Use of the Syrian Hamster as a New Model of Ebola Virus Disease and Other Viral Hemorrhagic Fevers

Abstract
:1. Introduction
2. Ebola Virus Disease (Humans): Clinical Presentation and Pathogenesis
3. Existing Animal Models of EVD

{kind=link}
{kind=link}
{kind=link}
| Coagulation Parameter | Rhesus Macaque b | Syrian Hamster c | Guinea Pig d | Mouse c |
|---|---|---|---|---|
| Increased prothrombin time (PT) | (++) [26,27] | (+++) | (+++) | (-) [20,27] |
| Increased activated partial thromboplastin time (aPTT) | (++) [26,27] | (+++) | (++) | (-)[20,27] |
| Increased thrombin time (TT) | (++) [26,27] | (++) | ND | ND |
| Late hypofibrinogenemia | (+++) [26,27] | (++) | (-) (increased fibrinogen) | (-/+) [20,27] |
| Decreased protein C activity % | (+++) [26,27] | (+++) | ND | ND |
| Thrombocytopenia | (++) [26,27] | (++) | (++) e [20] / (+++) f [21] | (++) [20,28] |
| Vaccines | Immunization Schedule | Mouse Model | Guinea Pig Model | NHP Model |
|---|---|---|---|---|
| Virus Vectors | ||||
| HPIV3 Immunogens | Guinea Pigs: |
|
| |
| HPIV3- NHPs: | ||||
| ||||
| VSV ∆GP Immunogens | Immunocompetent Mice: |
|
|
|
| EBOV GP attenuated [32,33,34,35,36,37,38] |
| |||
| NOD-SCID Mice: IP 2 × 105 PFU of VSV∆GP/EBOV GP | ||||
| Guinea Pigs: IP 2 × 105 PFU-1–2 doses of VSV∆GP/EBOV GP [32] | ||||
| HIV + NHPs: IM 1 × 107 PFU [34] | ||||
| Immunocompetent NHPs: | ||||
| ||||
| VV Immunogens | Guinea Pigs: SC 107 of VV/EBOV GP–3 doses [39] |
|
| |
| NHPs: SC of VV/EBOV GP–3 doses [24] | ||||
| Virus-like Particles (VLPs) | ||||
| VEEV RNA (VRP) encoding: | Mice: |
| ||
| Guinea Pigs: SC 107 IU of VRP EBOV GP, NP, or GP+NP–2 or 3 doses [41,44] | ||||
| NHPs: SC 2 x 106 FFU of VRP EBOV GP, NP or GP+NP–3 doses [24] | ||||
| Ebola Virus Vaccines | ||||
| EBOV | Mice | |||
| ||||
| NHPs: | ||||
| Peri- exposure Treatment | Dose and Route of Administration | Mouse Model | Guinea Pig Model | NHP Model |
|---|---|---|---|---|
| Virus Vectors | ||||
| VSV ∆GP Immunogens | Mice: IP 2 x 105 VSV ∆GP/EBOV GP PFU -1 day before or +30 minutes or +1 day after challenge [51] |
|
|
|
| ||||
| Guinea pigs: IP 2 x 105 VSV ∆GP/EBOV GP PFU -24 hours or +1 or 24 hours [51] | ||||
| NHPs: IM 2 x 107 PFU of VSV ∆GP/EBOV GP [51] +20–30 minutes | ||||
| Passive Immunity | ||||
| Pooled immune serum to live EBOV [46,52] | Mice: IP 1 mL of antisera (anti-EBOV IgG titers of ≥6,400) -1 day or + 1 day [46] |
|
| |
| NHPs: IV 6 mL/kg whole blood immediately after challenge and +3 or 4 days (anti-EBOV IgG ELISA titers of 100,000) [52] | ||||
| Passive Immunity | ||||
| Purified polyclonal IgG antibody against: | Mice: |
|
|
|
| ||||
| Guinea Pigs: IM 1 mL/kg of horse IgG + several minutes and +3 days, or +4 days only [54] | ||||
| NHPs: | ||||
| mAb EBOV GP-specific | Guinea Pigs: |
|
| |
| ||||
| NHPs: IV 50 mg/kg -1 day and +4 days [56] | ||||
| Antiviral Agents | ||||
| Antisense Phosphorodiamidate morpholino oligomers (PMO) [57,58,59,60] | Mice: |
|
| |
| ||||
| Guinea Pigs: IP 10 mg of each PMO targeting VP24, VP35 or L -1 day or +1 or 6 days after challenge [58] | Reduced viremia and release of IL-6 and MCP-1 with PMOs targeting VP24 and VP35 | |||
