2.1. Effect of Aphidius Colemani on Aphid Dispersal and the Spread of Cucumber Mosaic Virus
The population density of adult morphs and nymphs in the CMV-infected source plant located in the middle of the cage where aphids were released was frequently higher in the control cages than in those containing the parasitoid
A. colemani, although no significant differences were found (
Table 1). Parasitoids successfully located a variable number of aphids in the virus-infected source plant and mummies could be observed (2.3 ± 1.5) 7 days after the release of parasitoids, whereas they could not be detected after 2 days, as mummies were not yet developed. There were fewer aphids on the peripheral test plants in the arenas (= cages) without
A. colemani after 2 days (
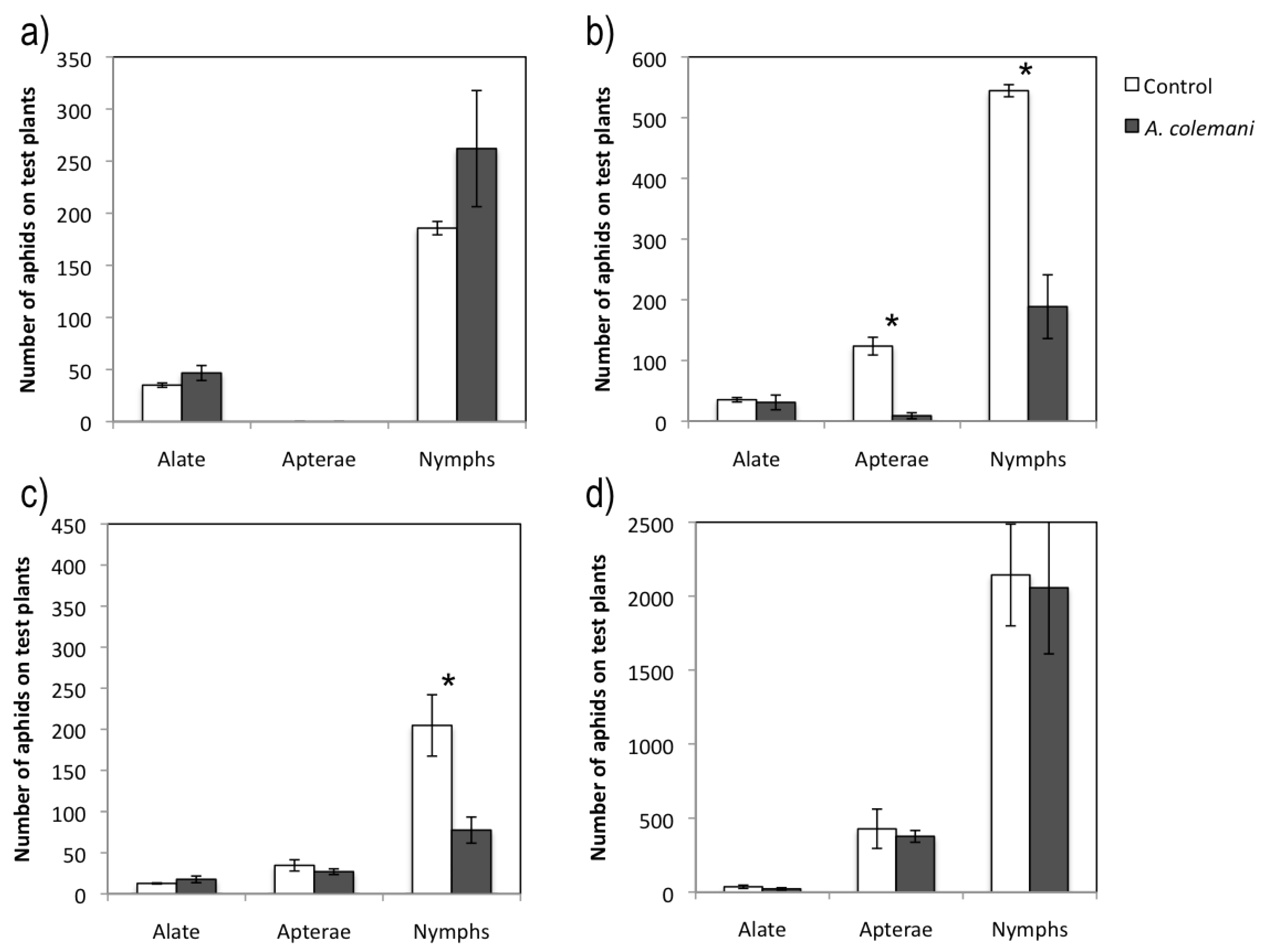
Figure 1a), but this trend was not repeated after 7 days, with significantly more apterae adults (
t = 6.775; 4 gl;
p = 0.002) and nymphs (
t = 11.864; 4 gl;
p < 0.001) in the test plants of control arenas (
Figure 1b). The number of nymphs increased considerably after 7 days (
Figure 1b). Besides, recognizable mummies could be detected in peripheral plants in CMV at 7 days (2.0 ± 2.0) but not at 2 days.
Table 1.
Mean ± S.E. density (number of individuals/plant) of adult morphs and nymphs in the CMV-infected virus source plant after 2 and 7 days in cages with and without (control) A. colemani followed by statistics according to Student’s t test (p < 0.05).
Table 1.
Mean ± S.E. density (number of individuals/plant) of adult morphs and nymphs in the CMV-infected virus source plant after 2 and 7 days in cages with and without (control) A. colemani followed by statistics according to Student’s t test (p < 0.05).
| | 2 days | 7 days |
|---|
| | Control | A. colemani | t | p | Control | A. colemani | t | p |
|---|
| Alate | 24.7 ± 5.5 | 18.0 ± 5.3 | 0.873 | 0.947 | 11.0 ± 3.2 | 9.0 ± 2.5 | 0.524 | 0.628 |
| Apterae | - | - | - | - | 10.7 ± 4.7 | 6.3 ± 3.0 | 0.882 | 0.428 |
| Nymphs | 176.3 ± 55.1 | 128.7 ± 26.6 | 0.745 | 0.498 | 170.0 ± 50.0 | 99.7 ± 33.3 | 1.227 | 0.287 |
Figure 1.
Mean ± S.E. values of total number of aphids on test plants in cages with (grey bars) and without (control cages, white bars) A. colemani. (a) CMV-infected source plant assay at 2 days. (b) CMV-infected source plant assay at 7 days. (c) CABYV-infected source plant assay at 7 days. (d) CABYV-infected source plant assay at 14 days. Bars with asterisks are significantly different according to Student’s t test (p < 0.05).
Figure 1.
Mean ± S.E. values of total number of aphids on test plants in cages with (grey bars) and without (control cages, white bars) A. colemani. (a) CMV-infected source plant assay at 2 days. (b) CMV-infected source plant assay at 7 days. (c) CABYV-infected source plant assay at 7 days. (d) CABYV-infected source plant assay at 14 days. Bars with asterisks are significantly different according to Student’s t test (p < 0.05).
The occupancy rates were calculated as the percentage of test plants with one or more aphids in their different morphs. These rates were consistent with aphid density in the peripheral test plants (
Figure 1a and 1b), as there were significantly fewer plants occupied by aphids in the control arenas than in the arenas with parasitoids after 2 days, but larger occupancy rate in the control after 7 days (
Table 2).
