Pathogenesis of Noroviruses, Emerging RNA Viruses

Center for Molecular and Tumor Virology, Department of Microbiology and Immunology, Louisiana State University Health Sciences Center, Shreveport, Louisiana 71130-3932, USA
Viruses 2010, 2(3), 748-781; https://doi.org/10.3390/v2030748
Submission received: 18 November 2009
/
Revised: 15 March 2010
/
Accepted: 15 March 2010
/
Published: 23 March 2010
(This article belongs to the Special Issue Pathogenesis of Emerging and Re-Emerging RNA Viruses)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Human noroviruses in the family Caliciviridae are a major cause of epidemic gastroenteritis. They are responsible for at least 95% of viral outbreaks and over 50% of all outbreaks worldwide. Transmission of these highly infectious plus-stranded RNA viruses occurs primarily through contaminated food or water, but also through person-to-person contact and exposure to fomites. Norovirus infections are typically acute and self-limited. However, disease can be much more severe and prolonged in infants, elderly, and immunocompromised individuals. Norovirus outbreaks frequently occur in semi-closed communities such as nursing homes, military settings, schools, hospitals, cruise ships, and disaster relief situations. Noroviruses are classified as Category B biodefense agents because they are highly contagious, extremely stable in the environment, resistant to common disinfectants, and associated with debilitating illness. The number of reported norovirus outbreaks has risen sharply since 2002 suggesting the emergence of more infectious strains. There has also been increased recognition that noroviruses are important causes of childhood hospitalization. Moreover, noroviruses have recently been associated with multiple clinical outcomes other than gastroenteritis. It is unclear whether these new observations are due to improved norovirus diagnostics or to the emergence of more virulent norovirus strains. Regardless, it is clear that human noroviruses cause considerable morbidity worldwide, have significant economic impact, and are clinically important emerging pathogens. Despite the impact of human norovirus-induced disease and the potential for emergence of highly virulent strains, the pathogenic features of infection are not well understood due to the lack of a cell culture system and previous lack of animal models. This review summarizes the current understanding of norovirus pathogenesis from the histological to the molecular level, including contributions from new model systems.
1. Introduction
Noroviruses constitute a genus within the family Caliciviridae. The human pathogens within this genus cause at least 95% of nonbacterial gastroenteritis outbreaks, and 50% of all gastroenteritis outbreaks, throughout the world. It is estimated that there are approximately 23 million norovirus infections per year in the United States alone, causing 50,000 hospitalizations and 300 deaths [1]. Based on a recent review of diagnostic studies testing for norovirus infections in clinical settings, it is estimated that they cause over one million hospitalizations and 200,000 deaths in young children in developing countries annually [2]. Noroviruses are highly transmissible and can spread via exposure to contaminated food or water sources, person-to-person contact, aerosolized vomitus particles, and fomites (Figure 1A). They are considered by CDC to be the most common cause of foodborne disease outbreaks [3]. Numerous features of norovirus infections have led to their classification as Category B biodefense agents, including their high infectivity, extreme stability, resistance to common disinfectants, and ability to cause incapacitating disease. Persons of all age groups are susceptible to infection and secondary infections are common [4,5,6]. In recent years, there has been a dramatic increase in the number of reported norovirus outbreaks. These emerging RNA viruses have historically been very difficult to study but numerous advances in the field have begun to contribute to the overall understanding of norovirus pathogenesis.
2. Clinical Disease and Epidemiology
2.1. Clinical Disease
In immunocompetent adults, the course of norovirus infection is rapid, with an incubation period of 24–48 hours and resolution of symptoms within 12–72 hours [7]. Symptoms include vomiting and diarrhea with or without nausea and abdominal cramps. Low-grade fever and malaise can also develop. While norovirus infection typically causes an acute bout of gastroenteritis that resolves within days of the onset of symptoms, norovirus-induced disease can be much more severe and prolonged in specific risk groups. For example, infants and young children can develop more severe gastroenteritis following norovirus infection, with symptoms lasting up to six weeks [8,9,10,11]. Noroviruses have been reported to be second only to rotaviruses in causing severe childhood gastroenteritis [10,12]; considering the recent success in vaccinating infants against rotavirus infections, noroviruses will likely become the most common cause of childhood diarrheal disease in the foreseeable future. In addition, prolonged symptomatic infection has been well-documented in transplant patients and other immunosuppressed individuals, with symptoms lasting over two years [13,14,15,16,17,18,19]. Similarly, norovirus infection can be particularly severe in the elderly, even resulting in death [20,21,22]. One recent study also reported that patients with inflammatory bowel disease may present with bloody diarrhea upon norovirus infection [24].
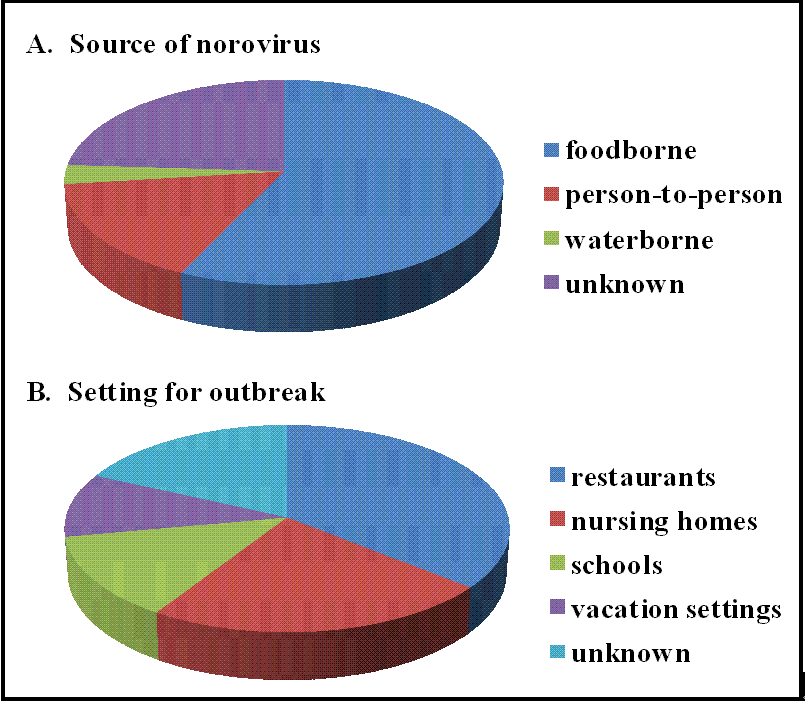
Figure 1.
Norovirus outbreak characteristics.The Centers for Disease Control collected data from 232 norovirus outbreaks between July 1997 and June 2000. The results of this surveillance are summarized here for the source of virus (A)and the type of setting affected by the outbreak (B). Data are reproduced from http://www.cdc.gov/ncidod/dvrd/revb/gastro/norovirus-factsheet.htm.
Figure 1.
Norovirus outbreak characteristics.The Centers for Disease Control collected data from 232 norovirus outbreaks between July 1997 and June 2000. The results of this surveillance are summarized here for the source of virus (A)and the type of setting affected by the outbreak (B). Data are reproduced from http://www.cdc.gov/ncidod/dvrd/revb/gastro/norovirus-factsheet.htm.

The complete scope of norovirus-induced morbidity worldwide and particularly in developing nations has been difficult to ascertain due to several factors (discussed in more detail elsewhere [20,25]). First, norovirus detection is difficult because these viruses cannot be propagated in cell culture and they are genetically variable, complicating RT-PCR-based detection assays. Second, there is a lack of reporting to health officials because of the acute nature of disease. Finally, national and international diagnostic/surveillance programs are not standardized, if present at all. Importantly though, there has recently been increased recognition of the burden of norovirus disease due to significant improvements in norovirus diagnostic assays and an increased awareness of the need for surveillance standardization. Along these lines, a global surveillance network called NoroNet (http://www.noronet.nl/noronet/) was recently established that aims to monitor norovirus epidemics in an effort to limit their scope. Continued efforts of this sort will be critical to delineate the global burden of norovirus disease.
As norovirus diagnostics have improved, there have been numerous reports of norovirus associations with clinical outcomes other than gastroenteritis. For example, one case report detected norovirus RNA in the serum and cerebrospinal fluid of a child with encephalopathy [26]. In addition, during a norovirus outbreak among military personnel in Afghanistan in May 2002, three infected patients presented with gastroenteritis, diminished alertness, headache, neck stiffness, and light sensitivity; one of these patients also displayed disseminated intravascular coagulation [27]. Noroviruses have also been implicated in necrotizing enterocolitis in premature infants [28], postinfectious irritable bowel syndrome [29], and benign infantile seizures [30,31]. While these case reports provide only anecdotal evidence that norovirus infection can have varied clinical outcomes, they do suggest that noroviruses should be considered as potential etiological agents of diseases other than gastroenteritis.
2.2. Genetic Diversity
Noroviruses are classified in five genogroups (GI-GV), three of which (GI, GII, and GIV) contain primarily human viruses [32,33]. Multiple porcine noroviruses have been placed in GII [34,35]. Genogroup III (GIII) contains bovine noroviruses [36,37] and GV contains murine noroviruses [38,39,40,41,42]. The genogroups are further subdivided into 31 distinct clusters or genotypes (8 GI, 19 GII, 2 GIII, 1 GIV, and 1 GV genotypes) [4,32]. Norovirus strains are typically named after the location in which they were first identified; e.g., the Farmington Hills strain was first shown to be responsible for gastroenteritis cases in Farmington Hills, Michigan [43]. They are also commonly referred to by their genogroup and genotype; e.g., the Farmington Hills strain is a GII.4 (genogroup II, genotype 4) strain. Noroviruses display a wide degree of genetic variability – members within a genogroup differ by 45–61% in their capsid genes, members within a genotype differ by 14–44%, and strains within a genotype differ by 0–14% [32]. This amount of intra-genus variation is high even compared to genera of other plus-strand RNA virus families [32]. This high degree of variability is undoubtedly one factor that complicates protective norovirus immunity.
2.3. Epidemiology
Norovirus outbreaks occur most commonly in semi-closed communities such as nursing homes, schools, hospitals, cruise ships, disaster relief/evacuation sites, and military settings (Figure 1B) [33,44,45,46,47]. Nosocomial norovirus outbreaks are one of the most common causes of hospital ward closures [48]. Outbreaks display wintertime seasonality, explaining why norovirus disease is referred to as the winter vomiting disease [49]. Several factors most likely contribute to the explosive nature of norovirus outbreaks, including the high infectivity of norovirus particles [50], the persistence of noroviruses in the environment [51,52,53], prolonged shedding of virus from both symptomatic and asymptomatic individuals [54,55,56], and a lack of lasting immunity [4,5,6]. In 2002, a sharp rise in norovirus outbreaks worldwide was recognized which correlated with the emergence of a new virus variant, specifically a GII.4 norovirus strain [51,57]. Interestingly, researchers have determined that GII.4 strains resulted in pandemics in 1995–1996 (US95/96 strain) [33,58,59], 2002 (Farmington Hills strain) [43], 2004 (Hunter strain) [60], and 2006 (Minerva/2006b and Laurens/2006a strains) [61]. These GII.4 variants spread rapidly and globally and are thought to account for 70–80% of all norovirus outbreaks at least since 2002 [62]. A recent study demonstrates that GII.4 strains associated with severe illness were circulating as early as 1974, and that the ancestral strain most likely emerged in the 1960s [63]. It is unclear why the GII.4 norovirus strains are so predominant but possibilities include increased environmental stability, transmissibility, and virulence [64].
While it is widely accepted that noroviruses cause a majority of epidemic gastroenteritis worldwide, they have not received significant attention as a cause of sporadic gastroenteritis. However, improved diagnostics has shed light on the unappreciated breadth of norovirus infection in this setting [12]. Based on numerous studies around the world, it is now recognized that noroviruses are an important cause of illness in hospitalized children (discussed in detail in several recent reviews [12,25,65]), second to or as prevalent as rotavirus infections. They are also a previously unappreciated cause of traveler’s diarrhea [66,67,68].
3. Pathophysiology
Because of the lack of a cell culture system and the historical lack of animal models of norovirus infection, the extent of our knowledge regarding pathogenesis of norovirus infection comes primarily from physical, histological, and biochemical studies of infected human volunteers. In recent years, work in porcine, bovine, and murine models has also begun to contribute to our understanding of norovirus pathogenesis.
3.1. Histological Alterations in the Intestine
Histological analysis of proximal intestinal biopsy samples from human volunteers that become ill after administration of either a GI (Norwalk; GI.1) or GII (Hawaii; GII.1) norovirus demonstrate an intact intestinal mucosa with specific histological changes, including broadening and blunting of the villi, shortening of the microvilli, enlarged and pale mitochondria, increased cytoplasmic vacuolization, and intercellular edema [69,70,71,72]. While abnormalities are apparent in intestinal epithelial cells of norovirus-infected volunteers, electron microscopy analysis reveals that these cells remain intact [69,70]. Crypt cell hyperplasia has also been reported following norovirus infection [71,73]. Only proximal intestinal biopsies from infected individuals were obtained in early volunteer studies so it remains to be determined whether the distal intestine is also affected by norovirus infection. In addition to enterocyte changes, norovirus infection results in a mild inflammatory infiltration into the lamina propria that has been observed in humans infected with the Norwalk [69,71] and Hawaii viruses [70,72], gnotobiotic calves infected with the human GII.4 norovirus HS66 [74], and mice infected with murine norovirus 1 (MNV-1) [75]. A recent study also reports an increased number of intraepithelial cytotoxic T cells in the duodenum of norovirus-infected patients 0–6 days after symptom onset [73]. While specific intestinal lesions are observed during the time of norovirus illness, they completely resolve within two weeks.
Several recent studies suggest that noroviruses cause apoptosis of enterocytes in humans [73], pigs [76], and mice [75]. It is unclear whether viral infection of enterocytes directly induces apoptosis or whether a viral component secreted from other cells acts upon bystander enterocytes to induce their programmed cell death. Murine noroviruses and feline caliciviruses have been demonstrated to cause apoptosis of infected macrophages and epithelial cells, respectively, in vitro through the mitochondrial pathway [77,78,79,80,81], suggesting that apoptosis in vivo may be due to direct effects of infection. Troeger et al. postulate that the observed influx of intraepithelial CD8+ lymphocytes during norovirus infection could cause enterocyte apoptosis upon perforin release [73]. Thus, it is possible that both direct and indirect mechanisms contribute to norovirus-induced apoptosis of enterocytes.
3.2. Physical and Biochemical Manifestations
In one human volunteer study, it was observed that norovirus infection causes a marked delay in gastric emptying thought to be related to the high incidence of vomiting episodes during norovirus infections [82]. Such a pathophysiologic outcome to infection could be caused by an alteration of gastric motor functions or inflammation of the pyloric junction between the stomach and intestine. While mice lack an emetic reflex and thus do not vomit, it has been demonstrated that STAT1−/− mice develop dramatic gastric bloating upon MNV-1 infection [75]. The recapitulation of delayed gastric emptying in a small animal model should facilitate a mechanistic dissection of the basis of this outcome of norovirus infection. A transient malabsorption of D-xylose, fat, and lactose also occurs during acute norovirus infection [71,83]. This malabsorption correlates with shortened microvilli and decreased activity of specific brush border enzymes on enterocytes, including alkaline phosphatase, sucrase, trehalase, and possibly lactase [69]. It is presently unclear whether these changes are due to direct virus-mediated damage to enterocytes, virus-induced alterations of brush border enzyme expression levels (as has been noted for rotavirus [84]), or bystander/immunopathologic effects. Moreover, it is unknown whether there is also a secretory component to norovirus-induced diarrhea.
