Satellite RNAs and Satellite Viruses of Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
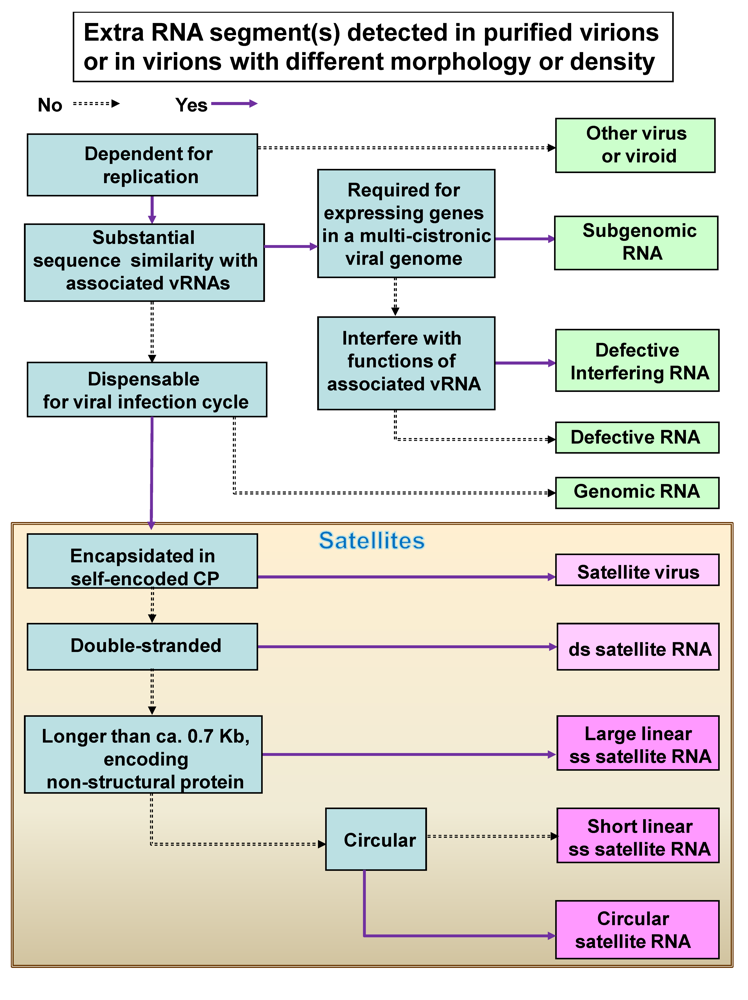
2. General classification of satRNAs and satellite viruses

2.1. Single-Stranded RNA Satellite Viruses
2.2. Single-stranded satRNAs
3. Newly identified satRNAs or satellite viruses
4. Recent advances regarding satellite viruses
5. Recent advances regarding satRNAs
5.1. Replication mechanisms
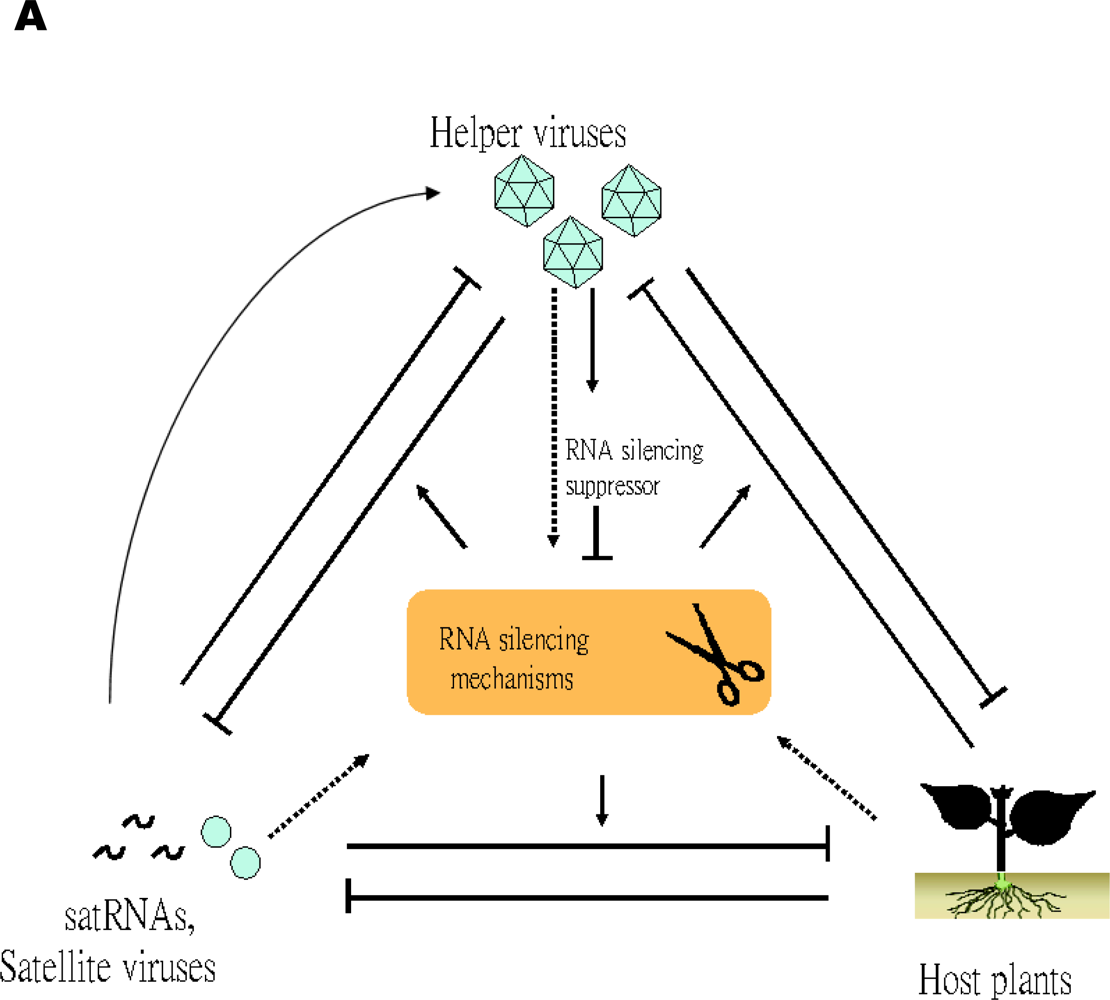
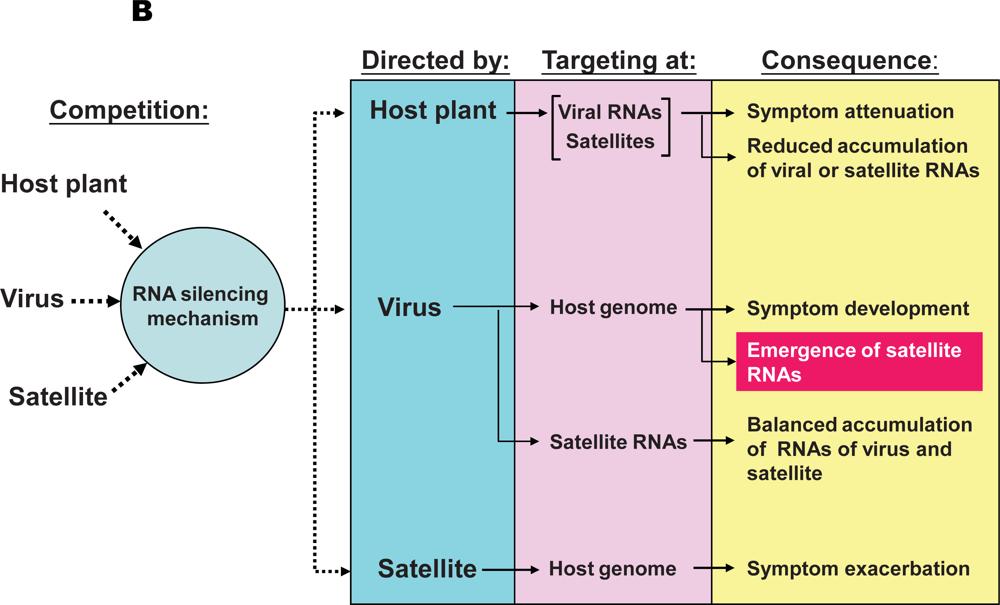
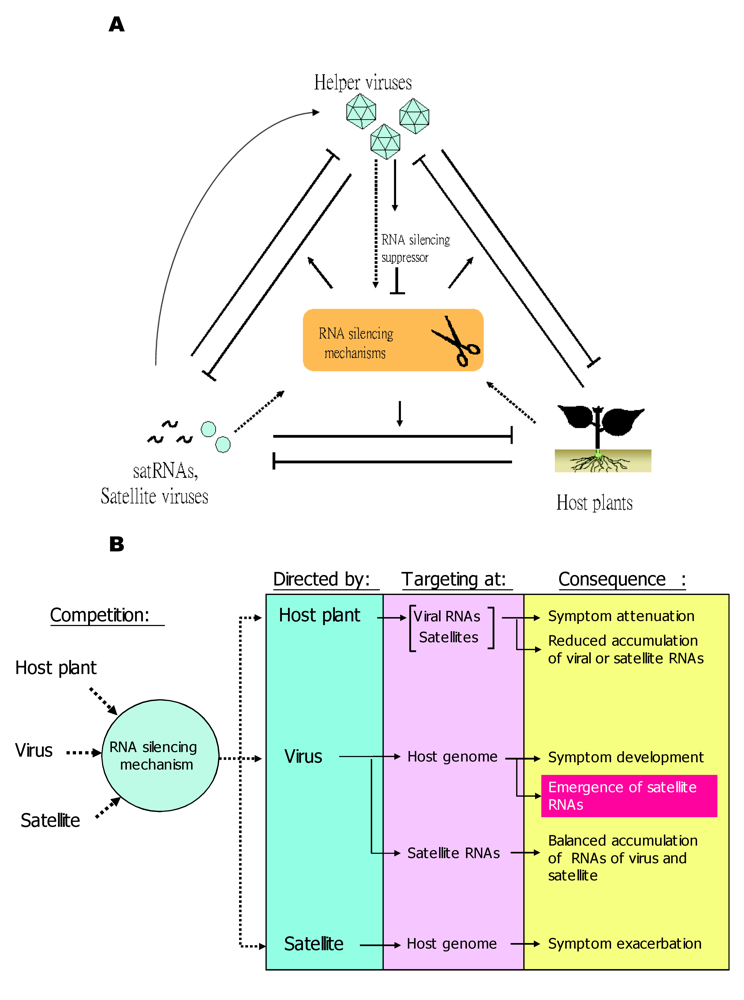
5.2. RNA silencing mechanisms of the host plants are involved in the pathogenicity of satRNAs and their co-evolution with their helper viruses
The pathogenicity of satRNAs is determined by a complex competition among host plants, helper viruses, and satRNAs in the targetting of the RNA silencing mechanism
A simplified model illustrating trilateral competition for the targeting of RNA silencing mechanisms
5.3. Possible Origin(s) of satRNAs

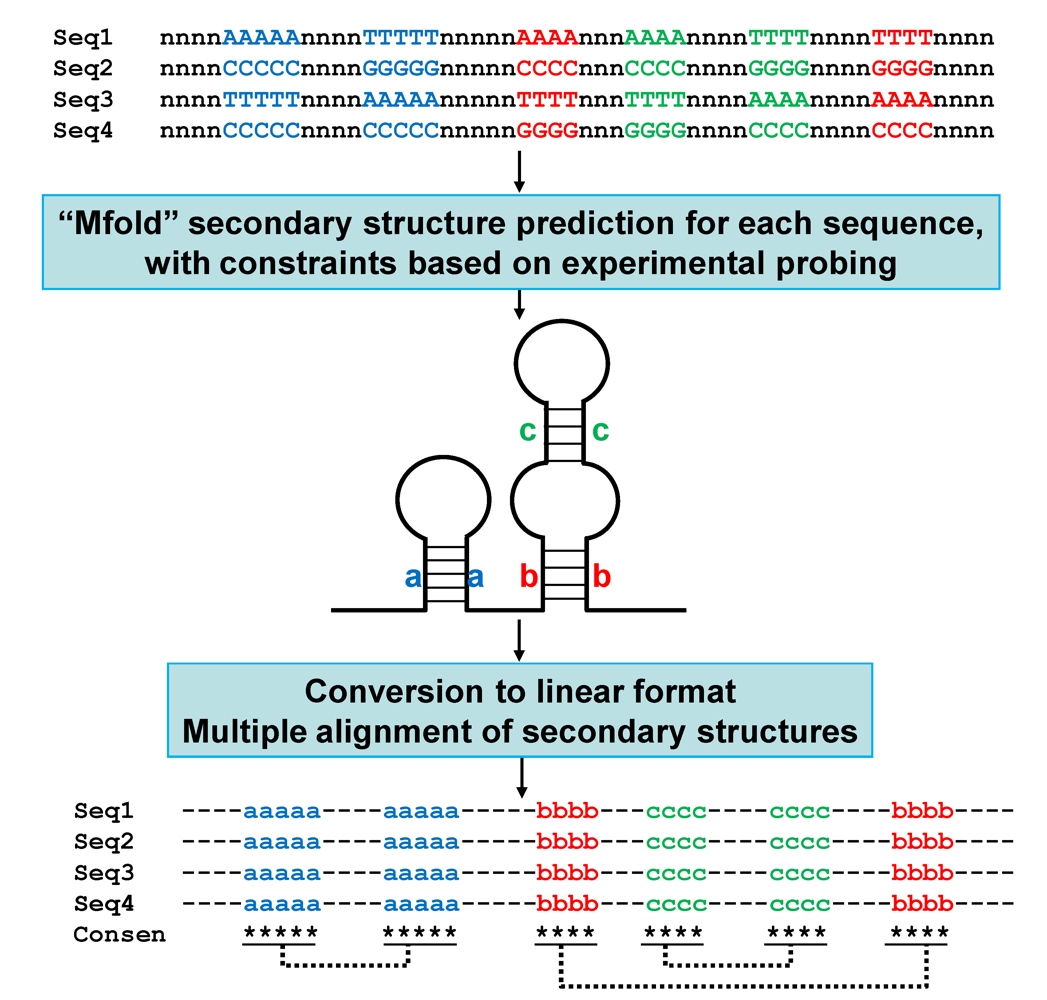
5.4. Molecular evolution: constraints on the structures of satRNAs; linear comparison of multiple secondary structures

6. Practical applications of satRNAs
7. Concluding Remarks
Acknowledgments
References
