Fire Season, Overstory Density and Groundcover Composition Affect Understory Hardwood Sprout Demography in Longleaf Pine Woodlands


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Statistical Analyses
2.3. Genet Survival
2.4. Sprouting
2.5. Stem Survival
3. Results
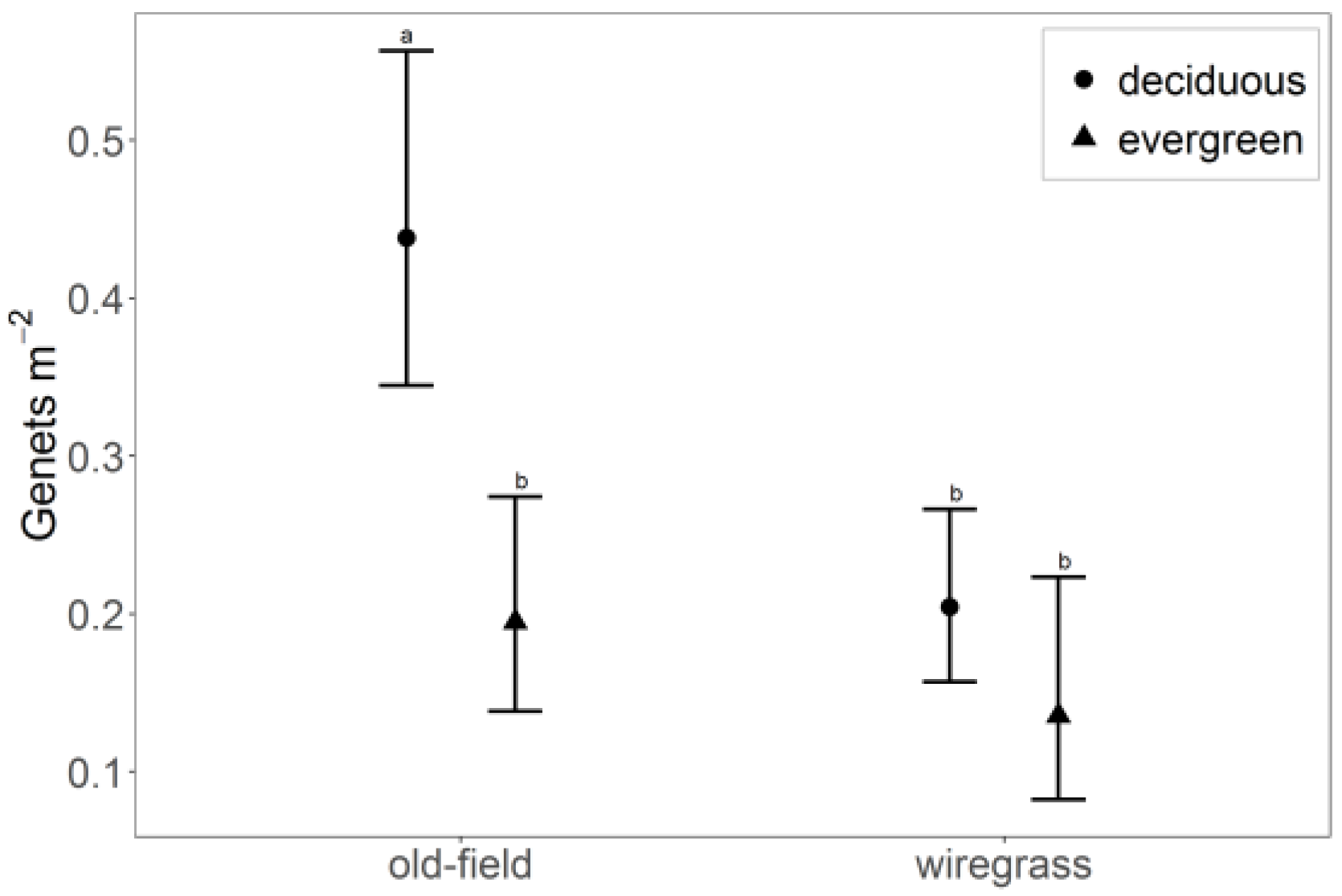
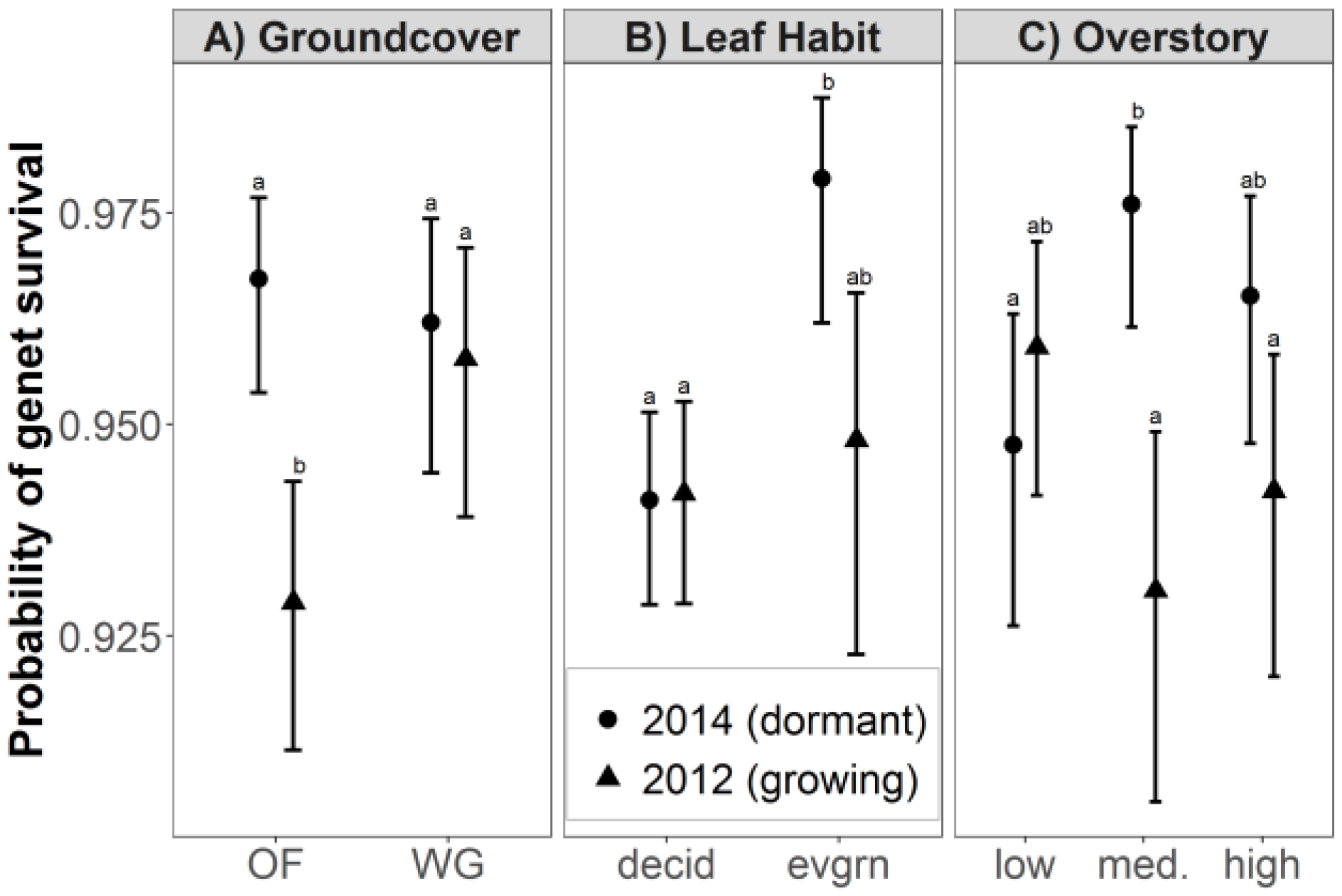
3.1. Genet Survival
3.2. New Sprout Counts
3.3. Stem Survival
3.4. Genet Height
4. Discussion
Management Implications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mitchard, E.T.A.; Flintrop, C.M. Woody encroachment and forest degradation in sub-Saharan Africa’s woodlands and savannas 1982–2006. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholes, R.J.; Archer, S.R. Tree-Grass Interactions in Savannas 1. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Van Dijk, A.I.J.M.; De Jeu, R.A.M.; Canadell, J.G.; McCabe, M.F.; Evans, J.P.; Wang, G. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Chang. 2015, 5, 470–474. [Google Scholar] [CrossRef]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Outcalt, K.W. Lightning, fire and longleaf pine: Using natural disturbance to guide management. For. Ecol. Manag. 2008, 255, 3351–3359. [Google Scholar] [CrossRef] [Green Version]
- Heyward, F. The relation of fire to stand composition of longleaf pine forests. Ecology 1939, 20, 287–304. [Google Scholar] [CrossRef]
- Christensen, N.L. Fire regimes in southeastern ecosystems. In Proceedings of the Conference Fire Regimes and Ecosystem Properties, Honolulu, HI, USA, 11–15 December 1978; Mooney, H.A., Bonnicksen, T.M., Christensen, N.L., Lotspeich, F.B., Reiners, W.A., Eds.; USDA Forest Service: Washington, DC, USA, 1981; pp. 112–136. [Google Scholar]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of fire regime and habitat on tree dynamics in north florida longleaf pine savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar] [CrossRef]
- Duncan, B.W.; Schmalzer, P.A. Anthropogenic influences on potential fire spread in a pyrogenic ecosystem of Florida, USA. Landsc. Ecol. 2004, 19, 153–165. [Google Scholar] [CrossRef]
