidly drawdown once black ash leaf expansion occurs (usually in mid-June). The influence of black ash transpiration on this pattern is readily apparent in the diurnal water table pattern once water tables are below the soil surface (inset).

Figure 4.
(a) Swamp white oak (Quercus bicolor) planted in riparian black ash forest two years prior to photo in southern Wisconsin as part of a strategy to underplant non-ash species to increase ecosystem resilience to emerald ash borer (EAB); (b) Balsam poplar (Populus balsamifera) planted five years prior to photo in 0.04 ha group selection harvest as part of an adaptive approach for addressing the threat of EAB to the extensive black ash swamps in north-central Minnesota. Arrows in photos identify terminal shoot of planted individuals, which was at 0.8 and 1.6 m height in (a) and (b), respectively.
Figure 4.
(a) Swamp white oak (Quercus bicolor) planted in riparian black ash forest two years prior to photo in southern Wisconsin as part of a strategy to underplant non-ash species to increase ecosystem resilience to emerald ash borer (EAB); (b) Balsam poplar (Populus balsamifera) planted five years prior to photo in 0.04 ha group selection harvest as part of an adaptive approach for addressing the threat of EAB to the extensive black ash swamps in north-central Minnesota. Arrows in photos identify terminal shoot of planted individuals, which was at 0.8 and 1.6 m height in (a) and (b), respectively.

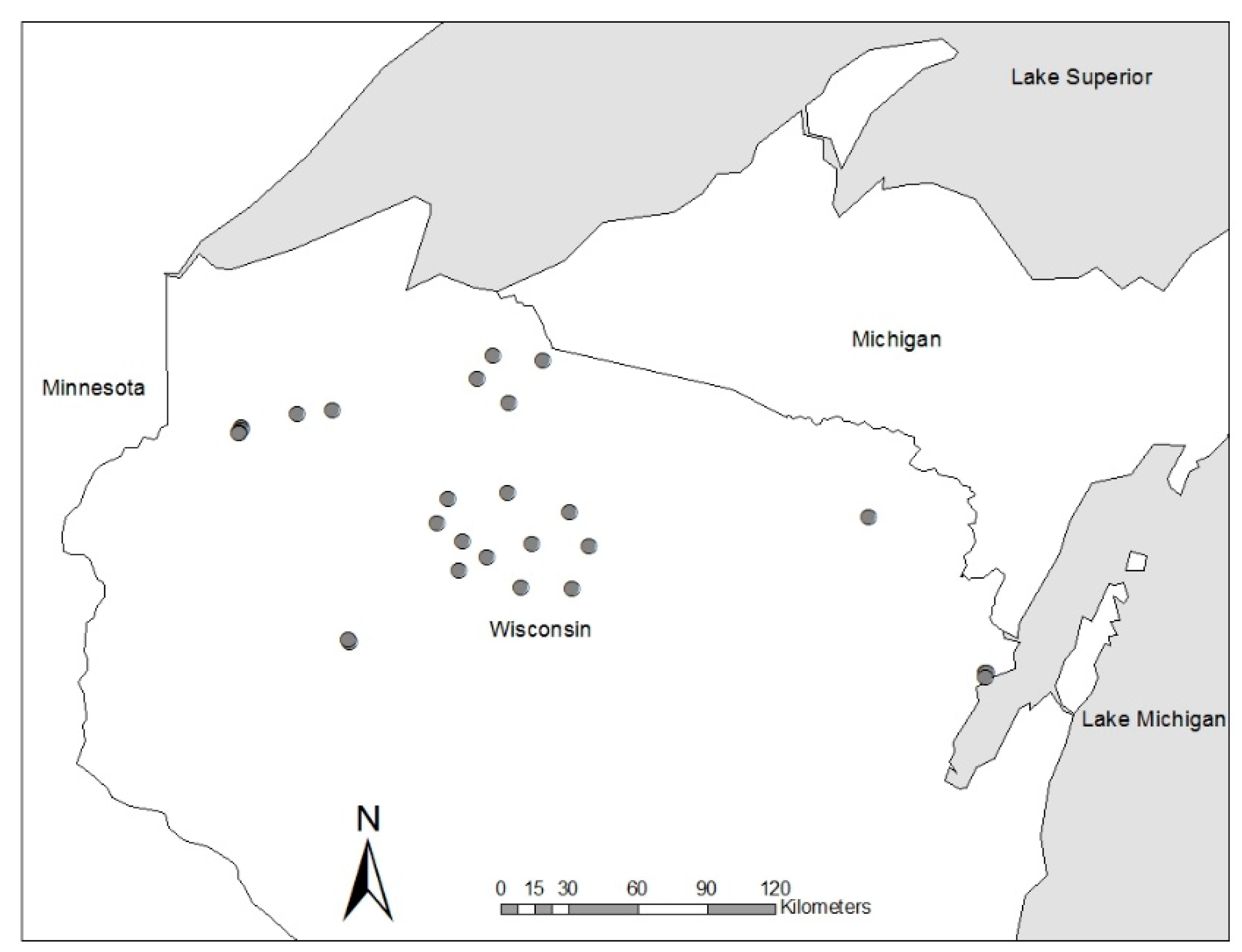
Figure 5.
Location of Swamp Hardwood Silviculture Trials maintained by the Wisconsin Department of Natural Resources. Trials are concentrated in north-central Wisconsin, which contains the greatest concentration of black ash wetlands in the state.
Figure 5.
Location of Swamp Hardwood Silviculture Trials maintained by the Wisconsin Department of Natural Resources. Trials are concentrated in north-central Wisconsin, which contains the greatest concentration of black ash wetlands in the state.

Figure 6.