- Murant, A.F; Mayo, M.A. Satellites of plant viruses. Annu. Rev. Phytopathol. 1982, 20, 49–70. [Google Scholar] [CrossRef]
- Tien, P.; Wu, G.S. Satellite RNA for the biocontrol of plant disease. Adv. Virus Res. 1991, 39, 321–39. [Google Scholar] [PubMed]
- Roossinck, M.J.; Sleat, D.; Palukaitis, P. Satellite RNAs of plant viruses: structures and biological effects. Microbiol. Rev. 1992, 56, 265–79. [Google Scholar] [PubMed]
- Garcia-Arenal, F.; Palukaitis, P. Structure and functional relationships of satellite RNAs of cucumber mosaic virus. Curr. Top. Microbiol. Immunol. 1999, 239, 37–63. [Google Scholar] [PubMed]
- Simon, A.E.; Roossinck, M.J.; Havelda, Z. Plant virus satellite and defective interfering RNAs: new paradigms for a new century. Annu. Rev. Phytopathol. 2004, 42, 415–37. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.M.; Waterworth, H.E. Cucumber mosaic virus associated RNA 5: causal agent for tomato necrosis. Science 1977, 196, 429–431. [Google Scholar] [PubMed]
- Mossop, D.W; Francki, R.I.B. Comparative studies on two satellite RNAs of cucumber mosaic virus. Virology 1979, 95, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, H.E.; Kaper, J.M.; Tousignant, M.E. CARNA 5, the small cucumber mosaic virus-dependent replicating RNA causal agent of lethal tomato necrosis, regulates disease expression. Science 1979, 204, 845–847. [Google Scholar] [PubMed]