- Kirkman, L.; Goebel, P.; Palik, B.; West, L. Predicting plant species diversity in a longleaf pine landscape. Ecoscience 2004, 11, 80–93. [Google Scholar] [CrossRef]
- Van Lear, D.H.; Carroll, W.D.; Kapeluck, P.R.; Johnson, R. History and restoration of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Drewa, P.B.; Platt, W.J.; Moser, A.E.B. Fire effects on resprouting of shrubs in headwaters of southeastern longleaf pine savannas. Ecology 2002, 83, 755–767. [Google Scholar] [CrossRef]
- Bond, W.J.; Van Wilgen, B.W. Fire and Plants, 1st ed.; Chapman & Hall: London, UK, 1996; ISBN 0412475405. [Google Scholar]
- Waldrop, T.A.; White, D.L.; Jones, S.M. Fire regimes for pine-grassland communities in the southeastern United States. For. Ecol. Manag. 1992, 47, 195–210. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Bond, W.J. What Limits Trees in C4 Grasslands and Savannas? Annu. Rev. Ecol. Evol. Syst. 2008, 39, 641–659. [Google Scholar] [CrossRef]
- Chao, W.S.; Foley, M.E.; Horvath, D.P.; Anderson, J. V Signals regulating dormancy in vegetative buds. Int. J. Plant Dev. Biol. 2007, 1, 49–56. [Google Scholar]
- Wade, D. Fuel moisture and prescribed burning. South. Fire Exch. Fact Sheet 2013, 5, 5–8. [Google Scholar]
- Boyer, W.D. Volume Growth Loss: A Hidden Cost of Periodic Prescribed Burning in Longleaf Pine? South. J. Appl. For. 1987, 11, 154–157. [Google Scholar]
- Streng, D.R.; Glitzenstein, J.S.; Platt, W.J. Evaluating effects of season of burn in longleaf pine forests: A critical literature review and some results from ongoing long-term study. In Proceedings of 18th Tall Timbers Fire Ecology Conference; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993; pp. 227–263. [Google Scholar]
- Robertson, K.M.; Hmielowski, T.L. Effects of fire frequency and season on resprouting of woody plants in southeastern US pine-grassland communities. Oecologia 2014, 174, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Melvin, M.; McIntyre, R.K. Air quality and human health challenges to prescribed fire. In Ecological Restoration and Management of Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 255–266. ISBN 9781498748186. [Google Scholar]