3.3. Systemic Infection
While it has long been assumed that norovirus infection is confined to the intestine, there is no direct proof for this claim and several recent findings suggest that this dogma be re-considered. First, a recent study detected norovirus RNA in the serum of 15% of infected individuals [85]. Second, work in animal models of norovirus infection support virus dissemination past the intestine: (i) a transient viremia was detected in 50% of gnotobiotic pigs infected with HS66 [76]; (ii) one of five HS66-infected gnotobiotic calves had detectable viral RNA in their serum [74]; and (iii) murine noroviruses are well-documented to spread to mesenteric lymph nodes (MLNs) that drain from the intestine [41,75,86,87,88,89] and to peripheral tissues [41,75,86,88,89]. In particular, MNV-1 replicates efficiently in the spleen and induces specific splenic histological changes, including activation of cells in the white pulp and hypertrophy of cells in the red pulp [75]. The physiological relevance of these splenic changes is unclear. The possibility that human noroviruses disseminate to peripheral sites is a clinically relevant question since mild or sporadic pathologies associated with human norovirus infection of peripheral tissues may have been missed due to the difficulties in their detection and the assumption that they are limited to the intestinal tract. Supporting this idea, there have been several recent associations between norovirus infection and extraintestinal pathologies including encephalopathy [26], disseminated intravascular coagulation [27], and benign infantile convulsions [30,31]. Interestingly, rotaviruses have also recently been confirmed to spread to peripheral tissues and cause histological changes at these sites [90,91,92]. The mechanism of norovirus dissemination is unknown but at least the murine noroviruses infect dendritic cells (DCs), a cell type known to actively migrate from tissues to draining lymph nodes. Thus, it is possible that noroviruses utilize DC infection to facilitate their extraintestinal spread.
3.4. The Course of Norovirus Infection
Norovirus infections are typically considered to be short-lived, lasting for only a few days; however, increasing evidence points to prolonged (or perhaps persistent) infections in some scenarios. Although the symptoms caused by human norovirus infection typically resolve within several days, virus particles can be shed from asymptomatic individuals for weeks after exposure [54,55,56,93]. Further, symptomatic infection has been documented in immunosuppressed individuals with symptoms lasting for over 2 years [13,14,15] and in immunocompetent children with symptoms lasting for up to 6 weeks [9,56]. Additionally, pigtail macaques infected with a human norovirus shed virus in their feces for at least 3 weeks [94]. It is also clear that murine noroviruses can infect wild-type hosts for weeks [41,86,87]. Interestingly, virus is detected not only in fecal samples of these mice but also in MLNs and in some cases intestines and spleens. Thus, there may be mucosal and extraintestinal sites of prolonged norovirus infection. Finally, it has long been recognized that feline caliciviruses establish chronic infections in cats, with animals continuing to shed virus for at least one month following infection and some even becoming life-long carriers [95]. Because these carrier cats are contagious, they represent an important reservoir for the maintenance of feline calicivirus in the feline population [96,97]. Overall, noroviruses can infect their hosts for weeks or months even in the presence of a fully functional immune system. Similar to the situation with feline caliciviruses, it is possible that individuals experiencing prolonged human norovirus infection act as reservoirs of virus, perpetuating its spread within a population.
The reason(s) for prolonged norovirus infection is currently unclear but mechanisms underlying the ability of other viruses to establish persistence may logically play a role. A fundamental feature of persistent viral infections is that the host immune response is ineffective at completely clearing infectious virus [98]. Consistent with this, adaptive immune cells are critical to limit the magnitude of norovirus infection since mice lacking B and T cells contain extremely high levels of MNV-1 within numerous tissues and feces for months following infection [39,99]. The inability to clear persistent infection with other viruses is commonly associated with viral evasion. For example, in persistent viruses such as hepatitis C virus (HCV) there is commonly an alteration of the antigenic epitopes encoded by the virus. Similarly, it has been demonstrated that the feline calicivirus genome acquires mutations in areas predicted to be important for immune recognition during chronic infection, suggesting that altered antigenicity plays a role in maintaining calicivirus persistence [100,101]. One study reports a similar accumulation of mutations within the hypervariable P2 capsid domain in a patient chronically infected with a human norovirus [15]. Interestingly, there is evidence for feline calicivirus that such progressive evolution of a single virus within a host accounts for only a minority of chronic infections; the majority appears to be instead due to cycles of sequential reinfection with either a variant of the same virus strain or with a different strain followed by a period of progressive evolution [102].
Persistent viral infections are also commonly associated with impairment of the normal functioning of immune cells upon infection. Although there is currently no direct evidence that noroviruses impair immune function, it is interesting to note that murine noroviruses infect DCs [103] since many persistent viruses target this cell type and specifically impair their ability to activate T and B cells [98]. Overall, the mechanisms by which noroviruses maintain prolonged infection remain incompletely understood, but they may play a critical role in impairing adaptive immune responses such that they fail to protect from secondary challenge. It will be interesting in future studies to examine the relationship between prolonged norovirus infection and the lack of protective immunity elicited by primary infection. Prolonged virus shedding from individuals in whom symptomatic infection has resolved also surely contributes to the difficulty in controlling norovirus outbreaks.
4. Cellular Determinants of Norovirus Pathogenesis
The precise cell tropism of human noroviruses is unknown. When intestinal biopsy sections from volunteers infected with either a GI.1 or a GII.1 norovirus were analyzed by electron microscopy, virus particles were not observed [70,83], leading investigators to postulate that human noroviruses infect very few cells in vivo. This hypothesis is supported by work in animal models (described below). Furthermore, numerous attempts to identify a relevant cell culture system for these viruses have been unsuccessful [104]. The inability to propagate human noroviruses has long been the major obstacle to studying this virus family. However, numerous recent advances in this field offer promise of overcoming this barrier. Moreover, recent work in animal models suggests that multiple intestinal cell types including enterocytes, macrophages (Mφs), and DCs may support at least low level in vivo replication.
4.1. Recent Advances of In Vitro Human Norovirus Infection Models
Extensive efforts have failed to detect norovirus replication in intestinal epithelial cell lines [104]. There has been a single report describing human norovirus infection of a 3-dimensional model of human small intestinal epithelium [105]; however, several independent laboratories failed to detect increased viral genome in these cells during the period of noted cytotoxicity, strongly suggesting that cellular pathology was not due to viral replication [106]. Based on the tropism of murine noroviruses for DCs and Mφs [103], in vitro infections of human DCs and Mφs derived from monocytes isolated from peripheral blood mononuclear cells (PBMCs), or myeloid DCs directly isolated from PBMCs, were performed with the G1.1 Norwalk virus. There was no evidence for viral replication in these cells (Lay MK, Atmar RL, Guix S, Bharadwaj U, He H, Neill FH, Sastry JK, Yao Q and Estes MK, manuscript in preparation). While these results demonstrate that at least one human norovirus does not infect peripheral DCs and Mφs in vitro, DCs and Mφs can differ significantly in pathogen susceptibility depending on their source and differentiation [107,108,109]; thus it remains possible that other types of DCs and Mφs (i.e., mucosal cells) are permissive to human noroviruses.
Based on several recent studies, it is now clear that the block to human norovirus propagation in vitro occurs at the level of receptor binding, internalization, or genome uncoating. Specifically, Guix et al. demonstrated that transfecting norovirus RNA into Huh-7 cells results in a complete single round of viral replication culminating in progeny virion release [110]. Furthermore, transfection of nonpermissive cells with infectious norovirus clones driven by either the T7 polymerase or the host RNA polymerase II results in genome replication and virion production [111,112,113,114]. Similarly, a norovirus replicon has been generated that results in genome replication and nonstructural protein production in stably transfected cells [115]. Overall, it can be concluded that bypassing the initial steps of virus entry and uncoating overcomes the block to norovirus replication in cultured cells.
4.2. Human Norovirus Cell Tropism in Animal Models
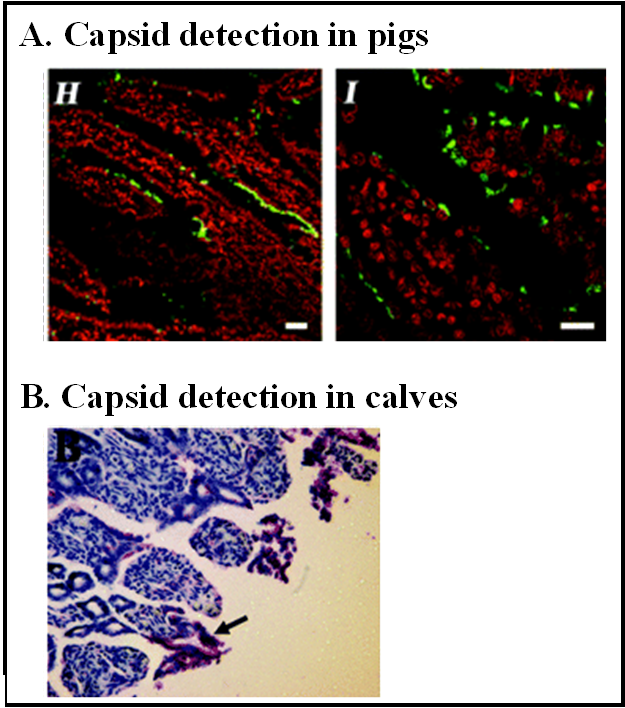
Both gnotobiotic pigs and gnotobiotic calves are susceptible to infection with at least one human norovirus, a GII.4 virus called HS66 [74,76]. Infected pigs and calves develop diarrhea and histopathological changes are evident in the intestines of infected calves. In HS66-infected gnotobiotic pigs, virus particles are apparent in cytoplasmic vesicles within enterocytes [76]. Moreover, viral capsid can be detected in enterocytes located at villous tips and along the sides of villi, but rarely in crypts, of the duodenum and jejunum (Figure 2A). Nonstructural protein indicative of viral replication can also be detected in a limited number of enterocytes. In HS66-infected gnotobiotic calves, viral structural protein is detectable in enterocytes (Figure 2B) and in lamina propria cells resembling Mφs along the jejunum and ileum, but not along the duodenum [74]. The discrepancy between these animal model studies and earlier studies that failed to detect virus in human intestinal sections could be due to (i) virus strain-dependent differences (GII.4 versus GI.1 and GII.1 viruses, respectively); (ii) virus dose differences; (iii) species-specific disparities in norovirus infection; or (iv) differences in assay sensitivity between studies.
Figure 2.
The intestinal cell tropism of HS66 in gnotobiotic pigs and calves.(A) Paraffin-embedded sections of the duodenum of HS66-infected gnotobiotic pigs prepared 4 dpi were stained with a monoclonal antibody against the GII human norovirus capsid protein (called NS14). Fluorescently conjugated anti-mouse secondary antibody facilitated visualization of capsid through immunofluorescent confocal microscopy (in green). Nuclei were stained with propidium iodide (in red). Data are reproduced from J. Virol.2006, 80, 10372–10381. (B) Paraffin-embedded sections of the jejunum of HS66-infected gnotobiotic calves prepared 3 dpi were stained with NS14. Viral capsid was visualized through immunohistochemistry. Data are reproduced from J. Virol.2008, 82, 1777–1786.
Figure 2.
The intestinal cell tropism of HS66 in gnotobiotic pigs and calves.(A) Paraffin-embedded sections of the duodenum of HS66-infected gnotobiotic pigs prepared 4 dpi were stained with a monoclonal antibody against the GII human norovirus capsid protein (called NS14). Fluorescently conjugated anti-mouse secondary antibody facilitated visualization of capsid through immunofluorescent confocal microscopy (in green). Nuclei were stained with propidium iodide (in red). Data are reproduced from J. Virol.2006, 80, 10372–10381. (B) Paraffin-embedded sections of the jejunum of HS66-infected gnotobiotic calves prepared 3 dpi were stained with NS14. Viral capsid was visualized through immunohistochemistry. Data are reproduced from J. Virol.2008, 82, 1777–1786.

4.3. Murine Norovirus Cell Tropism
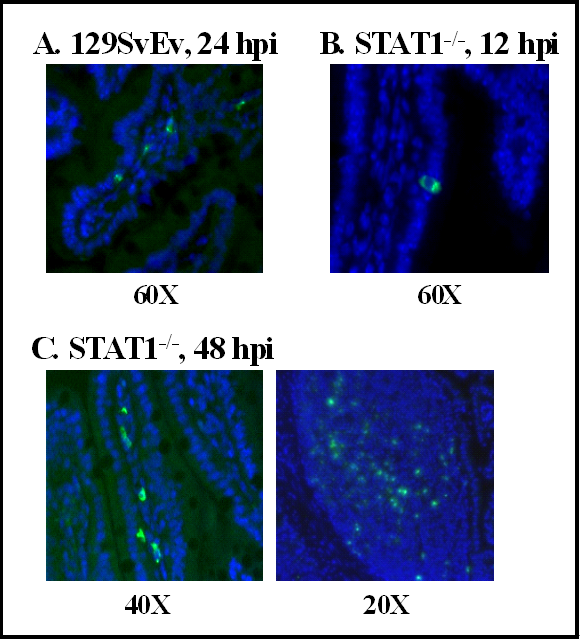
A number of murine norovirus strains have now been isolated from mouse colonies in academic research settings around the world [39,40,41,42,86]. Murine noroviruses are enteric in nature: (i) most strains were originally isolated from fecal samples or MLNs; (ii) they are infectious orally and can be detected in mucosal sites (intestines, MLNs) during infection; and (iii) they are shed in feces [39,40,41,42,75,86]. At least one murine norovirus causes decreased fecal consistency and mild intestinal inflammation in infected wild-type mice, and severe gastroenteritis in STAT1−/− mice [75,88]. Murine noroviruses are known to replicate in DCs and Mφs, but not other cell types, in vitro [39,40,41,86,103,116,117]. Moreover, in MNV-1-infected wild-type mice, viral nonstructural protein can be detected in lamina propria cells morphologically resembling DCs and Mφs of rare intestinal villi at 24–48 hpi (Figure 3A) [75]. The in vivo infection of DCs and Mφs by MNV-1 is further supported by the immunohistochemical detection of MNV-1 antigen in cells that resemble Mφs in the liver, and Mφs and DCs in the spleen [103]. It has also been reported that immunodeficient mice naturally infected with a murine norovirus contain viral antigen in the cytoplasm of inflammatory cells in the liver, splenic red and white pulp, intestinal lamina propria, intestinal lymphoid follicles, lung, pleural and peritoneal cavities, and MLNs [89,118]. In the absence of an intact interferon signaling pathway, MNV-1 antigen is detected more abundantly and in additional intestinal cell types of infected mice. Specifically, viral antigen can be detected in enterocytes of STAT1−/− mice at 12 hpi (Figure 3B), and lamina propria and Peyer’s patch cells at 24–48 hpi (Figure 3C) [75]. Thus, interferon may restrict the cell tropism of noroviruses, as has been shown for a number of other viruses [119,120,121]. If this is the case, norovirus strains with differing sensitivity to interferon responses may display disparate cell tropism.