- Gossele, V.; Metzlaff, M. Using satellite tobacco mosaic virus vectors for gene silencing. Protoc. Microbiol. 2005, Chapter 16, Unit 16I.5. [Google Scholar]
- Lin, N.S.; Lee, Y.S.; Lin, B.Y.; Lee, C.W.; Hsu, Y.H. The open reading frame of bamboo mosaic potexvirus satellite RNA is not essential for its replication and can be replaced with a bacterial gene. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 3138–3142. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.B.; Bian, X.Y.; Wu, L.M.; Liu, L.X.; Smith, N.A.; Isenegger, D.; Wu, R.M.; Masuta, C.; Vance, V.B.; Watson, J.M.; Rezaian, A.; Dennis, E.S.; Waterhouse, P.M. On the role of RNA silencing in the pathogenicity and evolution of viroids and viral satellites. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 3275–3280. [Google Scholar] [CrossRef] [PubMed]
- Pantaleo, V.; Burgyan, J. Cymbidium ringspot virus harnesses RNA silencing to control the accumulation of virus parasite satellite RNA. J. Virol. 2008, 82, 11851–11858. [Google Scholar] [CrossRef] [PubMed]
- Satellites and Defective Viral RNAs, 2nd; Vogt, P.K.; Jackson, A.O.E. (Eds.) Springer-Verlag: Berlin, Germany, 1999; Volume 69, pp. 179–.
