Epidemiology, Molecular Epidemiology and Evolution of Bovine Respiratory Syncytial Virus


{kind=link}
{kind=link}
Abstract
:1. Introduction
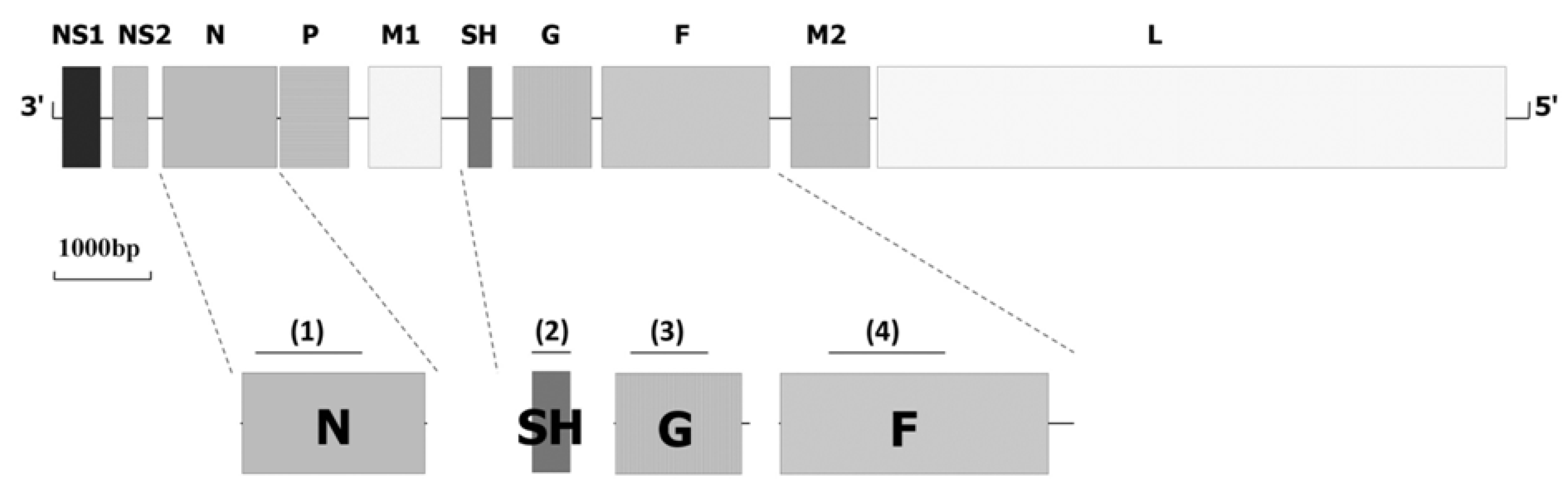
2. Epidemiology of BRSV
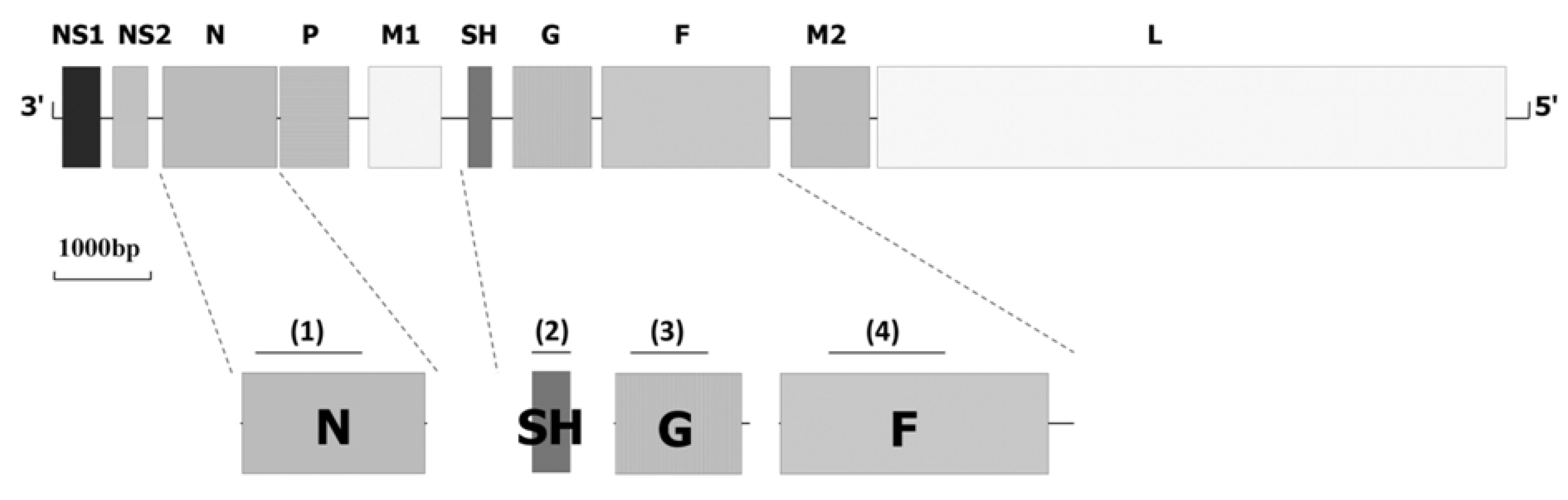
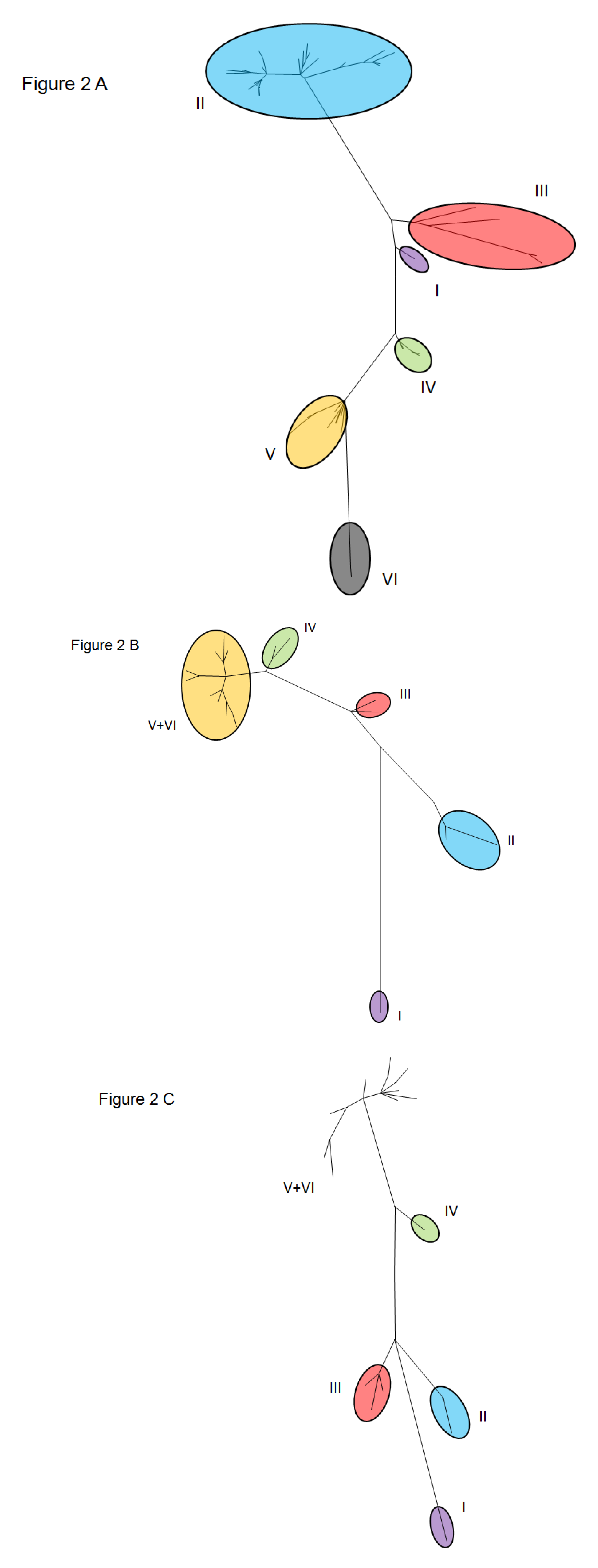
3. Molecular Epidemiology and Evolution of BRSV
4. Concluding Remarks
Acknowledgments
References and Notes
- Paccaud, M.F.; Jacquier, C. A respiratory syncytial virus of bovine origin. Arch. Gesamte Virusforsch. 1970, 30, 327–342. [Google Scholar] [CrossRef]
- Wellemans, G.; Leunen, J.; Luchsinger, E. Respiratory ailments of cattle: isolation of a virus (220/69) with serologic resemblance to the human respiratory syncytial virus. Ann. Med. Vet. 1970, 114, 89–93. [Google Scholar]
- Smith, R.A. Effects of feedlot disease on economics, production and carcass value. The American Association of Bovine Practitioners Proceedings 2000, 33, 125–128. [Google Scholar]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae: The Viruses and their Replication. In Fields Virology, 5th; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Williams: Philadelphia, PA, USA, 2007; Volume 2, pp. 1449–1496. [Google Scholar]
- Van der Poel, W.H.; Brand, A.; Kramps, J.A.; Van Oirschot, J.T. Respiratory syncytial virus infections in human beings and in cattle. J. Infection 1994, 29, 215–228. [Google Scholar] [CrossRef]
- Ames, T.R. The epidemiology of BRSV infection. Vet. Med. 1993, 88, 881–885. [Google Scholar]
- Masot, A.J.; Kelling, C.L.; Lopez, O.; Sur, J.H.; Redondo, E. In situ hybridization detection of bovine respiratory syncytial virus in the lung of experimentally infected lambs. Vet. Pathol. 2000, 37, 618–625. [Google Scholar] [CrossRef]
- Ohlson, A.; Emanuelson, U.; Traven, M.; Alenius, S. The relationship between antibody status to bovine corona virus and bovine respiratory syncytial virus and disease incidence, reproduction and herd characteristics in dairy herds. Acta Vet. Scand. 2010, 52, 37. [Google Scholar] [CrossRef]
- Ohlson, A.; Heuer, C.; Lockhart, C.; Traven, M.; Emanuelson, U.; Alenius, S. Risk factors for seropositivity to bovine coronavirus and bovine respiratory syncytial virus in dairy herds. Vet. Rec. 2010, 167, 201–266. [Google Scholar] [CrossRef]
- Saa, L.R.; Perea, A.; Jara, D.V.; Arenas, A.J.; Garcia-Bocanegra, I.; Borge, C.; Carbonero, A. Prevalence of and risk factors for bovine respiratory syncytial virus (BRSV) infection in non-vaccinated dairy and dual-purpose cattle herds in Ecuador. Trop. Anim. Health Prod. 2012, 44, 1423–1427. [Google Scholar] [CrossRef]