Figure 3.
The intestinal cell tropism of MNV-1 in wild-type and STAT1−/− mice.Intestinal sections prepared from MNV-1-infected 129SvEv(A) or STAT1−/− (B and C)mice were stained with guinea pig polyclonal antibody raised against the MNV-1 Pro:Pol nonstructural protein. Fluorescently conjugated secondary antibody was then used to visualize viral protein through immunofluoresence, in which viral protein is pseudocolored in green and nuclei in blue. Data are reproduced from J. Virol.2007, 81, 3251–3262.
Figure 3.
The intestinal cell tropism of MNV-1 in wild-type and STAT1−/− mice.Intestinal sections prepared from MNV-1-infected 129SvEv(A) or STAT1−/− (B and C)mice were stained with guinea pig polyclonal antibody raised against the MNV-1 Pro:Pol nonstructural protein. Fluorescently conjugated secondary antibody was then used to visualize viral protein through immunofluoresence, in which viral protein is pseudocolored in green and nuclei in blue. Data are reproduced from J. Virol.2007, 81, 3251–3262.

4.4. Norovirus Receptors
Human noroviruses recognize histo-blood group antigens (HBGAs) that are expressed on the surface of mucosal epithelial cells [122]. HBGAs are neutral carbohydrates linked to proteins or lipids on cell surfaces. The glycosyltransferases that control their synthesis are encoded by the highly polymorphic ABO, Lewis, and secretor gene families. The association of noroviruses with HBGAs has been demonstrated to be essential for certain virus strains. This is best exemplified by the strict correlation between secretor status and susceptibility to Norwalk virus infection – Norwalk virus recognizes α1,2-linked fucose residues [63] whose expression on gut epithelial cells and in body fluids is dependent on a wild-type FUT2 gene; while individuals with a wild-type FUT2 gene (~80% of the population; referred to as secretors) are susceptible to Norwalk virus infection, individuals that contain a null FUT2 allele (~20% of the population; referred to as nonsecretors) are completely resistant [123].
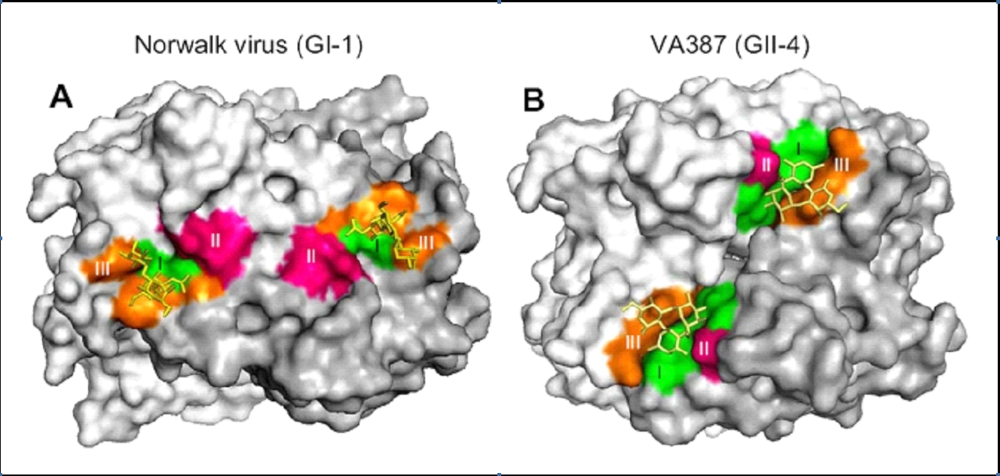
A number of distinct norovirus strain-specific binding profiles have been reported [124,125,126]. As described recently by Tan et al., strains can be segregated generally into those that bind A/B epitopes and those that bind Lewis epitopes [127]. While GI and GII strains are contained within both binding categories, there are genogroup-specific differences in the receptor binding interface of the norovirus capsid that interacts with the HBGA. Thus, even if a GI and a GII norovirus interact with the same HBGA, the precise residues involved in this binding will most likely be different (Figure 4). In terms of the highly prevalent GII.4 noroviruses, strains circulating in 1974, 1977, and 1987 all display a common HBGA binding pattern (H type 3, LewisY, and B antigens); a strain circulating in 1997 binds the same set of HBGAs in addition to A and LewisB antigens; a strain isolated from a 2002 outbreak displays a unique HBGA pattern (LewisA, LewisX, and A antigens); and another 2002 strain in addition to strains isolated from 2004 and 2005 outbreaks fail to bind efficiently to any HBGA suggesting they may bind other carbohydrates [63,128]. Interestingly, a strain circulating in 2006 displays a similar HBGA binding profile to the 1997 pandemic strain [129]. While norovirus strains display distinct HBGA binding properties, collectively they can infect nearly all individuals due to their high genetic variability [130]. This highlights the highly adaptive nature of noroviruses and the likelihood of a long co-evolution of human noroviruses with their human host.
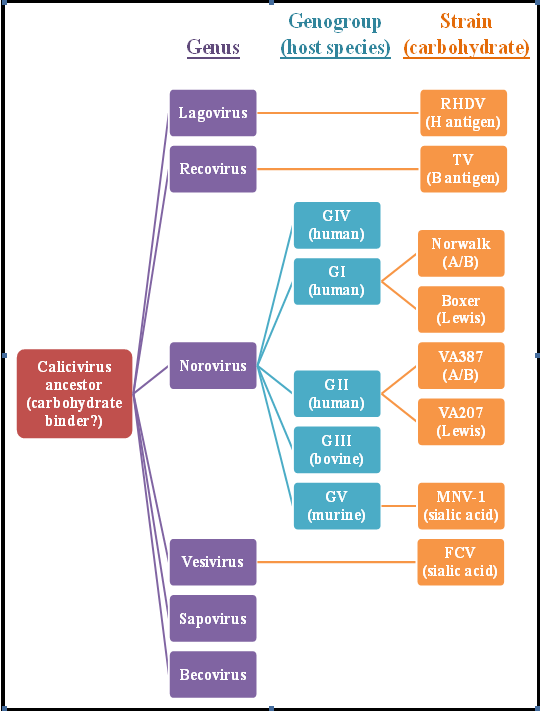
Several animal caliciviruses also bind HBGAs, including the highly virulent rabbit hemorrhagic disease virus that segregates within the lagovirus genus [131] and the Tulane virus isolated from rhesus macaques that defines the newly proposed recovirus genus [127,132]. Conversely, other animal caliciviruses, including multiple feline calicivirus strains [133] and murine norovirus strains [134], bind sialic acid on permissive cells. Thus, carbohydrate binding appears to be conserved across the Caliciviridae family while specific members can bind neutral or negatively charged residues (Figure 5). There are examples of human noroviruses that do not bind HBGAs, including recent GII.4 pandemic strains [125,128]; it has yet to be determined whether these strains instead bind negatively charged sugars similar to feline caliciviruses and murine noroviruses. Evidence supporting this possibility is provided by studies demonstrating binding of GII noroviruses to negatively charged heparan sulfate [135] and sialylated Lewisx carbohydrate [136].
5. Molecular Determinants of Norovirus Pathogenesis
Understanding the molecular aspects of viral replication can facilitate the development of rational therapies. In this section, we summarize recent findings of norovirus replication mechanisms that may contribute to norovirus pathogenesis and ultimately lead to the design of novel antiviral strategies.
Figure 4.
The HBGA binding interface on norovirus particles is genogroup-specific.Crystal structures have been determined for P dimers of a representative GI norovirus (A) and a representative GII norovirus (B) complexed with HBGA. Surface models are shown here, with P dimers colored gray (one monomer is darker gray than the other) and the three major components of the HBGA binding interface colored green, red, and orange. The HBGA is colored yellow. While the binding sites on both viruses lie within the exposed P2 domain, the exact residues and HBGA binding mode differ. Moreover, note that the HBGA binding interface of Norwalk virus is located within a single monomer while the interface of VA387 spans both monomers. Data are reproduced from PLoS ONE2009, 4, e5058.
Figure 4.
The HBGA binding interface on norovirus particles is genogroup-specific.Crystal structures have been determined for P dimers of a representative GI norovirus (A) and a representative GII norovirus (B) complexed with HBGA. Surface models are shown here, with P dimers colored gray (one monomer is darker gray than the other) and the three major components of the HBGA binding interface colored green, red, and orange. The HBGA is colored yellow. While the binding sites on both viruses lie within the exposed P2 domain, the exact residues and HBGA binding mode differ. Moreover, note that the HBGA binding interface of Norwalk virus is located within a single monomer while the interface of VA387 spans both monomers. Data are reproduced from PLoS ONE2009, 4, e5058.

Figure 5.
The known carbohydrate binding partners of caliciviruses. At least one member from four of the six calicivirus genera (colored in purple) has been demonstrated to bind a carbohydrate, suggesting that the original calicivirus ancestor was a carbohydrate binder. Similarly, three of the five norovirus genogroups (colored in blue) have been shown to contain members that bind carbohydrates. Two of the three norovirus genogroups containinghuman members have been shown to specifically bind HBGAs, either A/B/H antigens (A/B) or Lewis antigens (Lewis).Representative virus strains and their known carbohydrate ligands are shown in orange. Data are adapted from PLoS ONE2009, 4, e5058.
Figure 5.
The known carbohydrate binding partners of caliciviruses. At least one member from four of the six calicivirus genera (colored in purple) has been demonstrated to bind a carbohydrate, suggesting that the original calicivirus ancestor was a carbohydrate binder. Similarly, three of the five norovirus genogroups (colored in blue) have been shown to contain members that bind carbohydrates. Two of the three norovirus genogroups containinghuman members have been shown to specifically bind HBGAs, either A/B/H antigens (A/B) or Lewis antigens (Lewis).Representative virus strains and their known carbohydrate ligands are shown in orange. Data are adapted from PLoS ONE2009, 4, e5058.

5.1. Norovirus Genomic Structures
Noroviruses have plus-stranded linear RNA genomes of 7.4–7.7 kb which are typically organized into three open reading frames (ORF1-3) (Figure 6) [138,139,140]. Murine noroviruses have a predicted fourth ORF overlapping ORF2 but in an alternate reading frame [41]; it has yet to be determined whether this ORF codes for a functional protein. The 5’ proximal region of the norovirus genome encodes all of the nonstructural proteins in a single ORF (ORF1) while the 3’ proximal region, transcribed into a subgenomic message, encodes the major and minor structural proteins in two separate ORFs (ORF2 and ORF3, respectively). Norovirus genomes are thought to be covalently associated with a viral protein called VPg at their 5’ ends and are polyadenlyated at their 3’ ends.
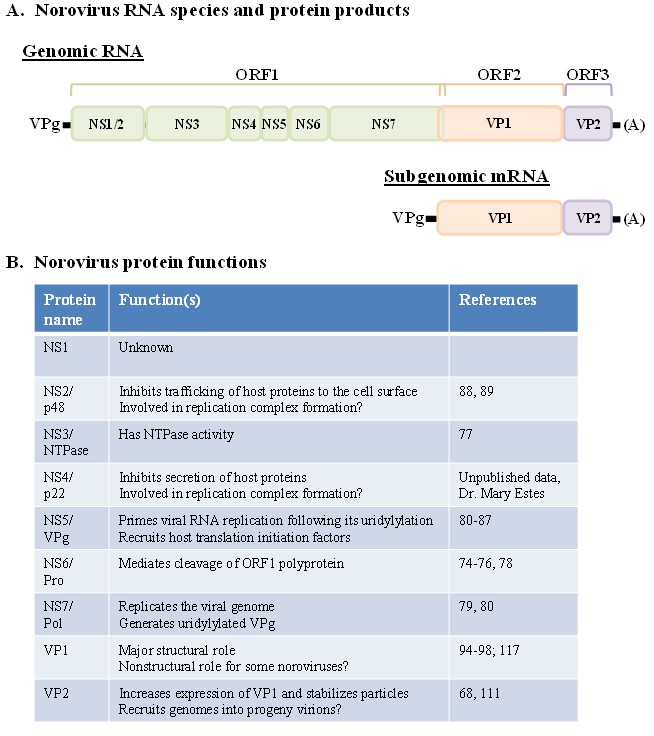
Figure 6.
Norovirus genomic organization and protein function. (A)Norovirus genomes are comprised of a single linear piece of positive-sense RNA between 7.4–7.7 kb. They contain three open reading frames, one encoding a polyprotein of 7 nonstructural protein products (colored in green), one encoding the major structural capsid protein VP1 (colored in orange), and one encoding the minor structural protein VP2 (colored in purple). There is a short overlap between ORFs 1 and 2. A subgenomic mRNA that is 3’ co-terminal with full-length genomes is produced during norovirus replication. This mRNA acts as a template for the production of structural proteins; similar to genomic RNA, it is covalently linked to VPg at its 5’ end and polyadenylated at its 3’ end. (B)Known and hypothesized functions of mature norovirus proteins.
Figure 6.
Norovirus genomic organization and protein function. (A)Norovirus genomes are comprised of a single linear piece of positive-sense RNA between 7.4–7.7 kb. They contain three open reading frames, one encoding a polyprotein of 7 nonstructural protein products (colored in green), one encoding the major structural capsid protein VP1 (colored in orange), and one encoding the minor structural protein VP2 (colored in purple). There is a short overlap between ORFs 1 and 2. A subgenomic mRNA that is 3’ co-terminal with full-length genomes is produced during norovirus replication. This mRNA acts as a template for the production of structural proteins; similar to genomic RNA, it is covalently linked to VPg at its 5’ end and polyadenylated at its 3’ end. (B)Known and hypothesized functions of mature norovirus proteins.

Essential viral genomic structures represent putative antisense drug targets. Moreover, identification of essential RNA elements within the viral genome may facilitate the construction of viruses with a dominant inhibitory effect on the replication of wild-type virus [141]. Recent bioinformatic analyses of calicivirus genomes have identified conserved secondary structures that include: (i) two or more 5’ terminal stem-loops; (ii) a 3’ terminal hairpin; (iii) a stem-loop just upstream of the ORF1/2 junction in the antigenomic strand proposed to be a component of the subgenomic promoter; and (iv) a stem-loop at the 5’ end of the polymerase coding region with a motif characteristic of picornavirus cre elements that dictate VPg uridylylation [142,143]. Several of these structures were determined to be critical for the replication of a murine norovirus [142].
5.2. Norovirus Nonstructural Proteins
Norovirus ORF1 encodes a large polyprotein that is cleaved into seven mature nonstructural proteins (NS1-NS7) [144,145,146]. A number of these proteins have defined activities, including an NTPase (NS3) [147], a protease (Pro; NS6) [148], and an RNA-dependent RNA polymerase (Pol; NS7) [149,150]. Noroviruses also encode a VPg (NS5) that, based on work with animal caliciviruses, is presumed to be covalently attached to the 5’ ends of viral genomes in place of a typical 5’ cap structure [151,152,153]. The norovirus VPg can function as a primer in viral RNA replication following its uridylylation [150,154]. However, the norovirus Pol can also initiate RNA polymerization in a primer-independent manner; it has been postulated that it uses uridylylated VPg primer-mediated polymerization to replicate polyadenylated genomes and primer-independent polymerization to copy antigenomes [149,150]. The norovirus VPg also interacts with host translation initiation factors, perhaps to recruit them to the 5’ end of the RNA genome for translation initiation [155,156,157].
