Antiviral Effects of Clinically-Relevant Interferon-α and Ribavirin Regimens against Dengue Virus in the Hollow Fiber Infection Model (HFIM)


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, and Compounds
2.2. Cytotoxicity Assay
2.3. Antiviral Evaluations
2.4. DENV Plaque Assay
2.5. Statistical Analysis
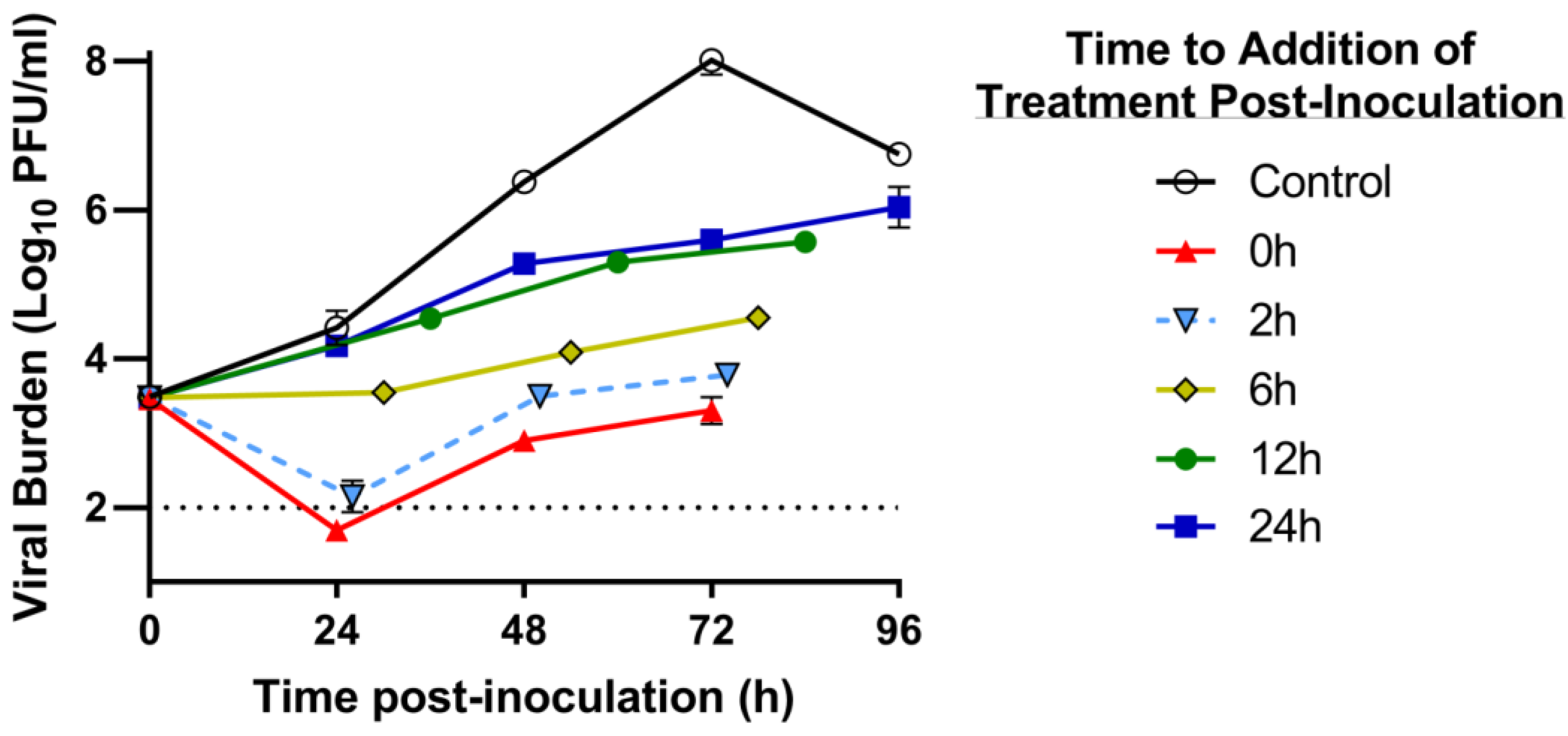
2.6. Delayed Treatment Studies in the HFIM System
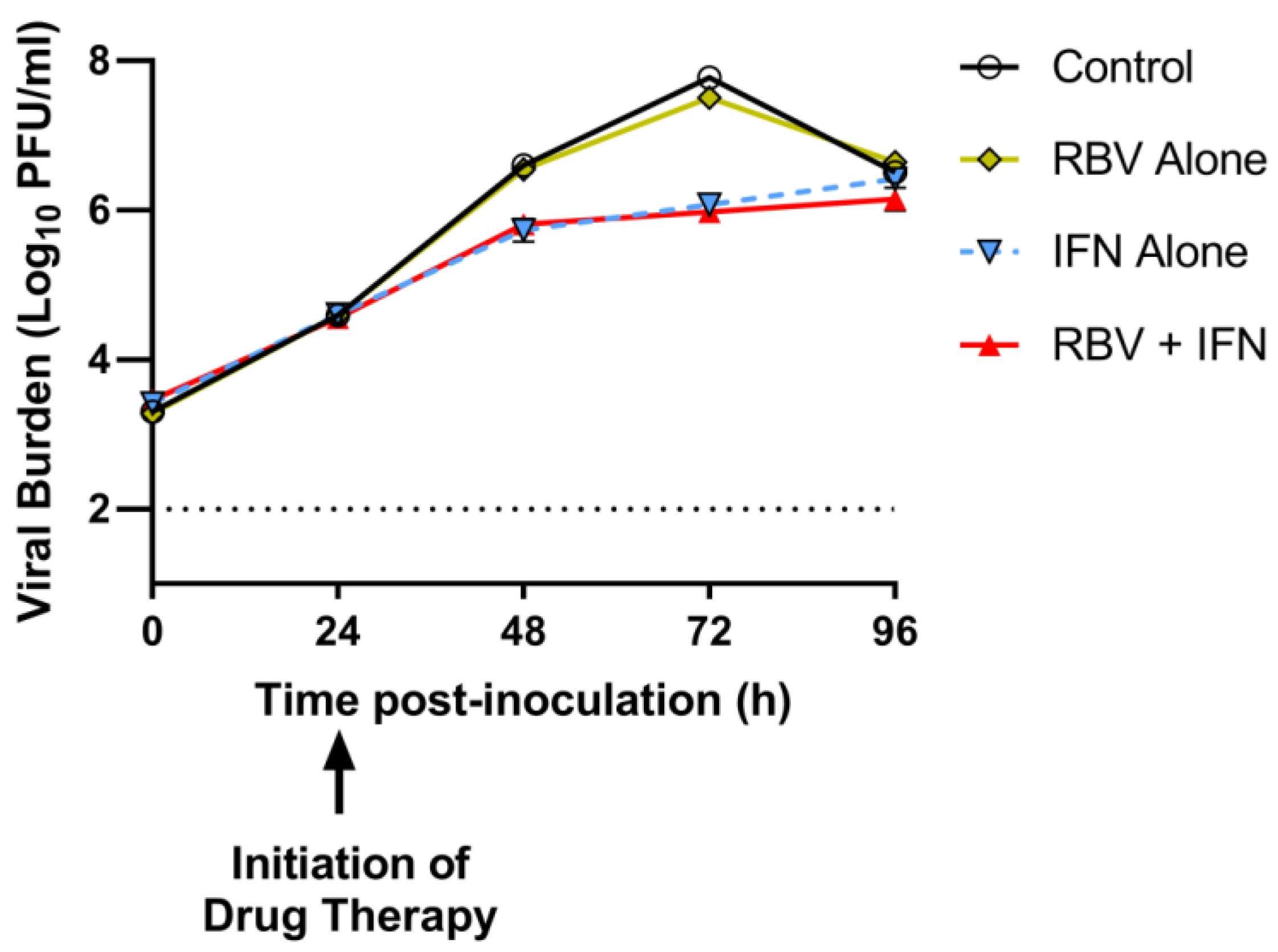
2.7. Antiviral Activity of Clinical Exposures of IFN and RBV after 24 h Post-Infection in the HFIM System
3. Results
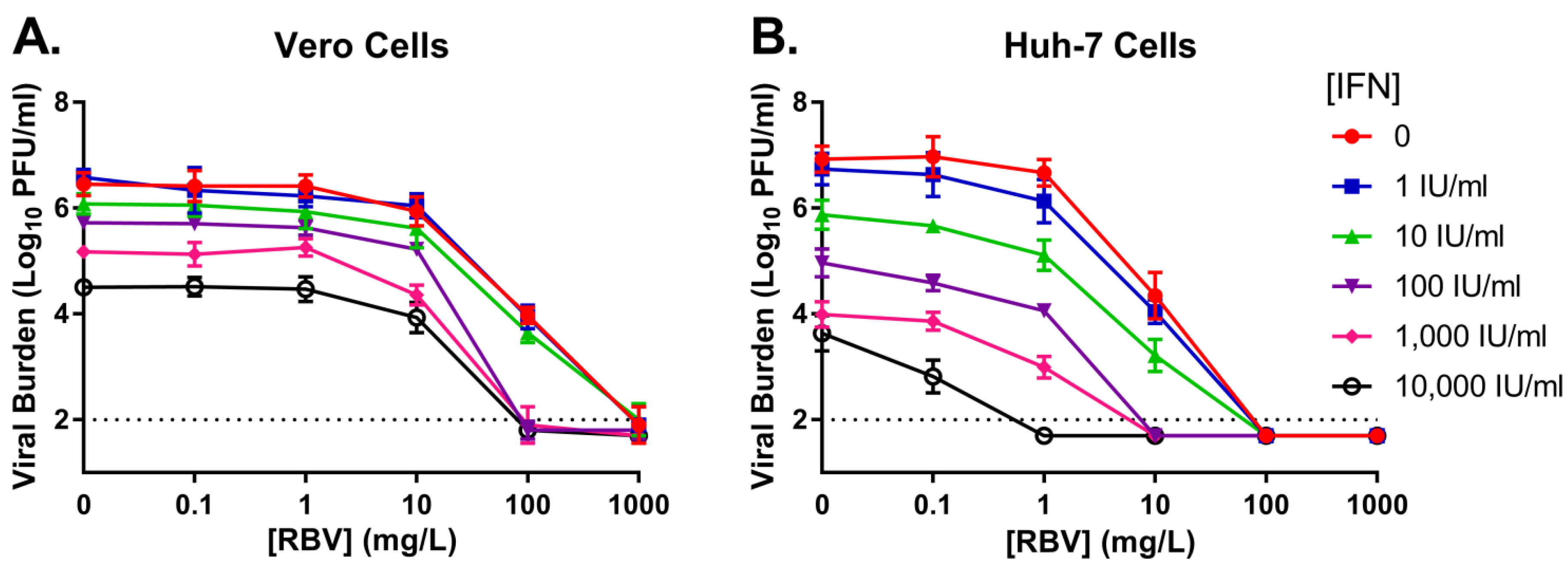
3.1. Monotherapy Antiviral Activity and Cytotoxicity
3.2. Combination Therapy against DENV in a Plate Assay
3.3. Delayed Treatment Studies in the HFIM System
3.4. Antiviral Evaluations of Clinical Exposures of IFN and/or RBV after 24 h p.i. in the HFIM System
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, C.P.; Farrar, J.J.; Nguyen, V.; Wills, B. Dengue. N. Engl. J. Med. 2012, 366, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Westaway, E.G.; Brinton, M.A.; Gaidamovich, S.Y.; Horzinek, M.C.; Igarashi, A.; Kaariainen, L.; Lvov, D.K.; Porterfield, J.S.; Russell, P.K.; Trent, D.W. Flaviviridae. Intervirology 1985, 24, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, M.S.; Rasotgi, V.; Jain, S.; Gupta, V. Discovery of fifth serotype of dengue virus (DENV-5): A new public health dilemma in dengue control. Med. J. Armed Forces India 2015, 71, 67–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Normile, D. Surprising new dengue virus throws a spanner in disease control efforts. Science 2013, 342, 415. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol. 2002, 10, 100–103. [Google Scholar] [CrossRef]
- Guzman, M.G.; Vazquez, S. The complexity of antibody-dependent enhancement of dengue virus infection. Viruses 2010, 2, 2649–2662. [Google Scholar] [CrossRef] [PubMed]
