Conserved Active-Site Residues Associated with OAS Enzyme Activity and Ubiquitin-Like Domains Are Not Required for the Antiviral Activity of goOASL Protein against Avian Tembusu Virus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. RNA Extraction and cDNA Preparation
2.3. Western Blotting Assay
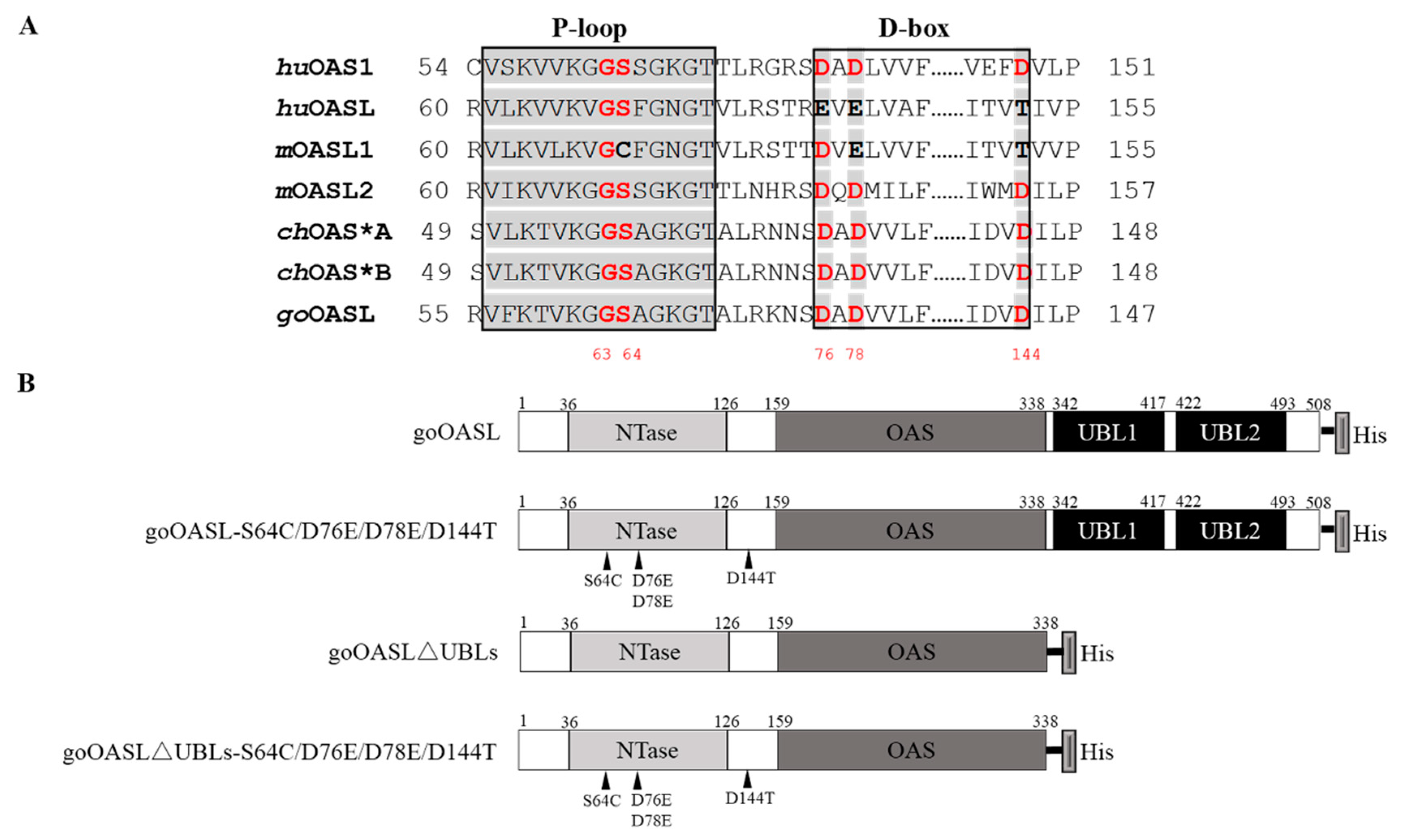
2.4. The Construction of goOASL-Mutant Plasmids
2.5. The Cytotoxicity Assay of goOASL-Mutant Proteins
2.6. Antiviral Activity Assay of goOASL and Its Mutant Proteins
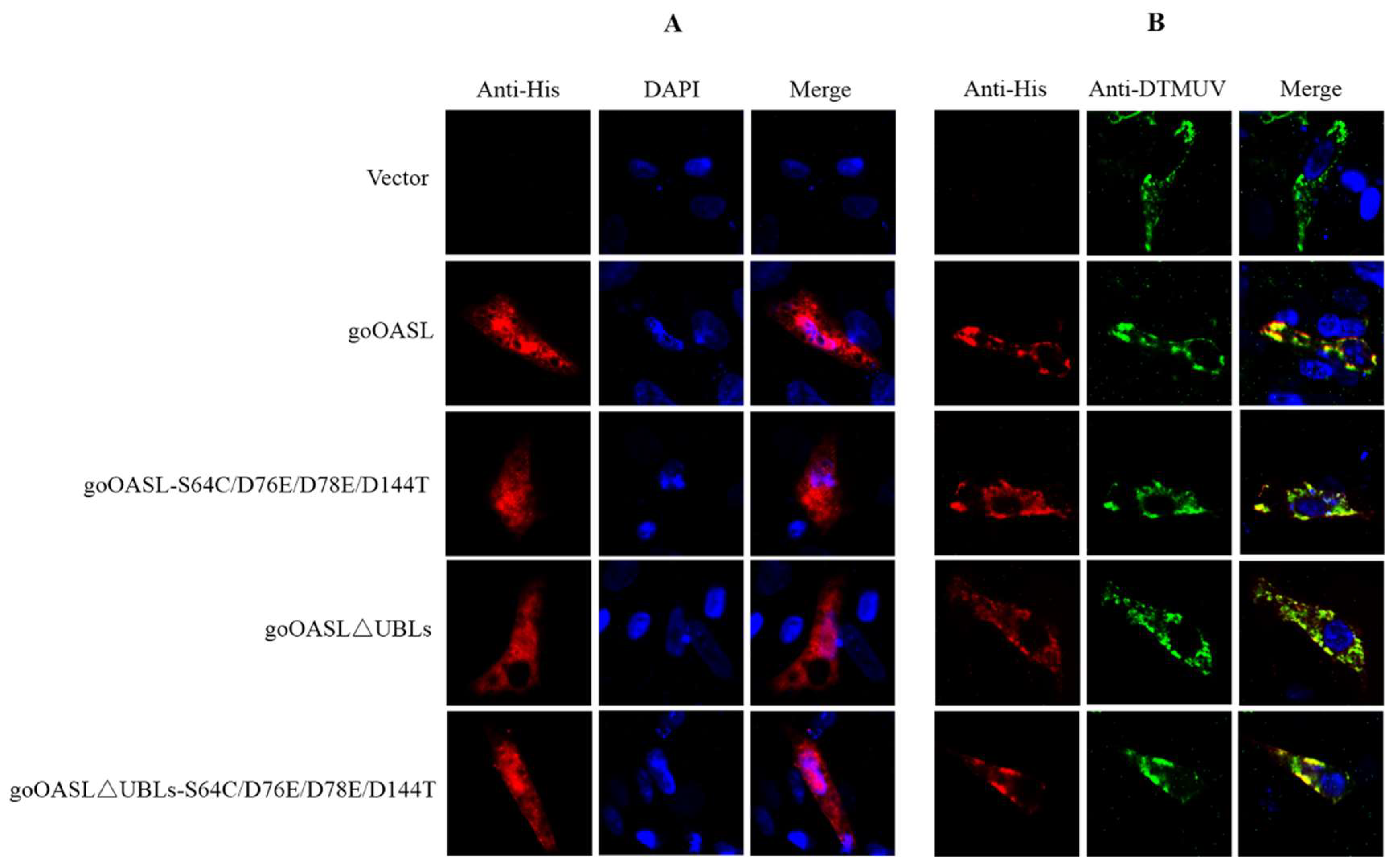
2.7. The Cellular Colocation of goOASL and Its Mutant Proteins with DTMUV
3. Results
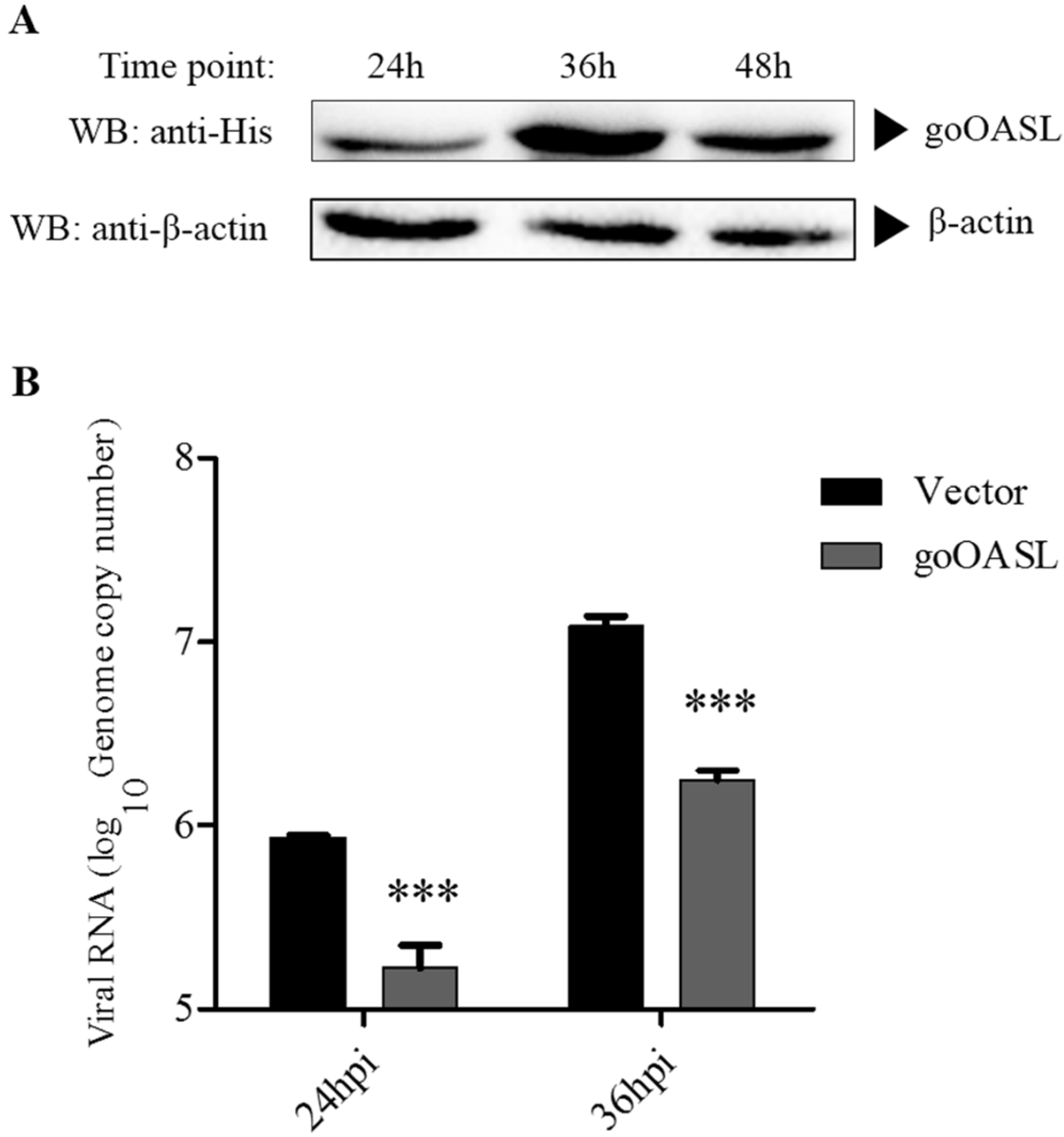
3.1. The Antiviral Activity of goOASL Protein against DTMUV in DEFs
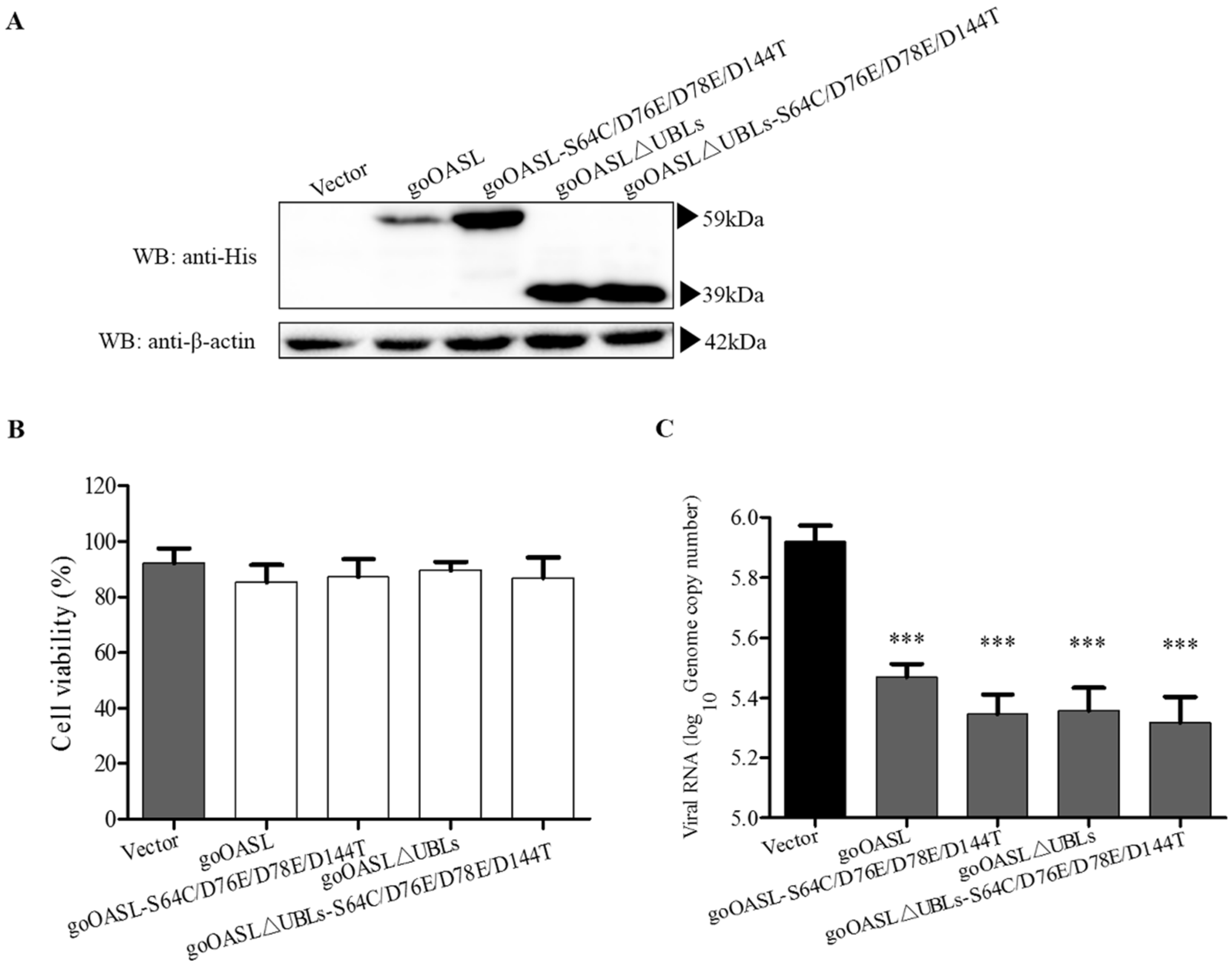
3.2. The Antiviral Activity of goOASL-Mutant Proteins against DTMUV in DEFs
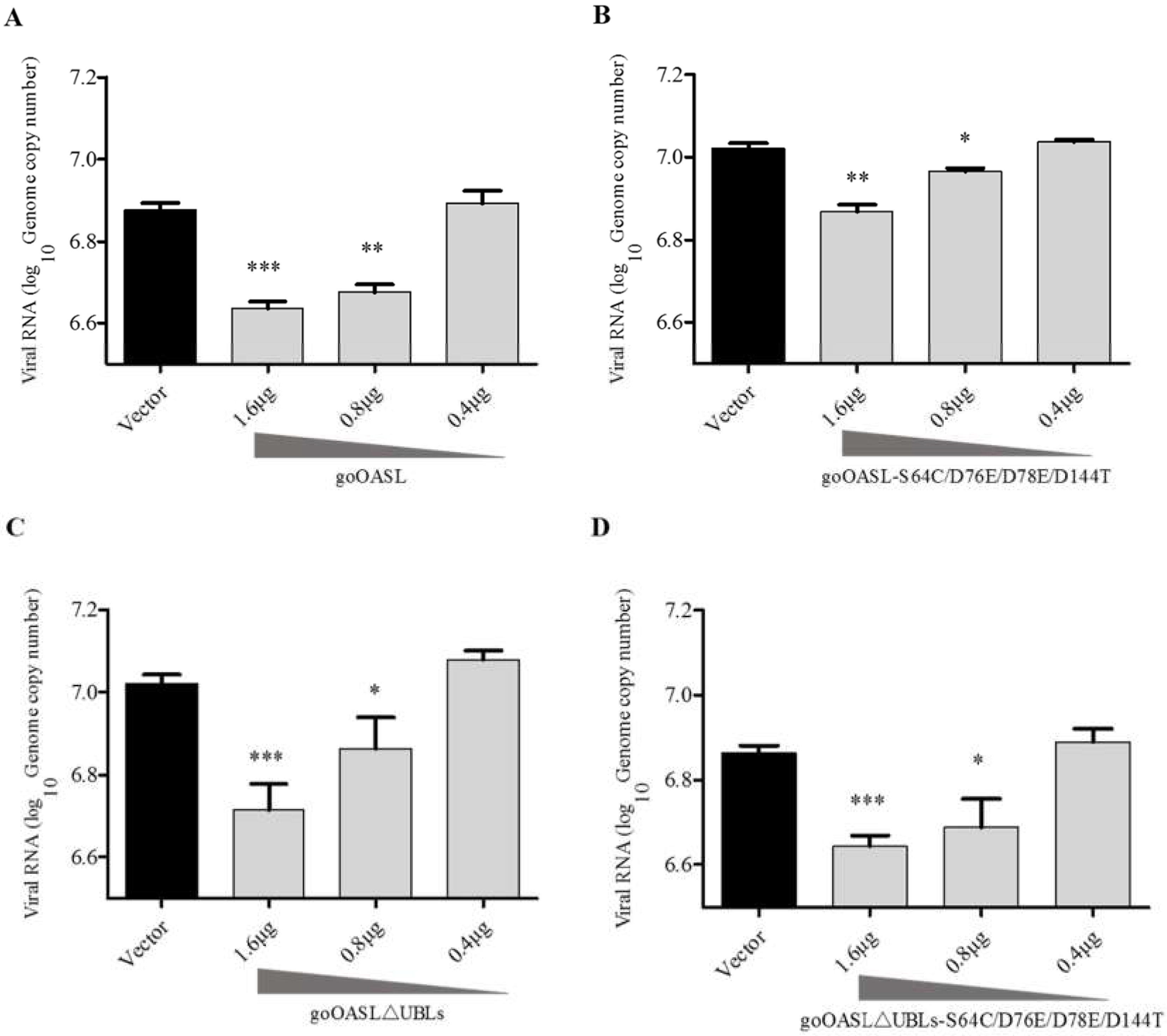
3.3. GoOASL and Its Mutant Proteins Exhibited Antiviral Activity against DTMUV in DEFs in A Dose-Dependent Manner
3.4. Cellular Colocalization of goOASL and Its Mutant Proteins with DTMUV in DEFs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pichlmair, A.; e Sousa, C.R. Innate recognition of viruses. Immunity 2007, 27, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Sander, C. DNA polymerase beta belongs to an ancient nucleotidyltransferase superfamily. Trends Biochem. Sci. 1995, 20, 345–347. [Google Scholar] [CrossRef]
- Zhu, J.; Ghosh, A.; Sarkar, S.N. OASL—A new player in controlling antiviral innate immunity. Curr. Opin. Virol. 