| Antiviral Agents | ||||
| Antisense PMOs (continued) | s: |
| ||
| ||||
4. History of Outbred Strains of Syrian Hamster in U.S.
5. Syrian Hamsters as a Model of Viral Hemorrhagic Fever Diseases

| Pathology and laboratory abnormalities | Virus |
|---|---|
| Spleen | |
| Punta Toro [83] |
| Pirital [84,86,87] | |
| Marburg [73] | |
| Ebola [27] |
| Gabek Forest [83] |
| Yellow fever [81,82,85] | |
| Ebola [27] |
| Andes [75] |
| Marburg [73] |
| Ebola [27] |
| Liver | |
| Punta Toro [83] |
| Ebola [27] |
| Gabek Forest [83] |
| Yellow fever [81,82,85] |
| Pirital [84,86,87] |
| Andes [75,76,79] | |
| Maporal [78] |
| Yellow Fever [81,85] |
| Pirital [84,86] | |
| Yellow Fever [81] |
| Punta Toro [83] |
| Gabek Forest [83] | |
| Yellow Fever [81,85] | |
| Pichinde [80] | |
| Pirital [84,86] | |
| Lymph nodes | |
| Punta Toro [83] |
| Ebola [27] |
| Ebola [27] |
| Andes [76] |
| Marburg [73] |
| Ebola [27] |
| Ebola [27] |
| Lung | |
| Yellow fever[81] |
| Andes[75,79] |
| Maporal [78] | |
| Punta Toro [83] |
| Pirital [87] | |
| Gabek Fores [83] |
| Pirital [84,86] | |
| Andes [76] | |
| Marburg [73] |
| Kidney | |
| Yellow Fever [82,85] |
| Marburg [73] |
| Gabek Forest [83] |
| Maporal [78] |
| Pirital [84,86] |
| Vascular dysregulation | |
| Andes [75,79] |
| Pichinde [80] | |
| Yellow fever [80] | |
| Maporal [78] | |
| Pichinde [80] |
| Yellow fever [81] | |
| Pirital [86] | |
| Coagulopathy | |
| Yellow Fever [81] |
| Pirital [84,86] | |
| Andes [76] | |
| Ebola [27] | |
| Pirital [86] |
| Ebola [27] | |
| Yellow fever [81] |
| Ebola [27] | |
| Andes [76] | |
| Pirital [84] | |
| Pirital [84] |
| Ebola [27] |
| Andes [76] |
| Pirital [84,86] |
| Andes [75] |
| Yellow fever [81] | |
| Hematological abnormalities | |
| Yellow fever [81] |
| Pichinde [80] |
| Pirital [86] | |
| Andes [75] | |
| Andes [75] |
| Cytokines | |
| Pichinde [80] |
| Andes [76] | |
| Andes [76] |
| Yellow fever [85] | |
| Ebola [27] | |
6. Syrian Hamsters as an Ebola Virus Disease Model
| Macaque | Guinea pig | Hamster | Mouse | |
|---|---|---|---|---|
| Hallmarks of Human Disease | ||||
| Target cells/organs | + | + | + | + |
| Cytokine dysregulation | + | ND a | + | +/- |
| Lymphocyte apoptosis | + | ND a | + | + |
| Coagulation dysfunction | ++ | + | ++ | +/- |
| Advantages/Disadvantages | ||||
| Availability | - | +/- | + | + |
| Ease of handling | - | +/- | + | + |
| Research reagents | ++ | - | +/++ | +++ |
| Pathogenicity of MA-EBOV | ++ [20] b | + | +++ | +++ |
| Liver | Spleen | |||||
|---|---|---|---|---|---|---|
| Hepatocellular degeneration and necrosis | Inflammation | Fibrin | Lymphoid necrosis and loss | Inflammation | Fibrin | |
