Vaccinia Virus in Blood Samples of Humans, Domestic and Wild Mammals in Brazil


Abstract
:1. Introduction
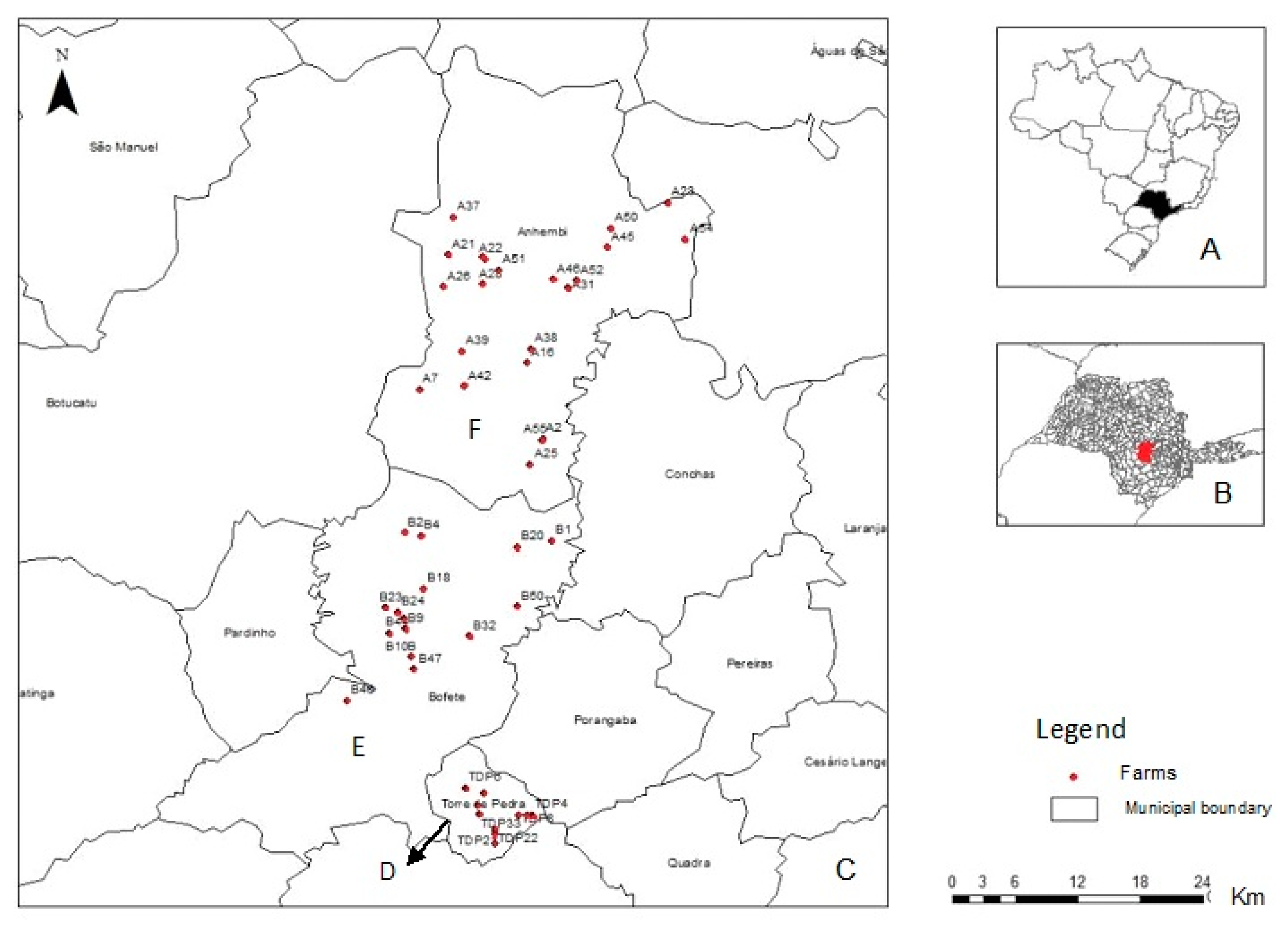
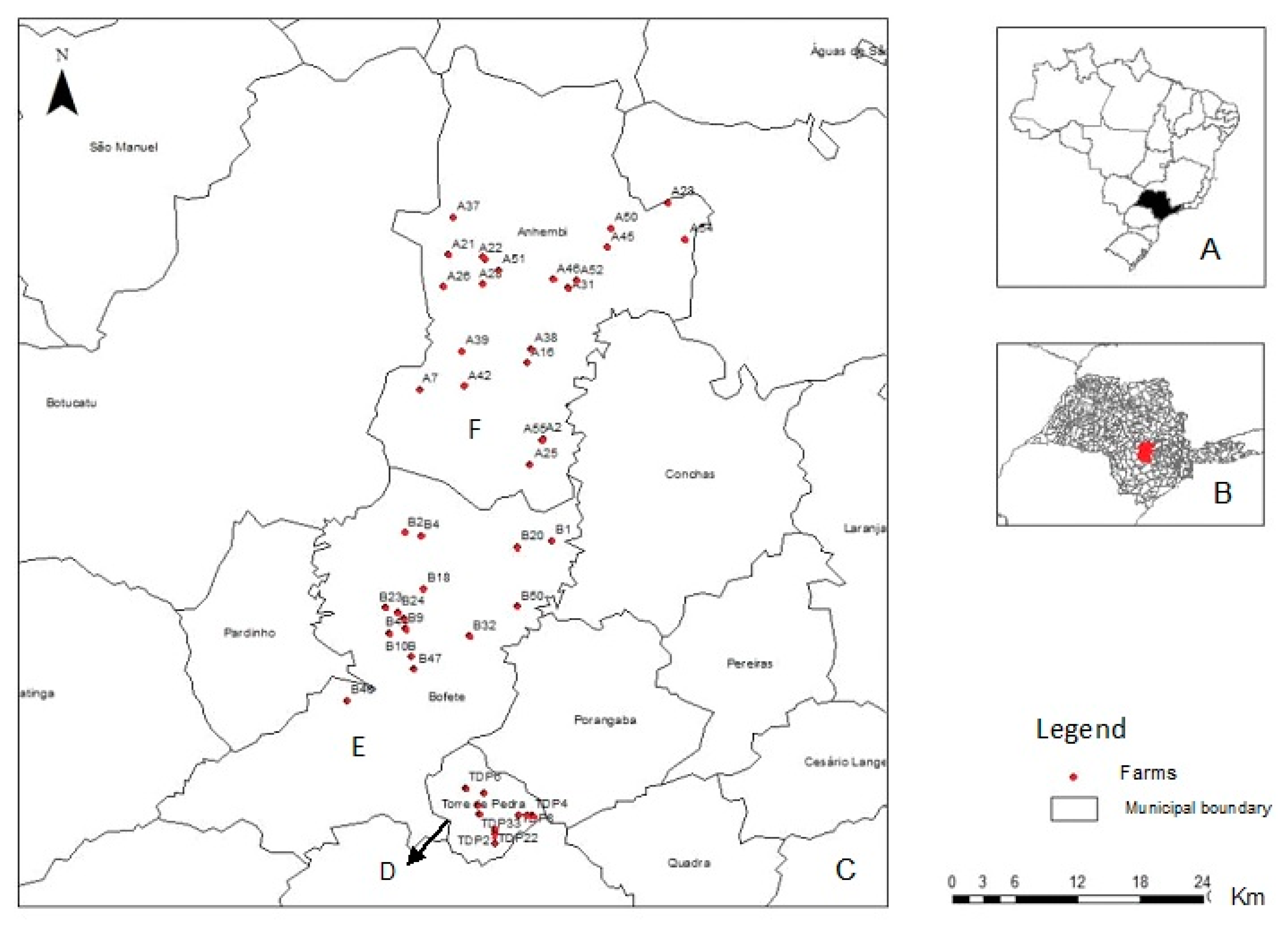
2. Materials and Methods
3. Results
3.1. Missing Values
3.2. Samples Collected and Positivity
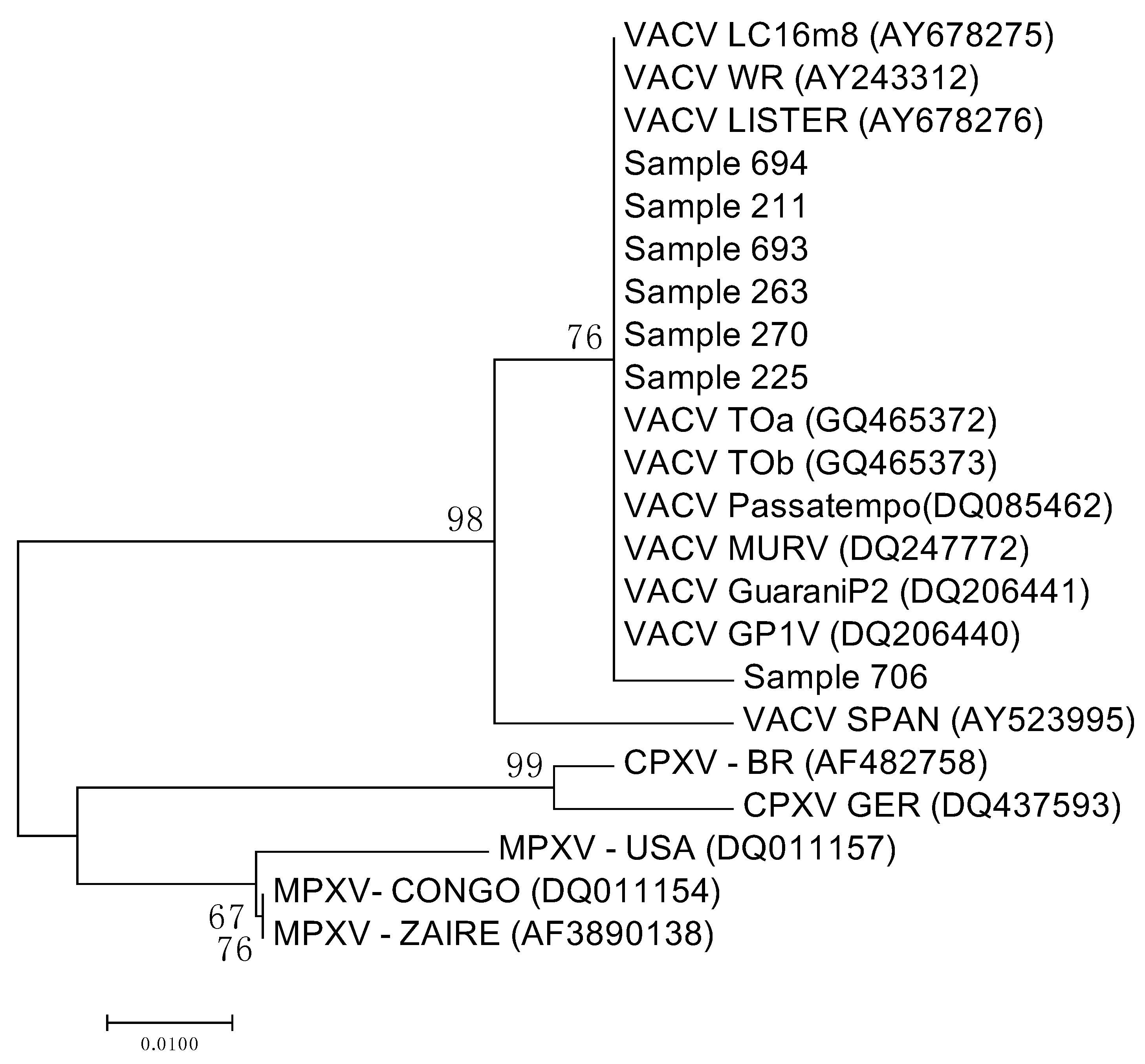
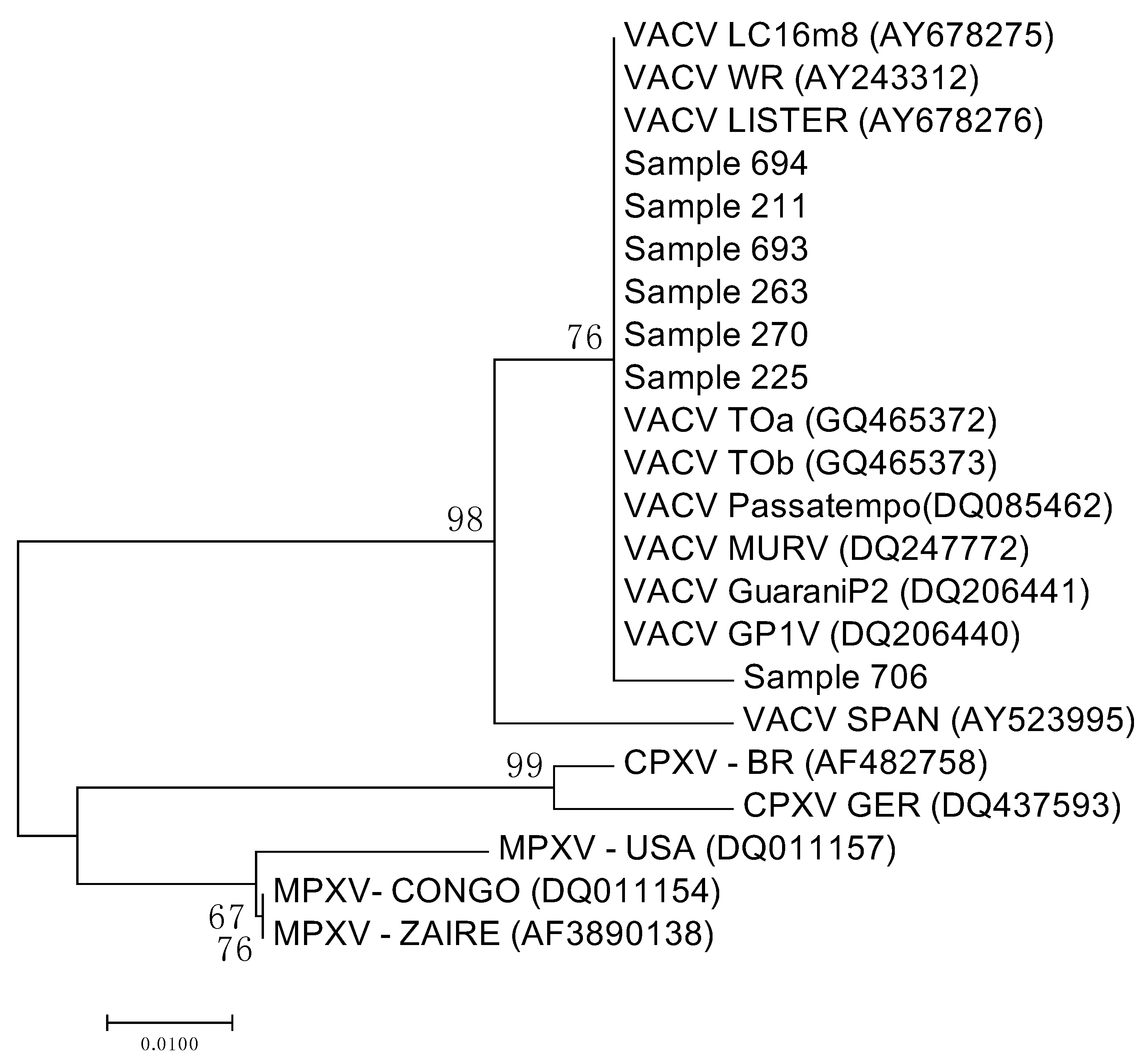
3.3. Sequencing and Analysis of the Sequences
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Damaso, C.R.A.; Esposito, J.J.; Condit, R.C.; Moussatche, N. An emergente poxvirus from humans and cattle in Rio de Janeiro State: Cantagalo Virus may derive from Brazilian Smallpox Vacine. Virology 2000, 277, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Negasse-segahara, T.K.; Kisielius, J.J.; Ueda-Ito, M.; Curtis, S.P.; Figueiredo, C.A.; Cruz, A.S.; Silv, M.M.J.; Ramos, C.H.; Silva, M.C.C.; Sakurati, T.; et al. Human Vaccinia-like outbreaks in São Paulo and Goiás States, Brazil: Vírus detection, isolation and identification. Rev. Inst. Med. Trop. São Paulo 2004, 46, 315–322. [Google Scholar] [CrossRef]
- Megid, J.; Appolinário, C.M.; Langoni, H.; Pituco, E.M.; Okuda, L.H. Short Report: Vaccinia virus in humans and cattle in southwest region of São Paulo State, Brazil. Am. J. Trop. Med. Hyg. 2008, 79, 647–651. [Google Scholar] [PubMed]
- Abrahão, J.S.; Sival-Fernandes, A.T.; Assis, F.L.; Guedes, M.I.; Drumond, B.P.; Leite, J.A.; Coelho, L.F.L.; Turrini, F.; Fonseca, F.G.; Lobato, Z.I.P.; et al. Human Vaccinia vírus and Pseudocowpox viru co-infection: Clinical description and phylogenetic characterization. J. Clin. Virol. 2010, 48, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Brum, M.C.S.; Anjos, B.L.; Nogueira, C.E.W.; Amaral, L.A.; Wiblen, R.; Flores, E.F. An outbreak of orthopoxvirus-associated disease in horse in Southern Brazil. J. Vet. Diagn. Investig. 2010, 22, 143–147. [Google Scholar]
