miRNA Profiles of Monocyte-Lineage Cells Are Consistent with Complicated Roles in HIV-1 Restriction

Abstract
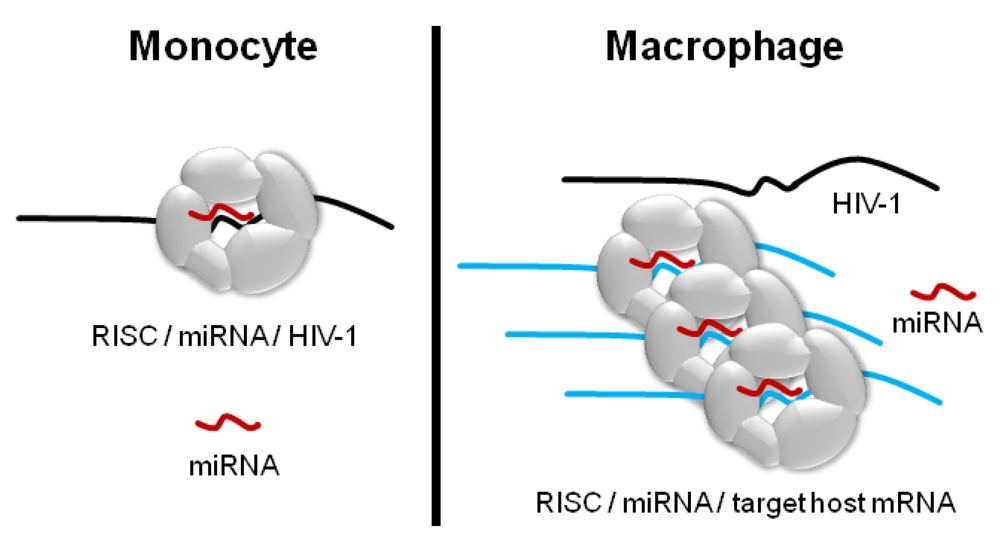
:1. Introduction

{kind=link}
{kind=link}
| miRNA | Wang | Coley | Forrest | Chen, Kasashima |
|---|---|---|---|---|
| U937 | U937 | THP-1 | HL-60 | |
| down | x | x | ||
| miR-17 | ||||
| up | x | x | ||
| miR-21 | ||||
| miR-22 | x | x | x | |
| miR-23a/b | x | x | x | |
| miR-24 | x | x | ||
| miR-26a/b | x | x | x | |
| miR-27a/b | x | x | ||
| miR-29a | x | x | x | |
| miR-29b | x | x | x | |
| miR-132 | x | x | ||
| miR-146a | x | x | x | |
| miR-146b | x | x | x | x |
| miR-221 | x | x | x | x |
| miR-222 | x | x | x | x |
| miR-424 | x | x | x | |
| miR-663 | x | x |
2. Results and Discussion
2.1. Differential Regulation of miRNAs: New Evidence
2.2. miR-29 Family
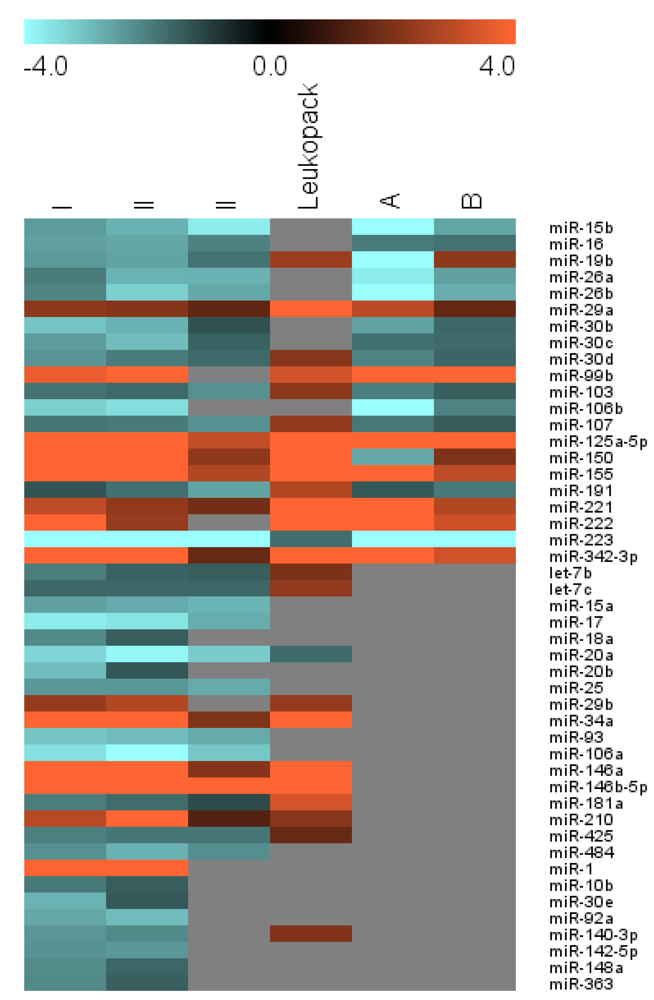
| Group | miRNA | B | Donor I FC | Donor II FC | Donor III FC | Leuko-pack | Sung/Rice FC | Novel inter-action | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MC | DS | MC | DS | MC | DS | DS | A | B | ||||
| I | miR-15b | 0.37 | −2.45 | −2.89 | −2.80 | −3.95 | −3.72 | −3.05 | −6.03 | −2.61 | ||
| miR-16 | 0.82 | −2.52 | −3.11 | −2.61 | −3.67 | −2.04 | −1.72 | −1.93 | −1.79 | |||
| miR-19b | −2.42 | −2.82 | −2.58 | −3.13 | −1.80 | −2.11 | 2.46 | −4.16 | 2.25 | |||
| miR-26a | −1.95 | −2.39 | −2.82 | −3.92 | −2.81 | −2.38 | −3.72 | −2.52 | * | |||
| miR-26b | −2.09 | −2.56 | −3.27 | −4.40 | −2.64 | −2.02 | −5.41 | −2.73 | * | |||
| miR-29a | 0.78 | 2.25 | 1.79 | 2.11 | 1.48 | 1.51 | 1.73 | 5.34 | 2.92 | 1.58 | ||
| miR-30b | −3.08 | −3.72 | −2.78 | −3.61 | −1.29 | −2.54 | −1.60 | |||||
| miR-30c | 0.55 | −2.46 | −2.91 | −2.97 | −3.77 | −1.55 | −1.75 | −1.67 | ||||
