Beclin-1 Targeting for Viral Immune Escape


Viral Immunobiology, Institute of Experimental Immunology, University of Zürich, Winterthurerstrasse 190, CH-8057 Zürich, Switzerland
Viruses 2011, 3(7), 1166-1178; https://doi.org/10.3390/v3071166
Submission received: 23 June 2011
/
Revised: 4 July 2011
/
Accepted: 5 July 2011
/
Published: 12 July 2011
(This article belongs to the Special Issue Autophagy and Viruses)
Abstract
:Macroautophagy is a catabolic pathway in eukaryotic cells that has recently been shown to facilitate pathogen detection, pathogen restriction and pathogen-derived antigen presentation to CD4+ T cells. Due to these protective functions during immune responses, several pathogens, including RNA and DNA viruses, have developed strategies to inhibit autophagosome generation or maturation. Interestingly, most of the respective viral proteins exert these functions via binding to Beclin-1, an essential macroautophagy protein that constitutes part of the phosphatidylinositol-3 kinase complexes that mark membranes for autophagosome generation and facilitate autophagosome fusion with lyososomes. The viruses that inhibit macroautophagy by this pathway include herpesviruses, HIV and influenza A virus. Inhibition either before or after autophagosome formation seems to benefit their viral replication by different mechanisms, which are discussed here.
1. Introduction
Lysosomes and proteasomes are the two main catabolic machineries in eukaryotic cells. Endocytosed, but also autophagocytosed material can reach lysosomes, thereby delivering extra- or intracellular material for degradation, respectively. At least three pathways of autophagy exist for the delivery of cytoplasmic constituents for lysosomal degradation. These are macro-, micro- and chaperone-mediated autophagy [1]. While micro- and chaperone-mediated autophagy occur directly at the lysosomal membrane, macroautophagy can originate at the rough endoplasmic reticulum, Golgi apparatus, outer nuclear or mitochondrial membrane, and the cell membrane [2,3,4,5,6,7,8,9,10]. During microautophagy a part of the lysosomal membrane invaginates into the lumen of these degradative vesicles and thereby delivers cytosolic substrates for lysosomal hydrolysis. In higher eukaryotes this seems to occur with the help of the chaperone HSC70 [11]. Chaperone-mediated autophagy, however, has been observed in higher eukaryotic cells only after prolonged starvation. Under these conditions, substrates with a signal motif, a permutation of the pentamer KFERQ, get transported across the lysosomal membrane with the help of the lysosome associated membrane protein (LAMP) 2a, cytosolic chaperones and luminal chaperones in the lysosomes. Signal peptide recognition seems to occur via the cytosolic chaperone, and therefore similar substrates are selected for microautophagy and chaperone mediated autophagy in higher eukaryotic cells [11]. Finally, most steady-state turn-over of cytosolic protein aggregates and organelles in lysosomes of higher eukaryotes seems to require macroautophagy for delivering these substrates. Macroautophagy is executed through the interaction of more than 30 autophagy related gene (atg) products, whose loss prevents yeast from surviving starvation conditions via this pathway. During this process a double membrane, called a phagophore, engulfs substrates, anchoring the inner autophagosomal membrane to the cargo via Atg8/LC3, which is covalently attached to phosphatidylethanolamine in the autophagosomal membrane via an ubiquitin-like conjugation reaction involving the E1-, E2- and E3-like enzymes Atg7, Atg3 and Atg12-Atg5/Atg16L1 [12,13]. Atg8/LC3 is either directly bound by a cargo protein, like the outer mitochondrial membrane protein NIX for mitophagy [14,15], or by anchor proteins, like p62/sequestosome 1, optineurin, NBR1 or NDP52 [16,17,18,19,20], that cross-link Atg8/LC3 with polyubiquitinated substrates. The completed autophagosome then fuses with late endosomes or directly with lysosomes, whereby the inner membrane and its cargo get degraded by lysosomal hydrolysis. Autophagosome generation and degradation is regulated by class III phosphatidylinositol-3-kinase (PI3K) complexes including Atg6/Beclin-1. Therefore, viruses often interfere with these complexes via targeting Beclin-1 in order to arrest autophagosome formation or maturation.
2. Beclin-1 Containing Complexes and Their Role in Macroautophagy
During autophagosome initiation Atg6/Beclin-1 is associated with the PI3K VPS34 and the adaptor proteins myristoylated VPS15 for membrane anchoring and Atg14L (Figure 1) [21,22,23]. This complex marks membranes for phagophore formation with PI3P, for example downstream of the starvation induced ULK complexes [24]. For incorporation into this complex Atg6/Beclin-1 has to dissociate from the anti-apoptotic Bcl-2 and Bcl-xL proteins, which anchor Atg6/Beclin-1 at the endoplasmic reticulum membrane. The Beclin-1 interacting protein AMBRA1 disrupts this interaction and thereby activates macroautophagy [25]. Vice versa, macroautophagy is compromised by mono-allelic deletion of Beclin-1 in 40–75% of ovarian and breast carcinomas [26,27]. These factors influence the generation of autophagosomes. In addition, the maturation, namely fusion of autophagosomes with lysosomes, is influenced by Beclin-1 containing VPS34 complexes. Instead of Atg14L, however, these contain the UV irradiation resistance-associated gene (UVRAG). This protein enhances autophagosome maturation, but again a negative regulator exists in the form of the Rubicon protein, which associates with the UVRAG containing Beclin-1 complex and arrests autophagosome maturation [21,22]. Thus, higher eukaryotic cells carry sophisticated molecular machineries for the regulation of PI3P deposition at future or formed autophagosomal membranes to react to metabolic needs, but also infections by regulating macroautophagy.