| Mouse | Diffuse, random | Neutrophilic | Little | Multifocal, mild | Neutrophilic | Little |
| Guinea pig | Diffuse, random | Neutrophilic and histiocytic | Little to moderate | Diffuse, severe | Neutrophilic | Little to moderate |
| Hamster | Diffuse, midzonal | Neutrophilic | Little | Diffuse, moderate to severe | Neutrophilic | Little |
| NHP | Diffuse, random | Neutrophilic | Abundant | Diffuse, severe | Neutrophilic | Abundant |

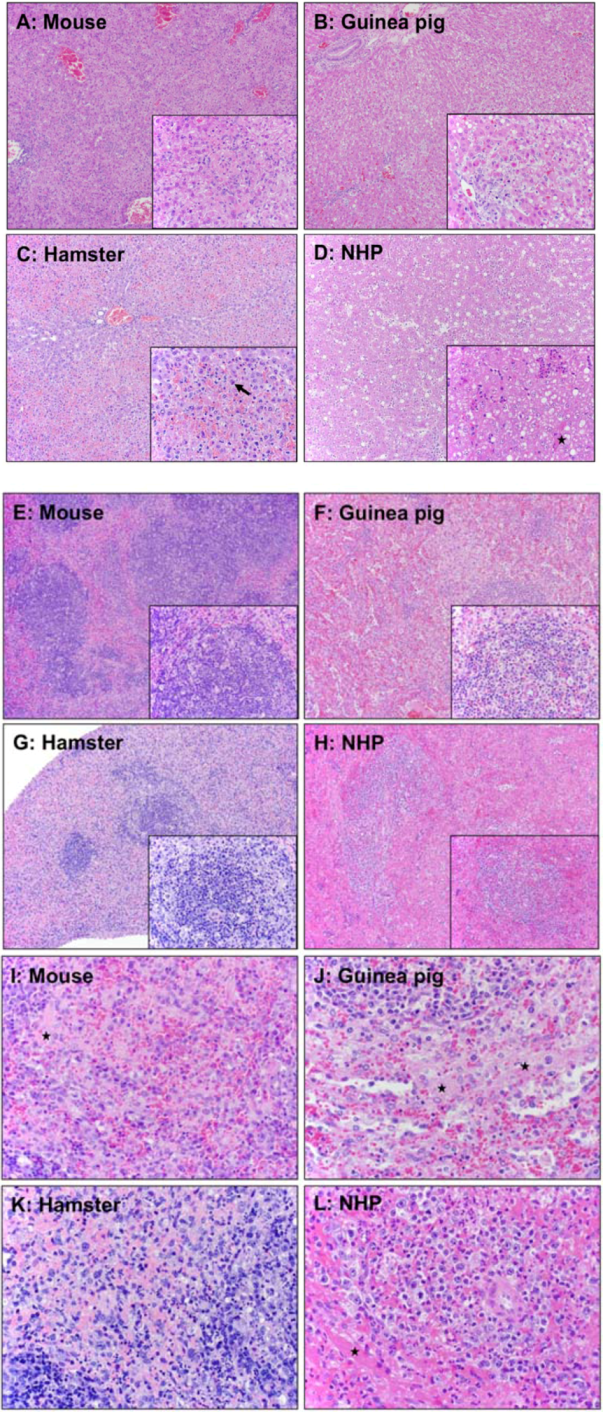
7. Reagent and Assay Development
| Cross-reactive antibodies | |
| Mouse anti-mouse/rat MHC II [74] | Rat anti-mouse CD4 [74,96,97] |
| Mouse anti-mouse Thy1.2 [74] | |
| Mouse anti-rat CD8, CD8β [74,97] | |
| Mouse, rabbit, and goat anti-hamster antibodies | |
| Mouse dendritic cell marker [95] | Mouse CD18 [95] |
| Mouse pan lymphocyte [95] | Mouse MHC II [95] |
| Mouse T cell [95] | Rabbit IgG [95] |
| Mouse B cell [95] | Rabbit IgM [74] |
| Mouse CD45 [95] | Goat IgG [74,78,96,97] |
| Cross-reactive cytokine, chemokine, and serum factor assays | |
| Rat GM-CSF [94] | Mouse / Rat VCAM-1 [76,80] |
| Rat Leptin [94] | Mouse / Rat vWF [80] |
| Rat GRO/KC [94] | Mouse / Rat VEGF [80] |
| Rat / Mouse IL-1α [80,94] | Mouse / Rat MDC [80] |
| Mouse MIG [94] | Mouse / Rat SCF [80] |
| Mouse IL-13 [94] | Mouse GCP-2 [80] |
| Mouse / Rat IP-10 [76,80] | Mouse MCP-3 [80] |
| Mouse /Rat M-CSF [76,80] | Mouse MIP-2 [80] |
| Mouse /Rat MCP-1 [76,80] | Mouse MIP-3β [80] |
| Mouse Fibrinogen [80] | Mouse AST [80] |
| Cross-reactive microarray hybridization | |
| DNA Microarrays | MicroRNA Microarrays |
| |
| Mouse genes [100,101] | |
8. Future Perspectives
Acknowledgments
Conflict of Interest
Supplementary Files
Supplementary File 1References
- Schnittler, H.J.; Feldmann, H. Viral hemorrhagic fever--a vascular disease? Thromb. Haemostasis 2003, 89, 967–972. [Google Scholar]
- Zaki, S.R.; Goldsmith, C.S. Pathologic features of filovirus infections in humans. Curr. Top. Microbiol. Immunol. 1999, 235, 97–116. [Google Scholar]
- Bwaka, M.A.; Bonnet, M.J.; Calain, P.; Colebunders, R.; De Roo, A.; Guimard, Y.; Katwiki, K.R.; Kibadi, K.; Kipasa, M.A.; Kuvula, K.J.; et al. Ebola hemorrhagic fever in Kikwit, Democratic Republic of the Congo: clinical observations in 103 patients. J. Infect. Dis. 1999, 179, S1–S7. [Google Scholar] [CrossRef]
- Formenty, P.; Hatz, C.; Le Guenno, B.; Stoll, A.; Rogenmoser, P.; Widmer, A. Human infection due to Ebola virus, subtype Cote d'Ivoire: Clinical and biologic presentation. J. Infect. Dis. 1999, 179, S48–S53. [Google Scholar]
- Kuhn, J.H. Filoviruses. A compendium of 40 years of epidemiological, clinical, and laboratory studies. Arch. Virol. Suppl. 2008, 20, 13–360. [Google Scholar]