- Chiodi, A.M.; Larkin, N.S.; Varner, J.M. An analysis of Southeastern US prescribed burn weather windows: Seasonal variability and El Niño associations. Int. J. Wildl. Fire 2018, 27, 176–189. [Google Scholar] [CrossRef]
- Fill, J.M.; Moule, B.M.; Varner, J.M.; Mousseau, T.A. Flammability of the keystone savanna bunchgrass Aristida stricta. Plant Ecol. 2016, 217, 331–342. [Google Scholar] [CrossRef]
- Fonda, R.W. Burning Characteristics of Needles from Eight Pine Species. For. Sci. 2001, 47, 390–396. [Google Scholar]
- Wenk, E.S.; Wang, G.G.; Walker, J.L. Understory fuel variation at the Carolina Sandhills National Wildlife Refuge: A description of chemical and physical properties. In Proceedings of the 15th Biennial Southern Silvicultural Research Conference, Asheville, USA, 2013; Guldin, J.M., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2013; Volume 175, pp. 351–356. [Google Scholar]
- Loudermilk, E.L.; O’Brien, J.J.; Mitchell, R.J.; Cropper, W.P.; Hiers, J.K.; Grunwald, S.; Grego, J.; Fernandez-Diaz, J.C. Linking complex forest fuel structure and fire behaviour at fine scales. Int. J. Wildl. Fire 2012, 21, 882–893. [Google Scholar] [CrossRef]
- Parrott, R.T. A Study of Wiregrass (Aristida Stricta Michx.) with Particular Reference to Fire. Master’s Thesis, Duke University, Durham, NC, USA, 1967. [Google Scholar]
- Hendricks, J.J.; Wilson, C.A.; Boring, L.R. Foliar litter position and decomposition in a fire-maintained longleaf pine-wiregrass ecosystem. Can. J. For. Res. 2002, 32, 928–941. [Google Scholar] [CrossRef]
- Williamson, G.B.; Black, E.M. High temperature of forest fires under pines as a selective advantage over oaks. Nature 1981, 293, 643–644. [Google Scholar] [CrossRef]