3. Macroautophagy in Pathogen Sensing
Pathogen sensing is the first step during the initiation of immune responses. Pathogen-associated molecular pattern (PAMP) receptors detect motifs in viruses, bacteria, fungi and parasites [28]. This recognition alarms the immune system by cytokine secretion and activation of antigen presenting cells. Different pattern recognition receptor (PRR) families monitor extracellular and intracellular space. RIG-I like (RLRs) and NOD like receptors (NLRs, including inflammasomes) detect cytosolic RNA and DNA, crystals or other danger molecules, respectively. In contrast, Toll-like (TLRs) and C-type lectin like receptors (CLRs) detect extracellular pathogen products. Interestingly, some TLRs signal from vesicular compartments and access to these endosomal TLRs (TLR3, 7, 8 and 9) can be regulated by vesicular transport. Along these lines it has been demonstrated that replication intermediates of Sendai and Vesicular Stomatitis virus require macroautophagy to be sensed by TLR7/8 [29]. Transport of these viral RNAs into TLR7/8 containing endosomes by autophagosomes was required for IFNα production by plasmacytoid dendritic cells (DCs) in response to infection. Similarly, macroautophagy has been shown to facilitate B cell receptor ligand transport to TLR9 containing endosomes for B cell activation [30]. Thus, PAMP sensing by vesicular TLRs of antigen presenting cells seems to benefit from macroautophagic transport of ligands to endosomes.
In contrast, cytosolic PRRs might be negatively regulated by macroautophagy. In the absence of Atg5, a component of the Atg8/LC3 E3-like ligase, dsRNA elicits increased IFNα production via RIG-I recognition in mouse embryonic fibroblasts [31]. Deficiency of macroautophagy leads to enhanced reactive oxygen species (ROS) production by dysfunctional mitochondria, which directly stimulates RLR activation [32]. ROS production does not only augment RLR, but also NLR signaling. Indeed Atg16L1 deficient macrophages produce elevated levels of inflammasome dependent IL-1 [33]. This elevated inflammasome activity is dependent on mitochondrial ROS, and inflammasomes and ROS contribute to mitochondrial DNA release into the cytosol, which augments IL-1 production [34,35]. Thus, PAMP recognition by vesicular TLRs of antigen presenting cells is supported by macroautophagy, while the same process dampens cytosolic PAMP recognition in somatic cells. This could benefit the initiation of immune responses by focusing activation towards antigen presenting cells, like DCs and B cells.
4. Contribution of Macroautophagy to Innate and Adaptive Immunity
During such immune responses macroautophagy restricts intracellular pathogens and allows for antigen presentation to the adaptive immune system. Indeed during the engulfment of group A Streptococci autophagosomes can reach ten-fold larger diameters (10 μm) than usually observed for metabolic autophagosomes (0.5–1 μm in diameter) [36]. Macroautophagy restricts these bacteria, which escape from endosomes to replicate in the cytosol. Interestingly, the formation of these large Streptococci containing autophagosomes requires Rab7, a GTPase that has previously been described to mediate autophagosome fusion to lysosomes [37]. In addition to bacteria, parasites like Toxoplasma gondii seems to be targeted by macroautophagy [38,39]. Toxoplasma conditions phagosomes to prevent their fusion with lysosomes, which allows the parasite to replicate in these vesicular compartments. The protective phagosome membrane, however, can be stripped by immunity related GTPase M (IRGM), which then exposes Toxoplasma to macroautophagy, leading to degradation in lysosomes [39]. Similar mechanisms might be at work during the restriction of Mycobacterium tuberculosis growth inhibition by macroautophagy [40,41]. Finally, neurotrophic Sindbis virus infection is also compromised by macroautophagy [42,43]. Enhancement of macroautophagy by Atg6/Beclin-1 overexpression [42] compromises Sindbis virus replication, while deficiency of Atg5 in neurons increases immunopathology of Sindbis virus infection in the central nervous system of mice [43]. While the detailed mechanism of viral restriction is not known so far, it might involve direct degradation of viral particles. In addition to simply using lysosomal hydrolysis for pathogen destruction, autophagosomes also deliver substrates to lysosomes, from which antibacterial peptides can be generated. These include ubiquitin [44] and the ubiquitin-domain containing ribosomal protein Fau [45]. Thus, macroautophagy can restrict bacteria, parasites and viruses during immune responses.
As a byproduct of this, pathogen degradation fragments are recycled for antigen presentation on MHC molecules to T cells. Especially, antigen processing for MHC class II presentation to CD4+ helper T cells utilizes lysosomal hydrolysis. Accordingly, intracellular antigen processing of some viral and bacterial antigens utilizes macroautophagy [46,47,48,49]. In good agreement with these examples of antigen processing by macroautophagy, autophagosomes have been found to frequently fuse with MHC class II loading compartments [50], and enhanced autophagosome formation due to starvation changes the repertoire of MHC class II presented peptides to contain more frequently ligands, which are derived from cytosolic and nuclear proteins [51]. This alteration of the MHC class II presented ligandome by macroautophagy leads also to deficient positive and negative T cell selection in the thymus [52]. Furthermore, even though mechanistically not very well understood so far, Atg5 deficiency in DCs compromises the development of CD4+ T cell responses upon herpes simplex virus (HSV) infection of mice [53], and loss of an HSV encoded macroautophagy inhibitor augments CD4+ T cell responses during infection [54]. These studies suggest that degradation of pathogens via macroautophagy leads to MHC class II presentation of their fragments for stimulation of CD4+ T cell responses.