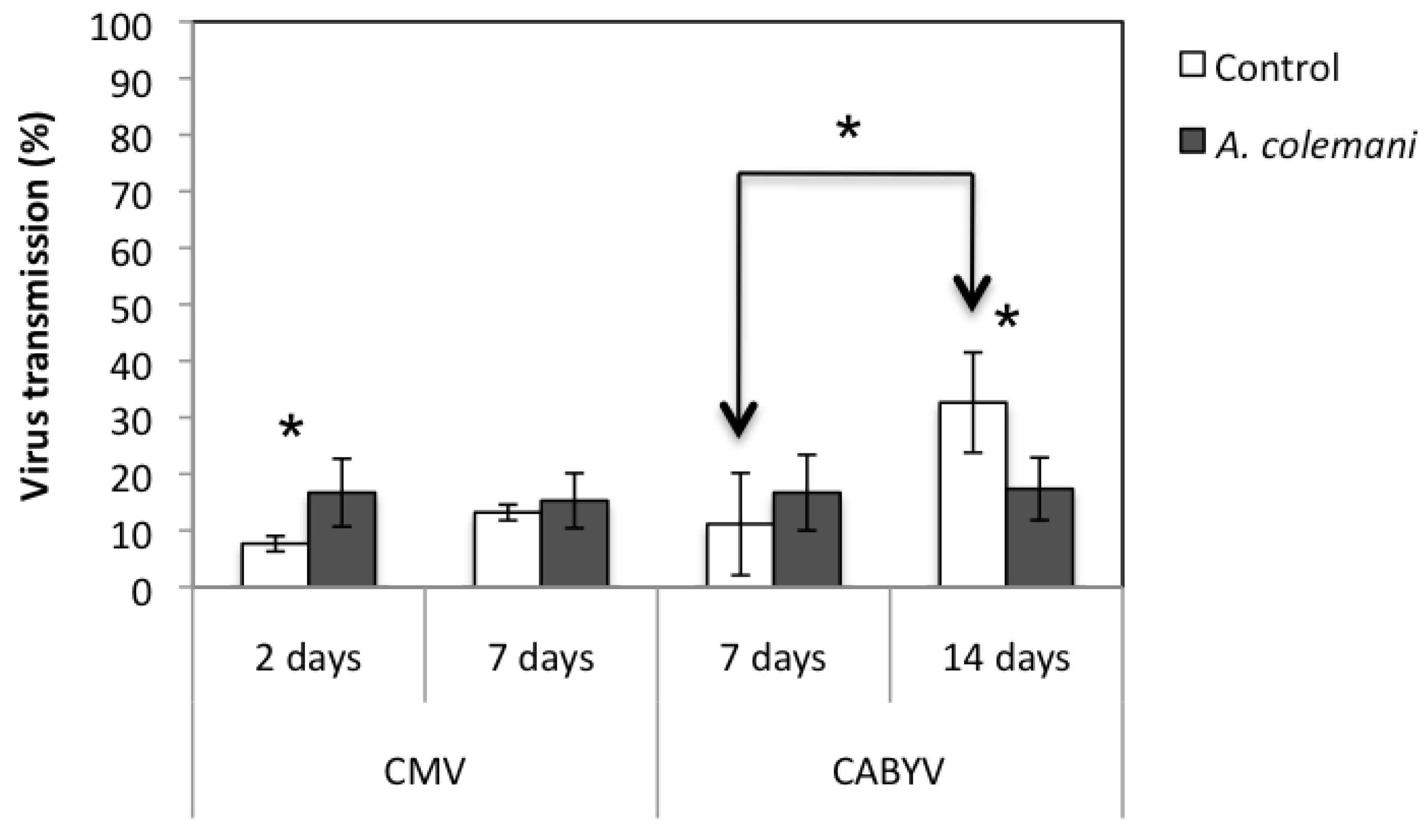
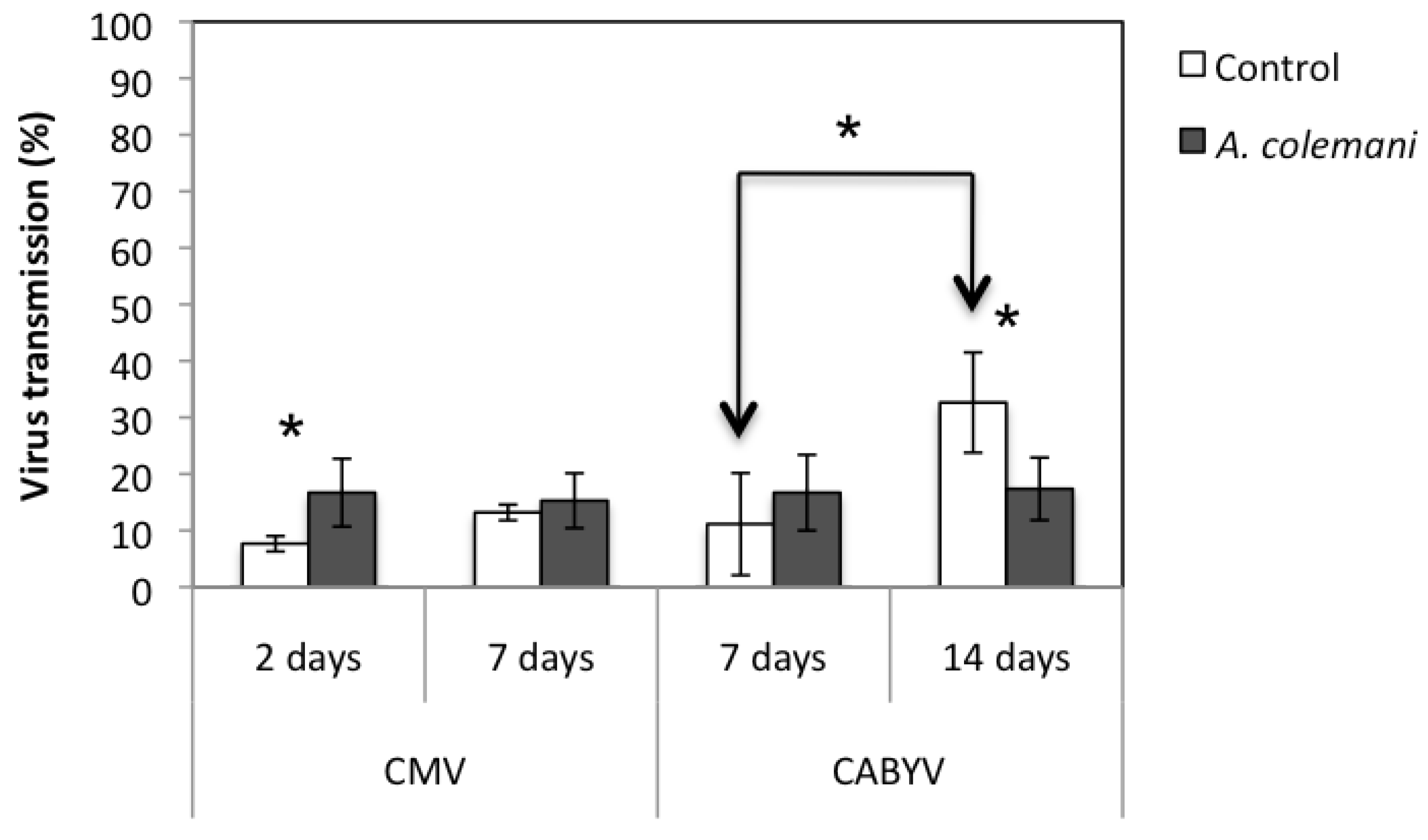
The incidence of CMV at short time (after 2 days) was significantly higher in arenas containing
A. colemani compared to that observed in the control arenas (
X2 = 5.497;
p = 0.020). When the same assay was assessed after 7 days, no significant differences in the incidence of CMV between treatments were detected (
Figure 2).
Table 2.
Mean ± S.E. percentage of test plants occupied by one or more alate, apterae or nymphs (occupancy rate) in CMV-infected source plant assays after 2 and 7 days in cages with and without (control) A. colemani. Means in the same row followed by statistics in bold are significantly different according to a Chi-square 2 × 2 goodness of fit test (p < 0.05).
Table 2.
Mean ± S.E. percentage of test plants occupied by one or more alate, apterae or nymphs (occupancy rate) in CMV-infected source plant assays after 2 and 7 days in cages with and without (control) A. colemani. Means in the same row followed by statistics in bold are significantly different according to a Chi-square 2 × 2 goodness of fit test (p < 0.05).
| | 2 days | 7 days |
|---|
| | Control | A. colemani | X 2 | p | Control | A. colemani | X 2 | p |
|---|
| Alate | 38.9 ± 2.5 | 54.2 ± 7.9 | 9.930 | 0.002 | 42.4 ± 3.5 | 33.3 ± 8.7 | 2.495 | 0.117 |
| Apterae | - | - | - | - | 38.2 ± 2.8 | 6.3 ± 3.2 | 42.509 | <0.001 |
| Nymphs | 38.2 ± 4.2 | 50.7 ± 8.2 | 4.556 | 0.044 | 50.7 ± 2.5 | 38.2 ± 7.8 | 4.556 | 0.034 |
Figure 2.