- Hiers, J.K.; O’Brien, J.J.; Mitchell, R.J.; Grego, J.M.; Loudermilk, E.L. The wildland fuel cell concept: An approach to characterize fine-scale variation in fuels and fire in frequently burned longleaf pine forests. Int. J. Wildl. Fire 2009, 18, 315–325. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.J.; Jack, S.B.; Engstrom, R.T. Silviculture that sustains: The nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Can. J. For. Res. 2006, 36, 2724–2736. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.; Starr, G. Ecological forestry in the Southeast: Understanding the ecology of fuels. J. For. 107391-397 2009, 107, 391–397. [Google Scholar]
- Nyland, R.D. Silviculture: Concepts and Applications, 2nd ed.; McGraw-Hill: Boston, MA, USA, 2002; ISBN 9780073661902. [Google Scholar]
- Kirkman, L.K.; Giencke, L.M. Restoring and managing a diverse ground cover. In Ecological Restoration and Management of Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 207–232. [Google Scholar]
- Miller, D.L.; Mora, C.I.; Grissino-Mayer, H.D.; Mock, C.J.; Uhle, M.E.; Sharp, Z. Tree-ring isotope records of tropical cyclone activity. Proc. Natl. Acad. Sci. USA 2006, 103, 14294–14297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hmielowski, T.L.; Robertson, K.M.; Platt, W.J. Influence of season and method of topkill on resprouting characteristics and biomass of Quercus nigra saplings from a southeastern U.S. pine-grassland ecosystem. Plant Ecol. 2014, 215, 1221–1231. [Google Scholar] [CrossRef]
- Platt, W.J.; Ellair, D.P.; Huffman, J.M.; Potts, S.E.; Beckage, B. Pyrogenic fuels produced by savanna trees can engineer humid savannas. Ecol. Monogr. 2016, 86, 352–372. [Google Scholar] [CrossRef]
- Palik, B.J.B.J.; Pederson, N. Overstory mortality and canopy disturbances in longleaf pine ecosystems. Can. J. For. Res. 1996, 26, 2035–2047. [Google Scholar] [CrossRef]
- Pederson, N.; Varner, J.M.; Palik, B.J. Canopy disturbance and tree recruitment over two centuries in a managed longleaf pine landscape. For. Ecol. Manag. 2008, 254, 85–95. [Google Scholar] [CrossRef]
- NCEI Monthly Normals, Albany SW Georgia Regional Airport, GA. Available online: https://www.ncdc.noaa.gov/cdo-web/datasets 9 (accessed on 30 November 2016).