- Smith, D.H.; Francis, D.P.; Simpson, D.I. African Haemorrhagic Fever in the Southern Sudan, 1976: The Clinical Manifestations. Ebola Virus Haemorrhagic Fever—Proceedings of an International Colloquium on Ebola Virus Infection and Other Haemorrhagic Fevers; Pattyn, S.R., Ed.; Elsevier North Holland Biomedical Press: Amsterdam, Netherlands, 1978; pp. 28–31. Available online: http://www.itg.be/internet/ebola/default.htm (accessed on 14 June 2012).
- Sureau, P.H. Firsthand clinical observations of hemorrhagic manifestations in Ebola hemorrhagic fever in Zaire. Rev. Infect. Dis. 1989, 11, S790–S793. [Google Scholar] [CrossRef]
- World Health Organization. Ebola haemorrhagic fever in Sudan, 1976. Bull. World Health Organ. 1978, 52, pp. 247–250. Available online: http://whqlibdoc.who.int/bulletin/1978/Vol56-No2/bulletin_1978_56%282%29_247–270.pdf (accessed on 19 June 2012).
- Rollin, P.E.; Bausch, D.G.; Sanchez, A. Blood chemistry measurements and D-Dimer levels associated with fatal and nonfatal outcomes in humans infected with Sudan Ebola virus. J. Infect. Dis. 2007, 196, S364–S371. [Google Scholar] [CrossRef]
- Richards, G.A.; Murphy, S.; Jobson, R.; Mer, M.; Zinman, C.; Taylor, R.; Swanepoel, R.; Duse, A.; Sharp, G.; De La Rey, I.C.; et al. Unexpected Ebola virus in a tertiary setting: clinical and epidemiologic aspects. Crit. Care Med. 2000, 28, 240–244. [Google Scholar]
- Dietrich, M.; Schumacher, H.H.; Peters, D.; Knobloch, J. Human pathology of Ebola (MARIDI) virus Infection in the Sudan. Ebola Virus Haemorrhagic Fever—Proceedings of an International Colloquium on Ebola Virus Infection and Other Haemorrhagic Fevers; Pattyn, S.R., Ed.; Elsevier North Holland Biomedical Press: Amsterdam, Netherlands, 1978; pp. 37–41. Available online: http://www.itg.be/internet/ebola/default.htm (accessed on 14 June 2012).
- Sanchez, A.; Lukwiya, M.; Bausch, D.; Mahanty, S.; Sanchez, A.J.; Wagoner, K.D.; Rollin, P.E. Analysis of human peripheral blood samples from fatal and nonfatal cases of Ebola (Sudan) hemorrhagic fever: Cellular responses, virus load, and nitric oxide levels. J. Virol. 2004, 78, 10370–10377. [Google Scholar]
- Kortepeter, M.G.; Bausch, D.G.; Bray, M. Basic clinical and laboratory features of filoviral hemorrhagic fever. J. Infect. Dis. 2011, 204, S810–S816. [Google Scholar] [CrossRef]
- Murphy, F.A. Pathology of Ebola virus infection. Ebola Virus Haemorrhagic Fever—Proceedings of an International Colloquium on Ebola Virus Infection and Other Haemorrhagic Fevers; Pattyn, S.R., Ed.; Elsevier North Holland Biomedical Press: Amsterdam, Netherlands, 1978; pp. 41–53. Available online: http://www.itg.be/internet/ebola/default.htm (accessed on 14 June 2012).