2015, 12, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Kakuta, S.; Shibata, S.; Iwakura, Y. Genomic structure of the mouse 2′,5′-oligoadenylate synthetase gene family. J. Interferon Cytokine Res. 2002, 22, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Eskildsen, S.; Justesen, J.; Schierup, M.H.; Hartmann, R. Characterization of the 2′-5′-oligoadenylate synthetase ubiquitin-like family. Nucleic Acids Res. 2003, 31, 3166–3173. [Google Scholar] [CrossRef] [PubMed]
- Kjaer, K.H.; Poulsen, J.B.; Reintamm, T.; Saby, E.; Martensen, P.M.; Kelve, M.; Justesen, J. Evolution of the 2′,5′-oligoadenylate synthetase family in eukaryotes and bacteria. J. Mol. Evol. 2009, 69, 612–624. [Google Scholar] [CrossRef] [PubMed]
- Justesen, J.; Hartmann, R.; Kjeldgaard, N.O. Gene structure and function of the 2′-5′-oligoadenylate synthetase family. Cell. Mol. Life Sci. 2000, 57, 1593–1612. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Silverman, R.H. 2-5A-dependent RNase molecules dimerize during activation by 2-5A. J. Biol. Chem. 1995, 270, 4133–4137. [Google Scholar] [CrossRef] [PubMed]
- Kuchta, K.; Knizewski, L.; Wyrwicz, L.S.; Rychlewski, L.; Ginalski, K. Comprehensive classification of nucleotidyltransferase fold proteins: Identification of novel families and their representatives in human. Nucleic Acids Res. 2009, 37, 7701–7714. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wu, J.; Du, F.; Chen, X.; Chen, Z.J. Cyclic GMP-AMP Synthase is a cytosolic DNA sensor that activates the type-I interferon pathway. Science 2013, 339, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Torralba, S.; Sojat, J. Hartmann R. 2′-5′ Oligoadenylate synthetase shares active site architecture with the archaeal CCA-adding enzyme. Cell Mol. Life Sci. 2008, 43, 2613–2620. [Google Scholar] [CrossRef] [PubMed]