- Astier, S.; Albouy, J.; Maury, Y.; Robaglia, C.; Lecoq, H. Science Publishers: Enfield, NH, USA, 2006; pp. 147–156.
- Mayo, M.A.; Leibowitz, M.J; Palukaitis, P.; Scholthof, K.-B.G.; Simon, A.E.; Stanley, J.; Taliansky, M. Satellites. Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A.E., Eds.; Elsevier Academic Press: New York, NY, USA, 2005; pp. 1163–1169. [Google Scholar]
- Satellites, 3rdBuchen-Osmond, C. (Ed.) Columbia University: New York, NY, USA, 2006.
- Saunders, K.; Bedford, I.D.; Briddon, R.W.; Markham, P.G.; Wong, S.M.; Stanley, J. A unique virus complex causes Ageratum yellow vein disease. Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 6890–6895. [Google Scholar] [CrossRef] [PubMed]
- Dry, I.B.; Krake, L.R.; Rigden, J.E.; Rezaian, M.A. A novel subviral agent associated with a geminivirus: the first report of a DNA satellite. Proc. Natl. Acad. Sci. U.S.A. 1997, 94, 7088–7093. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.J.; Zhou, X.P. Interaction between a nanovirus-like component and the Tobacco curly shoot virus/satellite complex. Acta. Biochim. Biophys. Sin. 2005, 37, 25–31. [Google Scholar] [CrossRef]
- Briddon, R.W.; Stanley, J. Subviral agents associated with plant single-stranded DNA viruses. Virology 2006, 344, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Briddon, R.W.; Brown, J.K.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X.; Fauquet, C.M. Recommendations for the classification and nomenclature of the DNA-beta satellites of begomoviruses. Arch. Virol. 2008, 153, 763–81. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.J.; Ryabov, E.V.; Raja, S.K.; Roberts, I.M.; Taliansky, M.E. Satellite RNA is essential for encapsidation of groundnut rosette umbravirus RNA by groundnut rosette assistor luteovirus coat protein. Virology 1999, 254, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Ban, N.; Larson, S.B.; McPherson, A. Structural comparison of the plant satellite viruses. Virology 1995, 214, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Olivier, V.; Blanchard, P.; Chaouch, S.; Lallemand, P.; Schurr, F.; Celle, O.; Dubois, E.; Tordo, N.; Thiery, R.; Houlgatte, R.; Ribiere, M. Molecular characterisation and phylogenetic analysis of Chronic bee paralysis virus, a honey bee virus. Virus Res. 2008, 132, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.l Zitter; Palukaitis, P. Helper virus-dependent replication, nucleotide sequence and genome organization of the satellite virus of maize white line mosaic virus. Virology 1991, 180, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Masuta, C.; Zuidema, D.; Hunter, B.G.; Heaton, L.A.; Sopher, D.S.; Jackson, A.O. Analysis of the genome of satellite panicum mosaic virus. Virology 1987, 159, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Mirkov, T.E.; Mathews, D.M.; Du Plessis, D.H,; Dodds, J.A. Nucleotide sequence and translation of satellite tobacco mosaic virus RNA. Virology 1989, 170, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.W.; Browning, K.S.; Heckman, J.E.; RajBhandary, U.L.; Clark, J.M.J. Nucleotide sequence of the 5' terminus of satellite tobacco necrosis virus ribonucleic acid. Biochemistry 1979, 18, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Ysebaert, M. Total nucleotide sequence of a nearly full-size DNA copy of satellite tobacco necrosis virus RNA. J. Mol. Biol. 1980, 143, 273–287. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Omarov, R.T.; Scholthof, K.B. The complex subcellular distribution of satellite panicum mosaic virus capsid protein reflects its multifunctional role during infection. Virology 2008, 376, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Dodds, J.A. Satellite tobacco mosaic virus. Annu. Rev. Phytopathol. 1998, 36, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Yassi, M.N.A.; Dodds, J.A. Specific sequence changes in the 5'-terminal region of the genome of satellite tobacco mosaic virus are required for adaptation to tobacco mosaic virus. J. Gen. Virol. 1998, 79, 905–913. [Google Scholar] [PubMed]
- Satellite ssRNA, 3rdBuchen-Osmond, C. (Ed.) Columbia University: New York, NY, USA, 2006.