Mean ± S.E. values of virus transmission (%) in the arenas with parasitoids (grey bars) and in those without them (control, white bars). Bars with asterisks indicate significant differences according to a Chi-square 2 × 2 goodness of fit test (p < 0.05).
Figure 2.
Mean ± S.E. values of virus transmission (%) in the arenas with parasitoids (grey bars) and in those without them (control, white bars). Bars with asterisks indicate significant differences according to a Chi-square 2 × 2 goodness of fit test (p < 0.05).
Overall, our results suggest that
A. colemani promoted early movement of
A. gossypii from the virus-infected source towards the peripheral plants. Parasitoids significantly increased the colonization of adjacent plants by both adults and nymphs. Consistently, the spread of CMV also increased when parasitoids interacted with aphids for 2 days. However, no differences in virus incidence could be found after 7 days between both treatments and aphids spread around and transmitted CMV equally well under the two treatments (in the presence and absence of
A. colemani). These results may be explained by the mode of transmission of non-persistent viruses and the particular behaviour of
A. gossypii. Transmission of non-persistent viruses is highly favoured if aphids acquire the viral particles during short periods, decreasing its efficiency with a longer acquisition access time [
4]. It is likely that soon after release, parasitoids caused a disturbance and their presence forced aphids to quickly disseminate, which led to an increase in aphid density on peripheral test plants and subsequent transmission of CMV.
A. colemani forced them to escape and, as non-persistent viruses do not require a latent period, aphids were able to inoculate CMV to peripheral plants readily after leaving the central plant. The incidence of CMV remained the same 2 and 7 days after the experiment started in arenas with
A. colemani, probably because parasitised aphids had reduced the distance they moved after few days and parasitoids were not in the attack mood any longer. The fact that mummies were observed in both central and test plants at 7 days seems to corroborate this hypothesis. Conversely, CMV transmission in control arenas increased with time as aphids continued colonizing test plants because their mobility was not jeopardised by parasitism. Similar alterations resulting in an increase in non-persistently and semipersistenly transmitted virus dispersal have been previously reported at short times (1–3 days) [
11,
13,
14,
17], but no information on the long term effects of parasitism on virus spread has been reported so far.
Natural enemies orientate towards plant-host complexes (PHC) by responding to host herbivore-induced plant volatiles and visual cues [
36,
37,
38,
39]. The emission of alarm signals by aphids causes conspecifics to disperse through different strategies such as ‘drop off’ [
18,
19]. That escaping behaviour may even modify virus spread and has been reported with the aphid
A. pisum and its parasitoid
A. ervi [
20]. Furthermore, damaged plants emit the “cry-for-help” signal that may indirectly benefit hosts under herbivore attack [
21]. Therefore, it is evident that there is a need to integrate all these multitrophic reactions between agents resulting in epidemiological consequences [