- Baize, S.; Leroy, E.M.; Georges, A.J.; Georges-Courbot, M.C.; Capron, M.; Bedjabaga, I.; Lansoud-Soukate, J.; Mavoungou, E. Inflammatory responses in Ebola virus-infected patients. Clin. Exp. Immunol. 2002, 128, 163–168. [Google Scholar] [CrossRef]
- Wauquier, N.; Becquart, P.; Padilla, C.; Baize, S.; Leroy, E.M. Human fatal zaire ebola virus infection is associated with an aberrant innate immunity and with massive lymphocyte apoptosis. PLoS Neglected Tropical Diseases 2010, 4, e837. [Google Scholar] [CrossRef]
- Villinger, F.; Rollin, P.E.; Brar, S.S.; Chikkala, N.F.; Winter, J.; Sundstrom, J.B.; Zaki, S.R.; Swanepoel, R.; Ansari, A.A.; Peters, C.J. Markedly elevated levels of interferon (IFN)-gamma, IFN-alpha, interleukin (IL)-2, IL-10, and tumor necrosis factor-alpha associated with fatal Ebola virus infection. J. Infect. Dis. 1999, 179, S188–S191. [Google Scholar]
- Hutchinson, K.L.; Rollin, P.E. Cytokine and chemokine expression in humans infected with Sudan Ebola virus. J. Infect. Dis. 2007, 196, S357–S363. [Google Scholar] [CrossRef]
- Food and Drug Administration. New Drug and Biological Drug Products; Evidence Needed to Demonstrate Effectiveness of New Drugs when Human Efficacy Studies are not Ethical or Feasible. Federal Register 2002, 67, 37988–37998.
- Bray, M. The role of the Type I interferon response in the resistance of mice to filovirus infection. J. Gen. Virol. 2001, 82, 1365–1373. [Google Scholar]
- Connolly, B.M.; Steele, K.E.; Davis, K.J.; Geisbert, T.W.; Kell, W.M.; Jaax, N.K.; Jahrling, P.B. Pathogenesis of experimental Ebola virus infection in guinea pigs. J. Infect. Dis. 1999, 179, S203–S217. [Google Scholar]
- Bray, M.; Davis, K.; Geisbert, T.; Schmaljohn, C.; Huggins, J. A mouse model for evaluation of prophylaxis and therapy of Ebola hemorrhagic fever. J. Infect. Dis. 1998, 178, 651–661. [Google Scholar]
- Cilloniz, C.; Ebihara, H.; Ni, C.; Neumann, G.; Korth, M.J.; Kelly, S.M.; Kawaoka, Y.; Feldmann, H.; Katze, M.G. Functional genomics reveals the induction of inflammatory response and metalloproteinase gene expression during lethal Ebola virus infection. J. Virol. 2011, 85, 9060–9068. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Pushko, P.; Anderson, K.; Smith, J.; Davis, K.J.; Jahrling, P.B. Evaluation in nonhuman primates of vaccines against Ebola virus. Emerg. Infect. Dis. 2002, 8, 503–507. [Google Scholar] [CrossRef]
- Reed, D.S.; Mohamadzadeh, M. Status and challenges of filovirus vaccines. Vaccine 2007, 25, 1923–1934. [Google Scholar] [CrossRef]
- Ebihara, H.; Rockx, B.; Marzi, A.; Feldmann, F.; Haddock, E.; Brining, D.; LaCasse, R.A.; Gardner, D.; Feldmann, H. Host response dynamics following lethal infection of rhesus macaques with Zaire ebolavirus. J. Infect. Dis. 2011, 204, S991–S999. [Google Scholar] [CrossRef]
- Ebihara, H.; Zivcec, M.; Gardner, D.; Falzarano, D.; Lacasse, R.; Rosenke, R.; Long, D.; Haddock, E.; Fischer, E.; Kawaoka, Y.; et al. A Syrian golden hamster model recapitulating Ebola hemorrhagic fever. J. Infect. Dis. 2012. [Google Scholar] [CrossRef]
- Michelow, I.C.; Lear, C.; Scully, C.; Prugar, L.I.; Longley, C.B.; Yantosca, L.M.; Ji, X.; Karpel, M.; Brudner, M.; Takahashi, K.; et al. High-dose mannose-binding lectin therapy for Ebola virus infection. J. Infect. Dis. 2011, 203, 175–179. [Google Scholar] [CrossRef]
- Bukreyev, A.; Marzi, A.; Feldmann, F.; Zhang, L.; Yang, L.; Ward, J.M.; Dorward, D.W.; Pickles, R.J.; Murphy, B.R.; Feldmann, H.; et al. Chimeric human parainfluenza virus bearing the Ebola virus glycoprotein as the sole surface protein is immunogenic and highly protective against Ebola virus challenge. Virology 2009, 383, 348–361. [Google Scholar] [CrossRef]
