Application of Trichoderma spp. Complex and Biofumigation to Control Damping-Off of Pinus radiata D. Don Caused by Fusarium circinatum Nirenberg and O’Donnell



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fusarium and Trichoderma Isolates
2.2. Microscopic Identification of F. circinatum
2.3. Biocontrol of F. circinatum by Trichoderma spp.
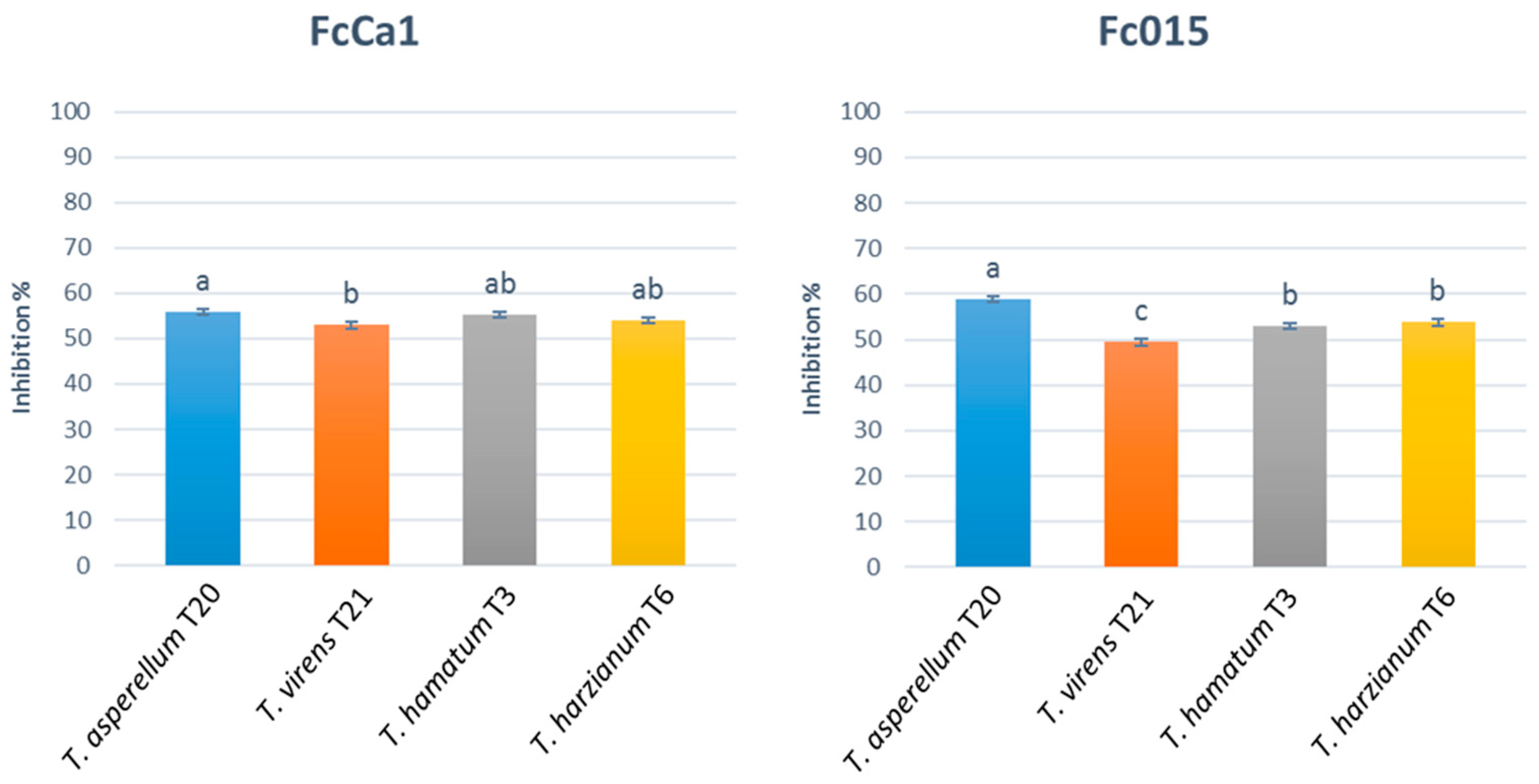
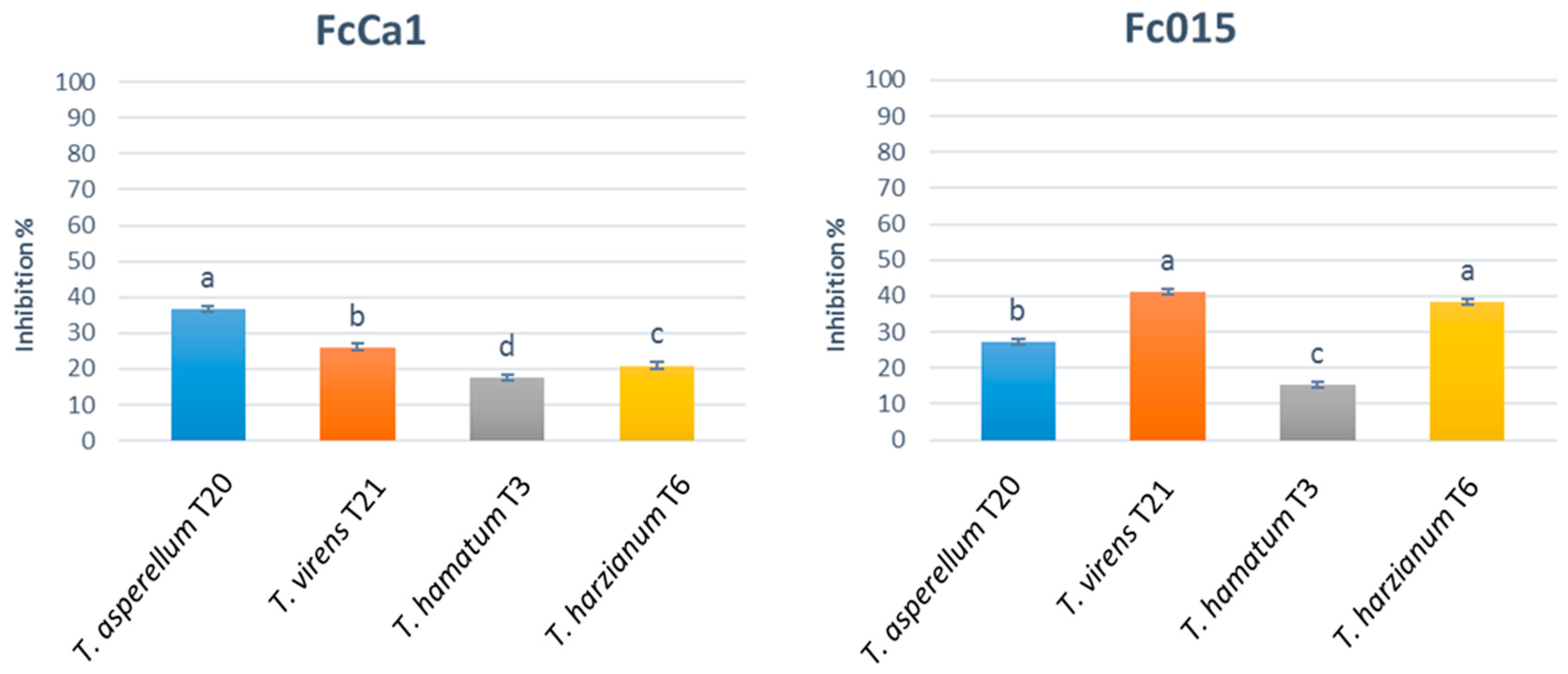
2.3.1. In Vitro Assessment of Antagonistic Activity by Dual Culture Assay and Antibiosis by Non-Volatile and Volatile Metabolites