- Megid, J.; Borges, I.A.; Abrahão, J.S.; Trindade, G.S.; Appolinário, C.M.; Ribeiro, M.G.; Allendorf, S.D.; Antunes, J.M.A.P.; Silva-Fernandes, A.T.; Kroon, E.G. Vaccinia Virus, Zoonotic Infection, São Paulo State, Brazil. J. Emerg. Infect. Dis. 2012, 18, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Assis, F.L.; Vinhote, W.M.; Barbosa, J.D.; Oliveira, C.H.S.; Oliveira, C.M.G.; Campos, K.F.; Silva, N.S.; Trindade, G.S.; Abrahão, J.S.; Kroon, E.G. Reemergence of Vaccinia vírus during zoonotic outbreak, Pará State, Brazil. Emerg. Infect. Dis. 2013, 19, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.P.; Fernandes, A.T.S.; Assis, F.L.; Alves, P.A.; Luiz, A.P.M.F.; Figueiredo, L.B.; Almeida, C.M.C.; Travassos, C.E.P.F.; Trindade, G.S.; Abrahão, J.S.; et al. Short Report: Intrafamilial transmission of Vaccinia vírus during a bovine Vaccinia outbreak in Brazil: A New Insight in Viral Transmission Chain. Am. J. Trop. Med. Hyg. 2014, 90, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; Campos, R.K.; Trindade, G.S.; Fonseca, F.G.; Ferreira, P.C.P.; Kroon, E.G. Outbreak of severe zoonotic Vaccinia vírus infection, Southeastern Brazil. Emerg. Infect. Dis. 2015, 21, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Moussatche, N.; Damaso, C.R.; McFadden, G. When good vaccine go wild: Feral Orthopoxvirus in developing countries and beyond. J. Infect. Dev. Ctries. 