20,
21].
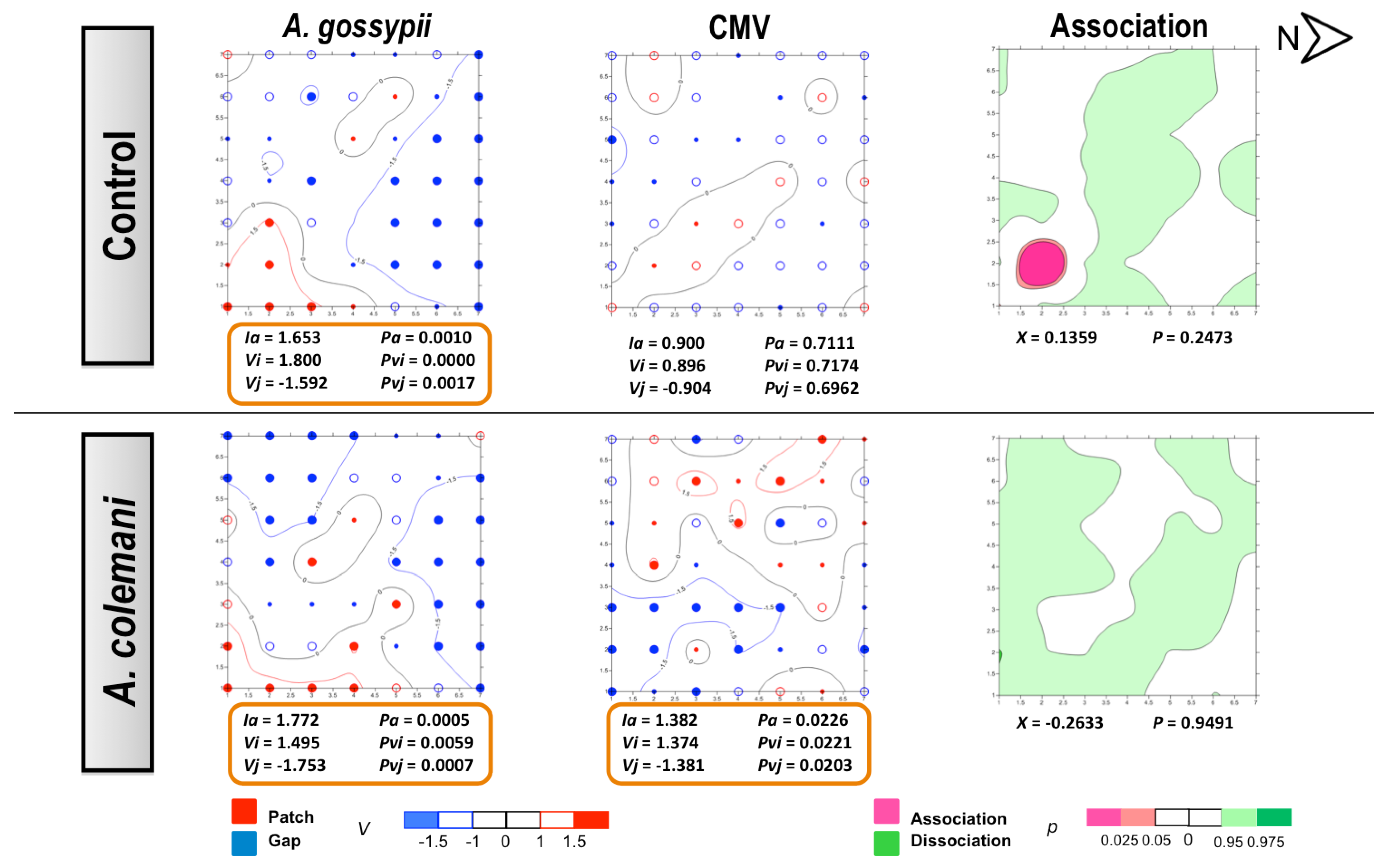
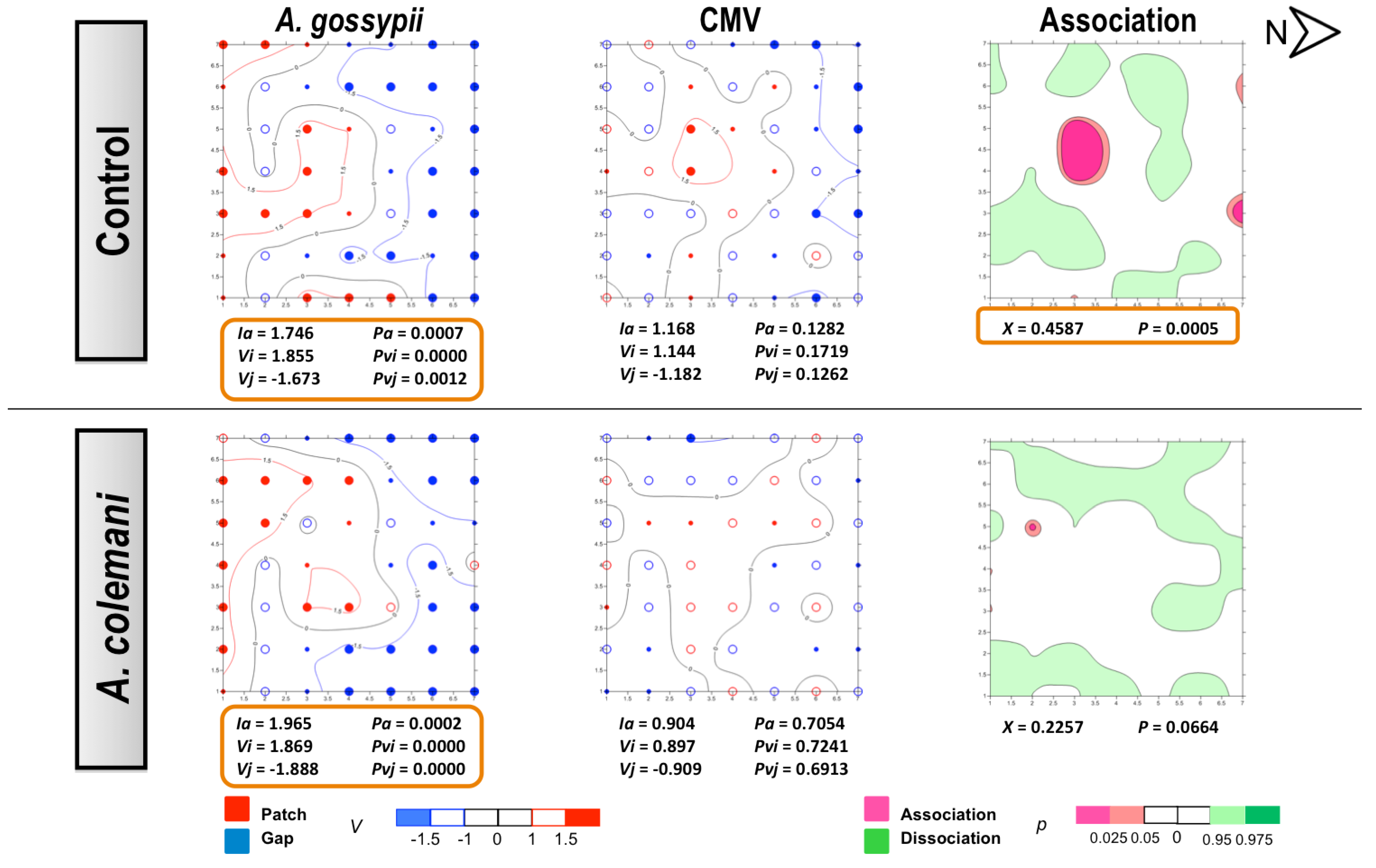
No differences could be found for the mean displacement (δ) of both aphids and CMV (
Table 3). The spatial analysis of CMV-infected source plant experiments showed a significant aggregated distribution of
A. gossypii in treatments with and whitout parasitoids after 2 and 7 days (
Figure 3 and
Figure 4). At 2 days, aphids were restricted to the lower right corner of the experimental arena (
Figure 3) but colonised the entire lower area of the arena after 7 days, with plants remaining unnoccupied in the upper side (
Figure 4). At short term, the spread of CMV followed a regular distribution in the control cages where few isolated plants became infected, although CMV distribution was significantly aggregated in the presence of parasitoids (
Figure 3). When the spatial distribution of aphids and CMV was studied at long term (7 days), opposite results were obtained (
Figure 4). The combination of aphid infestation and virus infection showed a significant association in the control arenas, that was statistically significant at 7 days (
Figure 4), and a significant dissociation in the the presence of parasitoids after 2 days (
Figure 3).
Table 3.
Mean ± S.E. values of the displacement (δ) of aphids and CMV after 2 and 7 days in arenas with and without (control) parasitoids, followed by statistics according to Student’s t test (p < 0.05).
Table 3.
Mean ± S.E. values of the displacement (δ) of aphids and CMV after 2 and 7 days in arenas with and without (control) parasitoids, followed by statistics according to Student’s t test (p < 0.05).
| | Aphids | Virus |
|---|
| | Control | A. colemani | t | p | Control | A. colemani | t | p |
|---|
| 2 days | 1.9 ± 0.2 | 1.6 ± 0.2 | 0.758 | 0.491 | 0.9 ± 0.1 | 1.4 ± 0.6 | –0.690 | 0.528 |
| 7 days | 1.3 ± 0.1 | 1.5 ± 0.4 | –0.539 | 0.619 | 1.2 ± 0.4 | 1.0 ± 0.4 | 0.215 | 0.840 |
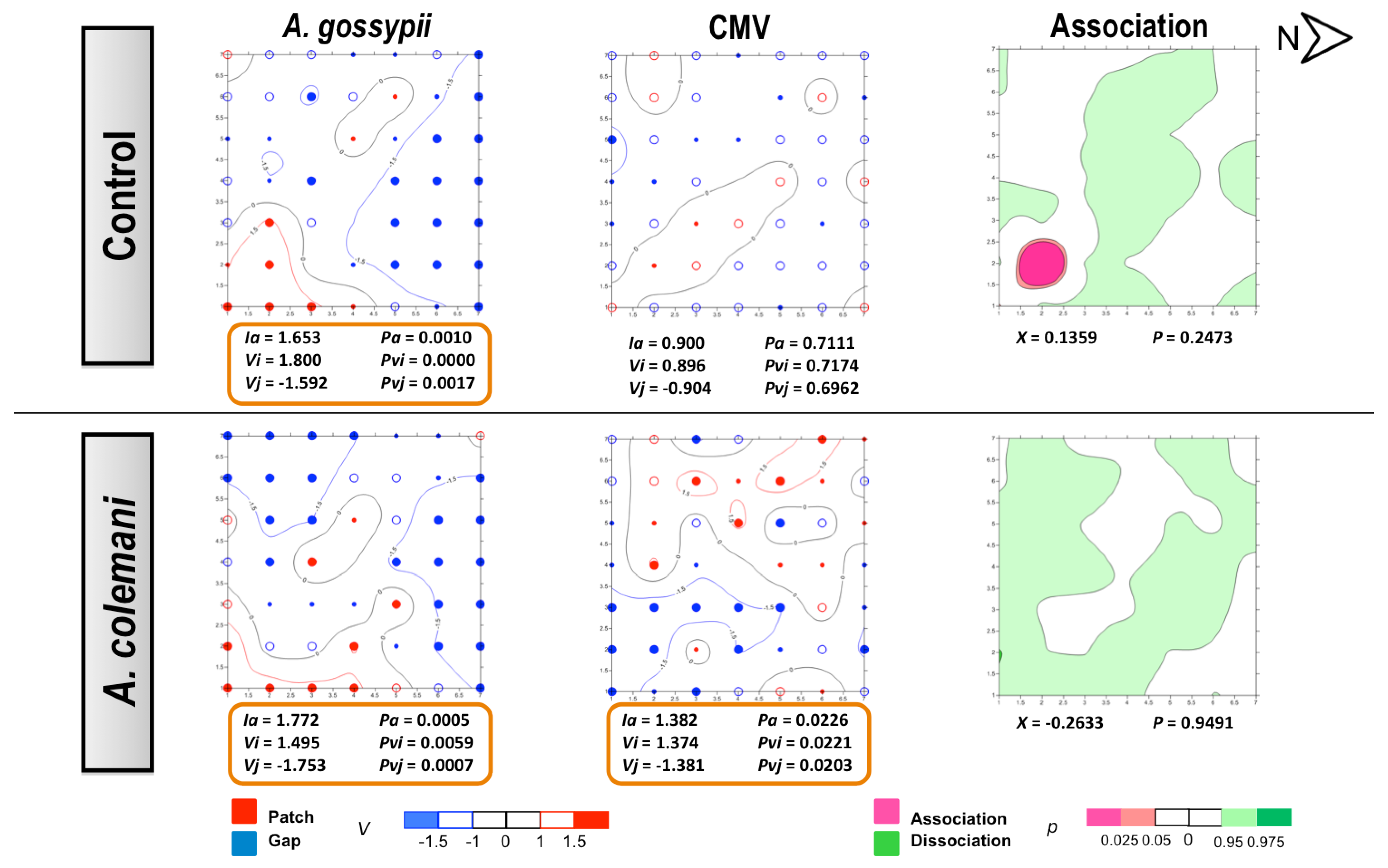
In our experiments,
A. gossypii showed the typical pattern of a colonizing aphid vector species because of its strong preference of cucumber as a host [
30]. However, the spread and incidence of the virus progressed differently at short and long times of evaluation. The contoured maps of CMV after 2 days revealed the typical pattern of a non-persistent virus in the absence of parasitoids (control cages). However, the spatial distribution was modified in the presence of
A. colemani and the clear consequence of the immediate disturbance of aphids, which promoted the distribution of CMV around the entire arena and its aggregation in several patches, in contrast with the few red spots indicating isolated infections under the control arenas. At 7 days, the infection in control arenas showed how the initial localised foci had merged in a larger patch, whereas a regular distribution (
Ia < 1) was found in the presence of parasitoids. The
A. gossypii-infested and CMV-infected plants were significantly associated in the control arenas, whilst
A. colemani induced dissociation between both agents, highlighting again the strong effect of natural enemies in the early dispersal of aphids as previously reported [
11,
14].