Natural regeneration response to different regeneration methods and diameter-limit cutting as part of the Wisconsin Department of Natural Resources silviculture trials in black ash lowland forests in north-central Wisconsin. Data in figures are based on field measurements of these trials in 2015 and values represent means ± 1 standard error. Sites were harvested between 2003 and 2013 and all exist on Black Ash–Balsam Fir–Red Maple/Sensitive Fern (FnAbArOn) habitat types. Sample sizes for the different regeneration methods and harvesting approaches are: strip shelterwood (n = 6), clearcut (n = 4), diameter limit (n = 1), and single-tree selection (n = 4). Non-ash species include Acer rubrum, Betula alleghaniensis, Abies balsamea, Ulmus americana, Betula papyrifera Marsh., and Thuja occidentalis. Shrub species are predominantly Alnus incana, Salix spp., and Corylus cornuta.
Figure 6.
Natural regeneration response to different regeneration methods and diameter-limit cutting as part of the Wisconsin Department of Natural Resources silviculture trials in black ash lowland forests in north-central Wisconsin. Data in figures are based on field measurements of these trials in 2015 and values represent means ± 1 standard error. Sites were harvested between 2003 and 2013 and all exist on Black Ash–Balsam Fir–Red Maple/Sensitive Fern (FnAbArOn) habitat types. Sample sizes for the different regeneration methods and harvesting approaches are: strip shelterwood (n = 6), clearcut (n = 4), diameter limit (n = 1), and single-tree selection (n = 4). Non-ash species include Acer rubrum, Betula alleghaniensis, Abies balsamea, Ulmus americana, Betula papyrifera Marsh., and Thuja occidentalis. Shrub species are predominantly Alnus incana, Salix spp., and Corylus cornuta.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Species planted as part of adaptive strategies for minimizing the impacts on emerald ash borer on black ash wetlands. General recommendation for species (R = recommended, NR = not recommended) are based on 3–5-year patterns in survival and growth and are presented based on performance in research trials in Minnesota [47,49], Michigan, and Wisconsin [50], as well as operational plantings.
Table 1.