| miR-30d | −2.35 | −2.74 | −1.95 | −2.65 | −1.67 | 2.14 | −2.07 | −1.59 | ||||
| miR-99b | 3.07 | 3.73 | 2.63 | 4.16 | 2.51 | 1.94 | 3.29 | 33.77 | 61.46 | |||
| miR-103 | −1.82 | −2.21 | −1.66 | −2.34 | −2.27 | −1.93 | 2.18 | −1.98 | −1.48 | # | ||
| miR-106b | 0.83 | −3.26 | −3.78 | −3.48 | −4.42 | −5.08 | −2.07 | |||||
| miR-107 | 0.10 | −1.86 | −2.26 | −1.92 | −2.73 | −2.28 | −1.91 | 2.31 | −1.87 | −1.45 | # | |
| miR-125a-5p | 3.19 | 6.65 | 5.59 | 5.50 | 6.10 | 3.05 | 3.90 | 6.06 | 8.91 | 30.89 | # | |
| miR-150 | 1.96 | 5.99 | 4.12 | 14.29 | 8.69 | 2.27 | 2.50 | 5.49 | −2.66 | 1.99 | ||
| miR-155 | 4.54 | 5.46 | 4.33 | 4.86 | 3.38 | 2.82 | 3.11 | 6.25 | 8.94 | 3.00 | ||
| miR-191 | −1.33 | −1.66 | −1.77 | −2.46 | −2.56 | −2.14 | 2.79 | −1.41 | −1.89 | * | ||
| miR-221 | 1.97 | 3.02 | 2.43 | 2.28 | 1.63 | 1.84 | 2.06 | 5.37 | 6.36 | 2.81 | # | |
| miR-222 | 1.32 | 4.36 | 3.47 | 2.37 | 1.63 | 5.48 | 7.26 | 3.22 | # | |||
| miR-223 | 0.89 | −4.23 | −5.31 | −6.95 | −9.88 | −5.15 | −4.18 | −1.72 | −5.96 | −5.61 | ||
| miR-342-3p | 5.41 | 7.01 | 5.40 | 6.16 | 4.23 | 1.62 | 1.80 | 7.39 | 8.40 | 3.27 | ||
| II | let-7b | −1.96 | −2.42 | −1.54 | −2.17 | −1.49 | 1.97 | # | ||||
| let-7c | −1.62 | −1.97 | −1.65 | −2.30 | −1.60 | 2.28 | # | |||||
| miR-15a | −2.52 | −3.07 | −2.68 | −3.65 | −2.84 | −2.19 | # | |||||
| miR-17 | 0.91 | −3.71 | −4.30 | −3.56 | −4.50 | −2.72 | −2.01 | |||||
| miR-18a | −2.15 | −2.25 | −1.49 | −2.30 | −1.63 | # | ||||||
| miR-20a | 2.61 | −3.36 | −4.01 | −3.89 | −5.09 | −3.20 | −2.52 | −1.67 | ||||
| miR-20b | 0.01 | −2.97 | −3.45 | −1.36 | −2.43 | −1.57 | # | |||||
| miR-25 | −2.40 | −2.84 | −2.38 | −3.22 | −2.67 | −2.18 | ||||||
| miR-29b | 0.89 | 2.37 | 1.81 | 2.83 | 1.79 | 2.37 | ||||||