Figure 3.
Classed post maps of the spatial distribution of mean number of A. gossypii and cumulative number of CMV-infected plants (total number of infected plants per treatment) at 2 days, and contoured map of the association between CMV-infected plants and its vector, A. gossypii. Spots indicate individual test plants. Small filled spots represent clustering indices of 0 to ±0.99 (clustering below expectation), unfilled spots ±1 to ±1.49 (clustering slightly exceeds expectation) and large filled spots >1.5 or <1.5 (more than half as much as expectation). Red lines enclosing patch clusters are contours of v = 1.5 and blue lines are of v = –1.5. Black lines are zero-value contours, representing boundaries between patch and gap regions. The index of aggregation, Ia, the positive patch cluster index, vi, the negative gap cluster index, vj, and the index of spatial association, X, enclosed by an orange line are statistically significant. Letter N and arrow indicate north orientation.
Figure 3.
Classed post maps of the spatial distribution of mean number of A. gossypii and cumulative number of CMV-infected plants (total number of infected plants per treatment) at 2 days, and contoured map of the association between CMV-infected plants and its vector, A. gossypii. Spots indicate individual test plants. Small filled spots represent clustering indices of 0 to ±0.99 (clustering below expectation), unfilled spots ±1 to ±1.49 (clustering slightly exceeds expectation) and large filled spots >1.5 or <1.5 (more than half as much as expectation). Red lines enclosing patch clusters are contours of v = 1.5 and blue lines are of v = –1.5. Black lines are zero-value contours, representing boundaries between patch and gap regions. The index of aggregation, Ia, the positive patch cluster index, vi, the negative gap cluster index, vj, and the index of spatial association, X, enclosed by an orange line are statistically significant. Letter N and arrow indicate north orientation.
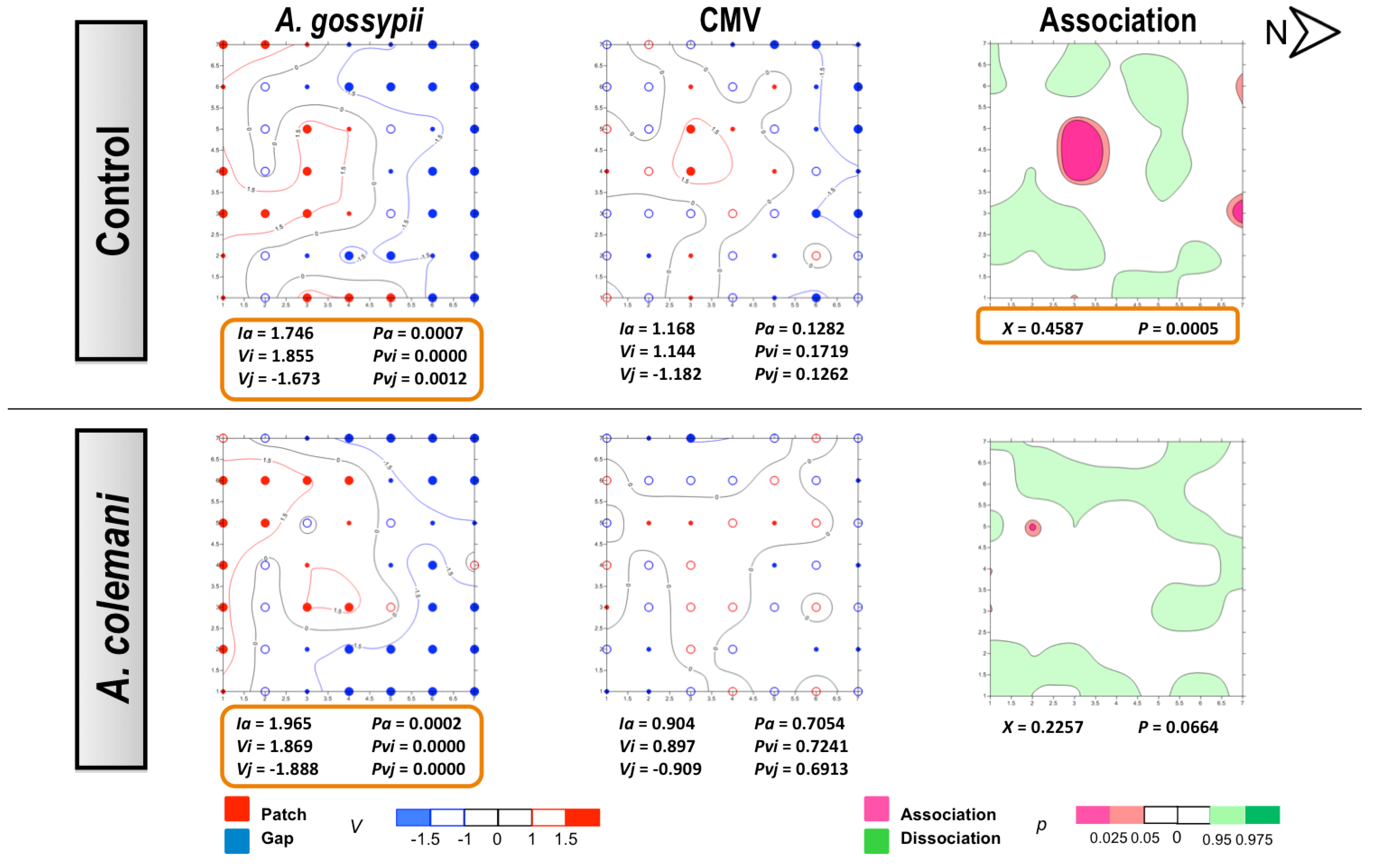
Figure 4.
Classed post maps of the spatial distribution of mean number of
A. gossypii and cumulative number of CMV-infected plants (total number of infected plants per treatment) at 7 days, and contoured map of the association between CMV-infected plants and its vector,
A. gossypii. Symbols and contours are as for
Figure 3.
Figure 4.
Classed post maps of the spatial distribution of mean number of
A. gossypii and cumulative number of CMV-infected plants (total number of infected plants per treatment) at 7 days, and contoured map of the association between CMV-infected plants and its vector,
A. gossypii. Symbols and contours are as for
Figure 3.