- Kalayanarooj, S. Clinical Manifestations and Management of Dengue/DHF/DSS. Trop. Med. Health 2011, 39, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhang, R.; Cheng, G. Progress towards understanding the pathogenesis of dengue hemorrhagic fever. Virol. Sin. 2017, 32, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Simmons, C.P.; McPherson, K.; Van Vinh, C.N.; Hoai Tam, D.T.; Young, P.; Mackenzie, J.; Wills, B. Recent advances in dengue pathogenesis and clinical management. Vaccine 2015, 33, 7061–7068. [Google Scholar] [CrossRef] [PubMed]
- Pang, T. SAGE committee advice on dengue vaccine. Lancet Infect. Dis. 2016, 16, 880–882. [Google Scholar] [CrossRef]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The Impact of the Newly Licensed Dengue Vaccine in Endemic Countries. PLoS Negl. Trop. Dis. 2016, 10, e0005179. [Google Scholar] [CrossRef] [PubMed]
- Low, J.G.; Sung, C.; Wijaya, L.; Wei, Y.; Rathore, A.P.S.; Watanabe, S.; Tan, B.H.; Toh, L.; Chua, L.T.; Hou, Y.; et al. Efficacy and safety of celgosivir in patients with dengue fever (CELADEN): A phase 1B, randomised, double-blind, placebo-controlled, proof-of-concept trial. Lancet Infect. Dis. 2014, 14, 706–715. [Google Scholar] [CrossRef]
- Sumarmo, D.; Talogo, W.; Asrin, A.; Isnuhandojo, B.; Sahudi, A. Failure of hydrocortisone to affect outcome in dengue shock syndrome. Pediatrics 1982, 69, 45–49. [Google Scholar] [PubMed]
- Tricou, V.; Minh, N.N.; Van, T.P.; Lee, S.J.; Farrar, J.; Wills, B.; Tran, H.T.; Simmons, C.P. A randomized controlled trial of chloroquine for the treatment of dengue in Vietnamese adults. PLoS Negl. Trop. Dis. 2010, 4, e785. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, J.; Nguyen, C.V.V.; Khanh, L.P.; Kien, D.T.H.; Quyen, N.T.H.; Tran, N.T.T.; Hang, N.T.; Truong, N.T.; Hue Tai, L.T.; Cam Huong, N.T.; et al. Lovastatin for the Treatment of Adult Patients With Dengue: A Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Infect. Dis. 2016, 62, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Borden, E.C.; Sen, G.C.; Uze, G.; Silverman, R.H.; Ransohoff, R.M.; Foster, G.R.; Stark, G.R. Interferons at age 50: Past, current and future impact on biomedicine. Nat. Rev. Drug Discov. 