- Valarcher, J.F.; Taylor, G. Bovine respiratory syncytial virus infection. Vet. Res. 2007, 38, 153–180. [Google Scholar]
- Bidokhti, M.R.; Traven, M.; Ohlson, A.; Zarnegar, B.; Baule, C.; Belak, S.; Alenius, S.; Liu, L. Phylogenetic analysis of bovine respiratory syncytial viruses from recent outbreaks in feedlot and dairy cattle herds. Arch. Virol. 2012, 157, 601–607. [Google Scholar] [CrossRef]
- Elvander, M. Severe respiratory disease in dairy cows caused by infection with bovine respiratory syncytial virus. Vet. Rec. 1996, 138, 101–105. [Google Scholar] [CrossRef]
- Woldemeskel, M.; Kebede, E.; Yigezu, L.; Potgieter, L.N. Prevalence of bovine respiratory syncytial virus (BRSV) and bovine herpesvirus-4 (BHV-4) in cattle from Ethiopia. Dtsch. Tierarztl. Wochenschr. 2000, 107, 464–466. [Google Scholar]
- Collins, J.K.; Teegarden, R.M.; MacVean, D.W.; Salman; Smith, G.H.; Frank, G.R. Prevalence and specificity of antibodies to bovine respiratory syncytial virus in sera from feedlot and range cattle. Am. J. Vet. Res. 1988, 49, 1316–1319. [Google Scholar]
- Elazhary, M.A.; Silim, A.; Dea, S. Prevalence of antibodies to bovine respiratory syncytial virus, bovine viral diarrhea virus, bovine herpesvirus-1, and bovine parainfluenza-3 virus in sheep and goats in Quebec. Am. J. Vet. Res. 1984, 45, 1660–1662. [Google Scholar]
- Elazhary, M.A.; Roy, R.S.; Champlin, R.; Higgins, R.; Marsolais, G. Bovine respiratory syncytial virus in Quebec: antibody prevalence and disease outbreak. Can. J. Comp. Med. 1980, 44, 299–303. [Google Scholar]
- Baker, J.C.; Ames, T.R.; Markham, R.J. Seroepizootiologic study of bovine respiratory syncytial virus in a dairy herd. Am. J. Vet. Res. 1986, 47, 240–245. [Google Scholar]
- Kimman, T.G.; Zimmer, G.M.; Westenbrink, F.; Mars, J.; van Leeuwen, E. Epidemiological study of bovine respiratory syncytial virus infections in calves: influence of maternal antibodies on the outcome of disease. Vet. Rec. 1988, 123, 104–109. [Google Scholar]
- Hagglund, S.; Svensson, C.; Emanuelson, U.; Valarcher, J.F.; Alenius, S. Dynamics of virus infections involved in the bovine respiratory disease complex in Swedish dairy herds. Vet. J. 2006, 172, 320–328. [Google Scholar]