2008, 2, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Trindade, G.S.; Emerson, G.L.; Carroli, D.S.; Kroon, E.G.; Damon, I.K. Brazilian Vaccinia virus and their origigns. Emerg. Infect. Dis. 2007, 13, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Drumond, B.P.; Leite, J.A.; da Fonseca, F.G.; Bonjardim, C.A.; Ferreira, P.C.P.; Kroon, E.G. Brzilian Vaccinia vírus strains are genetically divergente and differ from the Lister vaccine strain. Microb. Infect. 2008, 10, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; Guedes, M.I.M.; Trindade, G.S.; Fonseca, F.G.; Campos, R.K.; Mota, B.F.; Lobato, Z.I.P.; Silva-Fernandes, A.T.; Rodrigues, G.O.L.; Lima, L.S.; et al. One more piece in the VACV ecological puzzle: Could peridomestic rodents be the link between wildlife and bovine Vaccinia outbreaks in Brazil? PLoS ONE 2009, 4, e7428. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Pfeffer, M.; Meyer, H. Zoonotic poxviruses. Vet. Microbiol. 2010, 140, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.M.S.; Abrahão, J.S.; Drumond, B.P.; Oliveira, F.M.; Alves, P.A.; Pascoal-Xavier, M.A.; Lobato, Z.I.P.; Bonjardim, C.A.; Ferreira, P.C.P.; Kroon, E.G. Vaccinia vírus: Shedding and horizontal transmission in a murine model. J. Gen. Virol. 2008, 89, 2986–2991. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; Trindade, G.S.; Ferreira, J.M.S.; Campos, R.K.; Bonjardim, C.A.; Ferreira, P.C.P.; Kroon, E.G. Long-lasting stability of Vaccinia virus strains in murine feces: Implications for virus circulation and environmental maintenance. Arch. Virol. 2009, 154, 1551–1553. [Google Scholar] [CrossRef] [PubMed]