2.3.2. Biocontrol Activity against F. circinatum in the Seedbed of P. radiata
2.4. Biocontrol of F. circinatum with Biofence®
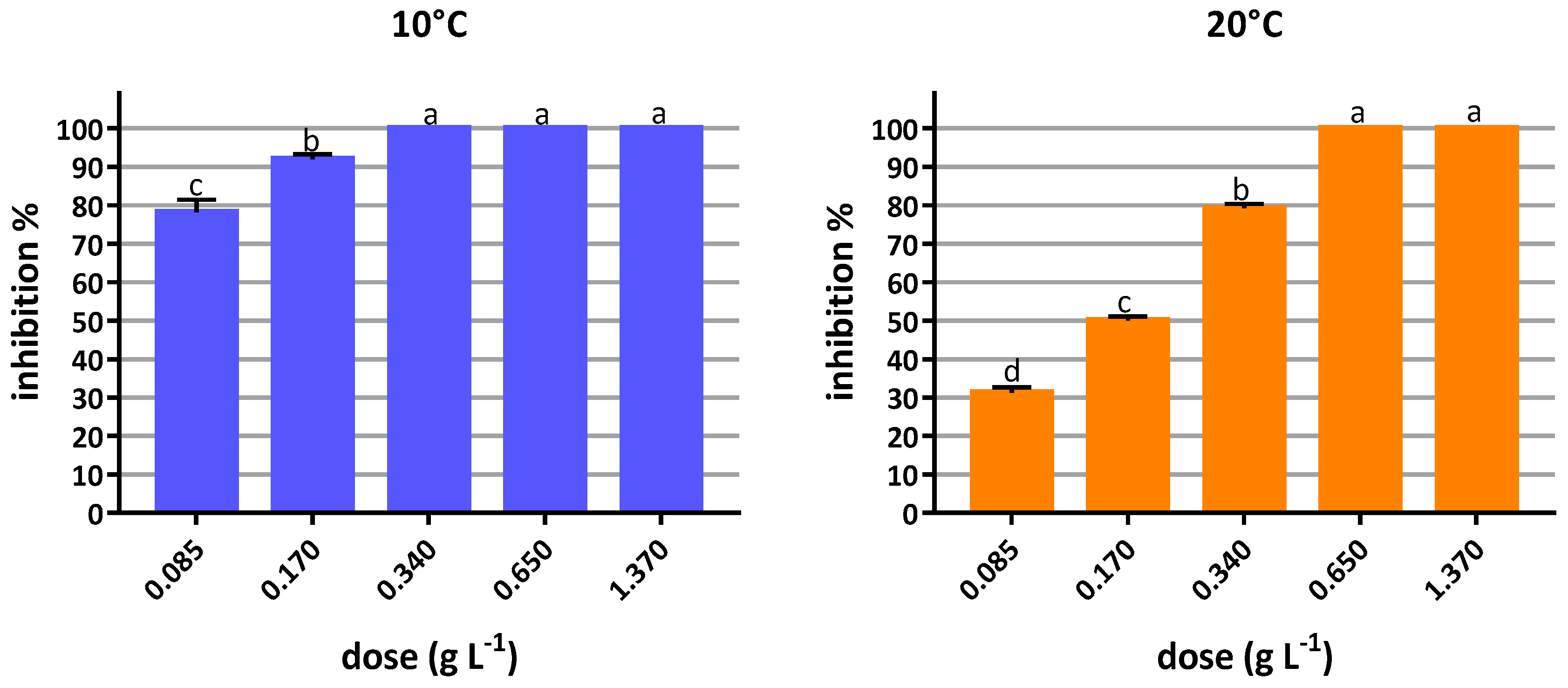
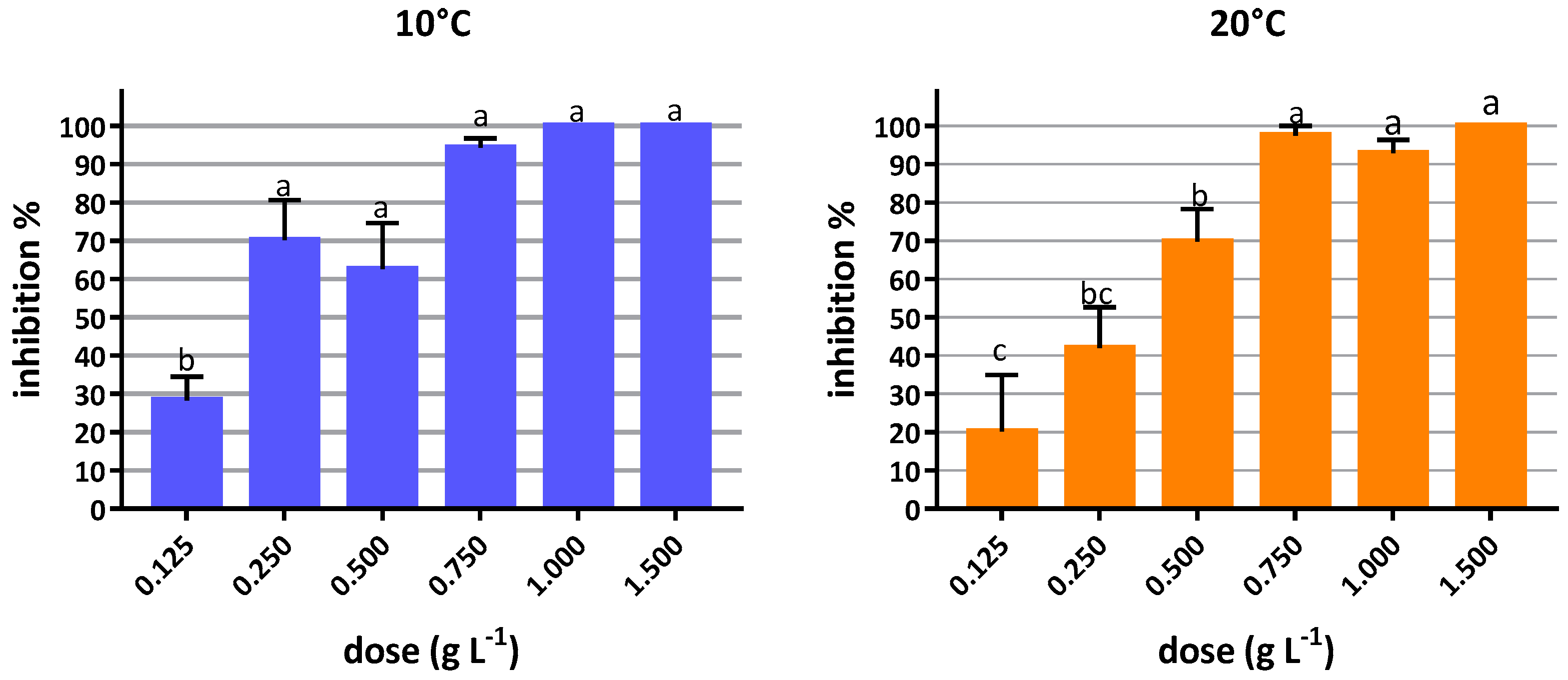
2.4.1. In Vitro Inhibition Tests
Mycelial Inhibition
Conidia Germination
2.4.2. Effect of Biofumigation Exposure Period on F. circinatum In Vitro
2.4.3. Effect of Biofumigation with Biofence® to Control F. circinatum on P. radiata Seeds
2.5. Statistical Analyses
3. Results
3.1. Biocontrol of F. circinatum by Trichoderma spp.
3.1.1. In Vitro Assessment of Antagonistic Activity by Dual Culture Assay and Antibiosis by Non-Volatile and Volatile Metabolites
3.1.2. Biocontrol Activity of F. circinatum in the Seedbed of Pinus radiata
3.2. Biocontrol of F. circinatum with BioFence®
3.2.1. In Vitro Inhibition Tests
3.2.2. Fungicide Effect and Application Timing of Biofence® on Fusarium circinatum In Vitro
3.2.3. Biocontrol in Pinus radiata Seeds with BioFence®
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ganley, R.J.; Watt, M.S.; Manning, L.; Iturritxa, E. A global climatic risk assessment of pitch canker disease. Can. J. For. Res. 2009, 39, 2246–2256. [Google Scholar] [CrossRef]
- Gordon, T.R.; Swett, C.L.; Wingfield, M.J. Management of Fusarium diseases affecting conifers. Crop Prot. 2015, 73, 28–39. [Google Scholar] [CrossRef]
- Evira-Recuenco, M.; Iturritxa, E.; Raposo, R. Impact of seed transmission on the infection and development of pitch canker disease in Pinus radiata. Forests 2015, 6, 3353–3368. [Google Scholar] [CrossRef]
- Dwinell, L.D.; Barrows-Broaddus, J.B.; Kuhlman, E.G. Pitch canker: A disease complex. Plant Dis. 1985, 69, 270–276. [Google Scholar] [CrossRef]
- Viljoen, A.; Wingfield, M.; Marasas, W. First report of Fusarium subglutinans f. sp. pini on pine seedlings in South Africa. Plant Dis. 1994, 78, 309–312. [Google Scholar] [CrossRef]
- Storer, A.; Gordon, T.; Clark, S. Association of the pitch canker fungus, Fusarium subglutinans f. sp. pini with Monterey pine seeds and seedlings in California. Plant Pathol. 1998, 47, 649–656. [Google Scholar] [CrossRef]
- Huang, J.; Kuhlman, E. Fungi associated with damping-off of slash pine seedlings in Georgia. Plant Dis. 1990, 74, 27–30. [Google Scholar] [CrossRef]