| miR-34a | 4.19 | 10.66 | 6.97 | 7.39 | 6.16 | 2.02 | 2.53 | 5.15 | * # | |||
| miR-93 | 1.65 | −3.11 | −3.77 | −2.96 | −4.00 | −2.68 | −2.26 | |||||
| miR-106a | 1.99 | −3.52 | −4.20 | −4.01 | −5.02 | −3.13 | −2.61 | |||||
| miR-146a | 5.75 | 8.40 | 6.47 | 9.75 | 6.43 | 2.10 | 2.24 | 7.68 | * | |||
| miR-146b-5p | 5.37 | 7.50 | 5.75 | 11.08 | 7.20 | 9.74 | 10.38 | 6.88 | * | |||
| miR-181a | −1.97 | −2.31 | −1.70 | −2.32 | −1.18 | 3.38 | * | |||||
| miR-210 | 1.32 | 2.89 | 2.71 | 4.39 | 3.88 | 1.31 | 1.54 | 2.14 | * # | |||
| miR-425 | −2.03 | −2.39 | −1.85 | −2.44 | −1.86 | −1.78 | 1.62 | # | ||||
| miR-484 | −2.26 | −2.71 | −2.79 | −3.55 | −2.23 | −1.85 | ||||||
| III | miR-1 | 4.90 | 11.14 | 7.19 | 4.98 | 4.90 | ||||||
| miR-10b | −1.91 | −2.02 | −1.46 | −1.74 | ||||||||
| miR-30e | −2.81 | −2.82 | −1.39 | −2.62 | ||||||||
| miR-92a | 0.37 | −2.63 | −3.17 | −2.95 | −4.00 | |||||||
| miR-140-3p | 0.07 | −2.36 | −2.59 | −2.15 | −2.58 | 2.04 | ||||||
| miR-142-5p | −2.31 | −2.39 | −2.39 | −2.91 | ||||||||
| miR-148a | −2.21 | −2.41 | −1.61 | −2.13 | ||||||||
| miR-363 | 1.62 | −2.15 | −2.89 | −1.50 | −2.76 | |||||||
| miRNA | Donor I | Donor II | Donor III | Leuko-pack | Sung/Rice | Wang et al. | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| MC | DS | MC | DS | MC | DS | DS | A | B | ||
| miR-28-3p | nc | −1.63 | nc | −1.65 | nc | nc | nc | nr | nr | ~−2 |
| miR-125b | bb | bb | bb | bb | bb | bb | bb | nr | 1.83 | nr |
| miR-125a-5p | 6.65 | 5.59 | 5.50 | 6.10 | 3.05 | 3.90 | 6.06 | 8.91 | 30.89 | nr |
| miR-150 | 6.00 | 4.12 | 14.29 | 8.69 | 2.27 | 2.50 | 5.49 | −2.66 | 1.99 | ~−10 |
| miR-223 | −4.23 | −5.31 | −6.95 | −9.88 | −5.15 | −4.18 | −1.72 | −5.96 | −5.61 | ~−10 |