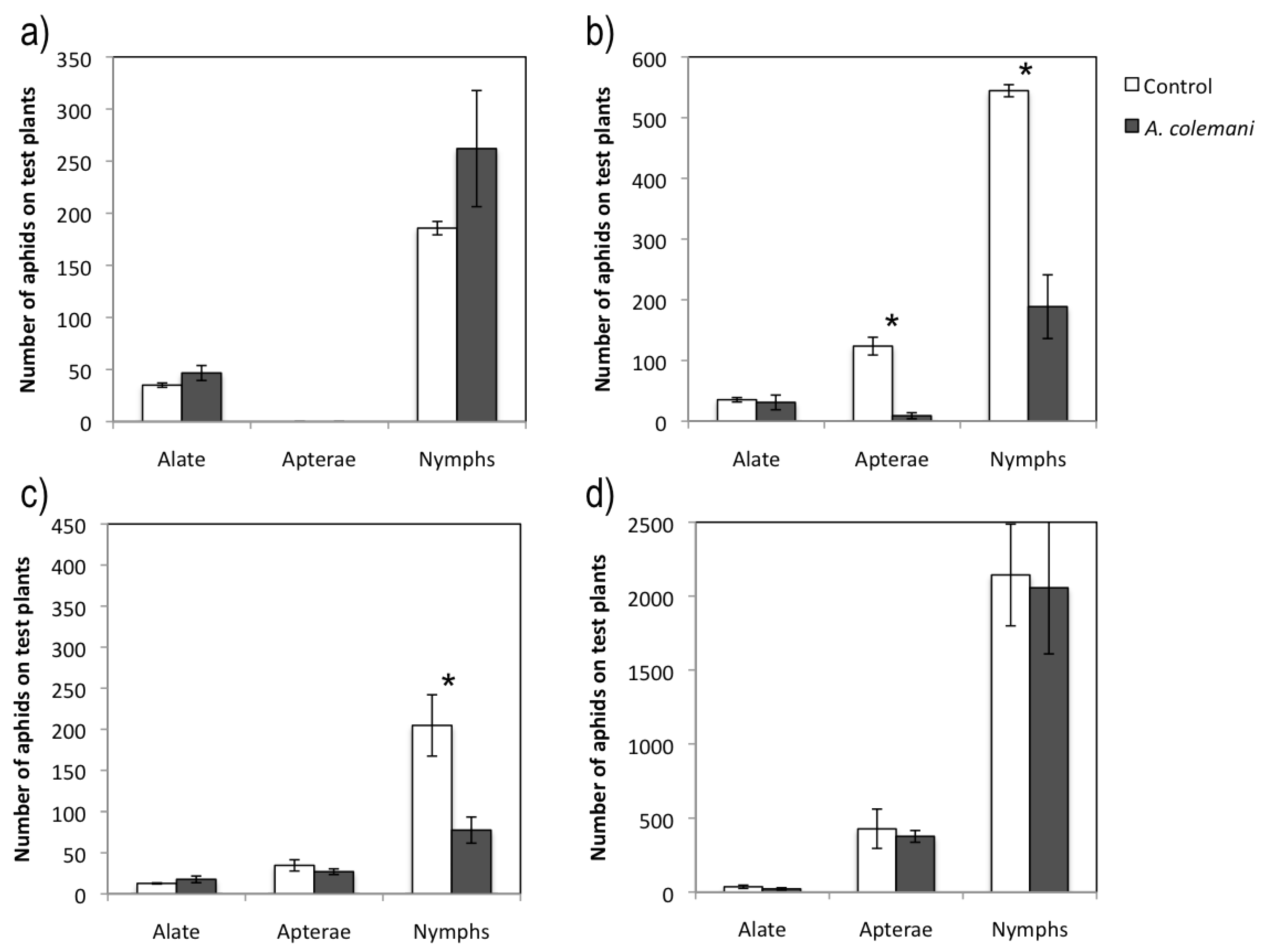
2.2. Effect of Aphidius Colemani on Aphid Dispersal and the Spread of Cucumber Aphid-Borne Yellows Virus
The short and long term experiments with CABYV was extended to 7 and 14 days, respectively, because its circulative mode of transmission requires longer acquisition and inoculation access periods than the non-circulative CMV. The central CABYV-infected plant contained more aphids in the control arenas than in those with
A. colemani, and significant differences in the number of apterae adults and nymphs were detected at 7 days, but not at 14 days (
Table 4). Moreover, it was possible to detect mummies in the virus source plant 14 days (8.3 ± 2.0) but not 7 days after parasitoid release. In general terms, there were more peripheral test plants occupied by a greater number of aphids in the control arenas than in those containing
A. colemani, with significant differences in the case of nymphs at 7 days (
t = 3.152; 4gl;
p = 0.034) (
Figure 1c). However these significant differences were not observed at 14 days (
Table 5,
Figure 1.d). As mentioned before for the CMV experiments, the number of offspring was much higher at long term (
Figure 1.d). Mummies (56.0 ± 21.6) and even parasitoid adults (3.3 ± 1.8) were found in peripheral test plants at 14 days again but not at 7 days. The mean incidence of CABYV when evaluated at short time (7 days) was higher in arenas containing
A. colemani compared to that observed in the control ones, although no significant differences were found. At long term, significantly fewer CABYV-infected plants were detected in arenas where
A. colemani were introduced (
X2 = 8.963;
p = 0.004) (
Figure 2).
The overall mean transmission rate of both viruses (CMV and CABYV) in the presence and absence of
A. colemani was not significantly different after 7 days. However, the presence of
A. colemani increased the rate the spread of CMV at short term (2 days) but reduced the spread of CABYV at long term (14 days) (
Figure 2). Moreover, the transmission rate of CABYV in control cages significantly increased at 14 days compared to 7 days from 11.1 ± 9.0% to 32.6 ± 8.9% (
X2 = 19.525;
p < 0.001) (
Figure 2).
Table 4.
Mean ± S.E. density (no. of individuals/plant) of adult morphs and nymphs in the CABYV-infected source plant at 7 and 14 days in cages with and without (control) A. colemani. Means in the same row followed by statistics in bold are significantly different according to Student’s t test (p < 0.05).
Table 4.
Mean ± S.E. density (no. of individuals/plant) of adult morphs and nymphs in the CABYV-infected source plant at 7 and 14 days in cages with and without (control) A. colemani. Means in the same row followed by statistics in bold are significantly different according to Student’s t test (p < 0.05).
| | 7 days | 14 days |
|---|
| | Control | A. colemani | t | p | Control | A. colemani | t | p |
|---|
| Alate | 36.0 ± 1.7 | 24.3 ± 7.8 | 1.443 | 0.222 | 20.0 ± 6.7 | 10.0 ± 1.2 | 1.359 | 0.246 |
| Apterae | 60.3 ± 9.9 | 17.0 ± 1.5 | 6.524 | 0.003 | 179.7 ± 54.3 | 185.0 ± 8.2 | 0.168 | 0.874 |
| Nymphs | 388.0 ± 39.1 | 160.0 ± 10.4 | 7.026 | 0.020 | 1624.7 ± 297.8 | 1212.0 ± 55.6 | 1.465 | 0.217 |
Table 5.
Mean±S.E. percentage of test plants occupied by one or more alate, apterae or nymphs (occupancy rate) in CABYV assays at 7 and 14 days in cages with and without (control) A. colemani followed by statistics according to a Chi-square 2x2 table test goodness of fit test (p <0.05).
Table 5.
Mean±S.E. percentage of test plants occupied by one or more alate, apterae or nymphs (occupancy rate) in CABYV assays at 7 and 14 days in cages with and without (control) A. colemani followed by statistics according to a Chi-square 2x2 table test goodness of fit test (p <0.05).
| | 7 days | 14 days |
|---|
| | Control | A. colemani | X 2 | p | Control | A. colemani | X 2 | p |
|---|
| Alate | 18.8 ± 1.2 | 24.3 ± 4.2 | 1.315 | 0.256 | 37.5 ± 9.8 | 30.6 ± 9.1 | 1.547 | 0.263 |
| Apterae | 20.9 ± 5.2 | 16.0 ± 3.0 | 1.133 | 0.293 | 63.9 ± 15.2 | 60.4 ± 1.2 | 0.369 | 0.627 |
| Nymphs | 27.8 ± 2.8 | 29.8 ± 4.6 | 0.152 | 0.670 | 75.0 ± 11.0 | 72.2 ± 0.7 | 0.286 | 0.596 |
Previous studies have described enhanced spread of persistent viruses in the presence of natural enemies [
12,
15], although the vector response might be influenced by the natural enemy’s searching habits [
16]. At 7 days, there were no differences in virus incidence between the two treatments, however significantly fewer CABYV-infected plants were observed at 14 days in the presence of
A. colemani, the same as seen by Smyrnioudis
et al. [
16] for both predators and parasitoids. Despite the fact that some studies show that virus transmission rate is not reduced by parasitoids [
40], Calvo and Fereres [
9] reported a reduction in the rate of spread of a circulative
Polerovirus due to a decrease in the life span of viruliferous aphids in the presence of parasitoids. In our study, we found, that mummification might have positively diminished the duration of aphids as active vectors after 14 days, so CABYV incidence was significantly reduced in comparison to that in the control cages. The recording of mummies and parasitoid adults provided evidence that
A. colemani was able to establish itself well in the experimental arenas. It has been reported an average of 10 days at a constant temperature of 25 °C for
A. colemani to complete its development [
41]. With the mean temperatures in which our experiments were performed, a new generation of parasitoids could have started to emerge after 14 days so that the reduction in vector populations may have possibly limited further virus spread.