- Dhar, J.; Cuevas, R.A.; Goswami, R.; Zhu, J.; Sarkar, S.N.; Barik, S. 2′-5′-Oligoadenylate Synthetase-Like Protein Inhibits Respiratory Syncytial Virus Replication and Is Targeted by the Viral Nonstructural Protein 1. J. Virol. 2015, 89, 10115–10119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, R.; Olsen, H.S.; Widder, S.; Jorgensen, R.; Justesen, J. P59OASL, a 2′-5′ oligoadenylate synthetase like protein: A novel human gene related to the 2′-5′ oligoadenylate synthetase family. Nucleic Acids Res. 1998, 26, 4121–4128. [Google Scholar] [CrossRef] [PubMed]
- Rebouillat, D.; Marié, I.; Hovanessian, A.G. Molecular cloning and characterization of two related and interferon-induced 56-kDa and 30-kDa proteins highly similar to 2′-5′ oligoadenylate synthetase. Eur. J. Biochem. 1998, 257, 319–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbahesh, H.; Jha, B.K.; Silverman, R.H.; Scherbik, S.V.; Brinton, M.A. The FLVR-encoded murine oligoadenylate synthetase 1B (OAS1B) suppresses 2-5A synthesis in intact cells. Virology 2011, 409, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Iwata, A.; Koh, Y.; Kawai, S.; Murayama, S.; Hamada, K.; Maekawa, S.; Ueda, S.; Sokawa, Y. Two types of chicken 2′,5′-oligoadenylate synthetase mRNA derived from alleles at a single locus 1. Biochim. Biophys. Acta 1998, 1395, 181–191. [Google Scholar] [CrossRef]
- Zhou, A.; Paranjape, J.M.; Hassel, B.A.; Nie, H.; Shah, S.; Galinski, B.; Silverman, R.H. Impact of RNase L overexpression on viral and cellular growth and death. J. Interferon Cytokine Res. 1998, 18, 953–961. [Google Scholar] [PubMed]