2007, 6, 975–990. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Harris, E. Interferon inhibits dengue virus infection by preventing translation of viral RNA through a PKR-independent mechanism. Virology 2001, 289, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Davidson, A.; Hibbert, L.; Gruenwald, P.; Schlaak, J.; Ball, S.; Foster, G.R.; Jacobs, M. Dengue virus inhibits alpha interferon signaling by reducing STAT2 expression. J. Virol. 2005, 79, 5414–5420. [Google Scholar] [CrossRef] [PubMed]
- Pires de Mello, C.P.; Tao, X.; Kim, T.H.; Bulitta, J.B.; Rodriquez, J.L.; Pomeroy, J.J.; Brown, A.N. Zika Virus Replication Is Substantially Inhibited by Novel Favipiravir and Interferon Alpha Combination Regimens. Antimicrob. Agents Chemother. 2018, 62, e01983-17. [Google Scholar] [CrossRef] [PubMed]
- Rattanaburee, T.; Junking, M.; Panya, A.; Sawasdee, N.; Songprakhon, P.; Suttitheptumrong, A.; Limjindaporn, T.; Haegeman, G.; Yenchitsomanus, P.T. Inhibition of dengue virus production and cytokine/chemokine expression by ribavirin and compound A. Antivir. Res. 2015, 124, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Feld, J.J.; Li, Q.; Hu, Z.; Fried, M.W.; Liang, T.J. Ribavirin potentiates interferon action by augmenting interferon-stimulated gene induction in hepatitis C virus cell culture models. Hepatology 2011, 53, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Greco, W.R.; Bravo, G.; Parsons, J.C. The search for synergy: A critical review from a response surface perspective. Pharmacol. Rev. 1995, 47, 331–385. [Google Scholar] [PubMed]
- Gallegos, K.M.; Drusano, G.L.; D’Argenio, D.Z.; Brown, A.N. Chikungunya Virus: In Vitro Response to Combination Therapy with Ribavirin and Interferon Alfa 2a. J. Infect. Dis. 2016, 214, 1192–1197. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.N.; McSharry, J.J.; Weng, Q.; Driebe, E.M.; Engelthaler, D.M.; Sheff, K.; Keim, P.S.; Nguyen, J.; Drusano, G.L. In vitro system for modeling influenza a virus resistance under drug pressure. Antimicrob. Agents Chemother. 