5. Inhibition of Autophagosome Generation by Viral Proteins Interacting with Beclin-1
However, viruses counteract these beneficial functions of macroautophagy for immune responses. They have developed strategies to both target autophagosome generation and autophagosome maturation. A rich source of inhibitors of autophagosome generation is the herpesvirus family (Figure 2 and Table 1) [55]. In fact, all three herpesvirus subfamilies encode inhibitors of macroautophagy, which primarily target the Atg14L containing Beclin-1 complex of VPS34 and VPS15. The α-herpesvirus HSV-1 encodes the ICP34.5 protein, which interacts with Beclin-1 via a 20 amino acid region (aa 68-87) and thereby compromises autophagosome generation [56]. HSV encoding ICP34.5, in which this region has been deleted, demonstrates attenuated neurovirulence in mice. In correlation with this attenuated neurovirulence, autophagosome accumulation can be observed in mutant virus infected neurons, and loss of the double-strand RNA-dependent kinase PKR, which stimulates macroautophagy in HSV infected cells, eliminates the protective function of elevated macroautophagy against mutant HSV. Without ICP34.5, HSV particles can be found in autophagosomes by electron microscopy [57,58]. In addition, loss of ICP34.5 also elevates HSV specific CD4+ T cell responses during infection [54]. Besides HSV, γ-herpesviruses also encode Beclin-1 interacting proteins. Kaposi sarcoma associated herpesvirus (KSHV) encodes a viral Bcl-2 homologue with its open reading frame (orf) 16. This binds Atg6/Beclin-1 with higher affinity than cellular Bcl-2 [59]. Binding of viral Bcl-2 inhibits macroautophagy by interrupting Beclin-1 incorporation in VPS34 complexes, probably via anchoring Beclin-1 at endoplasmic reticulum membranes [60]. Similarly, the viral Bcl-2 homologue M11 of the murine γ-herpesvirus 68 (MHV-68) interacts with Beclin-1 and inhibits autophagosome formation [61]. It binds with higher affinity to Beclin-1 than cellular Bcl-2 and Bcl-xL. Three amino acids at positions 85-87 are required for Beclin-1 binding by M11, and a C-terminally, but not N-terminally truncated molecule can still interact with Beclin-1 [62]. Interestingly, recombinant viruses lacking either the N-terminal α1 helix or with mutated amino acids 85-87 no longer inhibited macroautophagy upon infection, and loss of macroautophagy inhibition compromised persistence of MHV-68 in vivo [62]. Thus, herpesviruses sequester Beclin-1 to inhibit autophagosome formation to enhance their neurovirulence and increase their persistence in vivo.
6. Immune Escape from Autophagosome Maturation via Targeting of Beclin-1
In contrast to the DNA viruses of the herpesvirus family that inhibit autophagosome generation, RNA viruses seem to stabilize autophagosomes by preventing their degradation. Indeed, both infections with the hepatitis C virus and poliovirus accumulate autophagosomes, and benefit for their replication from this macroautophagy regulation [63,64,65,66]. However, for these and most other viral infections, this autophagosome accumulation is mechanistically not well understood. However, in two instances, influenza and human immunodeficiency virus (HIV) infection, some insights into mechanisms of autophagosome accumulation have been gained, and both pathogens target Beclin-1, presumably stabilizing complexes of Beclin-1, VPS34, VPS15, UVRAG and Rubicon that inhibit autophagosome maturation (Figure 2 and Table 1). Influenza A virus infection stabilizes autophagosomes, which fuse to large perinuclear vesicles and contain viral proteins, but not any lysosomal markers [67]. Matrix protein 2 (M2) of influenza A virus can induce this autophagosome accumulation, is localized to these accumulating autophagosomes, and M2 deficient virus does no longer establish this block of macroautophagy. Interestingly, the main function of M2, its proton channel activity that is required for viral core acidification during infection, does not seem to be involved in autophagosome stabilization. Instead full-length and a truncated M2 version (aa 1-60) are able to interact with Beclin-1. Inhibition of autophagosome maturation compromised survival of influenza A virus infected cells, thereby enhancing the pro-apoptotic effect of the viral protein PB-F1 by blocking the pro-survival pathway macroautophagy. Under certain infection conditions, this regulation of macroautophagy might also benefit viral replication [68]. Furthermore, consistent with inhibition of autophagosome maturation, influenza A virus antigens are not processed intracellularly for MHC class II presentation to CD4+ T cells [69]. Instead, proteasomal degradation facilitates intracellular influenza antigen processing for MHC class II presentation. This suggests that proteasomal degradation takes over when macroautophagy is blocked, and vice versa an increase in macroautophagy can be detected in cells treated with proteasomal inhibitors [66,70]. Interestingly, autophagosome generation facilitates antigen transfer from influenza A virus infected cells to neighboring DCs [71]. It is tempting to speculate that exocytosis of viral protein carrying autophagosomes, which accumulate due to the M2 inducted block of their fusion with lysosomes, might contribute to this cross-presentation. Similar to influenza A virus, HIV blocks autophagosome maturation in infected macrophages [72]. HIV gag proteins accumulate in the stabilized autophagosomes and this autophagosome accumulation enhances viral replication in macrophages. The virus achieves this autophagosome accumulation via its nef protein, which co-localizes with autophagosomes and binds to Beclin-1 via its DD motif (aa 174-175) and this motif is required for autophagosome accumulation by nef. This fusion block is also established in DCs upon HIV infection, and prevents the formation of so-called immunoamphisomes [73]. These studies suggest that HIV and influenza A virus block autophagosome maturation by encoding Beclin-1 interacting proteins.

{kind=link}
{kind=link}
Table 1.