- Raaperi, K.; Bougeard, S.; Aleksejev, A.; Orro, T.; Viltrop, A. Association of herd BRSV and BHV-1 seroprevalence with respiratory disease and reproductive performance in adult dairy cattle. Acta Vet. Scand. 2012, 54, 4. [Google Scholar] [CrossRef]
- Bidokhti, M.R.; Traven, M.; Fall, N.; Emanuelson, U.; Alenius, S. Reduced likelihood of bovine coronavirus and bovine respiratory syncytial virus infection on organic compared to conventional dairy farms. Vet. J. 2009, 182, 436–440. [Google Scholar] [CrossRef]
- Stott, E.J.; Thomas, L.H.; Collins, A.P.; Crouch, S.; Jebbett, J.; Smith, G.S.; Luther, P.D.; Caswell, R. A survey of virus infections of the respiratory tract of cattle and their association with disease. J. Hyg. (Lond.) 1980, 85, 257–270. [Google Scholar] [CrossRef]
- Brodersen, B.W. Bovine respiratory syncytial virus. Vet. Clin. North. Am. Food Anim. Pract. 2010, 26, 323–333. [Google Scholar] [CrossRef]
- Baker, J.C.; Werdin, R.E.; Ames, T.R.; Markham, R.J.; Larson, V.L. Study on the etiologic role of bovine respiratory syncytial virus in pneumonia of dairy calves. J. Am. Vet. Med. Assoc. 1986, 189, 66–70. [Google Scholar]
- De Jong, M.C.; van der Poel, W.H.; Kramps, J.A.; Brand, A.; van Oirschot, J.T. Quantitative investigation of population persistence and recurrent outbreaks of bovine respiratory syncytial virus on dairy farms. Am. J. Vet. Res. 1996, 57, 628–633. [Google Scholar]
- Rossi, C.R.; Kiesel, G.K. Serological evidence for the association of bovine respiratory syncytial virus with respiratory tract disease in Alabama cattle. Infect. Immun. 1974, 10, 293–298. [Google Scholar]
- Smith, M.H.; Frey, M.L.; Dierks, R.E. Isolation, characterization, and pathogenicity studies of a bovine respiratory syncytial virus. Arch. Virol. 1975, 47, 237–247. [Google Scholar] [CrossRef]
- Baker, J.C.; Ames, T.R.; Markham, R.J. Serologic studies of bovine respiratory syncytial virus in Minnesota cattle. Am. J. Vet. Res. 1985, 46, 891–892. [Google Scholar]
- Lehmkuhl, H.D.; Gough, P.M. Investigation of causative agents of bovine respiratory tract disease in a beef cow-calf herd with an early weaning program. Am. J. Vet. Res. 1977, 38, 1717–1720. [Google Scholar]
- Grubbs, S.T.; Kania, S.A.; Potgieter, L.N. Prevalence of ovine and bovine respiratory syncytial virus infections in cattle determined with a synthetic peptide-based immunoassay. J. Vet. Diagn. Invest. 2001, 13, 128–132. [Google Scholar] [CrossRef]