2010, 54, 3442–3450. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.N.; Bulitta, J.B.; McSharry, J.J.; Weng, Q.; Adams, J.R.; Kulawy, R.; Drusano, G.L. Effect of half-life on the pharmacodynamic index of zanamivir against influenza virus delineated by a mathematical model. Antimicrob. Agents Chemother. 2011, 55, 1747–1753. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.N.; McSharry, J.J.; Weng, Q.; Adams, J.R.; Kulawy, R.; Drusano, G.L. Zanamivir, at 600 milligrams twice daily, inhibits oseltamivir-resistant 2009 pandemic H1N1 influenza virus in an in vitro hollow-fiber infection model system. Antimicrob. Agents Chemother. 2011, 55, 1740–1746. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.N.; Adams, J.R.; Baluya, D.L.; Drusano, G.L. Pharmacokinetic determinants of virological response to raltegravir in the in vitro pharmacodynamic hollow-fiber infection model system. Antimicrob. Agents Chemother. 2015, 59, 3771–3777. [Google Scholar] [CrossRef] [PubMed]
- Gutterman, J.U.; Fine, S.; Quesada, J.; Horning, S.J.; Levine, J.F.; Alexanian, R.; Bernhardt, L.; Kramer, M.; Spiegel, H.; Colburn, W.; et al. Recombinant leukocyte A interferon: Pharmacokinetics, single-dose tolerance, and biologic effects in cancer patients. Ann. Intern. Med. 1982, 96, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Preston, S.L.; Drusano, G.L.; Glue, P.; Nash, J.; Gupta, S.K.; McNamara, P. Pharmacokinetics and absolute bioavailability of ribavirin in healthy volunteers as determined by stable-isotope methodology. Antimicrob. Agents Chemother. 1999, 43, 2451–2456. [Google Scholar] [PubMed]