- D’Anunciação, L.; Guedes, M.I.M.; Oliveira, T.L.; Rehfeld, I.; Bonjardim, C.A.; Ferreira, P.P.; Trindade, G.S.; Lobato, Z.P.; Kroon, E.G.; Abrahão, J.S. Filling one more gap: Experimental evidence of horizontal transmission of Vaccinia virus between bovines and rodents. Vector Borne Zoonotic Dis. 2012, 12, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Peres, M.G.; Bacchiega, T.S.; Appolinário, C.M.; Vicente, A.F.; Allendorf, S.D.; Antunes, J.M.A.P. Serological study of vaccínia virus reservoirs in areas with and without official reports of outbreaks in cattle and humans in São Paulo, Brazil. Arch. Virol. 2013, 158, 2433–2441. [Google Scholar] [CrossRef] [PubMed]
- Peres, M.G.; Barros, C.B.; Appolinário, C.M.; Antunes, J.M.A.P.; Mioni, M.S.R.; Bacchiega, T.S.; Allendorf, S.D.; Vicente, A.F.; Fonseca, C.R.; Megid, J. Dogs and opossums positive for vaccinia virus during outbreak affecting cattle and humans, São Paulo State, Brazil. J. Emerg. Infect. Dis. 2016, 22, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.B.; Borges, I.A.; Alves, P.A.; Miranda, J.B.; Luiz, A.P.M.F.; Ferreira, P.C.P.; Abrahão, J.S.; Moreno, E.C.; Kroon, E.G.; Trindade, G.S. Alternative routes of zoonotic Vaccinia virus transmission, Brazil. Emerg. Infect. Dis. 2015, 15, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I.; Hohman, P.; Preuss, J.C.; Li, L.; Fischer, S.H.; Fedorko, D.P. Detection of Vaccinia virus DNA, but not infectious virus in the blood of smallpox vaccine recipients. Vaccine 2007, 25, 4571–4574. [Google Scholar] [CrossRef] [PubMed]