- Hemmer, O.; Oncino, C.; Fritsch, C. Efficient replication of the in vitro transcripts from cloned cDNA of tomato black ring virus satellite RNA requires the 48K satellite RNA-encoded protein. Virology 1993, 194, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Hans, F.; Pinck, M.; Pinck, L. Location of the replication determinants of the satellite RNA associated with grapevine fanleaf nepovirus (strain F13). Biochimie 1993, 75, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Cooper, J.I. The multiplication in plants of arabis mosaic virus satellite RNA requires the encoded protein. J. Gen. Virol. 1993, 74, 1471–1474. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.S.; Lin, B.Y.; Lo, N.W.; Hu, C.C.; Chow, T;Y.; Hsu, Y.H. Nucleotide sequence of the genomic RNA of bamboo mosaic potexvirus. J. Gen. Virol. 1994, 75, 2513–2518. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.S.; Lee, Y.S.; Lin, B.Y.; Lee, C.W.; Hsu, Y.H. The open reading frame of bamboo mosaic potexvirus satellite RNA is not essential for its replication and can be replaced with a bacterial gene. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 3138–3142. [Google Scholar] [CrossRef] [PubMed]
- Palani, P.V.; Kasiviswanathan, V.; Chen, J.C.; Chen, W.; Hsu, Y.H.; Lin, N.S. The arginine-rich motif of bamboo mosaic virus satellite RNA-encoded P20 mediates self-interaction, intracellular targeting, and cell-to-cell movement. Mol. Plant Microbe Interact. 2006, 19, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.S.; Hsu, Y.H.; Lin, N.S. Bamboo mosaic potexvirus satellite RNA (satBaMV RNA)-encoded P20 protein preferentially binds to satBaMV RNA. J. Virol. 1999, 73, 3032–3039. [Google Scholar] [PubMed]
- Palani, P.V.; Chiu, M.; Chen, W.; Wang, C.C.; Lin, C.C.; Hsu, C.C.; Cheng, C.P.; Chen, C.M.; Hsu, Y.H.; Lin, N.S. Subcellular localization and expression of bamboo mosaic virus satellite RNA-encoded protein. J. Gen. Virol. 2009, 90, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Latvala-Kilby, S.; Lemmetty, A.; Lehto, K. Molecular characterization of a satellite RNA associated with blackcurrant reversion nepovirus. Arch. Virol. 2000, 145, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.E. Replication, recombination, and symptom-modulation properties of the satellite RNAs of turnip crinkle virus. Curr. Top. Microbiol. Immunol. 1999, 239, 19–36. [Google Scholar] [PubMed]
- Simon, A.E.; Howell, S.H. The virulent satellite RNA of turnip crinkle virus has a major domain homologous to the 3' end of the helper virus genome. EMBO J. 1986, 5, 3423–3428. [Google Scholar] [PubMed]
- Etscheid, M.; Tousignant, M.E.; Kaper, J.M. Small satellite of arabis mosaic virus: autolytic processing of in vitro transcripts of (+) and (-) polarity and infectivity of (+) strand transcripts. J. Gen. Virol. 1995, 76, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Song, S.I.; Silver, S.L.; Aulik, M.A.; Rasochova, L.; Mohan, B.R.; Miller, W.A. Satellite cereal yellow dwarf virus-RPV (satRPV) RNA requires a double hammerhead for self-cleavage and an alternative structure for replication. J. Mol. Biol. 1999, 293, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Song, S.I.; Miller, W.A. Cis and trans requirements for rolling circle replication of a satellite RNA. J. Virol. 2004, 78, 3072–3082. [Google Scholar] [CrossRef] [PubMed]
- Menzel, W.; Maiss, E.; Vetten, H.J. Nucleotide sequence of a satellite RNA associated with carrot motley dwarf in parsley and carrot. Virus Genes 2009, 38, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Hajimorad, M.R.; Ghabrial, S.A.; Roossinck, M.J. De novo emergence of a novel satellite RNA of cucumber mosaic virus following serial passages of the virus derived from RNA transcripts. Arch. Virol. 2009, 154, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.H.; Cao, Y.H.; Li, D.W.; Niu, S.N.; Cai, Z.N.; Han, C.G.; Zhai, Y.F.; Yu, J.L. Analysis of nucleotide sequences and multimeric forms of a novel satellite RNA associated with beet black scorch virus. J. Virol. 2005, 79, 3664–3674. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Simon, A.E. Enhanced viral pathogenesis associated with a virulent mutant virus or a virulent satellite RNA correlates with reduced virion accumulation and abundance of free coat protein. Virology 2003, 312, 8–13. [Google Scholar] [CrossRef]
- Manfre, A.J.; Simon, A.E. Importance of coat protein and RNA silencing in satellite RNA/virus interactions. Virology 2008, 379, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Scholthof, K.B. Satellite panicum mosaic virus capsid protein elicits symptoms on a nonhost plant and interferes with a suppressor of virus-induced gene silencing. Mol. Plant Microbe Interact. 2004, 17, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Omarov, R.T.; Qi, D.; Scholthof, K.B. The capsid protein of satellite panicum mosaic virus contributes to systemic invasion and interacts with its helper virus. J. Virol. 2005, 79, 9756–9764. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.G. A synergism induced by satellite panicum mosaic virus. Mol. Plant-Microbe Interact. 1999, 12, 163–166. [Google Scholar] [CrossRef]