Viral proteins that interact with Atg6/Beclin-1.
| Virus | Protein | Function | Reference |
|---|---|---|---|
| HSV-1 | ICP34.5 | blocks autophagosome generation | [54] |
| KSHV | orf16 | blocks autophagosome generation | [58] |
| MHV-68 | M11 | blocks autophagosome generation | [59,60] |
| HIV | nef | blocks autophagosome maturation | [70] |
| Influenza A | M2 | blocks autophagosome maturation | [65] |
7. Conclusions
Macroautophagy, a pathway that has been originally appreciated for its pro-survival function in nutrient recycling during starvation, has in recent years been recognized for its functions during immune responses. Both pathogen sensing by innate and adaptive immune compartment, as well as pathogen destruction through this pathway are supported by macroautophagy. Accordingly, successful viral pathogens have learned to interfere with this pathway for their own benefit. While herpesviruses inhibit autophagosome generation, many RNA viruses, including influenza virus and HIV, block autophagosome maturation and degradation, but seem to benefit from stabilization of these vesicles. Interestingly, both inhibition of autophagosome generation and degradation seems to be often achieved through viral protein binding to Beclin-1, probably regulating complexes of this essential macroautophagy protein that are either involved in phagophore generation or autophagosome fusion with lysosomes. How this is achieved by the different viral proteins remains to be seen. However, these viral modulators of macroautophagy already now give us tools to experimentally regulate this catabolic pathway, and interference with their function might hopefully suggest some therapeutic interventions against the encoding viruses in the future.
Conflict of Interest
The author declares no conflict of interest.
Acknowledgments
Work in my laboratory is in part supported by the National Cancer Institute (R01CA108609), Cancer Research Switzerland (KFS-02652-08-2010), the Association for International Cancer Research (11-0516), the Sassella Foundation (10/02), the Vontobel Foundation and the Swiss National Science Foundation (310030_126995).
References and Notes
- Mizushima, N.; Klionsky, D.J. Protein turnover via autophagy: Implications for metabolism. Annu. Rev. Nutr. 2007, 27, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Hayashi-Nishino, M.; Fujita, N.; Noda, T.; Yamaguchi, A.; Yoshimori, T.; Yamamoto, A. A subdomain of the endoplasmic reticulum forms a cradle for autophagosome formation. Nat. Cell Biol. 2009, 11, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Yla-Anttila, P.; Vihinen, H.; Jokitalo, E.; Eskelinen, E.L. 3D tomography reveals connections between the phagophore and endoplasmic reticulum. Autophagy 2009, 5, 1180–1185. [Google Scholar] [CrossRef]
- Yen, W.L.; Shintani, T.; Nair, U.; Cao, Y.; Richardson, B.C.; Li, Z.; Hughson, F.M.; Baba, M.; Klionsky, D.J. The conserved oligomeric Golgi complex is involved in double-membrane vesicle formation during autophagy. J. Cell Biol. 2010, 188, 101–114. [Google Scholar] [CrossRef]
- Lynch-Day, M.A.; Bhandari, D.; Menon, S.; Huang, J.; Cai, H.; Bartholomew, C.R.; Brumell, J.H.; Ferro-Novick, S.; Klionsky, D.J. Trs85 directs a Ypt1 GEF, TRAPPIII, to the phagophore to promote autophagy. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 7811–7816. [Google Scholar] [CrossRef]
- English, L.; Chemali, M.; Duron, J.; Rondeau, C.; Laplante, A.; Gingras, D.; Alexander, D.; Leib, D.; Norbury, C.; Lippe, R.; et al. Autophagy enhances the presentation of endogenous viral antigens on MHC class I molecules during HSV-1 infection. Nat. Immunol. 2009, 10, 480–487. [Google Scholar] [CrossRef]
- Hailey, D.W.; Rambold, A.S.; Satpute-Krishnan, P.; Mitra, K.; Sougrat, R.; Kim, P.K.; Lippincott-Schwartz, J. Mitochondria supply membranes for autophagosome biogenesis during starvation. Cell 2010, 141, 656–667. [Google Scholar] [CrossRef]
- He, C.; Song, H.; Yorimitsu, T.; Monastyrska, I.; Yen, W.L.; Legakis, J.E.; Klionsky, D.J. Recruitment of Atg9 to the preautophagosomal structure by Atg11 is essential for selective autophagy in budding yeast. J. Cell Biol. 2006, 175, 925–935. [Google Scholar] [CrossRef]
- Ravikumar, B.; Moreau, K.; Jahreiss, L.; Puri, C.; Rubinsztein, D.C. Plasma membrane contributes to the formation of pre-autophagosomal structures. Nat. Cell Biol. 2010, 12, 747–757. [Google Scholar] [CrossRef]
- Sahu, R.; Kaushik, S.; Clement, C.C.; Cannizzo, E.S.; Scharf, B.; Follenzi, A.; Potolicchio, I.; Nieves, E.; Cuervo, A.M.; Santambrogio, L. Microautophagy of cytosolic proteins by late endosomes. Dev. Cell 2011, 20, 131–139. [Google Scholar] [CrossRef]
- Hanada, T.; Noda, N.N.; Satomi, Y.; Ichimura, Y.; Fujioka, Y.; Takao, T.; Inagaki, F.; Ohsumi, Y. The Atg12-Atg5 conjugate has a novel E3-like activity for protein lipidation in autophagy. J. Biol. Chem. 2007, 282, 37298–37302. [Google Scholar] [CrossRef]