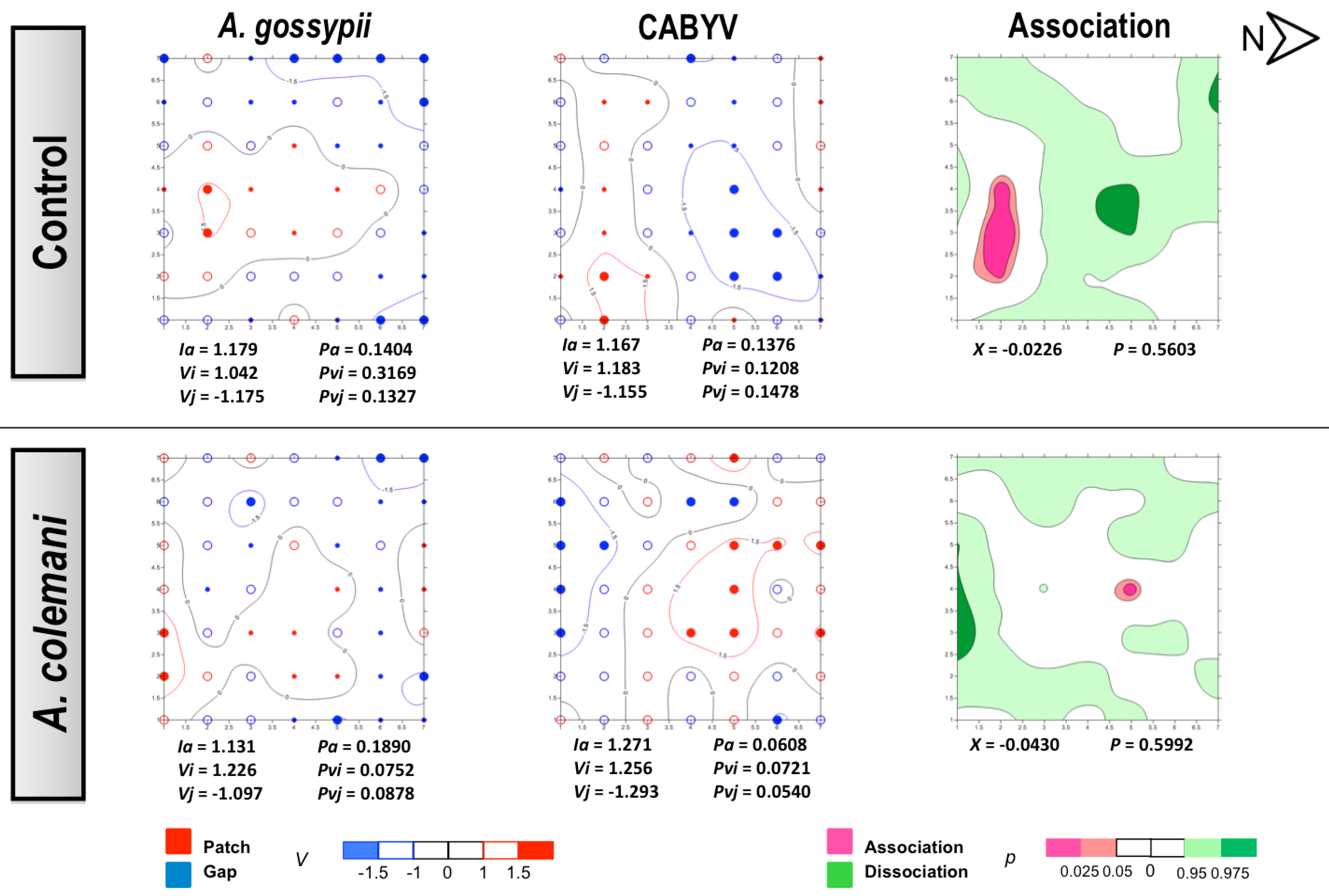
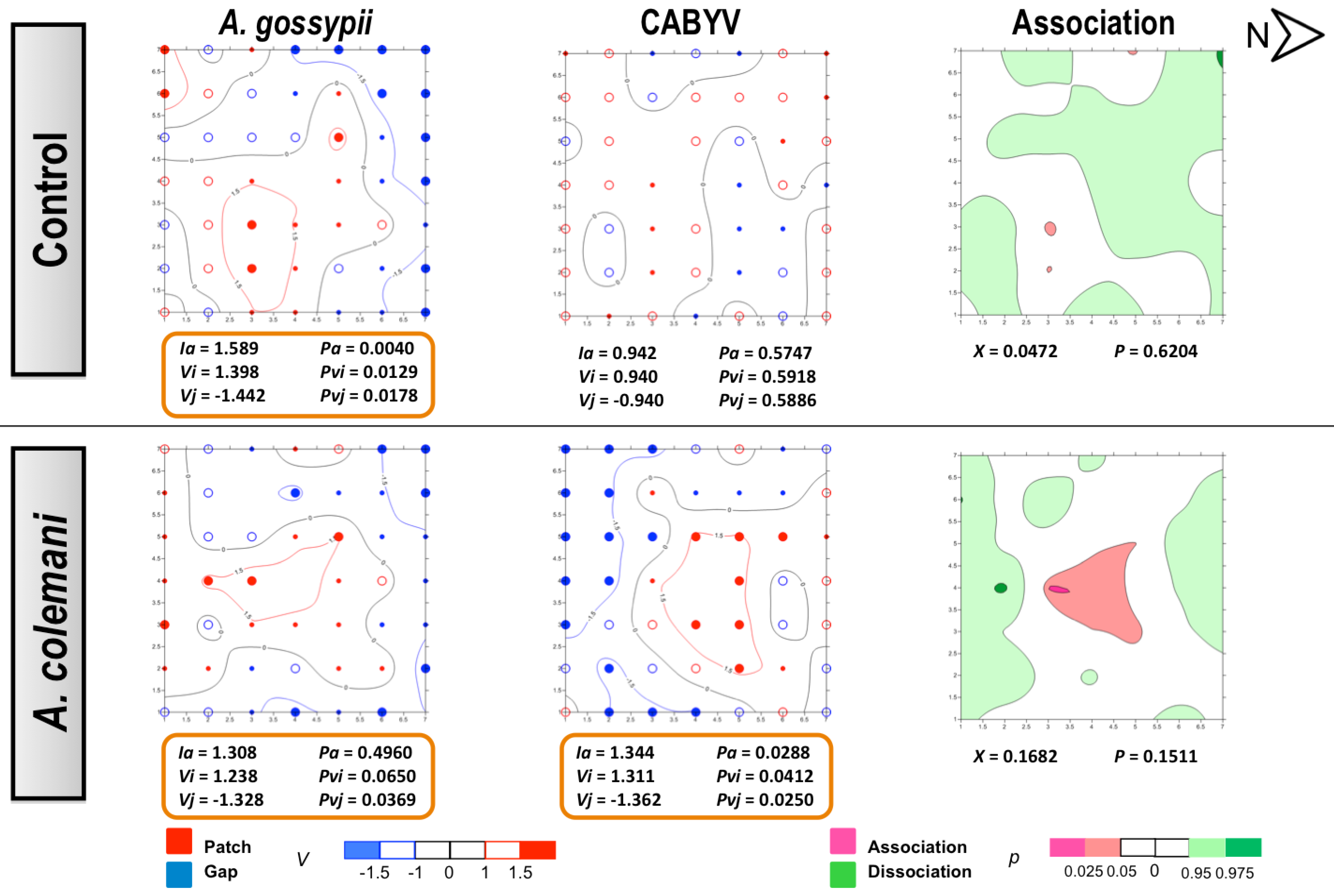
The same as in CMV experiments, no differences could be found for the mean displacement δ of both aphids and CABYV-infected plants under the two treatments (
Table 6). Aphids also showed an aggregated distribution in the CABYV assays, being significantly aggregated at 14 days (
Figure 6) as cucumber is an excellent host plant for
A. gossypii and large colonies are produced in a short period of time [
30]. Aphid spatial distribution showed a very similar pattern in the CABYV experiments when parasitoids were present, with population moving to the southern area of the cages and increasing the number of occupied plants occupied as time progressed (
Figure 5 and
Figure 6). In the control arenas, aphid distribution was limited to the south and center of the experimental cage at short times (
Figure 5) but reached almost all edges when aphids were allowed to stay 14 days, in parallel to what happened with the spread of CABYV (
Figure 6). Conversely, virus-infected plants were located in the northern area in arenas with parasitoids at 7 days (
Figure 5), and continued being restricted in the same area with a significantly aggregated distribution at 14 days (
Figure 6). When the spatial distribution of the virus and the vector were combined, a dissociation at 7 days and association at 14 days between aphid location and the position of CABYV-infected plants were recorded in both treatments (
Figure 5 and
Figure 6).