A number of the norovirus nonstructural proteins do not share significant sequence homology to proteins of known function, including NS1, NS2, and NS4 [4], and their functions in viral replication are only beginning to be unraveled. While there is no information available on the role played by the most amino-terminal protein NS1 in norovirus replication, NS2 (also referred to as p48 and the N terminal protein) has been shown to co-localize with the Golgi complex in transfected cells and induce rearrangement of the Golgi membrane [158]. It also interacts with a host protein involved in regulating vesicle transport and can inhibit cell surface expression of proteins [159]. Expression of NS4 (also referred to as p22 and the 3A-like protein) in transfected cells has recently been shown to induce Golgi disruption. Moreover, NS4 contains a novel ER export signal that results in aberrant trafficking of COPII-coated vesicles and ultimately in inhibition of host protein secretion (Sharp TM, Guix S, Katayama K, Crawford SE and Estes MK, manuscript submitted). Redistribution of COPII-coated vesicles was not observed during MNV-1 infection of RAW264.7 cells [160], suggesting that human and murine noroviruses may differentially affect secretory pathways. Overall, available data suggest that norovirus NS2 and NS4 proteins both contribute to viral replication complex formation on intracellular membranes including that of the Golgi apparatus. This is supported by work with MNV-1, demonstrating that all nonstructural proteins localize to the ER, Golgi bodies, and early endosomes, but not to late endosomes or lysosomes [160]. One can postulate that NS2 and NS4 disrupt intracellular host protein trafficking, which may play an important role in the pathogenesis of norovirus infections by blocking surface expression (i.e., MHC molecules) and secretion (i.e., cytokines) of immune mediators in infected cells.
Any of the essential activities encoded by norovirus nonstructural proteins could potentially be targeted with antiviral compounds for intervention. In addition, immunotherapeutic strategies based on enhancing the host cell’s normal antiviral mechanisms may be engineered to disrupt the function of one or more norovirus nonstructural proteins. For example, during MNV-1 infection of permissive cells, translation of nonstructural proteins can be inhibited by both type I and type II interferon responses [161]. A host innate immune mediator induced by interferon may thus block required interactions between norovirus VPg and translation initiation factors, representing a candidate immunotherapeutic to treat norovirus infections. Both type I and type II interferon also inhibit the replication of a human norovirus expressed from a replicon in transfected cells [115,162]; it has yet to be determined whether these responses target norovirus translation as they do in the murine system.
5.3. Norovirus Structural Proteins
Noroviruses are nonenveloped icosahedral viruses between 27–30 nm in diameter. Norovirus ORF2 encodes the major structural protein called VP1 or capsid. Virions contain 180 copies, or 90 dimers, of VP1 that assemble into icosahedral particles exhibiting T = 3 symmetry [163,164,165,166,167]. Each capsid dimer forms an arch-like protrusion extending from the internal shell of the particle. The capsid protein itself can be divided into a conserved internal shell domain (S) and a more variable protruding domain (P) that forms the arch-like protrusions. The P domain can be further subdivided into P1 and P2 domains, with P1 forming the sides of the arches and P2 located at the exposed tops of the arches. Expression of VP1 independent of other viral components results in the self-assembly of virus-like particles (VLPs) [168], and expression of solely the P domain of VP1 results in the formation of P dimers as well as P particles composed of 12 P dimers [169]. These structures retain the binding properties of native norovirus virions, at least in terms of carbohydrate association, and have been extremely useful in binding and antigenicity studies in the absence of a cell culture system [170]. Consistent with its exposed location on the virion surface, the P2 domain of VP1 comprises a hypervariable region that is suspected to contain receptor binding and antigenic sites. Indeed, numerous structural and mutagenesis studies of VLPs and P domain structures have confirmed that the binding interface for HBGA association with norovirus particles resides in the P2 domain of capsid [169,171,172,173,174,175]. Furthermore, monoclonal antibodies have been demonstrated to recognize conformational epitopes located in the P2 domain [176,177]. Identifying critical determinants of norovirus receptor binding and antigenicity may be instrumental in the design of effective therapeutics and vaccines. Interestingly, a single amino acid change in the P2 domain of a murine norovirus has been associated with its in vivo attenuation [103,178].
Norovirus ORF3 encodes a minor structural protein called VP2 that is present in only 1–2 copies per virion [138,179]. VP2 is small, basic, and quite divergent in both size and sequence between viruses within this family. Its function in viral replication is currently undefined but there is evidence that it increases the expression level of VP1 and that it stabilizes VLPs [180]. The basic charge of VP2 suggests that it may function in encapsidation of the viral genome.
Both VP1 and VP2 are translated from a subgenomic message that is polyadenylated and, based on work with feline caliciviruses [152], is presumed to be covalently attached to VPg at its 5’ end. While expression of VP1 is presumed to occur via VPg-directed translation initiation (similar to expression of nonstructural proteins from genomic RNA), the mechanism for translation initiation of the downstream VP2 protein was unclear until recently. There is now evidence from several animal caliciviruses that translation of VP2 proceeds via a translation termination re-initiation (TTR) mechanism. In this process, posttermination ribosomes from VP1 translational events remain associated with subgenomic RNA through interactions with termination upstream ribosomal binding sites (TURBSs) in the 3’ proximal end of VP1, facilitating recruitment of translation initiation factors to the 5’ end of VP2 [181,182,183,184]. At least one of the TURBS is conserved across the Caliciviridae family, suggesting that noroviruses rely on the same mechanism for VP2 expression [181]. A recent study also suggests that TTR is used by at least one norovirus to express VP1 independent of subgenomic message production [185]; while this most likely contributes minimal amounts of capsid compared to the levels produced from translation of subgenomic RNA, the authors speculate that VP1 may play an additional nonstructural role(s) in the viral replication cycle.
6. Norovirus Immunity and Vaccination
6.1. Immunity
Immunity to noroviruses is complicated by the heterogeneous responses of the human population and the transient nature of immunity in some individuals. These atypical responses are best highlighted by early studies in which volunteers were repeatedly exposed to homologous virus inoculum comprised of filtered stool from norovirus outbreak patients [5,6]. In these studies, several patterns of susceptibility were noted – a proportion of volunteers were resistant to primary and subsequent infections, and a proportion of individuals were equally susceptible to primary and secondary infections when there was at least a six-month interval between challenges. One possible explanation for the continued susceptibility of a subset of individuals is that they fail to generate a virus-specific immune response upon norovirus infection. However, data gathered from many human volunteer challenge studies and natural norovirus outbreaks conclusively show that people do develop a virus-specific antibody response and that the presence of this response does not correlate with protection [5,6,93,123,186,187,188,189]. Specifically, virus-specific serum IgG is induced and persists for months following infection, while serum IgA and IgM responses are more short-lived. Mucosal IgA is induced as well but its duration has not been determined [190]. Because there is no cell culture system for human noroviruses, it has not been possible to determine whether norovirus-specific antibodies are neutralizing. However, a surrogate assay based on inhibition of VLP binding to HBGA ligands has been used to demonstrate that virus-specific serum antibodies generated during experimental and natural infections neutralize carbohydrate binding [128,129,191,192]. Moreover, traditional neutralization assays are possible for murine noroviruses because they are cultivable. MNV-1 infection of mice induces neutralizing serum antibodies [103] and one MNV-1-neutralizing monoclonal antibody has been shown to bind each of the 180 hypervariable P2 domains of native MNV-1 virions without inducing a major conformational change, suggesting that neutralization in this case is mediated by abrogation of cellular attachment [167]. Nevertheless, the duration of norovirus antibody responses has not been clearly defined, so it remains possible that waning humoral immunity is related to the susceptibility of some previously exposed individuals to repeat norovirus infections.
While there have been no direct studies of T cell responses to human norovirus infection, surrogate cytokine studies suggest that virus-specific T cells are also induced in infected individuals. In one study, peripheral blood mononuclear cells (PBMCs) were collected from norovirus-infected volunteers at pre-challenge and post-challenge time points, incubated with norovirus VLPs, and secretion of cytokines from the PBMCs was measured [193]. The authors noted increases in the Th1 cytokines IFNγ and IL-2, as well as the Th2 cytokine IL-5. Notably though, similar increases in cytokine levels were observed when pre-challenge and post-challenge PBMCs from uninfected individuals were compared to each other, presumably indicating that controls in this study were previously exposed to natural norovirus infection. This finding highlights the difficulty of studying primary immune responses to human noroviruses where infection histories are impossible to discern. Recent studies in the gnotobiotic pig and calf models also report moderate increases in several Th1 and Th2 cytokines as well as interferon in the serum and intestinal contents of infected animals, suggestive of a T cell response upon primary challenge [74,194]. Overall, the available information on secondary human norovirus infections supports an atypical pattern of immunity in which some previously exposed individuals are susceptible to repeat infections even in the face of virus-specific memory immune responses. Another subset of individuals is resistant to infection, presumably due to genetic resistance (i.e., nonsecretors) or preexisting immunity.
In the early volunteer challenge studies, it was noted that susceptible individuals were protected from disease when re-challenged soon after secondary exposure (<6 months) in contrast to a more distant re-challenge (>6 months) [5,6], suggesting that short-term norovirus immunity is elicited whereas long-term immunity is not. Studies of murine norovirus infection lend support to this explanation – a primary MNV-1 exposure results in reduced virus loads upon re-challenge when the interval between primary and secondary infections is short (Figure 7; 6-week challenge), although it is important to note that this magnitude of protection is insufficient to protect the host from disease [88]. While protective peripheral immunity in this setting is maintained over time, protective mucosal responses wane (Figure 7, 9-month challenge; Karst laboratory, unpublished data), indicating that memory immune responses at mucosal sites are not efficiently maintained. There is also evidence from the murine model of norovirus infection that repeated norovirus exposure boosts the long-term immune response – repeated exposure to high-dose MNV-1 elicits long-lasting protective immunity [99], whereas single high-dose infection does not [88]. The immunity elicited by repeated exposure to MNV-1 does appear to be lasting [99]. Interestingly, studies of murine norovirus infection also signify that lower doses of virus elicit stronger memory immune responses than high doses [88]. It has yet to be determined whether virus dose similarly displays an inverse correlation to protection upon human norovirus infection.
In apparent contradiction to the results of early human volunteer studies, some investigators have recently suggested that noroviruses can elicit herd immunity [62,123,129,193]. This supposition is based on the epochal pattern of GII.4 pandemics, with emergence of a dominant virus strain for a 1–2 year period followed by a period of relative GII.4 quiescence prior to emergence of a new dominant strain (Figure 8) [62]. The emergence of new pandemic GII.4 strains is postulated to result from (i) antigenic drift-mediated altered carbohydrate usage and (ii) altered antigenicity facilitating virus escape of herd immunity [128,195]. Consistent with this, only a minimal number of amino acid changes in the capsid P domain are required to alter HBGA usage, enabling norovirus strains great flexibility in evolving to infect new populations [195]. It has also been reported in a recent human norovirus challenge study that a subset of secretor-positive volunteers were resistant to Norwalk virus infection, and these individuals displayed a modest increase in norovirus-specific salivary IgA early after infection [123]. While interesting, interpretation of these studies is complicated by several factors: the previous exposure histories of the subjects are unknown, and it is not possible to distinguish between short- and long-term immunity. Moreover, these studies do not offer an explanation for the observation that some individuals are susceptible to repeated infections with homologous virus [5,6].
Figure 7.
Mucosal norovirus immunity wanes over time.Groups of 129SvEv mice (5–14 mice per group) were mock-infected (1°) or inoculated perorally with 104 pfu MNV-1.CW3 (2°). Six weeks or nine months later, all mice were infected with 107 pfu MNV-1.CW3. One day after secondary challenge, animals were perfused, organs were harvested, and viral burden was determined by plaque assay. The data for all mice in each group are averaged. Limits of detection are indicated by dashed lines. Fold-reductions for titers in mice receiving secondary challenge compared to those in mice receiving primary challenge are listed above the black bars.
Figure 7.
Mucosal norovirus immunity wanes over time.Groups of 129SvEv mice (5–14 mice per group) were mock-infected (1°) or inoculated perorally with 104 pfu MNV-1.CW3 (2°). Six weeks or nine months later, all mice were infected with 107 pfu MNV-1.CW3. One day after secondary challenge, animals were perfused, organs were harvested, and viral burden was determined by plaque assay. The data for all mice in each group are averaged. Limits of detection are indicated by dashed lines. Fold-reductions for titers in mice receiving secondary challenge compared to those in mice receiving primary challenge are listed above the black bars.

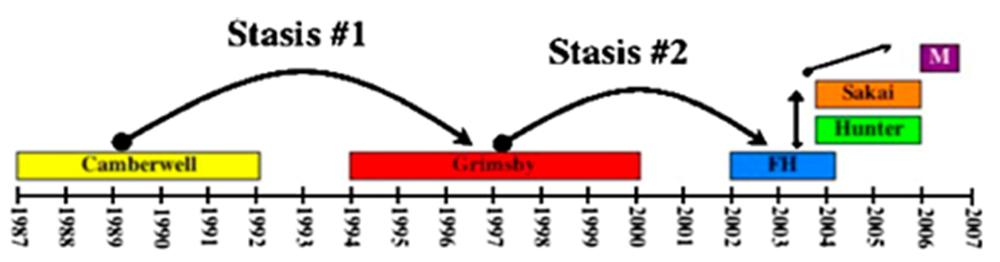
Figure 8.
Epochal evolution of the GII.4 noroviruses. Early GII.4 norovirus strains (Camberwell and Grimsby) persisted for long periods and were followed by shorter periods of stasis before being replaced by a new dominant strain. The evolution of GII.4 viruses appears to have become much more rapid in recent years, with pandemic strains being replaced by a new dominant strain in 1–2 years. FH, Farmington Hills; M, Minerva.
Figure 8.
Epochal evolution of the GII.4 noroviruses. Early GII.4 norovirus strains (Camberwell and Grimsby) persisted for long periods and were followed by shorter periods of stasis before being replaced by a new dominant strain. The evolution of GII.4 viruses appears to have become much more rapid in recent years, with pandemic strains being replaced by a new dominant strain in 1–2 years. FH, Farmington Hills; M, Minerva.

One explanation that would be inclusive of all norovirus immunity observations to date is that short-term herd immunity drives the evolution of emerging norovirus strains, but this immunity wanes over time such that genetically susceptible individuals can be repeatedly infected with homologous virus over the long-term. Studies of GII.4 carbohydrate usage may support this idea – while 2002–2005 strains display distinct binding profiles compared to earlier 1974–1997 strains, the binding profile of the more recent 2006 GII.4 strain is similar to those of 1974–1997 strains [63,195].