- Wingfield, M.; Hammerbacher, A.; Ganley, R.; Steenkamp, E.; Gordon, T.; Wingfield, B.; Coutinho, T. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Berbegal, M.; Landeras, E.; Sánchez, D.; Abad-Campos, P.; Pérez-Sierra, A.; Armengol, J. Evaluation of Pinus radiata seed treatments to control Fusarium circinatum: Effects on seed emergence and disease incidence. For. Pathol. 2015, 45, 525–533. [Google Scholar] [CrossRef]
- Garbelotto, M. Molecular analyses to study invasions by forest pathogens: Examples from Mediterranean ecosystems. Phytopathol. Mediterr. 2009, 47, 183–203. [Google Scholar]
- Berbegal, M.; Pérez-Sierra, A.; Armengol, J.; Grünwald, N. Evidence for multiple introductions and clonality in Spanish populations of Fusarium circinatum. Phytopathology 2013, 103, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Carey, W.; Oak, S.; Enebak, S. Pitch canker ratings of longleaf pine clones correlate with Fusarium circinatum infestation of seeds and seedling mortality in containers. For. Pathol. 2005, 35, 205–212. [Google Scholar] [CrossRef]
- Du Fretay, G.; Fritsch, F. Alterbromide, dissemination of sustainable alternatives to methyl bromide in soil disinfestation and in post-harvest. Acta Hortic. 2009, 883, 113–116. [Google Scholar] [CrossRef]
- Cooney, J.; Lauren, D. Trichoderma/pathogen interactions: Measurement of antagonistic chemicals produced at the antagonist/pathogen interface using a tubular bioassay. Lett. Appl. Microbiol. 1998, 27, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Khan, S.M.; Mohiddin, F.A. Biological control of Fusarium wilt of chickpea through seed treatment with the commercial formulation of Trichoderma harzianum and/or Pseudomonas fluorescens. Phytopathol. Mediterr. 2004, 43, 20–25. [Google Scholar]
- Howell, C. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef]
- Sivasithamparam, K.; Ghisalberti, E. Secondary metabolism in Trichoderma. In Trichoderma and Gliocladium. Volume 1: Basic Biology, Taxonomy and Genetics; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Aleandri, M.P.; Chilosi, G.; Bruni, N.; Tomassini, A.; Vettraino, A.M.; Vannini, A. Use of nursery potting mixes amended with local Trichoderma strains with multiple complementary mechanisms to control soil-borne diseases. Crop Prot. 2015, 67, 269–278. [Google Scholar] [CrossRef]
- Martín-Pinto, P.; Pajares, J.; Nanos, N.; Diez, J. Site and seasonal influences on the fungal community on leaves and stems of Pinus and Quercus seedlings in forest nurseries. Sydowia 2004, 56, 243–257. [Google Scholar]