- Scherbik, S.V.; Paranjape, J.M.; Stockman, B.M.; Silverman, R.H.; Brinton, M.A. RNase L Plays a Role in the Antiviral Response to West Nile Virus. J. Virol. 2006, 80, 2987–2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Zhang, Y.; Ghosh, A.; Cuevas, R.; Forero, A.; Dhar, J.; Ibsen, M.; Schmid-Burgk, J.L.; Schmidt, T.; Ganapathiraju, M. Antiviral Activity of Human OASL Protein Is Mediated by Enhancing Signaling of the RIG-I RNA Sensor. Immunity 2014, 40, 936–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibsen, M.S.; Gad, H.H.; Andersen, L.L.; Hornung, V.; Julkunen, I.; Sarkar, S.N.; Hartmann, R. Structural and functional analysis reveals that human OASL binds dsRNA to enhance RIG-I signaling. Nucleic Acids Res. 2015, 43, 5236–5248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashimo, T.; Simon-Chazottes, D.; Guénet, J.L. Innate Resistance to Flavivirus Infections and the Functions of 2′-5′ Oligoadenylate Synthetases. Curr. Top. Microbiol. Immunol. 2008, 321, 85–100. [Google Scholar] [PubMed]
- Lin, R.J.; Yu, H.P.; Chang, B.L.; Tang, W.C.; Liao, C.L.; Lin, Y.L. Distinct antiviral roles for human 2′,5′-oligoadenylate synthetase family members against dengue virus infection. J. Immunol. 2009, 183, 8035–8043. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zhu, D.; Lian, X.; Liu, W.; Cao, R.; Chen, P. Porcine 2′,5′-oligoadenylate synthetases inhibit Japanese encephalitis virus replication in vitro. J. Med. Virol. 2015, 88, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Courtney, S.C.; Di, H.; Stockman, B.M.; Liu, H.; Scherbik, S.V.; Brinton, M.A. Identification of novel host cell binding partners of OAS1B, the protein conferring resistance to Flavivirus-induced disease in mice. J. Virol. 2012, 86, 7953–7963. [Google Scholar] [CrossRef] [PubMed]