- Everett, A.L.; Scholthof, H.B.; Scholthof, K.B.G. Satellite panicum mosaic virus coat protein enhances the performance of plant virus gene vectors. Virology 2010, 396, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Scholthof, K.B. Multiple activities associated with the capsid protein of satellite panicum mosaic virus are controlled separately by the N- and C-terminal regions. Mol. Plant Microbe Interact. 2008, 21, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Danthinne, X.; Seurinck, J.; Meulewaeter, F.; Van Montagu, M.; Cornelissen, M. The 3' untranslated region of satellite tobacco necrosis virus RNA stimulates translation in vitro. Mol. Cell Biol. 1993, 13, 3340–3349. [Google Scholar] [PubMed]
- Timmer, R.T.; Benkowski, L.A.; Schodin, D.; Lax, S.R.; Metz, A.M.; Ravel, J.M.; Browning, K.S. The 5' and 3' untranslated regions of satellite tobacco necrosis virus RNA affect translational efficiency and dependence on a 5' cap structure. J. Biol. Chem. 1993, 268, 9504–9510. [Google Scholar] [PubMed]
- Meulewaeter, F.; Danthinne, X.; Van Montagu, M.; Cornelissen, M. 5'- and 3'-sequences of satellite tobacco necrosis virus RNA promoting translation in tobacco. Plant J. 1998, 14, 169–76. [Google Scholar] [CrossRef] [PubMed]
- van Lipzig, R.; van Montagu, M.; Cornelissen, M.; Meulewaeter, F. Functionality of the STNV translational enhancer domain correlates with affinity for two wheat germ factors. Nucleic Acids Res. 2001, 29, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- van Lipzig, R.; Gultyaev, A.P.; Pleij, C.W.; van Montagu, M.; Cornelissen, M.; The 5' and 3' extremities of the satellite tobacco necrosis virus translational enhancer domain contribute differentially to stimulation of translation. RNA 2002, 8, 229–236. [Google Scholar] [CrossRef]
- Gazo, B.M.; Murphy, P.; Gatchel, J.R.; Browning, K.S. A novel interaction of Cap-binding protein complexes eukaryotic initiation factor (eIF) 4F and eIF(iso)4F with a region in the 3'-untranslated region of satellite tobacco necrosis virus. J. Biol. Chem. 2004, 279, 13584–13592. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Miller, W.A. The 3' untranslated region of tobacco necrosis virus RNA contains a barley yellow dwarf virus-like cap-independent translation element. J. Virol. 2004, 78, 4655–4664. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Simon, A.E. 3'-End stem-loops of the subviral RNAs associated with turnip crinkle virus are involved in symptom modulation and coat protein binding. J. Virol. 2000, 74, 6528–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, G.; Guo, R.; Shapiro, B.A.; Simon, A.E. A pseudoknot in a preactive form of a viral RNA is part of a structural switch activating minus-strand synthesis. J. Virol. 2006, 80, 9181–9191. [Google Scholar] [CrossRef] [PubMed]
- Zhang J, Zhang G. Evolution of virus-derived sequences for high-level replication of a subviral RNA. Virology 2006, 351, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Shi, K.; Meskauskas, A.; Simon, A.E. The 3' end of turnip crinkle virus contains a highly interactive structure including a translational enhancer that is disrupted by binding to the RNA-dependent RNA polymerase. RNA 2009, 15, 1849–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, J.; George, A.T.; Baumstark, T.; Simon, A.E. Conformational changes involved in initiation of minus-strand synthesis of a virus-associated RNA. RNA 2006, 12, 147–62. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Lin, W.; Zhang, J.; Simon, A.E. Kushner. J. Virol. 2009, 83, 927–939. [CrossRef] [PubMed]
- Stupina, V.A.; Meskauskas, A.; McCormack, J.C.; Yingling, Y.G.;; Simon, A.E. The 3' proximal translational enhancer of turnip crinkle virus binds to 60S ribosomal subunits. RNA 2008, 14, 2379–93. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.E.; Gehrke, L. RNA conformational changes in the life cycles of RNA viruses, viroids, and virus-associated RNAs. Biochim. Biophys. Acta 2009. [Google Scholar]
- Annamalai, P.; Hsu, Y.H.; Liu, Y.P.; Tsai, C.H.; Lin, N.S. Structural and mutational analyses of cis-acting sequences in the 5'-untranslated region of satellite RNA of bamboo mosaic potexvirus. Virology 2003, 311, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.W.; Hu, C.C.; Lin, C.A.; Liu, Y.P.; Tsai, C.H.; Lin, N.S.; Hsu, Y.H. Structural and functional analyses of the 3' untranslated region of bamboo mosaic virus satellite RNA. Virology 2009, 386, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.W.; Hsu, Y.H.; Tsai, C.H. Specificity analysis of the conserved hexanucleotides for the replication of bamboo mosaic potexvirus RNA. Virus Res. 2002, 83, 159–167. [Google Scholar] [CrossRef] [PubMed]
- White, K.A.; Bancroft, J.B.; Mackie, G.A. Mutagenesis of a hexanucleotide sequence conserved in potexvirus RNAs. Virology 1992, 189, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Rubino, L.; Pantaleo, V.; Navarro, B.; Russo, M. Expression of tombusvirus open reading frames 1 and 2 is sufficient for the replication of defective interfering, but not satellite, RNA. J. Gen. Virol. 2004, 85, 3115–3122. [Google Scholar] [CrossRef] [PubMed]
- Takanami, Y. A striking change in symptoms on cucumber mosaic virus-infected tobacco plants induced by a satellite RNA. Virology 1981, 109, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Gonsalves, D.; Provvidenti, R.; Edwards, M.C. Tomato white leaf: the relation of an apparent satellite RNA and cucumber mosaic virus Lycopersicon esculentum, new disease detected in 1970. Phytopathology 1982, 72, 1533–1538. [Google Scholar] [CrossRef]