- Bukreyev, A.; Rollin, P.E.; Tate, M.K.; Yang, L.; Zaki, S.R.; Shieh, W.J.; Murphy, B.R.; Collins, P.L.; Sanchez, A. Successful topical respiratory tract immunization of primates against Ebola virus. J. Virol. 2007, 81, 6379–6388. [Google Scholar] [CrossRef]
- Bukreyev, A.; Yang, L.; Zaki, S.R.; Shieh, W.J.; Rollin, P.E.; Murphy, B.R.; Collins, P.L.; Sanchez, A. A single intranasal inoculation with a paramyxovirus-vectored vaccine protects guinea pigs against a lethal-dose Ebola virus challenge. J. Virol. 2006, 80, 2267–2279. [Google Scholar] [CrossRef]
- Marzi, A.; Ebihara, H.; Callison, J.; Groseth, A.; Williams, K.J.; Geisbert, T.W.; Feldmann, H. Vesicular stomatitis virus-based Ebola vaccines with improved cross-protective efficacy. J. Infect. Dis. 2011, 204, S1066–S10674. [Google Scholar] [CrossRef]
- Garbutt, M.; Liebscher, R.; Wahl-Jensen, V.; Jones, S.; Moller, P.; Wagner, R.; Volchkov, V.; Klenk, H.D.; Feldmann, H.; Stroher, U. Properties of replication-competent vesicular stomatitis virus vectors expressing glycoproteins of filoviruses and arenaviruses. J. Virol. 2004, 78, 5458–5465. [Google Scholar]
- Geisbert, T.W.; Daddario-Dicaprio, K.M.; Lewis, M.G.; Geisbert, J.B.; Grolla, A.; Leung, A.; Paragas, J.; Matthias, L.; Smith, M.A.; Jones, S.M.; et al. Vesicular stomatitis virus-based ebola vaccine is well-tolerated and protects immunocompromised nonhuman primates. PLoS Pathogens 2008, 4, e1000225. [Google Scholar] [CrossRef]
- Jones, S.M.; Stroher, U.; Fernando, L.; Qiu, X.; Alimonti, J.; Melito, P.; Bray, M.; Klenk, H.D.; Feldmann, H. Assessment of a vesicular stomatitis virus-based vaccine by use of the mouse model of Ebola virus hemorrhagic fever. J. Infect. Dis. 2007, 196, S404–S412. [Google Scholar] [CrossRef]
- Qiu, X.; Fernando, L.; Alimonti, J.B.; Melito, P.L.; Feldmann, F.; Dick, D.; Stroher, U.; Feldmann, H.; Jones, S.M. Mucosal immunization of cynomolgus macaques with the VSVDeltaG/ZEBOVGP vaccine stimulates strong ebola GP-specific immune responses. PloS One 2009, 4, e5547. [Google Scholar]
- Geisbert, T.W.; Daddario-Dicaprio, K.M.; Geisbert, J.B.; Reed, D.S.; Feldmann, F.; Grolla, A.; Stroher, U.; Fritz, E.A.; Hensley, L.E.; Jones, S.M.; et al. Vesicular stomatitis virus-based vaccines protect nonhuman primates against aerosol challenge with Ebola and Marburg viruses. Vaccine 2008, 26, 6894–6900. [Google Scholar]
- Jones, S.M.; Feldmann, H.; Stroher, U.; Geisbert, J.B.; Fernando, L.; Grolla, A.; Klenk, H.D.; Sullivan, N.J.; Volchkov, V.E.; Fritz, E.A.; et al. Live attenuated recombinant vaccine protects nonhuman primates against Ebola and Marburg viruses. Nat. Med. 2005, 11, 786–790. [Google Scholar] [CrossRef]
- Gilligan, K.; Geisbert, J.; Jahrling, P.; Anderson, K. Assessment of Protective Immunity Conferred by Recombinant Vaccinia Viruses to Guinea Pigs Challenged with Ebola Virus. In Vaccines 97: Molecular Approaches to the Control of Infectious Diseases; Brown, F., Burton, D., Doherty, P., Mekalanos, J., Norrby, E., Eds.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1997; pp. 87–92. [Google Scholar]
- Wilson, J.A.; Hart, M.K. Protection from Ebola virus mediated by cytotoxic T lymphocytes specific for the viral nucleoprotein. J. Virol. 2001, 75, 2660–2664. [Google Scholar] [CrossRef]
- Pushko, P.; Bray, M.; Ludwig, G.V.; Parker, M.; Schmaljohn, A.; Sanchez, A.; Jahrling, P.B.; Smith, J.F. Recombinant RNA replicons derived from attenuated Venezuelan equine encephalitis virus protect guinea pigs and mice from Ebola hemorrhagic fever virus. Vaccine 2000, 19, 142–153. [Google Scholar] [CrossRef]