- World Health Organization. Dengue and Severe Dengue; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Whitehead, S.S.; Blaney, J.E.; Durbin, A.P.; Murphy, B.R. Prospects for a dengue virus vaccine. Nat. Rev. Microbiol. 2007, 5, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Zachariah, M.; Harris, E. Mycophenolic acid inhibits dengue virus infection by preventing replication of viral RNA. Virology 2002, 304, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Koff, W.C.; Elm, J.L., Jr.; Halstead, S.B. Antiviral effects if ribavirin and 6-mercapto-9-tetrahydro-2-furylpurine against dengue viruses in vitro. Antivir. Res. 1982, 2, 69–79. [Google Scholar] [CrossRef]
- Diamond, M.S.; Roberts, T.G.; Edgil, D.; Lu, B.; Ernst, J.; Harris, E. Modulation of Dengue virus infection in human cells by alpha, beta, and gamma interferons. J. Virol. 2000, 74, 4957–4966. [Google Scholar] [CrossRef] [PubMed]
- Emeny, J.M.; Morgan, M.J. Regulation of the interferon system: Evidence that Vero cells have a genetic defect in interferon production. J. Gen. Virol. 1979, 43, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Liu, W.J.; Prow, N.A.; Geertsema, C.; Ligtenberg, M.; Vanlandingham, D.L.; Schnettler, E.; Vlak, J.M.; Suhrbier, A.; Khromykh, A.A.; et al. Chikungunya virus nonstructural protein 2 inhibits type I/II interferon-stimulated JAK-STAT signaling. J. Virol. 2010, 84, 10877–10887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The genome landscape of the African green monkey kidney-derived vero cell line. DNA Res. 2014, 21, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Fried, M.W.; Shiffman, M.L.; Reddy, K.R.; Smith, C.; Marinos, G.; Goncales, F.L., Jr.; Haussinger, D.; Diago, M.; Carosi, G.; Dhumeaux, D.; et al. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N. Engl. J. Med. 2002, 347, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Fernando, S.; Wijewickrama, A.; Gomes, L.; Punchihewa, C.T.; Madusanka, S.D.; Dissanayake, H.; Jeewandara, C.; Peiris, H.; Ogg, G.S.; Malavige, G.N. Patterns and causes of liver involvement in acute dengue infection. BMC Infect. Dis. 2016, 16, 319. [Google Scholar] [CrossRef] [PubMed]