- Tag-El-Din-Hassan, H.T.; Sasaki, N.; Torigoe, D.; Morimatsu, M.; Agui, T. Analysis of the relationship between enzymatic and antiviral activities of the chicken oligoadenylate synthetase-like. J. Interferon Cytokine Res. 2016, 37, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liu, F.; Chen, S.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Sun, K.; Yang, Q.; Wu, Y. Identification of 2′-5′-oligoadenylate synthetase-like gene in goose: gene structure, expression patterns and antiviral activity against newcastle disease virus. J. Interferon Cytokine Res. 2016, 36, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhang, W.; Wu, Z.; Zhang, J.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Sun, K.; Yang, Q.; et al. Goose MX and OASL play vital roles in the antiviral effects of type I, II and III interferon against newly emerging avian Flavivirus. Front. Immunol. 2017, 8, 1006. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zhang, C.; Liu, Y.; Liu, Y.; Ye, W.; Han, J.; Ma, G.; Zhang, D.; Xu, F.; Gao, X. Tembusu virus in ducks, China. Emerg. Infect. Dis. 2011, 17, 1873–1875. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Zhao, Y.; Zhang, X.; Xu, D.; Dai, X.; Teng, Q.; Yan, L.; Zhou, J.; Ji, X.; Zhang, S. An infectious disease of ducks caused by a newly emerged Tembusu virus strain in mainland China. Virology 2011, 417, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chen, S.; Chen, Y.; Liu, C.; Chen, S.; Yin, X.; Li, G.; Zhang, Y. Adapted Tembusu-like virus in chickens and geese in China. J. Clin. Microbiol. 2012, 50, 2807–2809. [Google Scholar] [CrossRef] [PubMed]