- Fulton, R.W.; Purdy, C.W.; Confer, A.W.; Saliki, J.T.; Loan, R.W.; Briggs, R.E.; Burge, L.J. Bovine viral diarrhea viral infections in feeder calves with respiratory disease: interactions with Pasteurella spp., parainfluenza-3 virus, and bovine respiratory syncytial virus. Can. J. Vet. Res. 2000, 64, 151–159. [Google Scholar]
- Martin, S.W.; Bateman, K.G.; Shewen, P.E.; Rosendal, S.; Bohac, J.E. The frequency, distribution and effects of antibodies, to seven putative respiratory pathogens, on respiratory disease and weight gain in feedlot calves in Ontario. Can. J. Vet. Res. 1989, 53, 355–362. [Google Scholar]
- Gagea, M.I.; Bateman, K.G.; van Dreumel, T.; McEwen, B.J.; Carman, S.; Archambault, M.; Shanahan, R.A.; Caswell, J.L. Diseases and pathogens associated with mortality in Ontario beef feedlots. J. Vet. Diagn. Invest. 2006, 18, 18–28. [Google Scholar]
- Figueroa-Chavez, D.; Segura-Correa, J.C.; Garcia-Marquez, L.J.; Pescador-Rubio, A.; Valdivia-Flores, A.G. Detection of antibodies and risk factors for infection with bovine respiratory syncytial virus and parainfluenza virus 3 in dual-purpose farms in Colima, Mexico. Trop. Anim. Health Prod. 2012, 44, 1417–1421. [Google Scholar] [CrossRef]
- Solis-Calderon, J.J.; Segura-Correa, J.C.; Aguilar-Romero, F.; Segura-Correa, V.M. Detection of antibodies and risk factors for infection with bovine respiratory syncytial virus and parainfluenza virus-3 in beef cattle of Yucatan, Mexico. Prev. Vet. Med. 2007, 82, 102–110. [Google Scholar] [CrossRef]
- Almeida, R.S.; Domingues, H.G.; Spilki, F.R.; Larsen, L.E.; Hagglund, S.; Belak, S.; Arns, C.W. Circulation of bovine respiratory syncytial virus in Brazil. Vet. Rec. 2006, 158, 632–634. [Google Scholar] [CrossRef]
- Arns, C.W.; Campalans, J.; Costa, S.C.; Domingues, H.G.; D'Arce, R.C.; Almeida, R.S.; Coswig, L.T. Characterization of bovine respiratory syncytial virus isolated in Brazil. Braz. J. Med. Biol. Res. 2003, 36, 213–218. [Google Scholar]
- Jacobs, J.W.; Edington, N. Isolation of respiratory syncytial virus from cattle in Britain. Vet. Rec. 1971, 88, 694. [Google Scholar]
- Beaudeau, F.; Bjorkman, C.; Alenius, S.; Frossling, J. Spatial patterns of bovine corona virus and bovine respiratory syncytial virus in the Swedish beef cattle population. Acta Vet. Scand. 2010, 52, 33. [Google Scholar]
- Uttenthal, A.; Larsen, L.E.; Philipsen, J.S.; Tjornehoj, K.; Viuff, B.; Nielsen, K.H.; Nielsen, T.K. Antibody dynamics in BRSV-infected Danish dairy herds as determined by isotype-specific immunoglobulins. Vet. Microbiol. 2000, 76, 329–341. [Google Scholar] [CrossRef]