- Fujita, N.; Itoh, T.; Omori, H.; Fukuda, M.; Noda, T.; Yoshimori, T. The Atg16L complex specifies the site of LC3 lipidation for membrane biogenesis in autophagy. Mol. Biol. Cell 2008, 19, 2092–2100. [Google Scholar] [CrossRef]
- Sandoval, H.; Thiagarajan, P.; Dasgupta, S.K.; Schumacher, A.; Prchal, J.T.; Chen, M.; Wang, J. Essential role for Nix in autophagic maturation of erythroid cells. Nature 2008, 454, 232–235. [Google Scholar] [CrossRef]
- Schweers, R.L.; Zhang, J.; Randall, M.S.; Loyd, M.R.; Li, W.; Dorsey, F.C.; Kundu, M.; Opferman, J.T.; Cleveland, J.L.; Miller, J.L.; et al. NIX is required for programmed mitochondrial clearance during reticulocyte maturation. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 19500–19505. [Google Scholar] [CrossRef]
- Bjorkoy, G.; Lamark, T.; Brech, A.; Outzen, H.; Perander, M.; Overvatn, A.; Stenmark, H.; Johansen, T. p62/SQSTM1 forms protein aggregates degraded by autophagy and has a protective effect on huntingtin-induced cell death. J. Cell Biol. 2005, 171, 603–614. [Google Scholar] [CrossRef]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.A.; Outzen, H.; Overvatn, A.; Bjorkoy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef]
- Kirkin, V.; Lamark, T.; Sou, Y.S.; Bjorkoy, G.; Nunn, J.L.; Bruun, J.A.; Shvets, E.; McEwan, D.G.; Clausen, T.H.; Wild, P.; et al. A role for NBR1 in autophagosomal degradation of ubiquitinated substrates. Mol. Cell 2009, 33, 505–516. [Google Scholar] [CrossRef]
- Thurston, T.L.; Ryzhakov, G.; Bloor, S.; von Muhlinen, N.; Randow, F. The TBK1 adaptor and autophagy receptor NDP52 restricts the proliferation of ubiquitin-coated bacteria. Nat. Immunol. 2009, 10, 1215–1221. [Google Scholar] [CrossRef]
- Wild, P.; Farhan, H.; McEwan, D.G.; Wagner, S.; Rogov, V.V.; Brady, N.R.; Richter, B.; Korac, J.; Waidmann, O.; Choudhary, C.; et al. Phosphorylation of the Autophagy Receptor Optineurin Restricts Salmonella Growth. Science 2011. [Google Scholar] [CrossRef]
- Matsunaga, K.; Saitoh, T.; Tabata, K.; Omori, H.; Satoh, T.; Kurotori, N.; Maejima, I.; Shirahama-Noda, K.; Ichimura, T.; Isobe, T.; et al. Two Beclin 1-binding proteins, Atg14L and Rubicon, reciprocally regulate autophagy at different stages. Nat. Cell Biol. 2009, 11, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Wang, Q.J.; Li, X.; Yan, Y.; Backer, J.M.; Chait, B.T.; Heintz, N.; Yue, Z. Distinct regulation of autophagic activity by Atg14L and Rubicon associated with Beclin 1-phosphatidylinositol-3-kinase complex. Nat. Cell Biol. 2009, 11, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Itakura, E.; Kishi, C.; Inoue, K.; Mizushima, N. Beclin 1 forms two distinct phosphatidylinositol 3-kinase complexes with mammalian Atg14 and UVRAG. Mol. Biol. Cell 2008, 19, 5360–5372. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nature 2011, 469, 323–335. [Google Scholar] [CrossRef]
- Strappazzon, F.; Vietri-Rudan, M.; Campello, S.; Nazio, F.; Florenzano, F.; Fimia, G.M.; Piacentini, M.; Levine, B.; Cecconi, F. Mitochondrial BCL-2 inhibits AMBRA1-induced autophagy. EMBO J. 2011, 30, 1195–1208. [Google Scholar] [CrossRef]
- Liang, X.H.; Jackson, S.; Seaman, M.; Brown, K.; Kempkes, B.; Hibshoosh, H.; Levine, B. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature 1999, 402, 672–676. [Google Scholar] [CrossRef]
- Aita, V.M.; Liang, X.H.; Murty, V.V.; Pincus, D.L.; Yu, W.; Cayanis, E.; Kalachikov, S.; Gilliam, T.C.; Levine, B. Cloning and genomic organization of beclin 1, a candidate tumor suppressor gene on chromosome 17q21. Genomics 1999, 59, 59–65. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Lee, H.K.; Lund, J.M.; Ramanathan, B.; Mizushima, N.; Iwasaki, A. Autophagy-dependent viral recognition by plasmacytoid dendritic cells. Science 2007, 315, 1398–1401. [Google Scholar] [CrossRef]
- Chaturvedi, A.; Dorward, D.; Pierce, S.K. The B cell receptor governs the subcellular location of Toll-like receptor 9 leading to hyperresponses to DNA-containing antigens. Immunity 2008, 28, 799–809. [Google Scholar] [CrossRef]
- Jounai, N.; Takeshita, F.; Kobiyama, K.; Sawano, A.; Miyawaki, A.; Xin, K.Q.; Ishii, K.J.; Kawai, T.; Akira, S.; Suzuki, K.; et al. The Atg5 Atg12 conjugate associates with innate antiviral immune responses. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 14050–14055. [Google Scholar] [CrossRef]
- Tal, M.C.; Sasai, M.; Lee, H.K.; Yordy, B.; Shadel, G.S.; Iwasaki, A. Absence of autophagy results in reactive oxygen species-dependent amplification of RLR signaling. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 2770–2775. [Google Scholar] [CrossRef]