- Rivetti, A.V., Jr.; Guedes, M.I.M.C.; Rehfekd, I.S.; Oliveira, T.M.L.; Matos, A.C.D.; Abrahão, J.S.; Kroon, E.G.; Lobato, Z.I.P. Bovine vaccinia, a systemic infection: Evidence of fecal shedding, viremia and detection in lymphoid organs. Vet. Microbiol. 2013, 162, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.L.V.; Cruz, M.L.; Cortopasso, S.R.G. Anestesiologia; Cubas, S.Z., Silva, I.C.R., Catão, J.L., Eds.; Tratado de Animais Selvagens: São Paulo, Brazil, 2006; pp. 1040–1067. [Google Scholar]
- Abrahão, J.S.; Lima, L.S.; Assis, F.L.; Alves, P.A.; Silva-Fernandes, A.T.; Cota, M.M.G.; Ferreira, V.M.; Campos, R.K.; Mazur, C.; Lobato, Z.I.P.; et al. Nested-multiplex PCR detection of Orthopoxvirus and Parapoxvirus directly from exanthematic clinical samples. Virol. J. 2009, 6, 140. [Google Scholar] [CrossRef] [PubMed]
- Assis, F.L.; Franco-Luiz, A.P.M.; Paim, L.M.; Oliveira, G.P.; Pereira, A.F.; Almeida, G.M.F.; Figueiredo, L.B.; Tanus, A.; Trindade, G.S.; Ferreira, P.P.; et al. Horizontal study of vaccinia virus infections in an endemic area: Epidemiologic, phylogenetic and economic aspects. Arch. Virol. 2015, 160, 2703–2708. [Google Scholar] [CrossRef] [PubMed]
- Peres, M.G.; Bacchiega, T.S. Vaccinia virus in feces and urine of wild rodents from São Paulo State, Brazil. Viruses 2018. in revision. [Google Scholar]
- Kermack, W.O.; McKendrick, A.G. A contribution to the mathematical theory of epidemics. R. Soc. Lond. Ser. A 1927, 115, 700–721. [Google Scholar] [CrossRef]
- Reis, N.L.; Perachi, A.L.; Pedro, W.A.; Lima, I.P. Mamíferos do Brasil; Universidade Estadual de Londrina: Londrina, Brazil, 2006; pp. 364–365. [Google Scholar]