Table 6.
Mean ± S.E. values of the displacement (δ) of aphids and CABYV after 7 and 14 days in arenas with and without (control) A. colemani, followed by statistics according to Student’s t test (p < 0.05).
Table 6.
Mean ± S.E. values of the displacement (δ) of aphids and CABYV after 7 and 14 days in arenas with and without (control) A. colemani, followed by statistics according to Student’s t test (p < 0.05).
| | Aphids | Virus |
|---|
| | Control | A. colemani | t | p | Control | A. colemani | t | p |
|---|
| 7 days | 0.8 ± 0.1 | 1.2 ± 0.2 | –1.496 | 0.209 | 2.2 ± 1.0 | 1.0 ± 0.3 | 1.154 | 0.313 |
| 14 days | 0.8 ± 0.3 | 0.8 ± 0.1 | –0.112 | 0.916 | 1.4 ± 0.4 | 1.0 ± 0.0 | 1.117 | 0.326 |
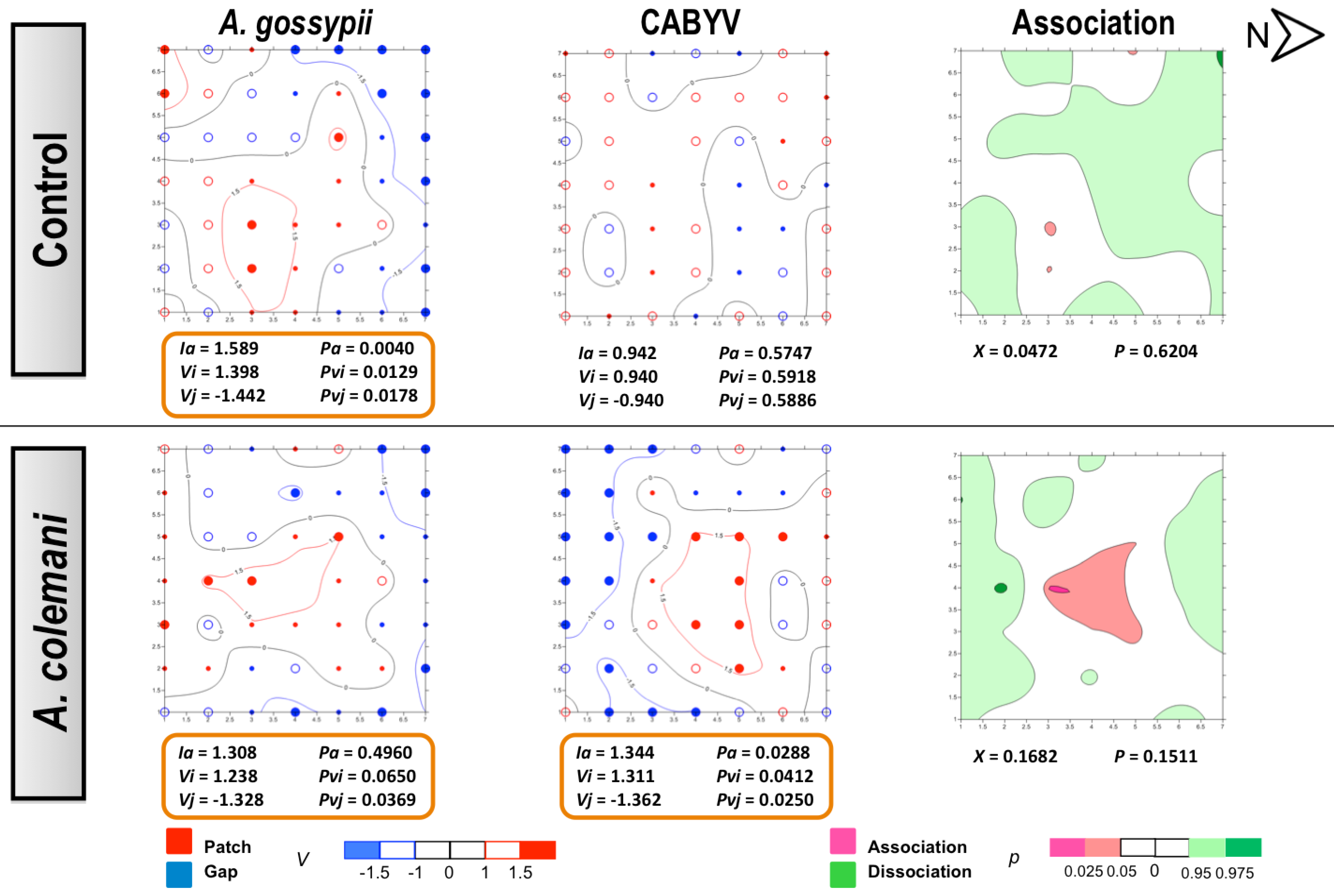
In the CABYV assays, there were major clustered areas of either patches or gaps of infected plants at 7 days in both treatments. Patch clusters seemed to be wider than gap ones in the
A. colemani treatment. Clusters of infected plants are frequently observed in viruses transmitted in a persistent circulative manner [
42]. Moreover, just two weeks were enough for aphids to expand CABYV to all the edges of the experimental arena in the absence of
A. colemani, whilst parasitoids limited the incidence of CABYV to specific patches. These results, together with the reduction in the rate of transmission of CABYV in the presence of
A. colemani after 14 days, prove the beneficial role of natural enemies at long term, specially when dealing with viruses transmitted in a persistent circulative manner.
The evaluation periods were selected depending on the type of transmission of each virus-vector combination and the life history of A. colemani. These periods cannot be comparable to the crop’s growth cycle that approximately lasts three to four months in commercial greenhouses. Making a prediction in a hypothetical scenario where the experiments had be run for a longer period is adventurous, but all plants would have possibly become infected by the persistent virus CABYV if parasitoids had not been released in the arena. Our data suggests that the spread of CABYV would be strongly constrained in the presence of parasitoids under a real field situation at long term because the mobility and population growth of aphid vectors would be jeopardised. However, it would be more difficult to predict the long-term consequences of parasitoid release in the case of the non-persistent virus CMV. Further research would be helpful to assess the effect of parasitism on aphid dispersal and virus transmission over time.
This study refers to a specific virus-vector-host-natural enemy complex, but natural enemy diversity occurring in cucurbit crops could influence the spread of both viruses in either direction. As a first approach, it will be desirable to carry out further studies with different natural enemies to study how this beneficial guild could modify viral dynamics and to investigate differences between their behaviour. As a second step, different virus-vector combinations could be evaluated to gain a better understanding of multitrophic interactions in pathosystems where whiteflies or thrips are present.
Figure 5.
Classed post maps of the spatial distribution of mean number of
A. gossypii and cumulative number of CABYV-infected plants (total number of infected plants per treatment) at 7 days, and contoured map of the association between CABYV-infected plants and its vector,
A. gossypii. Symbols and contours are as for
Figure 3.
Figure 5.
Classed post maps of the spatial distribution of mean number of
A. gossypii and cumulative number of CABYV-infected plants (total number of infected plants per treatment) at 7 days, and contoured map of the association between CABYV-infected plants and its vector,
A. gossypii. Symbols and contours are as for
Figure 3.
Figure 6.
Classed post maps of the spatial distribution of mean number of
A. gossypii and cumulative number of CABYV-infected plants (total number of infected plants per treatment) at 14 days, and contoured map of the association between CABYV-infected plants and its vector,
A. gossypii. Symbols and contours are as for
Figure 3.
Figure 6.
Classed post maps of the spatial distribution of mean number of
A. gossypii and cumulative number of CABYV-infected plants (total number of infected plants per treatment) at 14 days, and contoured map of the association between CABYV-infected plants and its vector,
A. gossypii. Symbols and contours are as for
Figure 3.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}