| miR-198 | bb | bb | bb | bb | bb | bb | bb | −8.87 | −59.18 | nr |
| miR-382 | bb | bb | bb | bb | bb | bb | −1.66 | nr | nr | ~−10 |
2.3. Ant-HIV-1 miRs-28-3p, -125b, -150, -223, and -382
2.4. miR-198
2.5. miR-17/92 Cluster
2.6. Additional miRNAs — Direct Interactions?
2.7. Additional miRNAs — Indirect Effects
3. Experimental Section
3.1. Cell Isolation and Culture
3.2. RNA Extraction
3.3. miRNA Microarrays
3.4. Analysis
3.5. Literature and Data Re-Analysis
3.6. Data Accessibility
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Shen, L.; Siliciano, R.F. Viral reservoirs, residual viremia, and the potential of highly active antiretroviral therapy to eradicate HIV infection. J. Allergy Clin. Immunol. 2008, 122, 22–28. [Google Scholar] [CrossRef]
- Richman, D.D.; Margolis, D.M.; Delaney, M.; Greene, W.C.; Hazuda, D.; Pomerantz, R.J. The challenge of finding a cure for HIV infection. Science 2009, 323, 1304–1307. [Google Scholar]
- Whitney, J.B.; Lim, S.Y.; Wainberg, M.A. Evolutionary mechanisms of retroviral persistence. AIDS Rev. 2011, 13, 234–239. [Google Scholar]
- Margolis, D.M. Eradication therapies for HIV infection: Time to begin again. AIDS Res. Hum. Retrovir. 2011, 27, 347–353. [Google Scholar] [CrossRef]
- Finzi, D.; Hermankova, M.; Pierson, T.; Carruth, L.M.; Buck, C.; Chaisson, R.E.; Quinn, T.C.; Chadwick, K.; Margolick, J.; Brookmeyer, R.; et al. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 1997, 278, 1295–1300. [Google Scholar] [CrossRef]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierson, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat. Med. 1999, 5, 512–517. [Google Scholar] [CrossRef]
- Alexaki, A.; Liu, Y.; Wigdahl, B. Cellular reservoirs of HIV-1 and their role in viral persistence. Curr. HIV Res. 2008, 6, 388–400. [Google Scholar] [CrossRef]
- Clements, J.E.; Gama, L.; Graham, D.R.; Mankowski, J.L.; Zink, M.C. A simian immunodeficiency virus macaque model of highly active antiretroviral treatment: Viral latency in the periphery and the central nervous system. Curr. Opin. HIV AIDS 2011, 6, 37–42. [Google Scholar] [CrossRef]
- Sharova, N.; Swingler, C.; Sharkey, M.; Stevenson, M. Macrophages archive HIV-1 virions for dissemination in trans. EMBO J. 2005, 24, 2481–2489. [Google Scholar] [CrossRef]
- Kedzierska, K.; Crowe, S.M. The role of monocytes and macrophages in the pathogenesis of HIV-1 infection. Curr. Med. Chem. 2002, 9, 1893–1903. [Google Scholar]
- Sonza, S.; Maerz, A.; Deacon, N.; Meanger, J.; Mills, J.; Crowe, S. Human immunodeficiency virus type 1 replication is blocked prior to reverse transcription and integration in freshly isolated peripheral blood monocytes. J. Virol. 1996, 70, 3863–3869. [Google Scholar]
- Fontana, L.; Pelosi, E.; Greco, P.; Racanicchi, S.; Testa, U.; Liuzzi, F.; Croce, C.M.; Brunetti, E.; Grignani, F.; Peschle, C. MicroRNAs 17-5p-20a-106a control monocytopoiesis through aml1 targeting and M-CSF receptor upregulation. Nat. Cell Biol. 2007, 9, 775–787. [Google Scholar] [CrossRef]
- Wang, X.; Ye, L.; Hou, W.; Zhou, Y.; Wang, Y.J.; Metzger, D.S.; Ho, W.Z. Cellular microRNA expression correlates with susceptibility of monocytes/macrophages to HIV-1 infection. Blood 2009, 113, 671–674. [Google Scholar] [CrossRef]