- Pardon, B.; De Bleecker, K.; Dewulf, J.; Callens, J.; Boyen, F.; Catry, B.; Deprez, P. Prevalence of respiratory pathogens in diseased, non-vaccinated, routinely medicated veal calves. Vet. Rec. 2011, 169, 278. [Google Scholar] [CrossRef]
- Hotchkiss, E.J.; Dagleish, M.P.; Willoughby, K.; McKendrick, I.J.; Finlayson, J.; Zadoks, R.N.; Newsome, E.; Brulisauer, F.; Gunn, G.J.; Hodgson, J.C. Prevalence of Pasteurella multocida and other respiratory pathogens in the nasal tract of Scottish calves. Vet. Rec. 2010, 167, 555–560. [Google Scholar] [CrossRef]
- Paton, D.J.; Christiansen, K.H.; Alenius, S.; Cranwell, M.P.; Pritchard, G.C.; Drew, T.W. Prevalence of antibodies to bovine virus diarrhoea virus and other viruses in bulk tank milk in England and Wales. Vet. Rec. 1998, 142, 385–391. [Google Scholar] [CrossRef]
- Raaperi, K.; Bougeard, S.; Aleksejev, A.; Orro, T.; Viltrop, A. Association of herd BHV-1 seroprevalence with respiratory disease in youngstock in Estonian dairy cattle. Res. Vet. Sci. 2012, 93, 641–648. [Google Scholar]
- Van Vuuren, M. Serological studies of bovine respiratory syncytial virus in feedlot cattle in South Africa. J. S. Afr. Vet. Assoc. 1990, 61, 168–169. [Google Scholar]
- Mahin, L.; Wellemans, G.; Shimi, A. Prevalence of antibodies to bovid herpesvirus 1 (IBR-IPV), bovine virus diarrhoea, bovine respiratory syncytial parainfluenza 3, adeno A and adeno B viruses in indigenous and imported Moroccan cattle. Ann. Rech. Vet. 1985, 16, 279–283. [Google Scholar]
- Mahin, L.; Wellemans, G. Serological evidence for the intervention of bovine respiratory syncytial virus in a respiratory disease outbreak in Moroccan cattle. Zentralbl. Veterinarmed. B 1982, 29, 76–79. [Google Scholar]
- Yesilbag, K.; Gungor, B. Seroprevalence of bovine respiratory viruses in North-Western Turkey. Trop. Anim. Health Prod. 2008, 40, 55–60. [Google Scholar] [CrossRef]
- Valarcher, J.F.; Schelcher, F.; Bourhy, H. Evolution of bovine respiratory syncytial virus. J. Virol. 2000, 74, 10714–10728. [Google Scholar]
- Deplanche, M.; Lemaire, M.; Mirandette, C.; Bonnet, M.; Schelcher, F.; Meyer, G. In vivo evidence for quasispecies distributions in the bovine respiratory syncytial virus genome. J. Gen. Virol. 2007, 88, 1260–1265. [Google Scholar] [CrossRef] [Green Version]