- Saitoh, T.; Fujita, N.; Jang, M.H.; Uematsu, S.; Yang, B.G.; Satoh, T.; Omori, H.; Noda, T.; Yamamoto, N.; Komatsu, M.; et al. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1beta production. Nature 2008, 456, 264–268. [Google Scholar] [CrossRef]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef]
- Nakahira, K.; Haspel, J.A.; Rathinam, V.A.; Lee, S.J.; Dolinay, T.; Lam, H.C.; Englert, J.A.; Rabinovitch, M.; Cernadas, M.; Kim, H.P.; et al. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol. 2011, 12, 222–230. [Google Scholar] [CrossRef]
- Nakagawa, I.; Amano, A.; Mizushima, N.; Yamamoto, A.; Yamaguchi, H.; Kamimoto, T.; Nara, A.; Funao, J.; Nakata, M.; Tsuda, K.; et al. Autophagy defends cells against invading group A Streptococcus. Science 2004, 306, 1037–1040. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Nakagawa, I.; Yamamoto, A.; Amano, A.; Noda, T.; Yoshimori, T. An initial step of GAS-containing autophagosome-like vacuoles formation requires Rab7. PLoS Pathog. 2009, 5, e1000670. [Google Scholar] [CrossRef]
- Andrade, R.M.; Wessendarp, M.; Gubbels, M.J.; Striepen, B.; Subauste, C.S. CD40 induces macrophage anti-Toxoplasma gondii activity by triggering autophagy-dependent fusion of pathogen-containing vacuoles and lysosomes. J. Clin. Invest. 2006, 116, 2366–2377. [Google Scholar] [CrossRef]
- Ling, Y.M.; Shaw, M.H.; Ayala, C.; Coppens, I.; Taylor, G.A.; Ferguson, D.J.; Yap, G.S. Vacuolar and plasma membrane stripping and autophagic elimination of Toxoplasma gondii in primed effector macrophages. J. Exp. Med. 2006, 203, 2063–2071. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy is a defense mechanism inhibiting BCG and mycobacterium tuberculosis survival in infected macrophages. Cell 2004, 119, 753–766. [Google Scholar] [CrossRef]
- Singh, S.B.; Davis, A.S.; Taylor, G.A.; Deretic, V. Human IRGM induces autophagy to eliminate intracellular mycobacteria. Science 2006, 313, 1438–1441. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.H.; Kleeman, L.K.; Jiang, H.H.; Gordon, G.; Goldman, J.E.; Berry, G.; Herman, B.; Levine, B. Protection against fatal Sindbis virus encephalitis by beclin, a novel Bcl-2-interacting protein. J. Virol. 1998, 72, 8586–8596. [Google Scholar] [CrossRef] [PubMed]
- Orvedahl, A.; MacPherson, S.; Sumpter, R., Jr.; Talloczy, Z.; Zou, Z.; Levine, B. Autophagy protects against Sindbis virus infection of the central nervous system. Cell Host Microbe 2010, 7, 115–127. [Google Scholar] [CrossRef]
- Alonso, S.; Pethe, K.; Russell, D.G.; Purdy, G.E. Lysosomal killing of Mycobacterium mediated by ubiquitin-derived peptides is enhanced by autophagy. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 6031–6036. [Google Scholar] [CrossRef] [PubMed]
- Ponpuak, M.; Davis, A.S.; Roberts, E.A.; Delgado, M.A.; Dinkins, C.; Zhao, Z.; Virgin, H.W.t.; Kyei, G.B.; Johansen, T.; Vergne, I.; et al. Delivery of cytosolic components by autophagic adaptor protein p62 endows autophagosomes with unique antimicrobial properties. Immunity 2010, 32, 329–341. [Google Scholar] [CrossRef]
- Paludan, C.; Schmid, D.; Landthaler, M.; Vockerodt, M.; Kube, D.; Tuschl, T.; Münz, C. Endogenous MHC class II processing of a viral nuclear antigen after autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef]
- Leung, C.S.; Haigh, T.A.; Mackay, L.K.; Rickinson, A.B.; Taylor, G.S. Nuclear location of an endogenously expressed antigen, EBNA1, restricts access to macroautophagy and the range of CD4 epitope display. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 2165–2170. [Google Scholar] [CrossRef]
- Jagannath, C.; Lindsey, D.R.; Dhandayuthapani, S.; Xu, Y.; Hunter, R.L., Jr.; Eissa, N.T. Autophagy enhances the efficacy of BCG vaccine by increasing peptide presentation in mouse dendritic cells. Nat. Med. 2009, 15, 267–276. [Google Scholar] [CrossRef]
- Russmann, H.; Panthel, K.; Kohn, B.; Jellbauer, S.; Winter, S.E.; Garbom, S.; Wolf-Watz, H.; Hoffmann, S.; Grauling-Halama, S.; Geginat, G. Alternative endogenous protein processing via an autophagy-dependent pathway compensates for Yersinia-mediated inhibition of endosomal major histocompatibility complex class II antigen presentation. Infect. Immun. 2010, 78, 5138–5150. [Google Scholar] [CrossRef]
- Schmid, D.; Pypaert, M.; Münz, C. MHC class II antigen loading compartments continuously receive input from autophagosomes. Immunity 2007, 26, 79–92. [Google Scholar] [CrossRef]
- Dengjel, J.; Schoor, O.; Fischer, R.; Reich, M.; Kraus, M.; Muller, M.; Kreymborg, K.; Altenberend, F.; Brandenburg, J.; Kalbacher, H.; et al. Autophagy promotes MHC class II presentation of peptides from intracellular source proteins. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 7922–7927. [Google Scholar] [CrossRef]
- Nedjic, J.; Aichinger, M.; Emmerich, J.; Mizushima, N.; Klein, L. Autophagy in thymic epithelium shapes the T-cell repertoire and is essential for tolerance. Nature 2008, 455, 396–400. [Google Scholar] [CrossRef]