- Palukaitis, P. Pathogenicity regulation by satellite RNAs of cucumber mosaic virus: minor nucleotide sequence changes alter host responses. Mol. Plant Microbe Interact. 1988, 1, 175–181. [Google Scholar] [PubMed]
- Ratcliff, F.; Harrison, B.D.; Baulcombe, D.C. A similarity between viral defense and gene silencing in plants. Science 1997, 276, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Vance, V.; Vaucheret, H. RNA silencing in plants -- defense and counterdefense. Science 2001, 292, 2277–2280. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ren, T.; Morris, T.J. The coat protein of turnip crinkle virus suppresses posttranscriptional gene silencing at an early initiation step. J. Virol. 2003, 77, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.L.; Leh, V.; Lederer, C.; Maul, A.J. Turnip crinkle virus coat protein mediates suppression of RNA silencing in Nicotiana benthamiana. Virology 2003, 206, 33–41. [Google Scholar] [CrossRef]
- Kaper, J.M.; Tousignant, M.E. Cucumber mosaic virus-associating RNA 5 I. Role of host plant and helper strain in determining amount of associated RNA 5 with virions. Virology 1977, 80, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Devic, M.; Jaegle, M.; Baulcombe, D. Cucumber mosaic virus satellite RNA (strain Y): analysis of sequences which affect systemic necrosis on tomato. J. Gen. Virol. 1990, 71, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Jaegle, M.; Devic, M.; Longstaff, M.; Baulcombe, D. Cucumber mosaic virus satellite RNA (Y strain): analysis of sequences which affect yellow mosaic symptoms on tobacco. J. Gen. Virol. 1990, 71, 1905–1912. [Google Scholar] [CrossRef] [PubMed]
- Sleat, D.E. Nucleotide sequence of a new satellite RNA of cucumber mosaic virus. Nucl. Acids Res. 1990, 18, 3416. [Google Scholar] [CrossRef]
- Sleat, D.E.; Palukaitis, P. Induction of tobacco chlorosis by certain cucumber mosaic virus satellite RNAs is specific to subgroup II helper strains. Virology 1990, 176, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Sleat, D.E.; Palukaitis, P. Site-directed mutagenesis of a plant viral satellite RNA changes its phenotype from ameliorative to necrogenic [published erratum appears in Proc Natl Acad Sci U S A 1991 Feb 1; 88(3):1084]. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 2946–2950. [Google Scholar] [CrossRef] [PubMed]
- Palukaitis, P.; Roossinck, M.J.; Dietzgen, R.G.; Francki, R.I. Cucumber mosaic virus. Adv. Virus. Res. 1992, 41, 281–348. [Google Scholar] [PubMed]
- Sleat, D.E.; Palukaitis, P. A single nucleotide change within a plant virus satellite RNA alters the host specificity of disease induction. Plant J. 1992, 2, 43–49. [Google Scholar] [PubMed]
- Naidu, R.A.; Collins, G.B.; Ghabrial, S.A. Symptom-modulating properties of peanut stunt virus satellite RNA sequence variants. Mol. Plant Microbe Interact. 1991, 4, 268–275. [Google Scholar] [PubMed]
- Jacquemond, M.; Lauquin, G.J. The cDNA of cucumber mosaic virus-associated satellite RNA has in vivo biological properties. Biochem. Biophys. Res. Commun. 1988, 151, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Masuta, C.; Takanami, Y. Determination of sequence and structural requirements for pathogenicity of a cucumber mosaic virus satellite RNA (Y-satRNA). Plant Cell 1989, 1, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Devic, M.; Jaegle, M; Baulcombe, D. Cucumber mosaic virus satellite RNA (strain Y): analysis of sequences which affect systemic necrosis on tomato. J. Gen. Virol. 1990, 71, 1443–1449. [Google Scholar] [CrossRef] [PubMed]
- Sleat, D.E.; Palukaitis, P. Site-directed mutagenesis of a plant viral satellite RNA changes its phenotype from ameliorative to necrogenic. Proc. Natl. Acad. Sci. U. S. A. 1990, 87, 2946–2950. [Google Scholar] [CrossRef] [PubMed]
- Naidu, R.A.; Collins, G.B.; Ghabrial, S.A. Peanut stunt virus satellite RNA: analysis of sequences that affect symptom attenuation in tobacco. Virology 1992, 189, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Irian, S.; Xu, P.; Dai, X.; Zhao, P.X.; Roossinck, M.J. Regulation of a virus-induced lethal disease in tomato revealed by LongSAGE analysis. Mol. Plant Microbe Interact. 2007, 20, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Wheeler, D.L. GenBank. Nucleic Acids Res. 2008, 36, D25–D30. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.S.; Duan, C.G.; Zhang, Z.H.; Fang, Y.Y.; Fang, R.X.; Xie, Q.; Guo, H.S. DCL4 targets cucumber mosaic virus satellite RNA at novel secondary structures. J. Virol. 2007, 81, 9142–9151. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Zhu, L. Satellite RNA-mediated reduction of cucumber mosaic virus genomic RNAs accumulation in Nicotiana tabacum. Acta Biochim. Biophys. Sin. (Shanghai) 2007, 39, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.H.; Chen, H.C.; Cheng, J.; Annamalai, P.; Lin, B.Y.; Wu, C.T.; Yeh, W.B.; Lin, N.S. Crucial role of the 5' conserved structure of bamboo mosaic virus satellite RNA in downregulation of helper viral RNA replication. J. Virol. 2006, 80, 2566–2574. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Hsu, Y.H.; Lin, N.S. Downregulation of bamboo mosaic virus replication requires the 5' apical hairpin stem loop structure and sequence of satellite RNA. Virology 2007, 365, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Ding, S.W. Viral suppressors of RNA silencing. Curr. Opin. Biotechnol. 2001, 12, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Moissiard, G.; Voinnet, O. Viral suppression of RNA silencing in plants. Mol. Plant Pathol. 2004, 5, 71–82. [Google Scholar] [CrossRef]