- Hohmann, P.; Jones, E.E.; Hill, R.A.; Stewart, A. Understanding Trichoderma in the root system of Pinus radiata: Associations between rhizosphere colonization and growth promotion for commercially grown seedlings. Fungal. Biol. 2011, 115, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Moraga-Suazo, P.; Opazo, A.; Zaldúa, S.; González, G.; Sanfuentes, E. Evaluation of Trichoderma spp. and Clonostachys spp. strains to control Fusarium circinatum in Pinus radiata seedlings. Chil. J. Agric. Res. 2011, 71, 412–417. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Alves-Santos, F.M.; Diez, J.J. In vitro and in vivo interactions between Trichoderma viride and Fusarium circinatum. Silva Fenn. 2012, 46, 303–316. [Google Scholar] [CrossRef]
- Martín-García, J.; Paraschiv, M.; Flores-Pacheco, J.A.; Chira, D.; Diez, J.J.; Fernández, M. Susceptibility of several northeastern conifers to Fusarium circinatum and strategies for biocontrol. Forests 2017, 8, 318. [Google Scholar] [CrossRef]
- Sarwar, M.; Kirkegaard, J. Biofumigation potential of brassicas: II. Effect of environment and ontogeny on glucosinolate production and implications for screening. Plant Soil 1998, 201, 91–101. [Google Scholar] [CrossRef]
- Brown, P.D.; Morra, M.J. Control of soil-borne plant pests using glucosinolate-containing plants. Adv. Agron. 1997, 61, 168–231. [Google Scholar]
- Kirkegaard, J.A.; Sarwar, M.; Matthiessen, J.N. Assessing the biofumigation potential of crucifers. Acta Hortic. 1998, 459, 105–112. [Google Scholar] [CrossRef]
- Mehra, L.; MacLean, D.; Shewfelt, R.; Smith, K.; Scherm, H. Effect of postharvest biofumigation on fungal decay, sensory quality, and antioxidant levels of blueberry fruit. Postharvest Biol. Technol. 2013, 85, 109–115. [Google Scholar] [CrossRef]
- Ugolini, L.; Martini, C.; Lazzeri, L.; D’Avino, L.; Mari, M. Control of postharvest grey mould (Botrytis cinerea Pers.: Fr.) on strawberries by glucosinolate-derived allyl-isothiocyanate treatments. Postharvest Biol. Technol. 2014, 90, 34–39. [Google Scholar] [CrossRef]
- Smolinska, U.; Morra, M.; Knudsen, G.; James, R. Isothiocyanates produced by Brassicaceae species as inhibitors of Fusarium oxysporum. Plant Dis. 2003, 87, 407–412. [Google Scholar] [CrossRef]
- Nirenberg, H.I.; O’Donnell, K. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 1998, 90, 434–458. [Google Scholar] [CrossRef]