- Lam, T.T.; Hon, C.C.; Tang, J.W. Use of phylogenetics in the molecular epidemiology and evolutionary studies of viral infections. Critical reviews in clinical laboratory sciences 2010, 47, 5–49. [Google Scholar] [CrossRef]
- Khudyakov, Y. Coevolution and HBV drug resistance. Antivir. Ther. 2010, 15, 505–515. [Google Scholar]
- Spilki, F.R.; Almeida, R.S.; Domingues, H.G.; D'Arce, R.C.F.; Ferreira, H.L.; Campalans, J.; Costa, S.C.B.; Arns, C.W. Phylogenetic relationships of Brazilian bovine respiratory syncytial virus isolates and molecular homology modeling of attachment glycoprotein. Virus Res. 2006, 116, 30–37. [Google Scholar] [CrossRef]
- Samal, S.K.; Zamora, M.; McPhillips, T.H.; Mohanty, S.B. Molecular cloning and sequence analysis of bovine respiratory syncytial virus mRNA encoding the major nucleocapsid protein. Virology 1991, 180, 453–456. [Google Scholar] [CrossRef]
- Kovarcik, K.; Valentova, V. Bovine respiratory syncytial virus strains currently circulating in the Czech Republic are most closely related to Danish strains from 1995. Acta Virol. 2004, 48, 57–62. [Google Scholar]
- Lerch, R.A.; Stott, E.J.; Wertz, G.W. Characterization of bovine respiratory syncytial virus proteins and mRNAs and generation of cDNA clones to the viral mRNAs. J. Virol. 1989, 63, 833–840. [Google Scholar]
- Baker, J.C.; Wilson, E.G.; McKay, G.L.; Stanek, R.J.; Underwood, W.J.; Velicer, L.F.; Mufson, M.A. Identification of subgroups of bovine respiratory syncytial virus. J. Clin. Microbiol. 1992, 30, 1120–1126. [Google Scholar]
- Mallipeddi, S.K.; Samal, S.K. Structural difference in the fusion protein among strains of bovine respiratory syncytial virus. Vet. Microbiol. 1993, 36, 359–367. [Google Scholar] [CrossRef]
- Prozzi, D.; Walravens, K.; Langedijk, J.P.; Daus, F.; Kramps, J.A.; Letesson, J.J. Antigenic and molecular analyses of the variability of bovine respiratory syncytial virus G glycoprotein. J. Gen. Virol. 1997, 78, 359–366. [Google Scholar]
- Furze, J.; Wertz, G.; Lerch, R.; Taylor, G. Antigenic heterogeneity of the attachment protein of bovine respiratory syncytial virus. J. Gen. Virol. 1994, 75, 363–370. [Google Scholar] [CrossRef]
- Meyer, G.; Deplanche, M.; Schelcher, F. Human and bovine respiratory syncytial virus vaccine research and development. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 191–225. [Google Scholar] [CrossRef] [Green Version]