- Roth, B.M.; Pruss, G.J.; Vance, V.B. Plant viral suppressors of RNA silencing. Virus Res. 2004, 102, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Howell, S.H. Nucleic acid species related to the satellite RNA of turnip crinkle virus in turnip plants and virus particles. Virology 1984, 134, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D.C.; Saunders, G.R.; Bevan, M.W.; Mayo, M.A.; Harrison, B.D. Expression of biologically active viral satellite RNA from the nuclear genome of transformed plants. Nature 1986, 321, 446–449. [Google Scholar] [CrossRef]
- Carpenter, C.D.; Simon, A.E. In vivo restoration of biologically active 3' ends of virus-associated RNAs by nonhomologous RNA recombination and replacement of a terminal motif. J. Virol. 1996, 70, 478–486. [Google Scholar] [PubMed]
- Carpenter, C.D.; Simon A.E. In vivo repair of 3'-end deletions in a TCV satellite RNA may involve two abortive synthesis and priming events. Virology 1996, 226, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Nagy, P.D.; Carpenter, C.D.; Simon, A.E. A novel 3'-end repair mechanism in an RNA virus. Proc. Natl. Acad. Sci. USA 1997, 94, 1113–1118. [Google Scholar] [CrossRef]
- Liu, J.S.; Lin, N.S. Satellite RNA associated with bamboo mosaic potexvirus shares similarity with satellites associated with sobemoviruses. Arch. Virol. 1995, 140, 1511–1514. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.S.; Hsu, Y.H.; Huang, T.Y.; Lin, N.S. Molecular evolution and phylogeny of satellite RNA associated with bamboo mosaic potexvirus. J. Mol. Evol. 1997, 44, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Llave, C.; Kasschau, K.D.; Rector, M.A.; Carrington, J.C. Endogenous and silencing-associated small RNAs in plants. Plant Cell 2002, 14, 1605–1619. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Reinhart, B.J.; Bartel, D.P.; Zamore, P.D. A biochemical framework for RNA silencing in plants. Genes Dev. 2003, 17, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Van Valen, L. A new evolutionary law. Evolutionary Theory 1973, 1, 1–30. [Google Scholar]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucl. Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignments through sequence weighting, position specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Sun, X.; Simon, A.E. A cis-replication element functions in both orientations to enhance replication of turnip crinkle virus. Virology 2006, 352, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Alvarado, G.; Roossinck, M.J. Structural analysis of a necrogenic strain of cucumber mosaic cucumovirus satellite RNA in planta. Virology 1997, 236, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.K.; Hu, C.C.; Lin, N.S.; Chang, B.Y.; Hsu, Y.H. Movement of potexviruses requires species-specific interactions among the cognate triple gene block proteins, as revealed by a trans-complementation assay based on the bamboo mosaic virus satellite RNA-mediated expression system. J. Gen. Virol. 2006, 87, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Hsu, Y.H.; Lin, N.S. Generation of subgenomic RNA directed by a satellite RNA associated with bamboo mosaic potexvirus: analyses of potexvirus subgenomic RNA promoter. J. Virol. 2000, 74, 10341–10348. [Google Scholar] [CrossRef] [PubMed]
- Gossele, V.; Fache, I.; Meulewaeter, F.; Cornelissen, M.; Metzlaff, M. SVISS - a novel transient gene silencing system for gene function discovery and validation in tobacco plants. Plant J. 2002, 32, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Tien, P.; Zhang, X.H.; Qiu, B.S.; Qin, B.Y.; Wu, G.S. Satellite RNA for the control of plant diseases caused by cucumber mosaic virus. Ann. Appl. Biol. 1987, 111, 143–153. [Google Scholar] [CrossRef]
- Harrison, B.D.; Mayo, M.A.; Baulcombe, D.C. Virus resistance in transgenic plants that express cucumber mosaic virus satellite RNA. Nature 1987, 328, 799–802. [Google Scholar] [CrossRef]
- Gerlach, W.L.; Llewellyn, D.; Haseloff, J. Construction of a plant disease resistance gene from the satellite RNA of tobacco ringspot virus. Nature 1987, 328, 802–805. [Google Scholar] [CrossRef]
- Jacquemond, M.; Amselem, J.; Tepfer, M. A gene coding for a monomeric form of cucumber mosaic virus satellite RNA confers tolerance to CMV. Mol. Plant Microbe Interact. 1988, 1, 311–316. [Google Scholar] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Hu, C.-C.; Hsu, Y.-H.; Lin, N.-S. Satellite RNAs and Satellite Viruses of Plants. Viruses 2009, 1, 1325-1350. https://doi.org/10.3390/v1031325
Hu C-C, Hsu Y-H, Lin N-S. Satellite RNAs and Satellite Viruses of Plants. Viruses. 2009; 1(3):1325-1350. https://doi.org/10.3390/v1031325
Chicago/Turabian StyleHu, Chung-Chi, Yau-Heiu Hsu, and Na-Sheng Lin. 2009. "Satellite RNAs and Satellite Viruses of Plants" Viruses 1, no. 3: 1325-1350. https://doi.org/10.3390/v1031325