- Morton, D.; Stroube, W. Antagonistic and stimulatory effects of soil microorganisms upon Sclerotium rolfsii. Phytopathology 1955, 45, 417–420. [Google Scholar]
- Lahlali, R.; Hijri, M. Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 2010, 311, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma: II. Production of volatile antibiotics. Trans. Br. Mycol. Soc. 1971, 57, 41–44. [Google Scholar] [CrossRef]
- Ioos, R.; Fourrier, C.; Iancu, G.; Gordon, T.R. Sensitive detection of Fusarium circinatum in pine seed by combining an enrichment procedure with a real-time polymerase chain reaction using dual-labeled probe chemistry. Phytopathology 2009, 99, 582–590. [Google Scholar] [CrossRef] [PubMed]
- EPPO. Pm 7/91(1): Gibberella circinata. EPPO Bull. 2009, 39, 298–309. [Google Scholar]
- Lazzeri, L.; Curto, G.; Leoni, O.; Dallavalle, E. Effects of glucosinolates and their enzymatic hydrolysis products via myrosinase on the root-knot nematode Meloidogyne incognita (Kofoid et White) chitw. J. Agric. Food Chem. 2004, 52, 6703–6707. [Google Scholar] [CrossRef] [PubMed]
- Morales-Rodríguez, C.; Vettraino, A.; Vannini, A. Efficacy of biofumigation with Brassica carinata commercial pellets (BioFence®) to control vegetative and reproductive structures of Phytophthora cinnamomi. Plant Dis. 2016, 100, 324–330. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Alves-Santos, F.M.; Díez, J.J.J. Efecto de Trichoderma harzianum sobre el damping off causado por Fusarium circinatum en plántulas de Pinus radiata. In Proceedings of the XV Congreso de la Sociedad Española de Fitopatología, Vitoria-Gasteiz, Spain, 27 September–1 October 2010. [Google Scholar]
- López-López, N.; Segarra, G.; Vergara, O.; López-Fabal, A.; Trillas, M.I. Compost from forest cleaning green waste and Trichoderma asperellum strain t34 reduced incidence of Fusarium circinatum in Pinus radiata seedlings. Biol. Control 2016, 95, 31–39. [Google Scholar] [CrossRef]
- Roberts, D.P.; Lohrke, S.M.; Meyer, S.L.; Buyer, J.S.; Bowers, J.H.; Baker, C.J.; Li, W.; de Souza, J.T.; Lewis, J.A.; Chung, S. Biocontrol agents applied individually and in combination for suppression of soilborne diseases of cucumber. Crop Prot. 2005, 24, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Gilardi, G.; Moizio, M.; Gullino, M.L.; Garibaldi, A. Use of different soil amendments to control fusarium wilt of lettuce. Prot. Delle Colt. 2013, 2, 74. [Google Scholar]