- Yun, T.; Zhang, A.D.; Ma, B.X.; Cao, C.Z.; Chen, B.L.; Ni, A.Z.; Ye, A.W.; Yu, A.B.; Hua, A.J.; Zhang, A.Y. Complete genome sequence of a novel Flavivirus, duck Tembusu virus, isolated from ducks and geese in China. J. Virol. 2012, 86, 3406–3407. [Google Scholar] [CrossRef] [PubMed]
- Ti, J.; Zhang, L.; Li, Z.; Zhao, D.; Zhang, Y.; Li, F.; Diao, Y. Effect of age and inoculation route on the infection of duck Tembusu virus in Goslings. Vet. Microbiol. 2015, 181, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, S.; Mahalingam, S.; Wang, M.; Cheng, A. An updated review of avian-origin Tembusu virus: A newly emerging avian Flavivirus. J. Gen. Virol. 2017, 98, 2413–2420. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Huang, J.; Jia, R.; Zhang, B.; Wang, M.; Zhu, D.; Chen, S.; Liu, M.; Yin, Z.; Cheng, A. Identification and molecular characterization of a novel duck Tembusu virus isolate from Southwest China. Arch. Virol. 2015, 160, 2781–2790. [Google Scholar] [CrossRef] [PubMed]
- Tag-El-Din-Hassan, H.T.; Morimatsu, M.; Agui, T. Function analysis of duck, goose and ostrich 2′-5′-oligoadenylate synthetase. Infect. Genet. Evol. 2018, 62, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Li, L.F.; Yu, J.; Zhang, Y.; Yang, Q.; Li, Y.; Zhang, L.; Wang, J.; Li, S.; Luo, Y.; Sun, Y. Interferon-inducible oligoadenylate synthetase-like protein acts as an antiviral effector against classical swine fever virus via the MDA5-mediated type I interferon signaling pathway. J. Virol. 2017, 91, e01514-16. [Google Scholar] [CrossRef] [PubMed]