- Lerch, R.A.; Anderson, K.; Wertz, G.W. Nucleotide sequence analysis and expression from recombinant vectors demonstrate that the attachment protein G of bovine respiratory syncytial virus is distinct from that of human respiratory syncytial virus. J. Virol. 1990, 64, 5559–5569. [Google Scholar]
- Walravens, K.; Kettmann, R.; Collard, A.; Coppe, P.; Burny, A. Sequence comparison between the fusion protein of human and bovine respiratory syncytial viruses. J. Gen. Virol. 1990, 71, 3009–3014. [Google Scholar] [CrossRef]
- Samal, S.K.; Zamora, M. Nucleotide sequence analysis of a matrix and small hydrophobic protein dicistronic mRNA of bovine respiratory syncytial virus demonstrates extensive sequence divergence of the small hydrophobic protein from that of human respiratory syncytial virus. J. Gen. Virol. 1991, 72, 1715–1720. [Google Scholar]
- Zamora, M.; Samal, S.K. Sequence analysis of M2 mRNA of bovine respiratory syncytial virus obtained from an F-M2 dicistronic mRNA suggests structural homology with that of human respiratory syncytial virus. J. Gen. Virol. 1992, 73, 737–741. [Google Scholar] [CrossRef]
- Mallipeddi, S.K.; Samal, S.K. Sequence comparison between the phosphoprotein mRNAs of human and bovine respiratory syncytial viruses identifies a divergent domain in the predicted protein. J. Gen. Virol. 1992, 73, 2441–2444. [Google Scholar] [CrossRef]
- Mallipeddi, S.K.; Samal, S.K. Sequence variability of the glycoprotein gene of bovine respiratory syncytial virus. J. Gen. Virol. 1993, 74, 2001–2004. [Google Scholar]
- Langedijk, J.P.; Meloen, R.H.; Taylor, G.; Furze, J.M.; van Oirschot, J.T. Antigenic structure of the central conserved region of protein G of bovine respiratory syncytial virus. J. Virol. 1997, 71, 4055–4061. [Google Scholar]
- Stine, L.C.; Hoppe, D.K.; Kelling, C.L. Sequence conservation in the attachment glycoprotein and antigenic diversity among bovine respiratory syncytial virus isolates. Vet. Microbiol. 1997, 54, 201–221. [Google Scholar] [CrossRef]
- Elvander, M.; Vilcek, S.; Baule, C.; Uttenthal, A.; Ballagi-Pordany, A.; Belak, S. Genetic and antigenic analysis of the G attachment protein of bovine respiratory syncytial virus strains. J. Gen. Virol. 1998, 79, 2939–2946. [Google Scholar]
- Larsen, L.E.; Uttenthal, A.; Arctander, P.; Tjornehoj, K.; Viuff, B.; Rontved, C.; Ronsholt, L.; Alexandersen, S.; Blixenkrone-Moller, M. Serological and genetic characterisation of bovine respiratory syncytial virus (BRSV) indicates that Danish isolates belong to the intermediate subgroup: no evidence of a selective effect on the variability of G protein nucleotide sequence by prior cell culture adaption and passages in cell culture or calves. Vet. Microbiol. 1998, 62, 265–279. [Google Scholar] [CrossRef]