- McIntyre, R.K.; Jack, S.B.; Mitchell, R.J.; Kevin Hiers, J.; Neel, W.L. Multiple Value Management: The Stoddard-Neel Approach to Ecological Forestry in Longleaf Pine Grasslands, 1st ed.; Joseph, W., Ed.; Jones Ecological Research Center: Newton, MA, USA, 2008. [Google Scholar]
- Palik, B.; Mitchell, R.J.; Pecot, S.; Battaglia, M.; Pu, M. Spatial Distribution of Overstory Retention Influences Resources and Growth of Longleaf Pine Seedlings. Ecol. Appl. 2003, 13, 674–686. [Google Scholar] [CrossRef]
- Stoll, P.; Weiner, J.; Schmid, B. Growth variation in a naturally established population of Pinus sylvestris. Ecology 1994, 75, 660–670. [Google Scholar] [CrossRef]
- Berg, E.E.; Hamrick, J.L. Spatial and Genetic Structure of Two Sandhills Oaks: Quercus laevis and Quercus margaretta (Fagaceae). Am. J. Bot. 1994, 81, 7–14. [Google Scholar] [CrossRef]
- Cavender-bares, A.J.; Kitajima, K.; Bazzaz, F.A. Multiple Trait Associations in Relation to Habitat Differentiation among 17 Floridian Oak Species. Ecol. Monogr. 2004, 74, 635–662. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 1. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 15 May 2017).
- Christensen, R.H.B. Ordinal—Regression Models for Ordinal Data. Available online: https://cran.r-project.org/package=ordinal (accessed on 15 May 2017).
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2000; ISBN 0470582472. [Google Scholar]
- Gilliam, F.S.; Platt, W.J. Effects of Long-Term Fire Exclusion on Tree Species Composition and Stand Structure in an Reviewed work (s): Effects of long-term fire exclusion on tree species composition and stand structure in an old-growth Pinus palustris (Longleaf pine) forest. Plant Ecol. 1999, 140, 15–26. [Google Scholar] [CrossRef]
- Jack, S.B.; Mitchell, R.J.; Pecot, S.D. Silvicultural Alternatives in a Longleaf Pine/Wiregrass Woodland in Southwest Georgia: Understory Hardwood Response To Harvest-Created Gaps. In Proceedings of the 13th Southern Silvicultural Research Conference, Asheville, NC, USA, 2006; Connor, K.F., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2006; pp. 85–89. [Google Scholar]
- Stambaugh, M.C.; Varner, J.M.; Noss, R.F.; Dey, D.C.; Christensen, N.L.; Baldwin, R.F.; Guyette, R.P.; Hanberry, B.B.; Harper, C.A.; Lindblom, S.G.; et al. Clarifying the role of fire in the deciduous forests of eastern North America: Reply to Matlack. Conserv. Biol. 2015, 29, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Grady, J.M.; Hoffmann, W.A. Caught in a fire trap: Recurring fire creates stable size equilibria in woody resprouters. Ecology 2012, 93, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.D.; Arthur, M.A.; Loftis, D.L.; Green, S.R. Survival and growth of upland oak and co-occurring competitor seedlings following single and repeated prescribed fires. For. Ecol. Manag. 2008, 256, 1021–1030. [Google Scholar] [CrossRef]
- Menaut, J.C.; Gignoux, J.; Prado, C.; Clobert, J. Tree Community Dynamics in a Humid Savanna of the Cote-d’Ivoire: Modelling the Effects of Fire and Competition with Grass and Neighbours. J. Biogeogr. 1990, 17, 471–481. [Google Scholar] [CrossRef]