- Bi, K.R.; Han, K.K.; Liu, Q.T.; Zhao, D.M.; Huang, X.M.; Liu, Y.Z.; Yang, J.; Li, Y. Molecular cloning, characterization and expression of duck 2′-5′-oligoadenylate synthetase-like gene. Gene 2017, 629, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, M.; Wakita, T.; Esumi, M. 2′,5′-Oligoadenylate synthetase-like gene highly induced by hepatitis C virus infection in human liver is inhibitory to viral replication in vitro. Biochem. Biophys. Res. Commun. 2010, 392, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.D. New insights into Flavivirus nonstructural protein 5. Adv. Virus Res. 2009, 74, 41–101. [Google Scholar] [PubMed]
- Diamond, M.S. Evasion of innate and adaptive immunity by Flaviviruses. Immunol. Cell Biol. 2003, 81, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.; Holden, K.L.; Edgil, D.; Polacek, C.; Clyde, K. Molecular biology of Flaviviruses. Novartis Found. Symp. 2006, 277, 23–39. [Google Scholar] [PubMed]
- Taguchi, T.; Naganofujii, M.; Akutsu, M.; Kadoya, H.; Ohgimoto, S.; Ishido, S.; Hotta, H. Hepatitis C virus NS5A protein interacts with 2′,5′-oligoadenylate synthetase and inhibits antiviral activity of IFN in an IFN sensitivity-determining region-independent manner. J. Gen. Virol. 2004, 85, 959–969. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′-3′) | Application |
|---|---|---|
| goOASL-N1-F goOASL-N1-R | CTGCGGGAGCCGCGATGGAG GGGCATCGTAGGTGGGCAGGA | Amplification of the N-terminus of goOASL-S64C/D76E/D78E/D144T |
| goOASL-C2-F goOASL-C2-R | TCCTGCCCACCTACGATGCCC CCAGGGAGAAATAAAAGGGGATG | Amplification of the rest of the sequence of goOASL-S64C/D76E/D78E/D144T |
| goOASL-His-F goOASL-His-R | TGGTGGAATTCTGCAGATATCGCCACCATGGAGCTGCGGGACGTG GCCCTCTAGACTCGAGCGGCCGCTCAGTGGTGGTGGTGGTGGTGGGAGGGCTGGCAGCAAGG | Amplification of goOASL-S64C/D76E/D78E/D144T and goOASL-D76A/D78A |