- Sung, T.L.; Rice, A.P. MiR-198 inhibits HIV-1 gene expression and replication in monocytes and its mechanism of action appears to involve repression of Cyclin T1. PLoS Pathog. 2009, 5, e1000263. [Google Scholar] [CrossRef]
- Coley, W.; van Duyne, R.; Carpio, L.; Guendel, I.; Kehn-Hall, K.; Chevalier, S.; Narayanan, A.; Luu, T.; Lee, N.; Klase, Z.; et al. Absence of Dicer in monocytes and its regulation by HIV-1. J. Biol. Chem. 2010, 285, 31930–31943. [Google Scholar]
- Jaskiewicz, L.; Filipowicz, W. Role of Dicer in posttranscriptional RNA silencing. Curr. Top. Microbiol. Immunol. 2008, 320, 77–97. [Google Scholar] [CrossRef]
- Klase, Z.; Kale, P.; Winograd, R.; Gupta, M.V.; Heydarian, M.; Berro, R.; McCaffrey, T.; Kashanchi, F. HIV-1 Tar element is processed by Dicer to yield a viral micro-RNA involved in chromatin remodeling of the viral LTR. BMC Mol. Biol. 2007, 8, 63. [Google Scholar] [CrossRef]
- Wang, J.; Xiang, G.; Mitchelson, K.; Zhou, Y. Microarray profiling of monocytic differentiation reveals miRNA-mRNA intrinsic correlation. J. Cell Biochem. 2011, 112, 2443–2453. [Google Scholar] [CrossRef]
- Brazma, A.; Hingamp, P.; Quackenbush, J.; Sherlock, G.; Spellman, P.; Stoeckert, C.; Aach, J.; Ansorge, W.; Ball, C.A.; Causton, H.C.; et al. Minimum information about a microarray experiment (MIAME)-toward standards for microarray data. Nat. Genet. 2001, 29, 365–371. [Google Scholar] [CrossRef]
- Wang, J.; Xiang, G.; Mitchelson, K.; Zhou, Y. Microarray profiling of monocytic differentiation reveals miRNA-mRNA intrinsic correlation (miRNA). J. Cell. Biochem. 2011, 112, 2443–2453. [Google Scholar] [CrossRef]
- Li, T.; Morgan, M.J.; Choksi, S.; Zhang, Y.; Kim, Y.S.; Liu, Z.G. MicroRNAs modulate the noncanonical transcription factor NF-kappab pathway by regulating expression of the kinase IKKalpha during macrophage differentiation. Nat. Immunol. 2010, 11, 799–805. [Google Scholar] [CrossRef]
- Schmeier, S.; MacPherson, C.R.; Essack, M.; Kaur, M.; Schaefer, U.; Suzuki, H.; Hayashizaki, Y.; Bajic, V.B. Deciphering the transcriptional circuitry of microRNA genes expressed during human monocytic differentiation. BMC Genomics 2009, 10, 595. [Google Scholar]
- Forrest, A.R.; Kanamori-Katayama, M.; Tomaru, Y.; Lassmann, T.; Ninomiya, N.; Takahashi, Y.; de Hoon, M.J.; Kubosaki, A.; Kaiho, A.; Suzuki, M.; et al. Induction of microRNAs, miR-155, miR-222, miR-424 and miR-503, promotes monocytic differentiation through combinatorial regulation. Leukemia 2010, 24, 460–466. [Google Scholar] [CrossRef]
- Ikeo, K.; Ishi-i, J.; Tamura, T.; Gojobori, T.; Tateno, Y. Cibex: Center for information biology gene expression database. C. R. Biol. 2003, 326, 1079–1082. [Google Scholar] [CrossRef]
- Chen, A.; Luo, M.; Yuan, G.; Yu, J.; Deng, T.; Zhang, L.; Zhou, Y.; Mitchelson, K.; Cheng, J. Complementary analysis of microRNA and mRNA expression during phorbol 12-myristate 13-acetate (TPA)-induced differentiation of HL-60 cells. Biotechnol. Lett. 2008, 30, 2045–2052. [Google Scholar] [CrossRef]
- Kasashima, K.; Nakamura, Y.; Kozu, T. Altered expression profiles of microRNAs during TPA-induced differentiation of HL-60 cells. Biochem. Biophys. Res. Commun. 2004, 322, 403–410. [Google Scholar] [CrossRef]
- Fontana, L.; Sorrentino, A.; Peschle, C. Regulation of monocytopoiesis by microRNAs. Methods Mol. Biol. 2010, 667, 165–176. [Google Scholar] [CrossRef]