- Kovarcik, K.; Valentova, V. Bovine respiratory syncytial virus strains currently circulating in the Czech Republic are most closely related to Danish strains from 1995. Acta Virol. 2004, 48, 57–62. [Google Scholar]
- Larsen, L.E.; Tjornehoj, K.; Viuff, B. Extensive sequence divergence among bovine respiratory syncytial viruses isolated during recurrent outbreaks in closed herds. J. Clin. Microbiol. 2000, 38, 4222–4227. [Google Scholar]
- Valentova, V.; Antonis, A.F.; Kovarcik, K. Restriction enzyme analysis of RT-PCR amplicons as a rapid method for detection of genetic diversity among bovine respiratory syncytial virus isolates. Vet. Microbiol. 2005, 108, 1–12. [Google Scholar] [CrossRef]
- Yaegashi, G.; Seimiya, Y.M.; Seki, Y.; Tsunemitsu, H. Genetic and antigenic analyses of bovine respiratory syncytial virus detected in Japan. J. Vet. Med. Sci. 2005, 67, 145–150. [Google Scholar]
- Spilki, F.R.; Almeida, R.S.; Domingues, H.G.; D'Arce, R.C.; Ferreira, H.L.; Campalans, J.; Costa, S.C.; Arns, C.W. Phylogenetic relationships of Brazilian bovine respiratory syncytial virus isolates and molecular homology modeling of attachment glycoprotein. Virus. Res. 2006, 116, 30–37. [Google Scholar] [CrossRef]
- Nettleton, P.F.; Gilray, J.A.; Caldow, G.; Gidlow, J.R.; Durkovic, B.; Vilcek, S. Recent isolates of bovine respiratory syncytial virus from Britain are more closely related to isolates from USA than to earlier British and current mainland European isolates. J. Vet. Med. B Infect. Dis. Vet. Public Health 2003, 50, 196–199. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sarmiento-Silva, R.E.; Nakamura-Lopez, Y.; Vaughan, G. Epidemiology, Molecular Epidemiology and Evolution of Bovine Respiratory Syncytial Virus. Viruses 2012, 4, 3452-3467. https://doi.org/10.3390/v4123452
Sarmiento-Silva RE, Nakamura-Lopez Y, Vaughan G. Epidemiology, Molecular Epidemiology and Evolution of Bovine Respiratory Syncytial Virus. Viruses. 2012; 4(12):3452-3467. https://doi.org/10.3390/v4123452
Chicago/Turabian StyleSarmiento-Silva, Rosa Elena, Yuko Nakamura-Lopez, and Gilberto Vaughan. 2012. "Epidemiology, Molecular Epidemiology and Evolution of Bovine Respiratory Syncytial Virus" Viruses 4, no. 12: 3452-3467. https://doi.org/10.3390/v4123452