- Keyser, T.L.; Loftis, D.L. Stump sprouting of 19 upland hardwood species 1 year following initiation of a shelterwood with reserves silvicultural system in the southern Appalachian Mountains, USA. New For. 2015, 46, 449–464. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. [Technical Coordinators] Silvics of North America: Volume 2. Hardwoods; Agricultural Handbook; United States Department of Agriculture (USDA), Forest Service: Washington, DC, USA, 1990; Volume 654.
- Meier, A.R.; Saunders, M.R.; Michler, C.H. Epicormic buds in trees: A review of bud establishment, development and dormancy release. Tree Physiol. 2012, 32, 565–584. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.; Starr, G.; Mack, M.C.; Martin, T.A.; Gholz, H.L. Effects of a prescribed fire on understory vegetation, carbon pools, and soil nutrients in a longleaf pine-slash pine forest in Florida. Nat. Areas J. 2010, 30, 82–94. [Google Scholar] [CrossRef]
- Varner, J.M.; Kane, J.M.; Hiers, J.K.; Kreye, J.K.; Veldman, J.W. Suites of fire-adapted traits of oaks in the Southeastern USA: Multiple strategies for persistence. Fire Ecol. 2016, 12, 48–64. [Google Scholar] [CrossRef]
- Graves, S.J.; Rifai, S.W.; Putz, F.E. Outer bark thickness decreases more with height on stems of fire-resistant than fire-sensitive Floridian oaks (Quercus spp.; Fagaceae). Am. J. Bot. 2014, 101, 2183–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawes, M.J.; Adie, H.; Russell-Smith, J.; Murphy, B.; Midgley, J.J. How do small savanna trees avoid stem mortality by fire? The roles of stem diameter, height and bark thickness. Ecosphere 2011, 2, 1–13. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Mitchell, R.J.; Mou, P.P.; Pecot, S.D. Light Transmittance Estimates in a Longleaf Pine Woodland. For. Sci. 2003, 49, 752–762. [Google Scholar]
- Pecot, S.D.; Mitchell, R.J.; Palik, B.J.; Moser, E.B.; Hiers, J.K. Competitive responses of seedlings and understory plants in longleaf pine woodlands: Separating canopy influences above and below ground. Can. J. For. Res. 2007, 37, 634–648. [Google Scholar] [CrossRef]
- Wright, A.; Schnitzer, S.A.; Reich, P.B. Living close to your neighbors-the importance of both competition and facilitation in plant communities. Ecology 2014, 95, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Werner, P.A. Growth of juvenile and sapling trees differs with both fire season and understorey type: Trade-offs and transitions out of the fire trap in an Australian savanna. Austral Ecol. 2012, 37, 644–657. [Google Scholar] [CrossRef]
- Fill, J.M.; Glitzenstein, J.S.; Streng, D.R.; Stowe, J.; Mousseau, T.A. Wiregrass ( Aristida beyrichiana ) May Limit Woody Plant Encroachment in Longleaf Pine ( Pinus palustris ) Ecosystems. Am. Midl. Nat. 2017. [Google Scholar] [CrossRef]
- Markewitz, D.; Sartori, F.; Craft, C. Soil change and carbon storage in longleaf pine stands planted on marginal agricultural lands. Ecol. Appl. 2002, 12, 1276–1285. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathic effects of Andropogon virginicus and its persistence in old fields. Am. J. Bot. 1972, 59, 752–755. [Google Scholar] [CrossRef]