6.2. Candidate Norovirus Vaccination Strategies
Current efforts to design human norovirus vaccines focus on expression of the norovirus capsid protein in various vectors, resulting in self-assembly of the capsid protein into immunogenic VLPs [4]. Such vaccine candidates elicit both serum IgG and mucosal IgA responses, but the duration of these responses and their protective potential have not been determined. A Phase I safety trial has been completed for a dry powder norovirus VLP vaccine delivered intranasally in conjunction with a TLR4 agonist adjuvant; the vaccine was determined to be immunogenic and was not associated with adverse side effects (http://www.ligocyte.com/downloads/Noro.pdf). Live virus challenge studies are now underway. While challenge studies following exposure to human norovirus VLPs have not been reported, results from animal models are encouraging: (i) intranasal inoculation of calves with bovine norovirus VLPs + adjuvant induces both mucosal and systemic humoral immunity, although protection from subsequent challenge is only partial and viral shedding is not prevented [196]; (ii) repeated mucosal exposure to HS66 VLPs + adjuvant elicits immune responses and protection from live HS66 in a majority of vaccinated gnotobiotic pigs [197]; (iii) footpad inoculation of mice with murine norovirus VLPs + adjuvant induces humoral immune responses and reduced virus loads, although protection from disease was not assessed in this study [198]. Additional work with murine norovirus suggests that a broad adaptive immune response including B cells, CD4 T cells, and CD8 T cells, is critical for full protection [99].
While available information on norovirus VLP-based vaccines is promising, several potential obstacles require serious consideration. Feline calicivirus vaccines, either modified live vaccines or inactivated vaccines given with adjuvants, have been used for many years [95]. These vaccines induce protective immunity in cats in terms of clinical disease prevention; however, they do not prevent infection or persistence. In such a scenario, the presence of non-neutralizing immunity could actually induce the emergence of virulent strains [199]. To this end, multiple virulent feline calicivirus isolates have recently emerged that cause systemic disease associated with high mortality; vaccinated cats have been prominently affected in these outbreaks [200,201]. These data highlight the potential necessity of designing norovirus vaccination strategies that target both clinical disease and prolonged infection. Other major obstacles in designing norovirus vaccines are the genetic variability within this virus family and the replacement of pandemic strains in short time intervals (see Figure 8). These factors may require a continually evolving vaccine preparation similar to the one currently in place for influenza viruses. Moreover, multiple norovirus strains may need to be included in this preparation to elicit effective immunity to each genogroup. A final possible hindrance to a successful norovirus vaccine is the inability of natural infection to elicit lasting protection in all individuals. It will be critical to delineate the basis for this lack of immunity and to determine whether a similar phenomenon occurs upon exposure to candidate norovirus vaccines.
7. Conclusions
Recent advances in the norovirus field have greatly facilitated progress in our understanding of how this emerging RNA virus family causes disease and persists worldwide. These advances include the identification and structural characterization of HBGAs as receptors, the development of animal models and a murine norovirus cell culture system, and the design of norovirus infectious clones and replicons. While much progress has been made in the human norovirus field, it is still limited by the lack of an efficient cell culture model. Designing efficacious norovirus vaccines and therapies with broad spectrum activity across the family also remains a major challenge due to several factors: (1) the extreme genetic variability within the norovirus family and within genogroups; (2) the rapid evolution of antigenically dissimilar pandemic GII.4 norovirus strains; and (3) the lack of lasting immunity upon natural exposure to noroviruses in at least a proportion of the population. Studies in the murine norovirus model suggest that immunity to natural infection does indeed wane over time but can be enhanced by repeat exposure. Moreover, the finding that the HBGA binding interface is conserved within a norovirus genogroup may facilitate the development of novel therapeutic strategies that target all members of a specific genogroup. As we continue to expand our understanding of norovirus replication strategies using newly developed in vitro models, additional novel targets for antiviral therapies will undoubtedly be revealed.
Acknowledgments
We sincerely apologize to those whose work was not cited in this review in our effort to concisely summarize a quickly growing field. We thank Christiane E. Wobus (University of Michigan) for critical reading of the manuscript and Mary K. Estes (Baylor College of Medicine) for allowing us to discuss data prior to publication.
The author was supported by Grant Number P20-RR018724 entitled ‘Center for Molecular and Tumor Virology’ from the National Center for Research Resources (NCRR), a component of the National Institutes of Health (NIH), and by the Louisiana Board of Regents through the Board of Regents Support Fund, Grant Number LEQSF(2008-11)-RD-A-16.
Referneces
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.M. Systematic Literature Review of Role of Noroviruses in Sporadic Gastroenteritis. Emerg. Infect. Dis. 2008, 14, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Surveillance for foodborne disease outbreaks - United States. 2006 . MMWR 2009, 58, 609–615. [Google Scholar] [PubMed]
- Green, K.Y. Caliciviridae: The Noroviruses. In Fields Virology; 2007; Lippincott Williams and Wilkins: Philadelphia, PA, USA. [Google Scholar]
- Parrino, T.A.; Schreiber, D.S.; Trier, J.S.; Kapikian, A.Z.; Blacklow, N.R. Clinical immunity in acute gastroenteritis caused by Norwalk agent. N. Engl. J. Med. 1977, 297, 86–89. [Google Scholar] [PubMed]
- Johnson, P.C.; Mathewson, J.J.; DuPont, H.L.; Greenberg, H.B. Multiple-challenge study of host susceptibility to Norwalk gastroenteritis in US adults. J. Infect. Dis. 1990, 161, 18–21. [Google Scholar] [PubMed]
- Estes, M.K.; Prasad, B.V.; Atmar, R.L. Noroviruses everywhere: has something changed? Curr. Opin. Infect. Dis. 2006, 19, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, C.D.and Streitberg. Calicivirus shedding in children after recovery from diarrhoeal disease . J. Clin. Virol. 2008, 43, 346–348. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Katsushima, N.; Mizuta, K.; Muraki, Y.; Hongo, S.; Matsuzaki, Y. Prolonged Norovirus Shedding in Infants - 6 Months of Age With Gastroenteritis. Pediatr. Infect. Dis. J. 2007, 26, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Sakai, Y.; Nakata, S.; Honma, S.; Tatsumi, M.; Numata-Kinoshita, K.; Chiba, S. Clinical severity of Norwalk virus and Sapporo virus gastroenteritis in children in Hokkaido, Japan. Pediatr. Infect. Dis. J. 2001, 20, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Zintz, C.; Bok, K.; Parada, E.; Barnes-Eley, M.; Berke, T.; Staat, M.A.; Azimi, P.; Jiang, X.; Matson, D.O. Prevalence and genetic characterization of caliciviruses among children hospitalized for acute gastroenteritis in the United States. Infect. Genet. Evol. 2005, 5, 281–290. [Google Scholar] [CrossRef]
- Glass, R.I.; Parashar, U.D.; Estes, M.K. Norovirus Gastroenteritis. N. Engl. J. Med. 2009, 361, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Gallimore, C.I.; Lewis, D.; Taylor, C.; Cant, A.; Gennery, A.; Gray, J.J. Chronic excretion of a norovirus in a child with cartilage hair hypoplasia (CHH). J. Clin. Virol. 2004, 30, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Morotti, R.A.; Kaufman, S.S.; Fishbein, T.M.; Chatterjee, N.K.; Fuschino, M.E.; Morse, D.L.; Magid, M.S. Calicivirus infection in pediatric small intestine transplant recipients: Pathological considerations. Hum. Pathol. 2004, 35, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Hedlund, K.; Thorhagen, M.; Larson, G.; Johansen, K.; Ekspong, A.; Svensson, L. Evolution of Human Calicivirus RNA In vivo: Accumulation of Mutations in the Protruding P2 Domain of the Capsid Leads to Structural Changes and Possibly a New Phenotype. J. Virol. 2003, 77, 13117–13124. [Google Scholar] [CrossRef] [PubMed]
- Gallimore, C.I.; Taylor, C.; Gennery, A.R.; Cant, A.J.; Galloway, A.; Iturriza-Gomara, M.; Gray, J.J. Environmental Monitoring for Gastroenteric Viruses in a Pediatric Primary Immunodeficiency Unit. J. Clin. Microbiol. 2006, 44, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, S.S.; Chatterjee, N.K.; Fuschino, M.E.; Magid, M.S.; Gordon, R.E.; Morse, D.L.; Herold, B.C.; LeLeiko, N.S.; Tschernia, A.; Florman, S.S.; Gondolesi, G.E.; Fishbein, T.M. Calicivirus Enteritis in an Intestinal Transplant Recipient. Am. J. Transplant. 2003, 3, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, S.S.; Chatterjee, N.K.; Fuschino, M.E.; Morse, D.L.; Morotti, R.A.; Magid, M.S.; Gondolesi, G.E.; Florman, S.S.; Fishbein, T.M. Characteristics of human calicivirus enteritis in intestinal transplant recipients. J. Pediatr. Gastroenterol. Nutr. 2005, 40, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, T.H.; Vergoulidou, M.; Loddenkemper, C.; Schwartz, S.; Hofmann, J.; Schneider, T.; Zidek, W.; van der Giet, M. Chronic norovirus infection in renal transplant recipients. Nephrol. Dial. Transplant. 2009, 24, 1051–1053. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Norovirus activity--United States, 2006â2007 . MMWR 2007, 56, 842–846. [Google Scholar] [PubMed]
- Lopman, B.A.; Adak, G.K.; Reacher, M.H.; Brown, D.W.G. Two epidemiologic patterns of norovirus outbreaks: surveillance in England and wales, 1992–2000. Emerg. Infect. Dis. 2003, 9, 71–77. [Google Scholar] [PubMed]
- Lopman, B.A.; Reacher, M.H.; Vipond, I.B.; Sarangi, J.; Brown, D.W.G. Clinical Manifestation of Norovirus Gastroenteritis in Health Care Settings. Clin. Infect. Dis. 2004, 39, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Story from REDORBIT NEWS. Norovirus outbreak kills 3 in Washington . 2006. Available online: http://www.redorbit.com/modules/news/tools.php?tool=print&id=462875 .
- Khan, R.R.; Lawson, A.D.; Minnich, L.L.; Martin, K.; Nasir, A.; Emmett, M.K.; Welch, C.A.; Udall, J.N. Gastrointestinal Norovirus Infection Associated With Exacerbation of Inflammatory Bowel Disease. J. Pediatr. Gastroenterol. Nutr. 2009, 48, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, M. Progress in understanding norovirus epidemiology. Curr. Opin. Infect. Dis. 2008, 21, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Takeshita, S.; Nezu, A.; Aihara, Y.; Usuku, S.; Noguchi, Y.; Yokota, S. Norovirus-associated encephalopathy. Pediatr. Infect. Dis. J. 2006, 25, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Outbreak of acute gastroenteritis associated with Norwalk-like viruses among British military personnel--Afghanistan, May 2002 . MMWR 2002, 51, 477–479. [Google Scholar] [PubMed]
- Turcios-Ruiz, R.M.; Axelrod, P.; John, K. St.; Bullitt, E.; Donahue, J.; Robinson, N.; Friss, H.E. Outbreak of Necrotizing Enterocolitis Caused by Norovirus in a Neonatal Intensive Care Unit. J. Pediatr. 2008, 153, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.K.; Thabane, M.; Borgaonkar, M.R.; James, C. Postinfectious Irritable Bowel Syndrome After a Food-Borne Outbreak of Acute Gastroenteritis Attributed to a Viral Pathogen. Clin. Gastroenterol. Hepatol. 2007, 5, 457–460. [Google Scholar] [CrossRef] [PubMed]
- Kawano, G.; Oshige, K.; Syutou, S.; Koteda, Y.; Yokoyama, T.; Kim, B.; Mizuochi, T.; Nagai, K.; Matsuda, K.; Ohbu, K.; Matsuishi, T. Benign infantile convulsions associated with mild gastroenteritis: A retrospective study of 39 cases including virological tests and efficacy of anticonvulsants. Brain Dev. 2007, 29, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tsai, C.; Lai, M.; Chen, C.; Lin, K.; Lin, T.; Chiu, C. Norovirus infection as a cause of diarrhea-associated benign infantile seizures. Clin. Infect. Dis. 2009, 48, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Ando, T.; Fankhauser, R.L.; Beard, R.S.; Glass, R.I.; Monroe, S.S. Norovirus classification and proposed strain nomenclature. Virology 2006, 346, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser, R.L.; Noel, J.S.; Monroe, S.S.; Ando, T.; Glass, R.I. Molecular Epidemiology of “Norwalk‐like Viruses” in Outbreaks of Gastroenteritis in the United States. J. Infect. Dis. 1998, 178, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Sugieda, M.; Nagaoka, H.; Kakishima, Y.; Ohshita, T.; Nakamura, S.; Nakajima, S. Detection of Norwalk-like virus genes in the caecum contents of pigs. Arch. Virol. 1998, 143, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Han, M.G.; Cheetham, S.; Souza, M.; Funk, J.A.; Saif, L.J. Porcine noroviruses related to human noroviruses. Emerg. Infect. Dis. 2005, 11, 1874–1881. [Google Scholar] [PubMed]
- Liu, B.L.; Lambden, P.R.; Gunther, H.; Otto, P.; Elschner, M.; Clarke, I.N. Molecular Characterization of a Bovine Enteric Calicivirus: Relationship to the Norwalk-Like Viruses. J. Virol. 1999, 73, 819–825. [Google Scholar] [PubMed]