- Rosa, A.; Ballarino, M.; Sorrentino, A.; Sthandier, O.; de Angelis, F.G.; Marchioni, M.; Masella, B.; Guarini, A.; Fatica, A.; Peschle, C.; et al. The interplay between the master transcription factor PU.1 and miR-424 regulates human monocyte/macrophage differentiation. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 19849–19854. [Google Scholar]
- Witwer, K.W.; Sisk, J.M.; Gama, L.; Clements, J.E. MicroRNA regulation of IFN-beta protein expression: Rapid and sensitive modulation of the innate immune response. J. Immunol. 2010, 184, 2369–2376. [Google Scholar] [CrossRef]
- Byerly, S.; Sundin, K.; Raja, R.; Stanchfield, J.; Bejjani, B.A.; Shaffer, L.G. Effects of ozone exposure during microarray posthybridization washes and scanning. J. Mol. Diagn. 2009, 11, 590–597. [Google Scholar] [CrossRef]
- Fare, T.L.; Coffey, E.M.; Dai, H.; He, Y.D.; Kessler, D.A.; Kilian, K.A.; Koch, J.E.; LeProust, E.; Marton, M.J.; Meyer, M.R.; et al. Effects of atmospheric ozone on microarray data quality. Anal. Chem. 2003, 75, 4672–4675. [Google Scholar] [CrossRef]
- Lusa, L.; McShane, L.M.; Reid, J.F.; de Cecco, L.; Ambrogi, F.; Biganzoli, E.; Gariboldi, M.; Pierotti, M.A. Challenges in projecting clustering results across gene expression-profiling datasets. J. Natl. Cancer Inst. 2007, 99, 1715–1723. [Google Scholar] [CrossRef]
- Kerr, M.K.; Martin, M.; Churchill, G.A. Analysis of variance for gene expression microarray data. J. Comput. Biol. 2000, 7, 819–837. [Google Scholar] [CrossRef]
- Witwer, K.W.; Sisk, J.M.; Clements, J.E. Johns Hopkins University, Baltimore, MD, USA, Unpublished work, 2010.
- Ahluwalia, J.K.; Khan, S.Z.; Soni, K.; Rawat, P.; Gupta, A.; Hariharan, M.; Scaria, V.; Lalwani, M.; Pillai, B.; Mitra, D.; et al. Human cellular microRNA hsa-miR-29a interferes with viral Nef protein expression and HIV-1 replication. Retrovirology 2008, 5, 117. [Google Scholar] [CrossRef]
- Nathans, R.; Chu, C.Y.; Serquina, A.K.; Lu, C.C.; Cao, H.; Rana, T.M. Cellular microRNA and P bodies modulate host-HIV-1 interactions. Mol. Cell 2009, 34, 696–709. [Google Scholar] [CrossRef]
- Hariharan, M.; Scaria, V.; Pillai, B.; Brahmachari, S.K. Targets for human encoded microRNAs in HIV genes. Biochem. Biophys. Res. Commun. 2005, 337, 1214–1218. [Google Scholar] [CrossRef]
- Chiang, K.; Sung, T.L.; Rice, A.P. Regulation of Cyclin T1 and HIV-1 replication by microRNAs in resting CD4+ T lymphocytes. J. Virol. 2012, 86, 3244–3252. [Google Scholar] [CrossRef]
- Sisk, J.M.; Witwer, K.; Clements, J.E. Interferon beta induces anti-SIV miRNA as part of the innate immune response. In Proceedings of CROI 2012 (Conference on Retroviruses and Opportunistic Infections), Seattle, WA, USA, 5–8 March 2012.
- Sisk, J.M.; Witwer, K.W.; Clements, J.E. Johns Hopkins University, Baltimore, MD, USA. Innate immune responses induce anti-SIV miRNAs. Unpublished work, to be submitted for publication, 2012.
- Sung, T.L.; Rice, A.P. MiR-198 inhibits HIV-1 gene expression and replication in monocytes and its mechanism of action appears to involve repression of Cyclin T1. Dataset. Available online: http://www.bcm.edu/molvir/ricelab/data (accessed on 20 September 2012).
- Simon, R.; Lam, A.; Li, M.C.; Ngan, M.; Menenzes, S.; Zhao, Y. Analysis of gene expression data using BRB-Array Tools. Cancer Inform. 2007, 3, 11–17. [Google Scholar]
- GenePix Pro, version 7.0; Molecular Devices, LLC: Sunnyvale, CA, USA, 2008.