- Daneshgar, P.; Jose, S. Role of species identity in plant invasions: Experimental test using Imperata cylindrica. Biol. Invasions 2009, 11, 1431–1440. [Google Scholar] [CrossRef]
- Gagnon, P.R.; Passmore, H.A.; Platt, W.J.; Myers, J.A.; Paine, C.E.T.; Harms, K.E. Does pyrogenicity protect burning plants? Ecology 2010, 91, 3481–3486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peet, R.; Allard, D. Longleaf pine vegetation of the southern Atlantic and eastern Gulf Coast regions: A preliminary classification. In Proceedings of 18th Tall Timbers Fire Ecology Conference, Tallahassee, USA, 1993; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993; pp. 45–81. [Google Scholar]
- Brockway, D.G.; Outcalt, K.W. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- McGuire, J.P.; Mitchell, R.J.; Moser, E.B.; Pecot, S.D.; Gjerstad, D.H.; Hedman, C.W. Gaps in a gappy forest: Plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Can. J. For. Res. 2001, 31, 765–778. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Groundcover | Temp. (C) | RH (%) | WS (m/s) | FMC (%) |
|---|---|---|---|---|---|
| 2010 | old-field | 26.3 | 51.1 | 1.7 | 8.1 |
| wiregrass | 21.1 | 55.1 | 2.0 | 9.6 | |
| 2012 | old-field | 27.1 | 54.9 | 1.7 | 8.5 |
| wiregrass | 24.8 | 48.0 | 2.1 | 10.9 | |
| 2014 | old-field | 11.3 | 30.9 | 1.7 | 12.6 |
| wiregrass | 14.9 | 37.6 | 2.1 | 12.0 | |
| 2016 | old-field | 27.7 | 43.1 | 1.9 | 7.0 |
| wiregrass | 23.9 | 48.5 | 2.1 | 8.0 |
| Common Name | Latin Name | Leaf Habit † | Family |
|---|---|---|---|
| laurel oak | Quercus hemisphaerica Michx. | brevideciduous | Fagaceae |
| live oak | Quercus virginiana Mill. | evergreen | Fagaceae |
| water oak | Quercus nigra L. | brevideciduous | Fagaceae |
| bluejack oak | Quercus incana W. Bartram | deciduous | Fagaceae |
| post oak | Quercus stellata Wangenh. | deciduous | Fagaceae |
| southern red oak | Quercus falcata Michx. | deciduous | Fagaceae |
| common persimmon | Diospyros virginiana L. | deciduous | Ebenaceae |
| sassafras | Sassafras albidum (Nutt.) Nees | deciduous | Lauraceae |
| winged sumac | Rhus copallinum L. | deciduous | Anacardiaceae |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whelan, A.W.; Bigelow, S.W.; Nieminen, M.F.; Jack, S.B. Fire Season, Overstory Density and Groundcover Composition Affect Understory Hardwood Sprout Demography in Longleaf Pine Woodlands. Forests 2018, 9, 423. https://doi.org/10.3390/f9070423
Whelan AW, Bigelow SW, Nieminen MF, Jack SB. Fire Season, Overstory Density and Groundcover Composition Affect Understory Hardwood Sprout Demography in Longleaf Pine Woodlands. Forests. 2018; 9(7):423. https://doi.org/10.3390/f9070423
Chicago/Turabian StyleWhelan, Andrew W., Seth W. Bigelow, Mary Frances Nieminen, and Steven B. Jack. 2018. "Fire Season, Overstory Density and Groundcover Composition Affect Understory Hardwood Sprout Demography in Longleaf Pine Woodlands" Forests 9, no. 7: 423. https://doi.org/10.3390/f9070423