- Oliver, S.L.; Dastjerdi, A.M.; Wong, S.; El-Attar, L.; Gallimore, C.; Brown, D.W.G.; Green, J.; Bridger, J.C. Molecular Characterization of Bovine Enteric Caliciviruses: a Distinct Third Genogroup of Noroviruses (Norwalk-Like Viruses) Unlikely To Be of Risk to Humans. J. Virol. 2003, 77, 2789–2798. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Riley, L.; Livingston, R. Molecular characterization of three novel murine noroviruses. Virus Genes. 2007, 34, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Karst, S.M.; Wobus, C.E.; Lay, M.; Davidson, J.; Virgin, H.W. STAT1-Dependent Innate Immunity to a Norwalk-Like Virus. Science 2003, 299, 1575–1578. [Google Scholar] [CrossRef] [PubMed]
- Müller, B.; Klemm, U.; Mas Marques, A.; Schreier, E. Genetic diversity and recombination of murine noroviruses in immunocompromised mice. Arch. Virol. 2007, 152, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Thackray, L.B.; Wobus, C.E.; Chachu, K.A.; Liu, B.; Alegre, E.R.; Henderson, K.S.; Kelley, S.T.; Virgin, H.W. Murine Noroviruses Comprising a Single Genogroup Exhibit Biological Diversity despite Limited Sequence Divergence. J. Virol. 2007, 81, 10460–10473. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Lee, H.; Chang, K.; Ko, G. Molecular Characterization of Murine Norovirus Isolates from South Korea. Virus Res. 2010, 147, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Widdowson, M.; Cramer, E.H.; Hadley, L.; Bresee, J.S.; Beard, R.S.; Bulens, S.N.; Charles, M.; Chege, W.; Isakbaeva, E.; Wright, J.G.; Mintz, E.; Forney, D.; Massey, J.; Glass, R.I.; Monroe, S.S. Outbreaks of Acute Gastroenteritis on Cruise Ships and on Land: Identification of a Predominant Circulating Strain of Norovirus—United States, 2002. J. Infect. Dis. 2004, 190, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Infectious disease and dermatologic conditions in evacuees and rescue workers after Hurricane Katrina--multiple states, August-September, 2005 . MMWR 2005, 54, 961–964. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention. Norovirus outbreak among evacuees from hurricane Katrina-Houston, Texas, September 2005 . MMWR 2005, 54, 1016–1018. [Google Scholar] [PubMed]
- McCarthy, M.; Estes, M.K.; Hyams, K.C. Norwalkâlike Virus Infection in Military Forces: Epidemic Potential, Sporadic Disease, and the Future Direction of Prevention and Control Efforts . J. Infect. Dis. 2000, 181, S387–S391. [Google Scholar] [CrossRef] [PubMed]
- Thornton, S.A.; Sherman, S.S.; Farkas, T.; Zhong, W.; Torres, P.; Jiang, X. Gastroenteritis in US Marines during Operation Iraqi Freedom. Clin. Infect. Dis. 2005, 40, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.; Stamm-Balderjahn, S.; Zuschneid, I.; Behnke, M.; Rüden, H.; Vonberg, R.; Gastmeier, P. Closure of medical departments during nosocomial outbreaks: data from a systematic analysis of the literature. J. Hosp. Infect. 2007, 65, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Rohayem, J. Norovirus seasonality and the potential impact of climate change. Clin. Microbiol. Infect. 2009, 15, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.; Moe, C.L.; Liu, P.; Miller, S.E.; Lindesmith, L.; Baric, R.S.; Pendu, J.L.; Calderon, R.L. Norwalk virus: How infectious is it? J. Med. Virol. 2008, 80, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Outbreaks of gastroenteritis associated with noroviruses on cruise ships--United States, 2002 . MMWR 2002, 51, 1112–1115. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention. Norovirus outbreak associated with ill food-service workers--Michigan, January-February 2006 . MMWR 2006, 56, 1212–1216. [Google Scholar]
- Centers for Disease Control and Prevention. Norovirus outbreak in an elementary school--District of Columbia, February 2007 . MMWR 2008, 56, 1340–1343. [Google Scholar] [PubMed]
- Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Estes, M.K.; Crawford, S.E.; Neill, F.H.; Graham, D.Y. Norwalk virus shedding after experimental human infection. Emerg. Infect. Dis. 2008, 14, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Patterson, T.; Hutchings, P.; Palmer, S. Outbreak of SRSV gastroenteritis at an international conference traced to food handled by a post-symptomatic caterer. Epidemiol. Infect. 1993, 111, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Rockx, B.; de Wit, M.; Vennema, H.; Vinjé, J.; de Bruin, E.; van Duynhoven, Y.; Koopmans, M. Natural History of Human Calicivirus Infection: A Prospective Cohort Study. Clin. Infect. Dis. 2002, 35, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Lopman, B.; Vennema, H.; Kohli, E.; Pothier, P.; Sanchez, A.; Negredo, A.; Buesa, J.; Schreier, E.; Reacher, M.; Brown, D.; Gray, J.; Iturriza, M.; Gallimore, C.; Bottiger, B.; Hedlund, K.; Torvén, M.; von Bonsdorff, C.; Maunula, L.; Poljsak-Prijatelj, M.; Zimsek, J.; Reuter, G.; Szücs, G.; Melegh, B.; Svennson, L.; van Duijnhoven, Y.; Koopmans, M. Increase in viral gastroenteritis outbreaks in Europe and epidemic spread of new norovirus variant. Lancet 2004, 363, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.S.; Fankhauser, R.L.; Ando, T.; Monroe, S.S.; Glass, R.I. Identification of a Distinct Common Strain of "Norwalk‐like Viruses" Having a Global Distribution. J. Infect. Dis. 1999, 179, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- White, P.A.; Hansman, G.S.; Li, A.; Dable, J.; Isaacs, M.; Ferson, M.; McIver, C.J.; Rawlinson, W.D. Norwalk-like virus 95/96-US strain is a major cause of gastroenteritis outbreaks in Australia. J. Med. Virol. 2002, 68, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Bull, R.A.; Tu, E.T.V.; McIver, C.J.; Rawlinson, W.D.; White, P.A. Emergence of a New Norovirus Genotype II.4 Variant Associated with Global Outbreaks of Gastroenteritis . J. Clin. Microbiol. 2006, 44, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Tu, E.T.; Bull, R.A.; Greening, G.E.; Hewitt, J.; Lyon, M.J.; Marshall, J.A.; McIver, C.J.; Rawlinson, W.D.; White, P.A. Epidemics of Gastroenteritis during 2006 Were Associated with the Spread of Norovirus GII.4 Variants 2006a and 2006b . Clin. Infect. Dis. 2008, 46, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Siebenga, J.J.; Vennema, H.; Zheng, D.; Vinjé, J.; Lee, B.E.; Pang, X.; Ho, E.C.M.; Lim, W.; Choudekar, A.; Broor, S.; Halperin, T.; Rasool, N.B.G.; Hewitt, J.; Greening, G.E.; Jin, M.; Duan, Z.; Lucero, Y.; O’Ryan, M.; Hoehne, M.; Schreier, E.; Ratcliff, R.M.; White, P.A.; Iritani, N.; Reuter, G.; Koopmans, G. Norovirus Illness Is a Global Problem: Emergence and Spread of Norovirus GII.4 Variants, 2001â2007 . J. Infect. Dis. 2009, 200, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Bok, K.; Abente, E.J.; Realpe-Quintero, M.; Mitra, T.; Sosnovtsev, S.V.; Kapikian, A.Z.; Green, K.Y. Evolutionary Dynamics of GII.4 Noroviruses over a Thirty-four Year Period . J. Virol. 2009, 83, 11890–11901. [Google Scholar] [PubMed]
- Friesema, I.; Vennema, H.; Heijne, J.; de Jager, C.; Teunis, P.; van der Linde, R.; Duizer, E.; van Duynhoven, Y. Differences in clinical presentation between norovirus genotypes in nursing homes. J. Clin. Virol. 2009, 46, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.M.; Hall, A.J.; Vinjé, J.; Parashar, U.D. Noroviruses: A comprehensive review. J. Clin. Virol. 2009, 44, 1–8. [Google Scholar] [CrossRef]
- Ko, G.; Garcia, C.; Jiang, Z.; Okhuysen, P.C.; Belkind-Gerson, J.; Glass, R.I.; DuPont, H.L. Noroviruses as a Cause of Traveler's Diarrhea among Students from the United States Visiting Mexico. J. Clin. Microbiol. 2005, 43, 6126–6129. [Google Scholar] [CrossRef] [PubMed]
- Chapin, A.R.; Carpenter, C.M.; Dudley, W.C.; Gibson, L.C.; Pratdesaba, R.; Torres, O.; Sanchez, D.; Belkind-Gerson, J.; Nyquist, I.; Karnell, A.; Gustafsson, B.; Halpern, J.L.; Bourgeois, A.L.; Schwab, K.J. Prevalence of Norovirus among Visitors from the United States to Mexico and Guatemala Who Experience Traveler's Diarrhea. J. Clin. Microbiol. 2005, 43, 1112–1117. [Google Scholar] [CrossRef] [PubMed]
- Ko, G.; Jiang, Z.; Okhuysen, P.C.; DuPont, H.L. Fecal cytokines and markers of intestinal inflammation in international travelers with diarrhea due to Noroviruses. J. Med. Virol. 2006, 78, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Acute infectious nonbacterial gastroenteritis: etiology and pathogenesis. Ann. Intern. Med. 1972, 76, 993–1008. [PubMed]
- Dolin, R.; Levy, A.G.; Wyatt, R.G.; Thornhill, T.S.; Gardner, J.D. Viral gastroenteritis induced by the Hawaii agent. Jejunal histopathology and serologic response. Am. J. Med. 1975, 59, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, D.S.; Blacklow, N.R.; Trier, J.S. The mucosal lesion of the proximal small intestine in acute infectious nonbacterial gastroenteritis. N. Engl. J. Med. 1973, 288, 1318–1323. [Google Scholar] [PubMed]
- Schreiber, D.S.; Blacklow, N.R.; Trier, J.S. The small intestinal lesion induced by Hawaii agent acute infectious nonbacterial gastroenteritis. J. Infect. Dis. 1974, 129, 705–708. [Google Scholar] [PubMed]
- Troeger, H.; Loddenkemper, C.; Schneider, T.; Schreier, E.; Epple, H.; Zeitz, M.; Fromm, M.; Schulzke, J. Structural and functional changes of the duodenum in human norovirus infection. Gut 2009, 58, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.; Azevedo, M.S.P.; Jung, K.; Cheetham, S.; Saif, L.J. Pathogenesis and Immune Responses in Gnotobiotic Calves after Infection with the Genogroup II.4-HS66 Strain of Human Norovirus . J. Virol. 2008, 82, 1777–1786. [Google Scholar] [CrossRef] [PubMed]
- Mumphrey, S.M.; Changotra, H.; Moore, T.N.; Heimann-Nichols, E.R.; Wobus, C.E.; Reilly, M.J.; Moghadamfalahi, M.; Shukla, D.; Karst, S.M. Murine Norovirus 1 Infection Is Associated with Histopathological Changes in Immunocompetent Hosts, but Clinical Disease Is Prevented by STAT1-Dependent Interferon Responses. J. Virol. 2007, 81, 3251–3263. [Google Scholar] [CrossRef] [PubMed]
- Cheetham, S.; Souza, M.; Meulia, T.; Grimes, S.; Han, M.G.; Saif, L.J. Pathogenesis of a Genogroup II Human Norovirus in Gnotobiotic Pigs. J. Virol. 2006, 80, 10372–10381. [Google Scholar] [CrossRef] [PubMed]
- Natoni, A.; Kass, G.E.N.; Carter, M.J.; Roberts, L.O. The mitochondrial pathway of apoptosis is triggered during feline calicivirus infection. J. Gen. Virol. 2006, 87, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Bok, K.; Prikhodko, V.G.; Green, K.Y.; Sosnovtsev, S.V. Apoptosis in Murine Norovirus-Infected RAW264.7 Cells Is Associated with Downregulation of Survivin . J. Virol. 2009, 83, 3647–3656. [Google Scholar] [PubMed]
- Roberts, L.O.; Al-Molawi, N.; Carter, M.J.; Kass, G.E.N. Apoptosis in Cultured Cells Infected with Feline Calicivirus. Ann. N. Y. Acad. Sci. 2003, 1010, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Sosnovtsev, S.V.; Prikhod'ko, E.A.; Belliot, G.; Cohen, J.I.; Green, K.Y. Feline calicivirus replication induces apoptosis in cultured cells. Virus Res. 2003, 94, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Furman, L.; Maaty, W.; Petersen, L.; Ettayebi, K.; Hardy, M.; Bothner, B. Cysteine protease activation and apoptosis in Murine norovirus infection. Virol. J. 2009, 6, 139. [Google Scholar] [CrossRef]
- Meeroff, J.C.; Schreiber, D.S.; Trier, J.S.; Blacklow, N.R. Abnormal gastric motor function in viral gastroenteritis. Ann. Intern. Med. 1980, 92, 370–373. [Google Scholar] [PubMed]
- Agus, S.G.; Dolin, R.; Wyatt, R.G.; Tousimis, A.J.; Northrup, R.S. Acute infectious nonbacterial gastroenteritis: intestinal histopathology. Histologic and enzymatic alterations during illness produced by the Norwalk agent in man. Ann. Intern. Medicine 1973, 79, 18–25. [Google Scholar]
- Boshuizen, J.A.; Reimerink, J.H.J.; Korteland-van Male, A.M.; van Ham, V.J.J.; Koopmans, M.P.G.; Buller, H.A.; Dekker, J.; Einerhand, A.W.C. Changes in Small Intestinal Homeostasis, Morphology, and Gene Expression during Rotavirus Infection of Infant Mice. J. Virol. 2003, 77, 13005–13016. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, S.; Hashira, S.; Matsunaga, T.; Yoshida, A.; Shiota, T.; Tung, P.G.; Khamrin, P.; Okitsu, S.; Mizuguchi, M.; Igarashi, T.; Ushijima, H. Detection, genetic characterization, and quantification of norovirus RNA from sera of children with gastroenteritis. J. Clin. Virol. 2009, 44, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Riley, L.K.; Wills, H.M.; Livingston, R.S. Persistent Infection with and Serologic Crossreactivity of Three Novel Murine Noroviruses. Comp. Med. 2006, 56, 247–251. [Google Scholar] [PubMed]
- Hsu, C.C.; Wobus, C.E.; Steffen, C.E.; Riley, L.K.; Livingston, R.S. “Development of a Microsphere-Based Serologic Multiplexed Fluorescent Immunoassay and a Reverse Transcriptase PCR Assay To Detect Murine Norovirus 1 Infection in Mice. Clin. Diagn. Lab. Immunol. 2005, 12, 1145–1151. [Google Scholar] [PubMed]
- Liu, G.; Kahan, S.M.; Jia, Y.; Karst, S.M. Primary High-Dose Murine Norovirus 1 Infection Fails To Protect from Secondary Challenge with Homologous Virus. J. Virol. 2009, 83, 6963–6968. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.M.; Wobus, C.E.; Thackray, L.B.; Erexson, C.R.; Faucette, L.G.; Belliot, G.; Barron, E.L.; Sosnovtsev, S.V.; Green, K.Y. Pathology of Immunodeficient Mice With Naturally Occurring Murine Norovirus Infection. Toxicol. Pathol. 2006, 34, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Blutt, S.E.; Fenaux, M.; Warfield, K.L.; Greenberg, H.B.; Conner, M.E. Active Viremia in Rotavirus-Infected Mice. J. Virol. 2006, 80, 6702–6705. [Google Scholar] [CrossRef] [PubMed]