- Khongphatthanayothin, A.; Mahayosnond, A.; Poovorawan, Y. Possible cause of liver failure in patient with dengue shock syndrome. Emerg. Infect. Dis. 2013, 19, 1161–1163. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Time (day) | IFN Cavg (IU/mL) a | RBV Cavg (mg/L) b |
|---|---|---|
| 1 | 182.10 | 0.394 |
| 2 | 250.78 | 0.616 |
| 3 | 259.88 | 0.76 |
| Vero a | Huh-7 a | |||||
|---|---|---|---|---|---|---|
| Parameter | RBV | IFN | RBV | IFN | Units | |
| Antiviral Effect | r2 b | 0.999 | 0.986 | 0.915 | 0.973 | -- |
| Econc | 6.46 | 6.61 | 6.82 | 7.04 | Log10 PFU/mL | |
| EC50d | 106.6 | 1381 | 10.02 | 58.34 | mg/L or IU/mL | |
| He | 0.934 | 0.356 | 14.44 | 0.315 | -- | |
| Cytotoxicity | r2 b | 0.995 | --f | 0.958 | --f | -- |
| Econc | 7.53 | 7.54 | 7.47 | 7.40 | Log10 RLU | |
| CC50d | 137.4 | >10,000 | 18.0 | >10,000 | mg/L or IU/mL | |
| He | 1.87 | --f | 1.76 | --f | ||
| Parameter | Final Estimate | Units | |
|---|---|---|---|
| Vero | Huh-7 | ||
| r2 a | 0.928 | 0.924 | -- |
| Econb | 6.072 | 6.73 | Log10 PFU/mL |
| EC50 RBVc | 69.41 | 6.06 | mg/L |
| m1d | 1 | 1 | -- |
| EC50 IFNe | >10,000 | 110.2 | IU/mL |
| m2f | 1 | 1 | -- |
| αg | 22.86 [−11.3–57.03] h | 34.87 [−30.54–100.3] h | -- |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pires de Mello, C.P.; Drusano, G.L.; Rodriquez, J.L.; Kaushik, A.; Brown, A.N. Antiviral Effects of Clinically-Relevant Interferon-α and Ribavirin Regimens against Dengue Virus in the Hollow Fiber Infection Model (HFIM). Viruses 2018, 10, 317. https://doi.org/10.3390/v10060317
Pires de Mello CP, Drusano GL, Rodriquez JL, Kaushik A, Brown AN. Antiviral Effects of Clinically-Relevant Interferon-α and Ribavirin Regimens against Dengue Virus in the Hollow Fiber Infection Model (HFIM). Viruses. 2018; 10(6):317. https://doi.org/10.3390/v10060317
Chicago/Turabian StylePires de Mello, Camilly P., George L. Drusano, Jaime L. Rodriquez, Ajeet Kaushik, and Ashley N. Brown. 2018. "Antiviral Effects of Clinically-Relevant Interferon-α and Ribavirin Regimens against Dengue Virus in the Hollow Fiber Infection Model (HFIM)" Viruses 10, no. 6: 317. https://doi.org/10.3390/v10060317