- Sarwar, M.; Kirkegaard, J.; Wong, P.; Desmarchelier, J. Biofumigation potential of brassicas. 3. In vitro toxicity of isothiocyanates to soil-borne fungal pathogens. Plant Soil (Netherlands) 1998, 201, 103–112. [Google Scholar] [CrossRef]
- Steffek, R.; Spornberger, A.; Stich, K.; Lazzeri, L. The use of brassicas to reduce inoculum potential of soil-borne pests in Austrian horticulture. In Proceedings of the International Symposium on Horticulture in Europe, Vienna, Austria, 17–20 Febuary 2008. [Google Scholar]
- Matthiessen, J.N.; Shackleton, M.A. Biofumigation: Environmental impacts on the biological activity of diverse pure and plant-derived isothiocyanates. Pest Manag. Sci. 2005, 61, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Price, A.J.; Charron, C.S.; Saxton, A.M.; Sams, C.E. Allyl isothiocyanate and carbon dioxide produced during degradation of Brassica juncea tissue in different soil conditions. HortScience 2005, 40, 1734–1739. [Google Scholar]
- Lim, L.T.; Tung, M.A. Vapor pressure of allyl isothiocyanate and its transport in pvdc/pvc copolymer packaging film. J. Food Sci. 1997, 62, 1061–1062. [Google Scholar] [CrossRef]
- Yulianti, T.; Sivasithamparam, K.; Turner, D. Response of different forms of propagules of Rhizoctonia solani AG2-1 (ZG5) exposed to the volatiles produced in soil amended with green manures. Ann. Appl. Biol. 2006, 148, 105–111. [Google Scholar] [CrossRef]
- Kurt, Ş.; Güneş, U.; Soylu, E.M. In vitro and in vivo antifungal activity of synthetic pure isothiocyanates against Sclerotinia sclerotiorum. Pest Manag. Sci. 2011, 67, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, J.; Mitchell, N. The involvement of flavor volatiles in the resistance to downy mildew of wild and cultivated forms of Brassica oleracea. New Phytol. 1976, 77, 391–398. [Google Scholar] [CrossRef]
- Mari, M.; Leoni, O.; Iori, R.; Cembali, T. Antifungal vapor-phase activity of allyl-isothiocyanate against Penicillium expansum on pears. Plant Pathol. 2002, 51, 231–236. [Google Scholar] [CrossRef]
- Mari, M.; Iori, R.; Leoni, O.; Marchi, A. Bioassays of glucosinolate-derived isothiocyanates against postharvest pear pathogens. Plant Pathol. 1996, 45, 753–760. [Google Scholar] [CrossRef]
- Gordon, T.; Storer, A.; Wood, D. The pitch canker epidemic in California. Plant Dis. 2001, 85, 1128–1139. [Google Scholar] [CrossRef]