- Crawford, S.E.; Patel, D.G.; Cheng, E.; Berkova, Z.; Hyser, J.M.; Ciarlet, M.; Finegold, M.J.; Conner, M.E.; Estes, M.K. Rotavirus Viremia and Extraintestinal Viral Infection in the Neonatal Rat Model. J. Virol. 2006, 80, 4820–4832. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, M.; Cuadras, M.A.; Feng, N.; Jaimes, M.; Greenberg, H.B. Extraintestinal Spread and Replication of a Homologous EC Rotavirus Strain and a Heterologous Rhesus Rotavirus in BALB/c Mice. J. Virol. 2006, 80, 5219–5232. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y.; Jiang, X.; Tanaka, T.; Opekun, A.R.; Madore, H.P.; Estes, M.K. Norwalk virus infection of volunteers: new insights based on improved assays. J. Infect. Dis. 1994, 170, 34–43. [Google Scholar] [PubMed]
- Subekti, D.; Tjaniadi, P.; Lesmana, M.; McArdle, J.; Iskandriati, D.; Budiarsa, I.; Walujo, P.; Suparto, I.; Winoto, I.; Campbell, J.; Porter, K.; Sajuthi, D.; Ansari, A.; Oyofo, B. Experimental infection of Macaca nemestrina with a Toronto Norwalk-like virus of epidemic viral gastroenteritis. J. Med. Virol. 2002, 66, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.D.; Dawson, S.; Coyne, K.P.; Porter, C.J.; Gaskell, R.M. The challenge for the next generation of feline calicivirus vaccines. Vet. Microbiol. 2006, 117, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Povey, R.C.; Wardley, R.C.; Jessen, H. Feline picornavirus infection: the in vivo carrier state. Vet. Rec. 1973, 92, 224–229. [Google Scholar] [PubMed]
- Johnson, R.P.; Povey, R.C. Feline calicivirus infection in kittens borne by cats persistently infected with the virus. Res. Vet. Sci. 1984, 37, 114–119. [Google Scholar] [PubMed]
- Oldstone, M.B. Viral persistence: Parameters, mechanisms and future predictions. Virology 2006, 344, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Chachu, K.A.; LoBue, A.D.; Strong, D.W.; Baric, R.S.; Virgin, H.W. Immune Mechanisms Responsible for Vaccination against and Clearance of Mucosal and Lymphatic Norovirus Infection . PLoS Pathog. 2008, 4, e1000236. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.P. Antigenic change in feline calicivirus during persistent infection. Can. J. Vet. Res. 1992, 56, 326–330. [Google Scholar] [PubMed]
- Radford, A.D.; Turner, P.C.; Bennett, M.; McArdle, F.; Dawson, S.; Glenn, M.A.; Williams, R.A.; Gaskell, R.M. Quasispecies evolution of a hypervariable region of the feline calicivirus capsid gene in cell culture and in persistently infected cats. J. Gen. Virol. 1998, 79, 1–10. [Google Scholar] [PubMed]
- Coyne, K.P.; Gaskell, R.M.; Dawson, S.; Porter, C.J.; Radford, A.D. Evolutionary Mechanisms of Persistence and Diversification of a Calicivirus within Endemically Infected Natural Host Populations. J. Virol. 2007, 81, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Wobus, C.E.; Karst, S.M.; Thackray, L.B.; Chang, K.; Sosnovtsev, S.V.; Belliot, G.; Krug, A.; Mackenzie, J.M.; Green, K.Y.; Virgin, H.W. Replication of Norovirus in Cell Culture Reveals a Tropism for Dendritic Cells and Macrophages . PLoS Biol. 2004, 2, e432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duizer, E.; Schwab, K.J.; Neill, F.H.; Atmar, R.L.; Koopmans, M.P.G.; Estes, M.K. Laboratory efforts to cultivate noroviruses. J. Gen. Virol. 2004, 85, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.; Honer zu, B.; Orosz-Coghlan, P.; Dohnalkova, A.; Mayer, B.; Bartholomew, R.; Valdez, C.; Bruckner-Lea, C.; Gerba, C.; Abbaszadegan, M.; Nickerson, C. In vitro cell culture infectivity assay for human noroviruses. Emerg. Infect. Dis. 2007, 13, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Papafragkou, E.; Hewitt, J.; Park, G.; Straub, T.; Greening, G.; Vinje, J. Challenges of Culturing Human Norovirus in a 3-D Organoid Cell Culture Model . In Proceedings of the ASM. 109th General Meeting; Philadephia, PA, USA, 2009; May. [Google Scholar]
- Hao, X.; Kim, T.S.; Braciale, T.J. Differential Response of Respiratory Dendritic Cell Subsets to Influenza Virus Infection. J. Virol. 2008, 82, 4908–4919. [Google Scholar] [CrossRef] [PubMed]
- Henri, S.; Curtis, J.; Hochrein, H.; Vremec, D.; Shortman, K.; Handman, E. Hierarchy of Susceptibility of Dendritic Cell Subsets to Infection by Leishmania major: Inverse Relationship to Interleukin-12 Production. Infect. Immun. 2002, 70, 3874–3880. [Google Scholar] [CrossRef] [PubMed]
- Latham, P.S.; Sepelak, S.B. Effect of macrophage source and activation on susceptibility in an age-dependent model of murine hepatitis caused by a phlebovirus, Punta Toro. Arch. Virol. 1992, 122, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Guix, S.; Asanaka, M.; Katayama, K.; Crawford, S.E.; Neill, F.H.; Atmar, R.L.; Estes, M.K. Norwalk Virus RNA Is Infectious in Mammalian Cells. J. Virol. 2007, 81, 12238–12248. [Google Scholar] [CrossRef] [PubMed]
- Asanaka, M.; Atmar, R.L.; Ruvolo, V.; Crawford, S.E.; Neill, F.H.; Estes, M.K. Replication and packaging of Norwalk virus RNA in cultured mammalian cells. Proc. Natl. Acad. Sci. USA 2005, 102, 10327–10332. [Google Scholar] [CrossRef]
- Chaudhry, Y.; Skinner, M.A.; Goodfellow, I.G. Recovery of genetically defined murine norovirus in tissue culture by using a fowlpox virus expressing T7 RNA polymerase. J. Gen. Virol. 2007, 88, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Katayama, K.; Hansman, G.S.; Oka, T.; Ogawa, S.; Takeda, N. Investigation of norovirus replication in a human cell line. Arch.Virol. 2006, 151, 1291–1308. [Google Scholar] [CrossRef] [PubMed]
- Ward, V.K.; McCormick, C.J.; Clarke, I.N.; Salim, O.; Wobus, C.E.; Thackray, L.B.; Virgin, H.W.; Lambden, P.R. Recovery of infectious murine norovirus using pol II-driven expression of full-length cDNA. Proc. Natl. Acad. Sci. USA 2007, 104, 11050–11055. [Google Scholar] [CrossRef]
- Chang, K.; Sosnovtsev, S.V.; Belliot, G.; King, A.D.; Green, K.Y. Stable expression of a Norwalk virus RNA replicon in a human hepatoma cell line. Virology 2006, 353, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Wobus, C.E.; Thackray, L.B.; Virgin, H.W. Murine Norovirus: a Model System To Study Norovirus Biology and Pathogenesis. J. Virol. 2006, 80, 5104–5112. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.W.; Taube, S.; Wobus, C.E. Murine norovirus-1 entry into permissive macrophages and dendritic cells is pH-independent. Virus Res. 2009, 143, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Perdue, K.A.; Green, K.Y.; Copeland, M.; Barron, E.; Mandel, M.; Faucette, L.J.; Williams, E.M.; Sosnovtsev, S.V.; Elkins, W.R.; Ward, J.M. Naturally Occurring Murine Norovirus Infection in a Large Research Institution. J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 39–45. [Google Scholar] [PubMed]
- Ryman, K.D.; Klimstra, W.B.; Nguyen, K.B.; Biron, C.A.; Johnston, R.E. Alpha/Beta Interferon Protects Adult Mice from Fatal Sindbis Virus Infection and Is an Important Determinant of Cell and Tissue Tropism. J. Virol. 2000, 74, 3366–3378. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Diamond, M.S. Alpha/Beta Interferon Protects against Lethal West Nile Virus Infection by Restricting Cellular Tropism and Enhancing Neuronal Survival. J. Virol. 2005, 79, 13350–13361. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ma, Y.; Barrett, J.W.; Gao, X.; Loh, J.; Barton, E.; Virgin, H.W.; McFadden, G. Disruption of Erk-dependent type I interferon induction breaks the myxoma virus species barrier. Nat. Immunol. 2004, 5, 1266–1274. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Jiang, X. Norovirus and its histo-blood group antigen receptors: an answer to a historical puzzle. Trends Microbiol. 2005, 13, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.; Moe, C.; Marionneau, S.; Ruvoen, N.; Jiang, X.; Lindblad, L.; Stewart, P.; LePendu, J.; Baric, R. Human susceptibility and resistance to Norwalk virus infection. Nat. Med. 2003, 9, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Farkas, T.; Marionneau, S.; Zhong, W.; Ruvoën‐Clouet, N.; Morrow, A.L.; Altaye, M.; Pickering, L.K.; Newburg, D.S.; LePendu, J.; Jiang, X. Noroviruses Bind to Human ABO, Lewis, and Secretor Histo–Blood Group Antigens: Identification of 4 Distinct Strain‐Specific Patterns. J. Infect. Dis. 2003, 188, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Farkas, T.; Zhong, W.; Tan, M.; Thornton, S.; Morrow, A.L.; Jiang, X. Norovirus and Histo-Blood Group Antigens: Demonstration of a Wide Spectrum of Strain Specificities and Classification of Two Major Binding Groups among Multiple Binding Patterns. J. Virol. 2005, 79, 6714–6722. [Google Scholar] [CrossRef] [PubMed]
- Shirato, H.; Ogawa, S.; Ito, H.; Sato, T.; Kameyama, A.; Narimatsu, H.; Xiaofan, Z.; Miyamura, T.; Wakita, T.; Ishii, K.; Takeda, N. Noroviruses Distinguish between Type 1 and Type 2 Histo-Blood Group Antigens for Binding. J. Virol. 2008, 82, 10756–10767. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Xia, M.; Chen, Y.; Bu, W.; Hegde, R.S.; Meller, J.; Li, X.; Jiang, X. Conservation of Carbohydrate Binding Interfaces â Evidence of Human HBGA Selection in Norovirus Evolution . PLoS ONE 2009, 4, e5058. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.C.; Donaldson, E.F.; LoBue, A.D.; Cannon, J.L.; Zheng, D.; Vinje, J.; Baric, R.S. Mechanisms of GII.4 Norovirus Persistence in Human Populations . PLoS Med. 2008, 5, e31. [Google Scholar] [CrossRef] [PubMed]
- Cannon, J.L.; Lindesmith, L.C.; Donaldson, E.F.; Saxe, L.; Baric, R.S.; Vinje, J. Herd Immunity to GII.4 Noroviruses Is Supported by Outbreak Patient Sera . J. Virol. 2009, 83, 5363–5374. [Google Scholar] [CrossRef] [PubMed]
- Le Pendu, J.; Ruvoën-Clouet, N.; Kindberg, E.; Svensson, L. Mendelian resistance to human norovirus infections. Sem. Immunol. 2006, 18, 375–386. [Google Scholar] [CrossRef]
- Ruvoen-Clouet, N.; Ganiere, J.P.; Andre-Fontaine, G.; Blanchard, D.; Le Pendu, J. Binding of Rabbit Hemorrhagic Disease Virus to Antigens of the ABH Histo-Blood Group Family. J. Virol. 2000, 74, 11950–11954. [Google Scholar] [CrossRef] [PubMed]
- Farkas, T.; Sestak, K.; Wei, C.; Jiang, X. Characterization of a Rhesus Monkey Calicivirus Representing a New Genus of Caliciviridae. J. Virol. 2008, 82, 5408–5416. [Google Scholar] [CrossRef] [PubMed]
- Stuart, A.D.; Brown, T.D.K. {alpha}2,6-Linked sialic acid acts as a receptor for Feline calicivirus. J. Gen. Virol. 2007, 88, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Taube, S.; Perry, J.W.; Yetming, K.; Patel, S.P.; Auble, H.; Shu, L.; Nawar, H.F.; Lee, C.H.; Connell, T.D.; Shayman, J.A.; Wobus, C.E. Ganglioside-Linked Terminal Sialic Acid Moieties on Murine Macrophages Function as Attachment Receptors for Murine Noroviruses. J. Virol. 2009, 83, 4092–4101. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Natori, K.; Kobayashi, M.; Miyamura, T.; Takeda, N. Genogroup II Noroviruses Efficiently Bind to Heparan Sulfate Proteoglycan Associated with the Cellular Membrane. J. Virol. 2004, 78, 3817–3826. [Google Scholar] [CrossRef] [PubMed]
- Rydell, G.E.; Nilsson, J.; Rodriguez-Diaz, J.; Ruvoen-Clouet, N.; Svensson, L.; Le Pendu, J.; Larson, G. Human noroviruses recognize sialyl Lewis x neoglycoprotein. Glycobiology 2009, 19, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Makino, A.; Shimojima, M.; Miyazawa, T.; Kato, K.; Tohya, Y.; Akashi, H. Junctional Adhesion Molecule 1 Is a Functional Receptor for Feline Calicivirus. J. Virol. 2006, 80, 4482–4490. [Google Scholar] [CrossRef] [PubMed]
- Glass, P.J.; White, L.J.; Ball, J.M.; Leparc-Goffart, I.; Hardy, M.E.; Estes, M.K. Norwalk Virus Open Reading Frame 3 Encodes a Minor Structural Protein. J. Virol. 2000, 74, 6581–6591. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, M.; Wang, K.; Estes, M.K. Sequence and Genomic Organization of Norwalk Virus. Virology 1993, 195, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lambden, P.; Caul, E.; Ashley, C.; Clarke, I. Sequence and genome organization of a human small round-structured (Norwalk-like) virus. Science 1993, 259, 516–519. [Google Scholar] [PubMed]
- Crowder, S.; Kirkegaard, K. Trans-dominant inhibition of RNA viral replication can slow growth of drug-resistant viruses. Nat. Genet. 2005, 37, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, P.; Karakasiliotis, I.; Bailey, D.; Chaudhry, Y.; Evans, D.J.; Goodfellow, I.G. Bioinformatic and functional analysis of RNA secondary structure elements among different genera of human and animal caliciviruses. Nucleic Acids Res. 2008, 36, 2530–2546. [Google Scholar] [CrossRef] [PubMed]
- Victoria, M.; Colina, R.; Miagostovich, M.; Leite, J.; Cristina, J. Phylogenetic prediction of cis-acting elements: a cre-like sequence in Norovirus genome? BMC Res. Notes 2009, 2, 176. [Google Scholar] [CrossRef]
- Sosnovtsev, S.V.; Belliot, G.; Chang, K.; Prikhodko, V.G.; Thackray, L.B.; Wobus, C.E.; Karst, S.M.; Virgin, H.W.; Green, K.Y. Cleavage Map and Proteolytic Processing of the Murine Norovirus Nonstructural Polyprotein in Infected Cells. J. Virol. 2006, 80, 7816–7831. [Google Scholar] [CrossRef] [PubMed]
- Belliot, G.; Sosnovtsev, S.V.; Mitra, T.; Hammer, C.; Garfield, M.; Green, K.Y. In vitro Proteolytic Processing of the MD145 Norovirus ORF1 Nonstructural Polyprotein Yields Stable Precursors and Products Similar to Those Detected in Calicivirus-Infected Cells. J. Virol. 2003, 77, 10957–10974. [Google Scholar] [CrossRef] [PubMed]
- Seah, E.L.; Marshall, J.A.; Wright, P.J. trans Activity of the Norovirus Camberwell Proteinase and Cleavage of the N-Terminal Protein Encoded by ORF1. J. Virol. 2003, 77, 7150–7155. [Google Scholar] [CrossRef] [PubMed]