Species planted as part of adaptive strategies for minimizing the impacts on emerald ash borer on black ash wetlands. General recommendation for species (R = recommended, NR = not recommended) are based on 3–5-year patterns in survival and growth and are presented based on performance in research trials in Minnesota [47,49], Michigan, and Wisconsin [50], as well as operational plantings.
| Species | Stock Type | MN | MI | WI | Notes |
|---|---|---|---|---|---|
| Acer rubrum | Bare-root (2+0), containerized (90 cm3) | NR | NR | R | Success observed with bare-root stock in riparian black ash forest |
| Acer saccharinum | Bare-root (4+0) | - | R | - | Success observed with large bare-root stock |
| Abies balsamea | Bare-root (2+0, 3+0) | - | NR | NR | Very low survival regardless of cultural treatment |
| Betula alleghaniensis | Bare-root (2+0), Containerized (90 cm3) | NR | NR | NR | Very low survival with containerized stock. Marginal survival (36%) for bare-root stock planted in clearcut prepared by fecon mower |
| Celtis occidentalis | Bare-root (2+0), Containerized (336 cm3) | R | - | R | Greatest success when planted in group selection harvests in depressional wetlands |
| Fraxinus mandschurica | Bare-root (3+0) | NR | - | - | Limited cold tolerance, despite high survival of sprouts |
| Larix laricina | Bare-root (2+0), containerized (60 cm3) | NR | NR | R | Success observed with bare-root and containerized stock in clearcut prepared with fecon mower |
| Picea glauca | Bare-root (3+0) | - | - | R | Success observed with bare-root stock in clearcut prepared with fecon mower |
| Picea mariana | Containerized (90 cm3) | NR | NR | R | Success observed with bare-root and containerized stock in clearcut prepared with fecon mower |
| Pinus strobus | Bare-root (3+0) | - | - | R | Success observed with bare-root stock in clearcut prepared with fecon mower |
| Platanus occidentalis | Bare-root (1+0) | - | - | R | Success observed with bare-root stock in riparian black ash forest |
| Populus balsamifera | Containerized (164 cm3) | R | - | - | Greatest success when planted in clearcut and group selection harvests in depressional wetlands |
| Populus deltoides | Bare-root (1+0) | NR | - | - | Very low survival regardless of cultural treatment |
| Populus tremuloides | Containerized (90 cm3) | NR | - | NR | Marginal success observed with containerized stock in clearcut prepared with fecon mower |
| Quercus bicolor | Bare-root (1+0) | R | - | R | Greatest success in group selection harvests in depressional wetlands; successful under intact canopy of riparian black ash forests |
| Quercus macrocarpa | Bare-root (3+0) | - | NR | - | Greatest success when planted on hummocks |
| Thuja occidentalis | Bare-root (2+0), Containerized (60 cm3) | R | R | R | Greatest success in intact canopies and group selection harvests; high incidence of browse on seedlings |
| Tilia americana | Bare-root (3+0) | - | R | - | Greatest success when planted on hummocks |
| Tsuga canadensis | Bare-root (3+0) | - | - | NR | |
| Ulmus americana | Containerized (1890 cm3) | R | R | - | Greatest success when planted in group selection harvests and on hummocks using DED1 tolerant cultivars |
1 Dutch elm disease (Ophiostoma novo-ulmi).
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
D’Amato, A.W.; Palik, B.J.; Slesak, R.A.; Edge, G.; Matula, C.; Bronson, D.R. Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion. Forests 2018, 9, 348. https://doi.org/10.3390/f9060348
AMA Style
D’Amato AW, Palik BJ, Slesak RA, Edge G, Matula C, Bronson DR. Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion. Forests. 2018; 9(6):348. https://doi.org/10.3390/f9060348
Chicago/Turabian StyleD’Amato, Anthony W., Brian J. Palik, Robert A. Slesak, Greg Edge, Colleen Matula, and Dustin R. Bronson. 2018. "Evaluating Adaptive Management Options for Black Ash Forests in the Face of Emerald Ash Borer Invasion" Forests 9, no. 6: 348. https://doi.org/10.3390/f9060348
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.