- Flood, J.S.; Prophyre, T.; Tindesley, M.J.; Wollhouse, M.E. The performance of approximations of farm contiguity compared to contiguity defined using detailed geographical information in two sample areas in Scotland: Implications for foot-and-mouth disease modelling. Vet. Res. 2013, 9, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Morse, S.S. Factor in the emergence of infectious diseases. Emerg. Infect. Dis. 1995, 1, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Medaglia, M.L.G.; Pessoa, L.C.G.D.; Sales, E.R.C.; Freitas, T.R.P.; Damaso, C.R. Spread of Cantagalo Vírus to Northern Brazil. Emerg. Infect. Dis. 2009, 15, 1142–1143. [Google Scholar] [CrossRef] [PubMed]
- Quixabeira-Santos, J.C.; Medaglia, M.L.G.; Pescador, C.A.; Damaso, C.R. Animal movement and establishment of Vaccinia virus Cantagalo strain in Amazon Biome Brazil. Emerg. Infect. Dis. 2011, 17, 726–729. [Google Scholar] [CrossRef] [PubMed]
- Damaso, C.R.A.; Reis, S.A.; Jesus, D.M.; Lima, P.S.F.; Moussatché, N. A PCR-based assay for detection of emerging vaccínia-like viroses isolated in Brazil. Diagn. Microbiol. Infect. Dis. 2007, 57, 39–46. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Species | n | p | (%) |
|---|---|---|---|
| Human | 148 | 0 | (0,0) |
| Cow | 688 | 4 | (0,6) |
| Sheep | 44 | 0 | (0,0) |
| Swine | 22 | 0 | (0,0) |
| Horse | 117 | 1 | (0,8) |
| Dog | 114 | 0 | (0,0) |
| Cat | 7 | 0 | (0,0) |
| Didelphis albiventris | 57 | 1 | (1,7) |
| Didelphis aurita | 16 | 2 | (12,5) |
| Gracilinanus microtarsus | 6 | 0 | (0,0) |
| Cerdocyon thous | 4 | 0 | (0,0) |
| Nasua nasua | 4 | 0 | (0,0) |
| Leopardus pardalis | 1 | 0 | (0,0) |
| Akodon montensis | 4 | 0 | (0,0) |
| Nectomys squamipes | 4 | 0 | (0,0) |
| Calomys tener | 4 | 0 | (0,0) |
| Sooretamys angouya | 13 | 0 | (0,0) |