- Pfister, T.; Wimmer, E. Polypeptide p41 of a Norwalk-Like Virus Is a Nucleic Acid-Independent Nucleoside Triphosphatase. J. Virol. 2001, 75, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Clarke, I.; Lambden, P. Polyprotein processing in Southampton virus: identification of 3C-like protease cleavage sites by in vitro mutagenesis. J. Virol. 1996, 70, 2605–2610. [Google Scholar] [PubMed]
- Fukushi, S.; Kojima, S.; Takai, R.; Hoshino, F.B.; Oka, T.; Takeda, N.; Katayama, K.; Kageyama, T. Poly(A)- and Primer-Independent RNA Polymerase of Norovirus. J. Virol. 2004, 78, 3889–3896. [Google Scholar] [CrossRef] [PubMed]
- Rohayem, J.; Robel, I.; Jager, K.; Scheffler, U.; Rudolph, W. Protein-Primed and De Novo Initiation of RNA Synthesis by Norovirus 3Dpol. J. Virol. 2006, 80, 7060–7069. [Google Scholar] [CrossRef] [PubMed]
- Burroughs, J.N.; Brown, F. Presence of a covalently linked protein on calicivirus RNA. J. Gen. Virol. 1978, 41, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Herbert, T.P.; Brierley, I.; Brown, T.D. Identification of a protein linked to the genomic and subgenomic mRNAs of feline calicivirus and its role in translation. J. Gen. Virol. 1997, 78, 1033–1040. [Google Scholar] [PubMed]
- Schaffer, F.L.; Ehresmann, D.W.; Fretz, M.K.; Soergel, M.I. A protein, VPg, covalently linked to 36S calicivirus RNA. J. Gen. Virol. 1980, 47, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Belliot, G.; Sosnovtsev, S.V.; Chang, K.; McPhie, P.; Green, K.Y. Nucleotidylylation of the VPg Protein of a Human Norovirus by its Proteinase-Polymerase Precursor Protein. Virology 2008, 374, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, Y.; Nayak, A.; Bordeleau, M.; Tanaka, J.; Pelletier, J.; Belsham, G.J.; Roberts, L.O.; Goodfellow, I.G. Caliciviruses Differ in Their Functional Requirements for eIF4F Components. J. Biol. Chem. 2006, 281, 25315–25325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daughenbaugh, K.F.; Fraser, C.S.; Hershey, J.W.; Hardy, M.E. The genome-linked protein VPg of the Norwalk virus binds eIF3, suggesting its role in translation initiation complex recruitment. EMBO J. 2003, 22, 2852–2859. [Google Scholar] [CrossRef] [PubMed]
- Daughenbaugh, K.; Wobus, C.; Hardy, M. VPg of murine norovirus binds translation initiation factors in infected cells. Virol. J. 2006, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Vega, V.; Sosnovtsev, S.V.; Belliot, G.; King, A.D.; Mitra, T.; Gorbalenya, A.; Green, K.Y. Norwalk Virus N-Terminal Nonstructural Protein Is Associated with Disassembly of the Golgi Complex in Transfected Cells. J. Virol. 2004, 78, 4827–4837. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Hardy, M.E. Norwalk Virus Nonstructural Protein p48 Forms a Complex with the SNARE Regulator VAP-A and Prevents Cell Surface Expression of Vesicular Stomatitis Virus G Protein. J. Virol. 2003, 77, 11790–11797. [Google Scholar] [CrossRef] [PubMed]
- Hyde, J.L.; Sosnovtsev, S.V.; Green, K.Y.; Wobus, C.; Virgin, H.W.; Mackenzie, J.M. Mouse Norovirus Replication Is Associated with Virus-Induced Vesicle Clusters Originating from Membranes Derived from the Secretory Pathway. J. Virol. 2009, 83, 9709–9719. [Google Scholar] [CrossRef] [PubMed]
- Changotra, H.; Jia, Y.; Moore, T.N.; Liu, G.; Kahan, S.M.; Sosnovtsev, S.V.; Karst, S.M. Type I and Type II Interferons Inhibit the Translation of Murine Norovirus Proteins. J. Virol. 2009, 83, 5683–5692. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.; George, D.W. Interferons and Ribavirin Effectively Inhibit Norwalk Virus Replication in Replicon-Bearing Cells. J. Virol. 2007, 81, 12111–12118. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Neill, J.D.; Noel, J.S.; Hutson, A.M.; Glass, R.I.; Estes, M.K.; Prasad, B.V.V. Inter- and Intragenus Structural Variations in Caliciviruses and Their Functional Implications. J. Virol. 2004, 78, 6469–6479. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.V.; Rothnagel, R.; Jiang, X.; Estes, M.K. Three-dimensional structure of baculovirus-expressed Norwalk virus capsids. J. Virol. 1994, 68, 5117–5125. [Google Scholar] [PubMed]
- Prasad, B.V.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray Crystallographic Structure of the Norwalk Virus Capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Prasad, B.V.; Crawford, S.; Lawton, J.A.; Pesavento, J.; Hardy, M.; Estes, M.K. Structural studies on gastroenteritis viruses . Novartis Found. Symp. 2001, 238 discussion 37â46, 26–37. [Google Scholar] [PubMed]
- Katpally, U.; Wobus, C.E.; Dryden, K.; Virgin, H.W.; Smith, T.J. Structure of Antibody-Neutralized Murine Norovirus and Unexpected Differences from Viruslike Particles. J. Virol. 2008, 82, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, M.; Graham, D.Y.; Estes, M.K. Expression, self-assembly, and antigenicity of the Norwalk virus capsid protein. J. Virol. 1992, 66, 6527–6532. [Google Scholar] [PubMed]
- Tan, M.; Fang, P.; Chachiyo, T.; Xia, M.; Huang, P.; Fang, Z.; Jiang, W.; Jiang, X. Noroviral P particle: Structure, function and applications in virus-host interaction. Virology 2008, 382, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wilton, N.; Zhong, W.M.; Farkas, T.; Huang, P.W.; Barrett, E.; Guerrero, M.; Ruiz‐Palacios, G.; Green, K.Y.; Green, J.; Hale, A.D.; Estes, M.K.; Pickering, L.K.; Matson, D.O. Diagnosis of Human Caliciviruses by Use of Enzyme Immunoassays . J. Infect. Dis. 2000, 181, S349–S359. [Google Scholar] [CrossRef] [PubMed]
- Bu, W.; Mamedova, A.; Tan, M.; Xia, M.; Jiang, X.; Hegde, R.S. Structural Basis for the Receptor Binding Specificity of Norwalk Virus. J. Virol. 2008, 82, 5340–5347. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Lou, Z.; Tan, M.; Chen, Y.; Liu, Y.; Zhang, Z.; Zhang, X.C.; Jiang, X.; Li, X.; Rao, Z. Structural Basis for the Recognition of Blood Group Trisaccharides by Norovirus. J. Virol. 2007, 81, 5949–5957. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Hutson, A.M.; Estes, M.K.; Prasad, B.V.V. Atomic resolution structural characterization of recognition of histo-blood group antigens by Norwalk virus. Proc. Natl. Acad. Sci. USA 2008, 105, 9175–9180. [Google Scholar] [CrossRef]
- Tan, M.; Huang, P.; Meller, J.; Zhong, W.; Farkas, T.; Jiang, X. Mutations within the P2 Domain of Norovirus Capsid Affect Binding to Human Histo-Blood Group Antigens: Evidence for a Binding Pocket. J. Virol. 2003, 77, 12562–12571. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Xia, M.; Cao, S.; Huang, P.; Farkas, T.; Meller, J.; Hegde, R.S.; Li, X.; Rao, Z.; Jiang, X. Elucidation of strain-specific interaction of a GII-4 norovirus with HBGA receptors by site-directed mutagenesis study. Virology 2008, 379, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Lochridge, V.P.; Jutila, K.L.; Graff, J.W.; Hardy, M.E. Epitopes in the P2 domain of norovirus VP1 recognized by monoclonal antibodies that block cell interactions. J. Gen. Virol. 2005, 86, 2799–2806. [Google Scholar] [CrossRef] [PubMed]
- Allen, D.J.; Noad, R.; Samuel, D.; Gray, J.J.; Roy, P.; Iturriza-Gomara, M. Characterisation of a GII-4 norovirus variant-specific surface-exposed site involved in antibody binding. Virol. J. 2009, 6, 150. [Google Scholar] [CrossRef]
- Bailey, D.; Thackray, L.B.; Goodfellow, I.G. A Single Amino Acid Substitution in the Murine Norovirus Capsid Protein Is Sufficient for Attenuation In vivo. J. Virol. 2008, 82, 7725–7728. [Google Scholar] [CrossRef] [PubMed]
- Glass, P.J.; Zeng, C.Q.; Estes, M.K. Two Nonoverlapping Domains on the Norwalk Virus Open Reading Frame 3 (ORF3) Protein Are Involved in the Formation of the Phosphorylated 35K Protein and in ORF3-Capsid Protein Interactions. J. Virol. 2003, 77, 3569–3577. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti-Ciarlet, A.; Crawford, S.E.; Hutson, A.M.; Estes, M.K. The 3' End of Norwalk Virus mRNA Contains Determinants That Regulate the Expression and Stability of the Viral Capsid Protein VP1: a Novel Function for the VP2 Protein. J. Virol. 2003, 77, 11603–11615. [Google Scholar] [CrossRef] [PubMed]
- Luttermann, C.; Meyers, G. A Bipartite Sequence Motif Induces Translation Reinitiation in Feline Calicivirus RNA. J. Biol. Chem. 2007, 282, 7056–7065. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G. Characterization of the Sequence Element Directing Translation Reinitiation in RNA of the Calicivirus Rabbit Hemorrhagic Disease Virus. J. Virol. 2007, 81, 9623–9632. [Google Scholar] [CrossRef] [PubMed]
- Meyers, G. Translation of the Minor Capsid Protein of a Calicivirus Is Initiated by a Novel Termination-dependent Reinitiation Mechanism. J. Biol. Chem. 2003, 278, 34051–34060. [Google Scholar] [CrossRef] [PubMed]
- Luttermann, C.; Meyers, G. The importance of inter- and intramolecular base pairing for translation reinitiation on a eukaryotic bicistronic mRNA. Genes Develop. 2009, 23, 331–344. [Google Scholar] [CrossRef]
- McCormick, C.J.; Salim, O.; Lambden, P.R.; Clarke, I.N. Translation Termination Reinitiation between Open Reading Frame 1 (ORF1) and ORF2 Enables Capsid Expression in a Bovine Norovirus without the Need for Production of Viral Subgenomic RNA. J. Virol. 2008, 82, 8917–8921. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.C.; Greenberg, H.B.; Cukor, G.; Blacklow, N.R. Serological responses among teenagers after natural exposure to Norwalk virus. J. Infect. Dis. 1984, 150, 531–534. [Google Scholar] [PubMed]
- Blacklow, N.R.; Cukor, G.; Bedigian, M.K.; Echeverria, P.; Greenberg, H.B.; Schreiber, D.S.; Trier, J.S. Immune response and prevalence of antibody to Norwalk enteritis virus as determined by radioimmunoassay. J. Clin. Microbiol. 1979, 10, 903–909. [Google Scholar] [PubMed]
- Cukor, G.; Nowak, N.A.; Blacklow, N.R. Immunoglobulin M responses to the Norwalk virus of gastroenteritis. Infect. Immun. 1982, 37, 463–468. [Google Scholar] [PubMed]
- Johnson, P.C.; Hoy, J.; Mathewson, J.J.; Ericsson, C.D.; DuPont, H.L. Occurrence of Norwalk virus infections among adults in Mexico. J. Infect. Dis. 1990, 162, 389–393. [Google Scholar] [PubMed]
- Agus, S.G.; Falchuk, Z.M.; Sessoms, C.S.; Wyatt, R.G.; Dolin, R. Increased jejunal IgA synthesis in vitro during acute infectious nonbacterial gastroenteritis. Am. J. Dig. Dis. 1974, 19, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Harrington, P.R.; Lindesmith, L.; Yount, B.; Moe, C.L.; Baric, R.S. Binding of Norwalk Virus-Like Particles to ABH Histo-Blood Group Antigens Is Blocked by Antisera from Infected Human Volunteers or Experimentally Vaccinated Mice. J. Virol. 2002, 76, 12335–12343. [Google Scholar] [CrossRef] [PubMed]
- Rockx, B.; Baric, R.S.; Grijs, I.D.; Duizer, E.; Koopmans, M.P. Characterization of the homo- and heterotypic immune responses after natural norovirus infection. J. Med. Virol. 2005, 77, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.; Moe, C.; LePendu, J.; Frelinger, J.A.; Treanor, J.; Baric, R.S. Cellular and Humoral Immunity following Snow Mountain Virus Challenge. J. Virol. 2005, 79, 2900–2909. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.; Cheetham, S.M.; Azevedo, M.S.P.; Costantini, V.; Saif, L.J. Cytokine and Antibody Responses in Gnotobiotic Pigs after Infection with Human Norovirus Genogroup II.4 (HS66 Strain) . J. Virol. 2007, 81, 9183–9192. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, E.F.; Lindesmith, L.C.; Lobue, A.D.; Baric, R.S. Norovirus pathogenesis: mechanisms of persistence and immune evasion in human populations. Immunol. Rev. 2008, 225, 190–211. [Google Scholar] [CrossRef] [PubMed]
- Han, M.G.; Cheetham, S.; Azevedo, M.; Thomas, C.; Saif, L.J. Immune responses to bovine norovirus-like particles with various adjuvants and analysis of protection in gnotobiotic calves. Vaccine. 2006, 24, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.; Costantini, V.; Azevedo, M.S.; Saif, L.J. A human norovirus-like particle vaccine adjuvanted with ISCOM or mLT induces cytokine and antibody responses and protection to the homologous GII.4 human norovirus in a gnotobiotic pig disease model . Vaccine 2007, 25, 8448–8459. [Google Scholar] [CrossRef] [PubMed]
- LoBue, A.D.; Thompson, J.M.; Lindesmith, L.; Johnston, R.E.; Baric, R.S. Alphavirus-Adjuvanted Norovirus-Like Particle Vaccines: Heterologous, Humoral, and Mucosal Immune Responses Protect against Murine Norovirus Challenge. J. Virol. 2009, 83, 3212–3227. [Google Scholar] [CrossRef] [PubMed]
- Mackinnon, M.J.; Read, A.F. Immunity Promotes Virulence Evolution in a Malaria Model . PLoS Biol. 2004, 2, e230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurley, K.E.; Pesavento, P.A.; Pedersen, N.C.; Poland, A.M.; Wilson, E.; Foley, J.E. An outbreak of virulent systemic feline calicivirus disease. J. Am. Vet. Med. Assoc. 2004, 224, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Elliott, J.B.; Glasgow, A.; Poland, A.; Keel, K. An isolated epizootic of hemorrhagic-like fever in cats caused by a novel and highly virulent strain of feline calicivirus. Vet. Microbiol. 2000, 73, 281–300. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
MDPI and ACS Style
Karst, S.M. Pathogenesis of Noroviruses, Emerging RNA Viruses. Viruses 2010, 2, 748-781. https://doi.org/10.3390/v2030748
AMA Style
Karst SM. Pathogenesis of Noroviruses, Emerging RNA Viruses. Viruses. 2010; 2(3):748-781. https://doi.org/10.3390/v2030748
Chicago/Turabian StyleKarst, Stephanie M. 2010. "Pathogenesis of Noroviruses, Emerging RNA Viruses" Viruses 2, no. 3: 748-781. https://doi.org/10.3390/v2030748