- Olinger, G.G.; Bailey, M.A.; Dye, J.M.; Bakken, R.; Kuehne, A.; Kondig, J.; Wilson, J.; Hogan, R.J.; Hart, M.K. Protective cytotoxic T-cell responses induced by venezuelan equine encephalitis virus replicons expressing Ebola virus proteins. J. Virol. 2005, 79, 14189–14196. [Google Scholar]
- Phoolcharoen, W.; Dye, J.M.; Kilbourne, J.; Piensook, K.; Pratt, W.D.; Arntzen, C.J.; Chen, Q.; Mason, H.S.; Herbst-Kralovetz, M.M. A Nonreplicating Subunit Vaccine Protects Mice Against Lethal Ebola Virus Challenge. In Proceedings of the National Academy of Sciences of the United States of America; 2011; 108, pp. 20695–20700. [Google Scholar]
- Pushko, P.; Geisbert, J.; Parker, M.; Jahrling, P.; Smith, J. Individual and bivalent vaccines based on alphavirus replicons protect guinea pigs against infection with Lassa and Ebola viruses. J. Virol. 2001, 75, 11677–11685. [Google Scholar]
- Wilson, J.A.; Bray, M.; Bakken, R.; Hart, M.K. Vaccine potential of Ebola virus VP24, VP30, VP35, and VP40 proteins. Virology 2001, 286, 384–390. [Google Scholar]
- Gupta, M.; Mahanty, S.; Bray, M.; Ahmed, R.; Rollin, P.E. Passive transfer of antibodies protects immunocompetent and imunodeficient mice against lethal Ebola virus infection without complete inhibition of viral replication. J. Virol. 2001, 75, 4649–4654. [Google Scholar]
- Gupta, M.; Mahanty, S.; Greer, P.; Towner, J.S.; Shieh, W.J.; Zaki, S.R.; Ahmed, R.; Rollin, P.E. Persistent infection with ebola virus under conditions of partial immunity. J. Virol. 2004, 78, 958–967. [Google Scholar]
- Warfield, K.L.; Bosio, C.M.; Welcher, B.C.; Deal, E.M.; Mohamadzadeh, M.; Schmaljohn, A.; Aman, M.J.; Bavari, S. Ebola virus-like particles protect from lethal Ebola virus infection. In Proceedings of the National Academy of Sciences of the United States of America; 2003; 100, pp. 15889–15894. [Google Scholar]
- Rao, M.; Bray, M.; Alving, C.R.; Jahrling, P.; Matyas, G.R. Induction of immune responses in mice and monkeys to Ebola virus after immunization with liposome-encapsulated irradiated Ebola virus: protection in mice requires CD4(+) T cells. J. Virol. 2002, 76, 9176–9185. [Google Scholar] [CrossRef]
- Warfield, K.L.; Swenson, D.L.; Olinger, G.G.; Kalina, W.V.; Viard, M.; Aitichou, M.; Chi, X.; Ibrahim, S.; Blumenthal, R.; Raviv, Y.; et al. Ebola virus inactivation with preservation of antigenic and structural integrity by a photoinducible alkylating agent. J. Infect. Dis. 2007, 196, S276–S283. [Google Scholar] [CrossRef]
- Feldmann, H.; Jones, S.M.; Daddario-DiCaprio, K.M.; Geisbert, J.B.; Stroher, U.; Grolla, A.; Bray, M.; Fritz, E.A.; Fernando, L.; Feldmann, F.; et al. Effective post-exposure treatment of Ebola infection. PLoS Pathogens 2007, 3, e2. [Google Scholar] [CrossRef]
- Jahrling, P.B.; Geisbert, J.B.; Swearengen, J.R.; Larsen, T.; Geisbert, T.W. Ebola hemorrhagic fever: Evaluation of passive immunotherapy in nonhuman primates. J. Infect. Dis. 2007, 196, S400–S403. [Google Scholar] [CrossRef]
- Jahrling, P.B.; Geisbert, J.; Swearengen, J.R.; Jaax, G.P.; Lewis, T.; Huggins, J.W.; Schmidt, J.J.; LeDuc, J.W.; Peters, C.J. Passive immunization of Ebola virus-infected cynomolgus monkeys with immunoglobulin from hyperimmune horses. Arch. Virol. Suppl. 1996, 11, 135–140. [Google Scholar]
- Jahrling, P.B.; Geisbert, T.W.; Geisbert, J.B.; Swearengen, J.R.; Bray, M.; Jaax, N.K.; Huggins, J.W.; LeDuc, J.W.; Peters, C.J. Evaluation of immune globulin and recombinant interferon-alpha2b for treatment of experimental Ebola virus infections. J. Infect. Dis. 1999, 179, S224–S234. [Google Scholar]