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef]
- Wang, X.; Ye, L.; Zhou, Y.; Liu, M.Q.; Zhou, D.J.; Ho, W.Z. Inhibition of anti-HIV microRNA expression: A mechanism for opioid-mediated enhancement of HIV infection of monocytes. Am. J. Pathol. 2011, 178, 41–47. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, X.; Liu, M.; Hu, Q.; Song, L.; Ye, L.; Zhou, D.; Ho, W. A critical function of toll-like receptor-3 in the induction of anti-human immunodeficiency virus activities in macrophages. Immunology 2010, 131, 40–49. [Google Scholar]
- Swaminathan, S.; Zaunders, J.; Wilkinson, J.; Suzuki, K.; Kelleher, A.D. Does the presence of anti-HIV miRNAs in monocytes explain their resistance to HIV-1 infection? Blood 2009, 113, 5029–5030, author reply 5030–5021. [Google Scholar] [CrossRef]
- Hulsmans, M.; Sinnaeve, P.; van der Schueren, B.; Mathieu, C.; Janssens, S.; Holvoet, P. Decreased miR-181a expression in monocytes of obese patients is associated with the occurrence of metabolic syndrome and coronary artery disease. J. Clin. Endocrinol. Metab. 2012, 97, E1213–E1218. [Google Scholar]
- Hashimi, S.T.; Fulcher, J.A.; Chang, M.H.; Gov, L.; Wang, S.; Lee, B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood 2009, 114, 404–414. [Google Scholar] [CrossRef]
- Murray, D.D.; Swaminathan, S. MicroRNA profiling of monocytes in LTNPs and chronic HIV patients (CHI). GEO dataset GSE38556, 2012. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE38556 (accessed on 20 September 2012). National Center for Biotechnology Information.
- Allantaz, F.; Cheng, D.T.; Bergauer, T.; Ravindran, P.; Rossier, M.F.; Ebeling, M.; Badi, L.; Reis, B.; Bitter, H.; D'Asaro, M.; et al. Expression profiling of human immune cell subsets identifies miRNA-mRNA regulatory relationships correlated with cell type specific expression. PLoS One 2012, 7, e29979. [Google Scholar]
- Gupta, A.; Nagilla, P.; Le, H.S.; Bunney, C.; Zych, C.; Thalamuthu, A.; Bar-Joseph, Z.; Mathavan, S.; Ayyavoo, V. Comparative expression profile of miRNA and mRNA in primary peripheral blood mononuclear cells infected with human immunodeficiency virus (HIV-1). PLoS One 2011, 6, e22730. [Google Scholar]
- Luers, A.J.; Loudig, O.D.; Berman, J.W. MicroRNAs are expressed and processed by human primary macrophages. Cell Immunol. 2010, 263, 1–8. [Google Scholar] [CrossRef]
- Witwer, K.W.; Watson, A.K.; Blankson, J.N.; Clements, J.E. Relationships of pbmc microRNA expression, plasma viral load, and CD4+ T-cell count in HIV-1-infected elite suppressors and viremic patients. Retrovirology 2012, 9, 5. [Google Scholar] [CrossRef]
- Van Eisden, R.G.; Ayoubi, T. MiRNA profiling of FACS sorted human leukocyte cell types. GEO Dataset GSE19183, 2009. Available online: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE19183 (accessed on 20 September 2012).
- Mendell, J.T. MiRiad roles for the miR-17-92 cluster in development and disease. Cell 2008, 133, 217–222. [Google Scholar] [CrossRef]
- Triboulet, R.; Mari, B.; Lin, Y.L.; Chable-Bessia, C.; Bennasser, Y.; Lebrigand, K.; Cardinaud, B.; Maurin, T.; Barbry, P.; Baillat, V.; et al. Suppression of microRNA-silencing pathway by HIV-1 during virus replication. Science 2007, 315, 1579–1582. [Google Scholar]
- Schopman, N.C.; van Montfort, T.; Willemsen, M.; Knoepfel, S.A.; Pollakis, G.; van Kampen, A.; Sanders, R.W.; Haasnoot, J.; Berkhout, B. Selective packaging of cellular miRNAs in HIV-1 particles. Virus Res. 2012. [Google Scholar]
- Althaus, C.F.; Vongrad, V.; Niederost, B.; Joos, B.; di Giallonardo, F.; Rieder, P.; Pavlovic, J.; Trkola, A.; Gunthard, H.F.; Metzner, K.J.; et al. Tailored enrichment strategy detects low abundant small noncoding RNAs in HIV-1 infected cells. Retrovirology 2012, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.S.; Wu, T.C.; Sang, W.W.; Ruan, Z. MiR-217 is involved in tat-induced HIV-1 long terminal repeat (LTR) transactivation by down-regulation of SIRT1. Biochim. Biophys. Acta 2012, 1823, 1017–1023. [Google Scholar] [CrossRef]
- Witwer, K.W.; Sarbanes, S.L.; Liu, J.; Clements, J.E. A plasma microRNA signature of acute lentiviral infection: Biomarkers of CNS disease. AIDS 2011, 204, 1104–1114. [Google Scholar]
- Smyth, G.K. Limma: Linear models for microarray data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Gentleman, R., Dudoit, V.C.S., Irizarry, R., Huber, W., Eds.; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar]
- NCode Profiler, version 2.0; previously Invitrogen; Life Technologies: Carlsbad, CA, USA, 2007.