- Lee, H.K.; Mattei, L.M.; Steinberg, B.E.; Alberts, P.; Lee, Y.H.; Chervonsky, A.; Mizushima, N.; Grinstein, S.; Iwasaki, A. In vivo requirement for Atg5 in antigen presentation by dendritic cells. Immunity 2010, 32, 227–239. [Google Scholar] [CrossRef]
- Leib, D.A.; Alexander, D.E.; Cox, D.; Yin, J.; Ferguson, T.A. Interaction of ICP34.5 with Beclin 1 modulates herpes simplex virus type 1 pathogenesis through control of CD4+ T-cell responses. J. Virol. 2009, 83, 12164–12171. [Google Scholar] [CrossRef]
- Taylor, G.S.; Mautner, J.; Münz, C. Autophagy in herpesvirus immune control and immune escape. Herpesviridae 2011, 2, 2. [Google Scholar] [CrossRef]
- Orvedahl, A.; Alexander, D.; Talloczy, Z.; Sun, Q.; Wei, Y.; Zhang, W.; Burns, D.; Leib, D.; Levine, B. HSV-1 ICP34.5 confers neurovirulence by targeting the Beclin 1 autophagy protein. Cell Host Microbe 2007, 1, 23–35. [Google Scholar] [CrossRef]
- Levine, B.; Deretic, V. Unveiling the roles of autophagy in innate and adaptive immunity. Nat. Rev. Immunol. 2007, 7, 767–777. [Google Scholar] [CrossRef]
- Talloczy, Z.; Virgin, H.W.t.; Levine, B. PKR-dependent autophagic degradation of herpes simplex virus type 1. Autophagy 2006, 2, 24–29. [Google Scholar] [CrossRef]
- Liang, C.; E, X.; Jung, J.U. Downregulation of autophagy by herpesvirus Bcl-2 homologs. Autophagy 2008, 4, 268–272. [Google Scholar] [CrossRef]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.H.; Mizushima, N.; Packer, M.; Schneider, M.D.; Levine, B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 2005, 122, 927–939. [Google Scholar] [CrossRef]
- Ku, B.; Woo, J.S.; Liang, C.; Lee, K.H.; Hong, H.S.; E, X.; Kim, K.S.; Jung, J.U.; Oh, B.H. Structural and biochemical bases for the inhibition of autophagy and apoptosis by viral BCL-2 of murine gamma-herpesvirus 68. PLoS Pathog. 2008, 4, e25. [Google Scholar] [CrossRef] [PubMed]
- E, X.; Hwang, S.; Oh, S.; Lee, J.S.; Jeong, J.H.; Gwack, Y.; Kowalik, T.F.; Sun, R.; Jung, J.U.; Liang, C. Viral Bcl-2-mediated evasion of autophagy aids chronic infection of gammaherpesvirus 68. PLoS Pathog. 2009, 5, e1000609. [Google Scholar] [CrossRef] [PubMed]
- Dales, S.; Eggers, H.J.; Tamm, I.; Palade, G.E. Electron microscopic study of the formation of poliovirus. Virology 1965, 26, 379–389. [Google Scholar] [CrossRef]
- Jackson, W.T.; Giddings, T.H., Jr.; Taylor, M.P.; Mulinyawe, S.; Rabinovitch, M.; Kopito, R.R.; Kirkegaard, K. Subversion of cellular autophagosomal machinery by RNA viruses. PLoS Biol. 2005, 3, e156. [Google Scholar] [CrossRef] [PubMed]
- Ait-Goughoulte, M.; Kanda, T.; Meyer, K.; Ryerse, J.S.; Ray, R.B.; Ray, R. Hepatitis C virus genotype 1a growth and induction of autophagy. J. Virol. 2008, 82, 2241–2249. [Google Scholar] [CrossRef]
- Dreux, M.; Gastaminza, P.; Wieland, S.F.; Chisari, F.V. The autophagy machinery is required to initiate hepatitis C virus replication. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 14046–14051. [Google Scholar] [CrossRef]
- Gannage, M.; Dormann, D.; Albrecht, R.; Dengjel, J.; Torossi, T.; Ramer, P.C.; Lee, M.; Strowig, T.; Arrey, F.; Conenello, G.; et al. Matrix protein 2 of influenza A virus blocks autophagosome fusion with lysosomes. Cell Host Microbe 2009, 6, 367–380. [Google Scholar] [CrossRef]
- Zhou, Z.; Jiang, X.; Liu, D.; Fan, Z.; Hu, X.; Yan, J.; Wang, M.; Gao, G.F. Autophagy is involved in influenza A virus replication. Autophagy 2009, 5, 321–328. [Google Scholar] [CrossRef]
- Comber, J.D.; Robinson, T.M.; Siciliano, N.A.; Snook, A.E.; Eisenlohr, L.C. Functional macroautophagy induction by influenza A virus without a contribution to MHC-class II Restricted presentation. J. Virol. 2011, 85, 6453–6463. [Google Scholar] [CrossRef]
- Zhu, K.; Dunner, K., Jr.; McConkey, D.J. Proteasome inhibitors activate autophagy as a cytoprotective response in human prostate cancer cells. Oncogene 2010, 29, 451–462. [Google Scholar] [CrossRef]
- Uhl, M.; Kepp, O.; Jusforgues-Saklani, H.; Vicencio, J.M.; Kroemer, G.; Albert, M.L. Autophagy within the antigen donor cell facilitates efficient antigen cross-priming of virus-specific CD8+ T cells. Cell Death Differ. 2009, 16, 991–1005. [Google Scholar] [CrossRef]
- Kyei, G.B.; Dinkins, C.; Davis, A.S.; Roberts, E.; Singh, S.B.; Dong, C.; Wu, L.; Kominami, E.; Ueno, T.; Yamamoto, A.; et al. Autophagy pathway intersects with HIV-1 biosynthesis and regulates viral yields in macrophages. J. Cell Biol. 2009, 186, 255–268. [Google Scholar] [CrossRef]
- Blanchet, F.P.; Moris, A.; Nikolic, D.S.; Lehmann, M.; Cardinaud, S.; Stalder, R.; Garcia, E.; Dinkins, C.; Leuba, F.; Wu, L.; et al. Human immunodeficiency virus-1 inhibition of immunoamphisomes in dendritic cells impairs early innate and adaptive immune responses. Immunity 2010, 32, 654–669. [Google Scholar] [CrossRef]
Figure 1.