| Oligoryzomys flavescens | 17 | 0 | (0,0) |
| Oligoryzomys nigripes | 61 | 0 | (0,0) |
| TOTAL | 1331 | 8 | (0,6) |
| Species | Anhembi | Bofete | Torre de Pedra | |||
|---|---|---|---|---|---|---|
| n | p (%) | n | p (%) | n | p (%) | |
| Human | 82 | 0 (0,0) | 38 | 0 (0,0) | 28 | 0 (0,0) |
| Cow | 332 | 3 (0,9) | 204 | 1 (0,5) | 152 | 0 (0,0) |
| Sheep | 33 | 0 (0,0) | 9 | 0 (0,0) | 2 | 0 (0,0) |
| Swine | 9 | 0 (0,0) | 12 | 0 (0,0) | 1 | 0 (0,0) |
| Horse | 71 | 1 (1,4) | 23 | 0 (0,0) | 23 | 0 (0,0) |
| Dog | 56 | 0 (0,0) | 35 | 0 (0,0) | 23 | 0 (0,0) |
| Cat | 3 | 0 (0,0) | 2 | 0 (0,0) | 2 | 0 (0,0) |
| D. albiventris | 31 | 1 (3,2) | 19 | 0 (0,0) | 7 | 0 (0,0) |
| D. aurita | 0 | 0 (0,0) | 13 | 2 (15,4) | 3 | 0 (0,0) |
| G. microtarsus | 2 | 0 (0,0) | 2 | 0 (0,0) | 2 | 0 (0,0) |
| C. thous | 3 | 0 (0,0) | 1 | 0 (0,0) | 0 | 0 (0,0) |
| N. nasua | 3 | 0 (0,0) | 1 | 0 (0,0) | 0 | 0 (0,0) |
| L. pardalis | 1 | 0 (0,0) | 0 | 0 (0,0) | 0 | 0 (0,0) |
| A. montensis | 0 | 0 (0,0) | 4 | 0 (0,0) | 0 | 0 (0,0) |
| N. squamipes | 3 | 0 (0,0) | 1 | 0 (0,0) | 0 | 0 (0,0) |
| C. tener | 0 | 0 (0,0) | 2 | 0 (0,0) | 2 | 0 (0,0) |
| S. angouya | 13 | 0 (0,0) | 0 | 0 (0,0) | 0 | 0 (0,0) |
| O. flavescens | 11 | 0 (0,0) | 3 | 0 (0,0) | 3 | 0 (0,0) |
| O. nigripes | 30 | 0 (0,0) | 26 | 0 (0,0) | 5 | 0 (0,0) |
| TOTAL | 683 | 5 (0,7) | 395 | 3 (0,7) | 253 | 0 (0,0) |
| Serological Condition | PCR Positive (%) | PCR Negative (%) | Total (%) |
|---|---|---|---|
| Seropositive 1 | 1 * (0,5) | 185 (99,5) | 186 (100) |
| Seronegative 1 | 7 ** (0,6) | 1138 (99,4) | 1145 (100) |
| Total | 8 (0,6) | 1323 (99,4) | 1331 (100) |