| goOASL∆UBLs-His-F goOASL∆UBLs-His-R | TGGTGGAATTCTGCAGATATCGCCACCATGGAGCTGCGGGACGTG GCCCTCTAGACTCGAGCGGCCGCTCAGTGGTGGTGGTGGTGGTGCGGCTGCACGTTCCAGGGGT | Amplification of goOASL∆UBLs, goOASL∆UBLs-S64C/D76E/D78E/D144T and goOASL∆UBLs-D76A/D78A |
| goOASL-N1′-F goOASL-N1′-R | CTGCGGGAGCCGCGATGGAG CCACGGCGGCGGCCGAGTTCT | Amplification of the N-terminus of goOASL-D76A/D78A |
| goOASL-C2′-F goOASL-C2′-R | AGAACTCGGCCGCCGCCGTGG CCAGGGAGAAATAAAAGGGGATG | Amplification of the rest of the sequence of goOASL-D76A/D78A |
| DTMUV(E)-F DTMUV(E)-R | CGCTGAGATGGAGGATTATGG ACTGATTGTTTGGTGGCGTG | qRT-PCR for detection of viral RNA of DTMUV |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Yang, C.; Zhang, J.; Wu, Z.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Yang, Q.; Wu, Y.; et al. Conserved Active-Site Residues Associated with OAS Enzyme Activity and Ubiquitin-Like Domains Are Not Required for the Antiviral Activity of goOASL Protein against Avian Tembusu Virus. Viruses 2018, 10, 371. https://doi.org/10.3390/v10070371
Chen S, Yang C, Zhang J, Wu Z, Wang M, Jia R, Zhu D, Liu M, Yang Q, Wu Y, et al. Conserved Active-Site Residues Associated with OAS Enzyme Activity and Ubiquitin-Like Domains Are Not Required for the Antiviral Activity of goOASL Protein against Avian Tembusu Virus. Viruses. 2018; 10(7):371. https://doi.org/10.3390/v10070371
Chicago/Turabian StyleChen, Shun, Chao Yang, Jinyue Zhang, Zhen Wu, Mingshu Wang, Renyong Jia, Dekang Zhu, Mafeng Liu, Qiao Yang, Ying Wu, and et al. 2018. "Conserved Active-Site Residues Associated with OAS Enzyme Activity and Ubiquitin-Like Domains Are Not Required for the Antiviral Activity of goOASL Protein against Avian Tembusu Virus" Viruses 10, no. 7: 371. https://doi.org/10.3390/v10070371