- Muñoz López, C.; Cuervo Sánchez, E.; Ampudia Díaz, M.; Gastón González, A.; Peñuelas Rubira, J.; Iglesias Sauce, S.; Herrero Sierra, N. Control químico de Fusarium circinatum Niremberg & O’Donnell en semillas del género Pinus. In Proceedings of the 5° Congreso Forestal Español, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Baker, R. Thermotherapy of planting material. Phytopathology 1962, 52, 1244–1255. [Google Scholar]
- Barnett, J.P. Sterilizing southern pine seeds with peroxide. Tree Plant. Notes 1976, 27, 17–19. [Google Scholar]
- Vettraino, A.M.; Potting, R.P.J.; Raposo, R. EU legislation on forest plant health: An overview with a focus on Fusarium circinatum. Forests. in press.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Germination ± SE | % Seedling Mortality ± SE 1 | |
|---|---|---|
| Non-inoculated control | 78.3 ± 5.2 b | 0 |
| T-complex | 66.6 ± 5.2 b | 0 |
| Fusarium and T-complex | 70.83 ± 7.52 b | 16.5 ± 3.7 a |
| Fusarium | 52.5 ± 8.51 a | 69 ± 3.7 b |
| 10 °C | 20 °C | Models Comparison 2 | |||
|---|---|---|---|---|---|
| EC50 (g L−1 ± SE) | Goodness of Fit Rsq (DF 1) | EC50 (g L−1 ± SE) | Goodness of Fit Rsq (DF) | ||
| Mycelial growth | 0.026 ± 0.003 | 0.98 (69) | 0.22 ± 0.02 | 0.97 (69) | F = 399.4; p < 0.0001 |
| Conidia germination | 0.3 ± 0.1 | 0.76 (52) | 0.47 ± 0.15 | 0.79 (53) | F = 1.22; p > 0.05 |
| % Germination ± SE | |
|---|---|
| Non-inoculated control | 75 ± 6.4 a |
| Non-inoculated control and BioFence® | 51.7 ± 11.38 b |
| Fusarium and BioFence® | 47.8 ± 11.7 b |
| Fusarium | 22.8 ± 7.2 c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Rodríguez, C.; Bastianelli, G.; Aleandri, M.; Chilosi, G.; Vannini, A. Application of Trichoderma spp. Complex and Biofumigation to Control Damping-Off of Pinus radiata D. Don Caused by Fusarium circinatum Nirenberg and O’Donnell. Forests 2018, 9, 421. https://doi.org/10.3390/f9070421
Morales-Rodríguez C, Bastianelli G, Aleandri M, Chilosi G, Vannini A. Application of Trichoderma spp. Complex and Biofumigation to Control Damping-Off of Pinus radiata D. Don Caused by Fusarium circinatum Nirenberg and O’Donnell. Forests. 2018; 9(7):421. https://doi.org/10.3390/f9070421
Chicago/Turabian StyleMorales-Rodríguez, Carmen, Giorgia Bastianelli, MariaPia Aleandri, Gabriele Chilosi, and Andrea Vannini. 2018. "Application of Trichoderma spp. Complex and Biofumigation to Control Damping-Off of Pinus radiata D. Don Caused by Fusarium circinatum Nirenberg and O’Donnell" Forests 9, no. 7: 421. https://doi.org/10.3390/f9070421