| Milking Farms | PCR Positive Animals | Previous Seropositive 1 | |||
|---|---|---|---|---|---|
| Species | p (%) | n | |||
| Anhembi | A16 | Opossum (D. albiventris) | Human | 1 (25) | 4 |
| Domestic dog | 1 (50) | 2 | |||
| A2 | Cow | Cow | 1 (5) | 20 | |
| A23 | Cow | Cow | 1 (6,6) | 15 | |
| Horse | |||||
| A7 | Cow | Human | 1 (16,6) | 6 | |
| Cow | 1 (5) | 20 | |||
| Domestic dog | 2 (33,3) | 6 | |||
| Wild rodent (O. nigripes) | 1 (5,5) | 18 | |||
| Bofete | B46 | Cow * | Horse | 1 (25) | 4 |
| Domestic dog | 1 (16,6) | 6 | |||
| Cows | 5 (25) | 20 | |||
| B47 | Opossum (two D. aurita) | Human | 1 (50) | 2 | |
| Swine | 2 (33,3) | 6 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peres, M.G.; Bacchiega, T.S.; Appolinário, C.M.; Vicente, A.F.; Mioni, M.D.S.R.; Ribeiro, B.L.D.; Fonseca, C.R.S.; Pelícia, V.C.; Ferreira, F.; Oliveira, G.P.; et al. Vaccinia Virus in Blood Samples of Humans, Domestic and Wild Mammals in Brazil. Viruses 2018, 10, 42. https://doi.org/10.3390/v10010042
Peres MG, Bacchiega TS, Appolinário CM, Vicente AF, Mioni MDSR, Ribeiro BLD, Fonseca CRS, Pelícia VC, Ferreira F, Oliveira GP, et al. Vaccinia Virus in Blood Samples of Humans, Domestic and Wild Mammals in Brazil. Viruses. 2018; 10(1):42. https://doi.org/10.3390/v10010042
Chicago/Turabian StylePeres, Marina G., Thais S. Bacchiega, Camila M. Appolinário, Acácia F. Vicente, Mateus De Souza Ribeiro Mioni, Bruna L. D. Ribeiro, Clóvis R. S. Fonseca, Vanessa C. Pelícia, Fernando Ferreira, Graziele P. Oliveira, and et al. 2018. "Vaccinia Virus in Blood Samples of Humans, Domestic and Wild Mammals in Brazil" Viruses 10, no. 1: 42. https://doi.org/10.3390/v10010042