Atg6/Beclin-1 containing PI3 kinase complexes catalyze autophagosome generation and fusion with lysosomes. The Beclin-1 containing complex with Atg14L, VPS34 and VPS15 marks membranes at which the phagophore forms, which expands around its substrates with the help of the two ubiquitin-like molecules Atg8 and 12 and upon closure constitutes double-membrane surrounded autophagosomes. Their fusion with lysosomes is assisted by the UVRAG, VPS34 and VPS15 containing Beclin-1 complex. Recruitment of Rubicon into this complex blocks autophagosome fusion with lysosomes.
Figure 1.
Atg6/Beclin-1 containing PI3 kinase complexes catalyze autophagosome generation and fusion with lysosomes. The Beclin-1 containing complex with Atg14L, VPS34 and VPS15 marks membranes at which the phagophore forms, which expands around its substrates with the help of the two ubiquitin-like molecules Atg8 and 12 and upon closure constitutes double-membrane surrounded autophagosomes. Their fusion with lysosomes is assisted by the UVRAG, VPS34 and VPS15 containing Beclin-1 complex. Recruitment of Rubicon into this complex blocks autophagosome fusion with lysosomes.

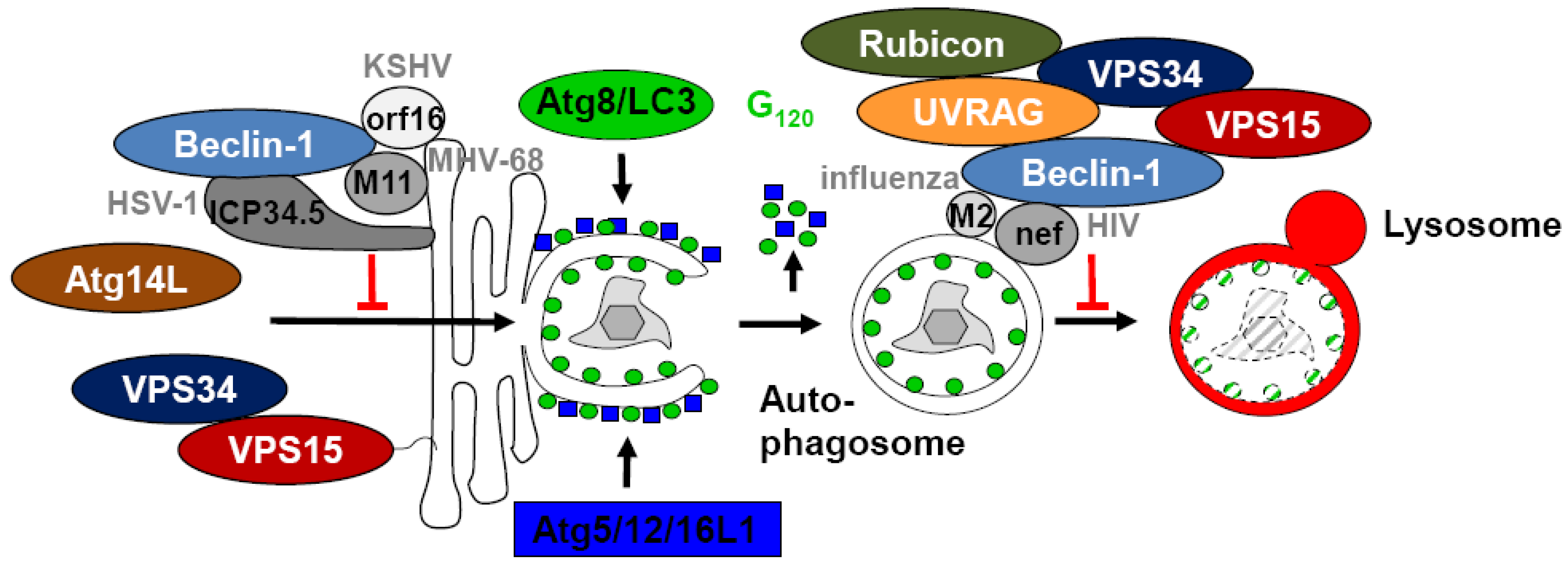
Figure 2.
Viral proteins block autophagosome generation and degradation via Beclin-1 binding. KSHV orf16, MHV-68 M11 and HSV-1 ICP34.5 anchor Beclin-1 at membranes and block its incorporation into the PI3 kinase complex with VPS34, VPS15 and Atg14L, which catalyzes autophagosome generation. Furthermore influenza A virus M2 and HIV nef presumably stabilize through their Beclin-1 binding inhibitory PI3 kinase complexes that prevent fusion between autophagosomes and lysosomes.
Figure 2.
Viral proteins block autophagosome generation and degradation via Beclin-1 binding. KSHV orf16, MHV-68 M11 and HSV-1 ICP34.5 anchor Beclin-1 at membranes and block its incorporation into the PI3 kinase complex with VPS34, VPS15 and Atg14L, which catalyzes autophagosome generation. Furthermore influenza A virus M2 and HIV nef presumably stabilize through their Beclin-1 binding inhibitory PI3 kinase complexes that prevent fusion between autophagosomes and lysosomes.

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Münz, C. Beclin-1 Targeting for Viral Immune Escape. Viruses 2011, 3, 1166-1178. https://doi.org/10.3390/v3071166
AMA Style
Münz C. Beclin-1 Targeting for Viral Immune Escape. Viruses. 2011; 3(7):1166-1178. https://doi.org/10.3390/v3071166
Chicago/Turabian StyleMünz, Christian. 2011. "Beclin-1 Targeting for Viral Immune Escape" Viruses 3, no. 7: 1166-1178. https://doi.org/10.3390/v3071166