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. Tm4 microarray software suite. Methods Enzymol. 2006, 411, 134–193. [Google Scholar] [CrossRef]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. Tm4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar]
- Larsson, E.; Sander, C.; Marks, D.S. MRNA turnover rate limits microRNA and siRNA efficacy. In MicroRNAs and Human Disease; Keystone Symposia on Molecular and Cellular Biology: Banff, Alberta, Canada, 2011; p. 208. [Google Scholar]
- Seitz, H. Redefining microRNA targets. Curr. Biol. 2009, 19, 870–873. [Google Scholar] [CrossRef]
- Bolstad, B.M.; Collin, F.; Simpson, K.M.; Irizarry, R.A.; Speed, T.P. Experimental design and low-level analysis of microarray data. Int. Rev. Neurobiol. 2004, 60, 25–58. [Google Scholar] [CrossRef]
- Witwer, K.; Clements, J. Evidence for CD4+ T-cell miRNA differences of HIV-1 treatment naïve patients and elite suppressors: A re-analysis. Blood 2012, 119, 6395–6396. [Google Scholar] [CrossRef]
- Kerr, M.K.; Churchill, G.A. Experimental design for gene expression microarrays. Biostatistics 2001, 2, 183–201. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. Roy. Stat. Soc. B Met. 1995, 57, 289–300. [Google Scholar]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene expression omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef]
- Brazma, A.; Parkinson, H.; Sarkans, U.; Shojatalab, M.; Vilo, J.; Abeygunawardena, N.; Holloway, E.; Kapushesky, M.; Kemmeren, P.; Lara, G.G.; et al. ArrayExpress—A public repository for microarray gene expression data at the EBI. Nucleic Acids Res. 2003, 31, 68–71. [Google Scholar]
- Lawrence, T.; Natoli, G. Transcriptional regulation of macrophage polarization: Enabling diversity with identity. Nat. Rev. Immunol. 2011, 11, 750–761. [Google Scholar] [CrossRef]
- Hristov, M.; Weber, C. Differential role of monocyte subsets in atherosclerosis. Thromb. Haemost 2011, 106, 757–762. [Google Scholar] [CrossRef]
- Gama, L.; Shirk, E.N.; Russell, J.N.; Carvalho, K.I.; Li, M.; Queen, S.E.; Kalil, J.; Zink, M.C.; Clements, J.E.; Kallas, E.G. Expansion of a subset of CD14highCD16negCCR2low/neg monocytes functionally similar to myeloid-derived suppressor cells during SIV and HIV infection. J. Leukoc. Biol. 2012, 91, 803–816. [Google Scholar] [CrossRef] [Green Version]
- Graff, J.W.; Dickson, A.M.; Clay, G.; McCaffrey, A.P.; Wilson, M.E. Identifying functional microRNAs in macrophages with polarized phenotypes. J. Biol. Chem. 2012, 287, 21816–21825. [Google Scholar] [CrossRef]
- Ng, Y.S.; Roca, H.; Fuller, D.; Sud, S.; Pienta, K.J. Chemical transfection of dye-conjugated microRNA precursors for microRNA functional analysis of M2 macrophages. J. Cell. Biochem. 2012, 113, 1714–1723. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sisk, J.M.; Clements, J.E.; Witwer, K.W. miRNA Profiles of Monocyte-Lineage Cells Are Consistent with Complicated Roles in HIV-1 Restriction. Viruses 2012, 4, 1844-1864. https://doi.org/10.3390/v4101844
Sisk JM, Clements JE, Witwer KW. miRNA Profiles of Monocyte-Lineage Cells Are Consistent with Complicated Roles in HIV-1 Restriction. Viruses. 2012; 4(10):1844-1864. https://doi.org/10.3390/v4101844
Chicago/Turabian StyleSisk, Jeanne M., Janice E. Clements, and Kenneth W. Witwer. 2012. "miRNA Profiles of Monocyte-Lineage Cells Are Consistent with Complicated Roles in HIV-1 Restriction" Viruses 4, no. 10: 1844-1864. https://doi.org/10.3390/v4101844