- Parren, P.W.; Geisbert, T.W.; Maruyama, T.; Jahrling, P.B.; Burton, D.R. Pre- and postexposure prophylaxis of Ebola virus infection in an animal model by passive transfer of a neutralizing human antibody. J. Virol. 2002, 76, 6408–6412. [Google Scholar] [CrossRef]
- Oswald, W.B.; Geisbert, T.W.; Davis, K.J.; Geisbert, J.B.; Sullivan, N.J.; Jahrling, P.B.; Parren, P.W.; Burton, D.R. Neutralizing antibody fails to impact the course of Ebola virus infection in monkeys. PLoS Pathogens 2007, 3, e9. [Google Scholar] [CrossRef]
- Enterlein, S.; Warfield, K.L.; Swenson, D.L.; Stein, D.A.; Smith, J.L.; Gamble, C.S.; Kroeker, A.D.; Iversen, P.L.; Bavari, S.; Muhlberger, E. VP35 knockdown inhibits Ebola virus amplification and protects against lethal infection in mice. Antimicrob. Agents Chemother. 2006, 50, 984–993. [Google Scholar]
- Warfield, K.L.; Swenson, D.L.; Olinger, G.G.; Nichols, D.K.; Pratt, W.D.; Blouch, R.; Stein, D.A.; Aman, M.J.; Iversen, P.L.; Bavari, S. Gene-specific countermeasures against Ebola virus based on antisense phosphorodiamidate morpholino oligomers. PLoS Pathogens 2006, 2, e1. [Google Scholar] [CrossRef]
- Warren, T.K.; Warfield, K.L.; Wells, J.; Swenson, D.L.; Donner, K.S.; Van Tongeren, S.A.; Garza, N.L.; Dong, L.; Mourich, D.V.; Crumley, S.; et al. Advanced antisense therapies for postexposure protection against lethal filovirus infections. Nat. Med. 2010, 16, 991–994. [Google Scholar]
- Swenson, D.L.; Warfield, K.L.; Warren, T.K.; Lovejoy, C.; Hassinger, J.N.; Ruthel, G.; Blouch, R.E.; Moulton, H.M.; Weller, D.D.; Iversen, P.L.; Bavari, S. Chemical modifications of antisense morpholino oligomers enhance their efficacy against Ebola virus infection. Antimicrob. Agents Chemothe.r 2009, 53, 2089–2099. [Google Scholar] [CrossRef]
- Adler, S. Origin of the golden hamster Cricetus auratus as a laboratory animal. Nature 1948, 162, 256. [Google Scholar] [CrossRef]
- Boschert, K. Hamsters: Biology, care, diseases & models. Available online: http://netvet.wustl.edu/species/hamsters/hamstbio.txt (accessed on 26 June 2012).
- Murphy, M.R. History of the Capture and Domestication of the Syrian Golden Hamster (Mesocricetus auratus Waterhouse). In The Hamster: Reproduction and Behavior; Siegel, H.I., Ed.; Plenum Press: New York, NY, USA, 1985; pp. 3–20. [Google Scholar]
- Smith, G.D. Taxonomy and History. In The Laboratory Rabbit, guinea pig, hamster, and other rodents.; Suckow M.A.;, Stevens, K.A.; Wilson, R.P., Eds.; Acedemic Press: Waltham MA, 2012. [Google Scholar]
- Simonsen Laboratories. Origin and Strain Designation: Syrian Hamsters. Available online: http://www.simlab.com/products.html (accessed on 20 March 2012).
- Charles River Laboratories International. Charles River History. Available online: http://www.criver.com/en-us/aboutus/history/Pages/home.aspx (accessed on 20 March 2012).
- Charles River Laboratories International. LVG golden syrian hamsters 2012 US pricing. Available online: http://www.criver.com/sitecollectiondocuments/rm_rm_c_lvg_golden_syrian_hamsters.pdf (accessed on 20 March 2012).
- Harlan Laboratories. Syrian Hamster. 3248 Harlan Research Models and Services (RMS) Datasheets v4. Available online: http://www.harlan.com/products_and_services/research_models_and_services/research_models (accessed on 20 March 2012).
- Kunstyr, I.; Schoeneberg, U.; Friedhoff, K.T. Host specificity of Giardia muris isolates from mouse and golden hamster. Parasitol. Res. 1992, 78, 621–622. [Google Scholar] [CrossRef]
- Bio Breeders. Hamster models and applications for human diseases. Available online: http://www.biobreeders.com/hamstermodels.html (accessed on 26 June 2012).
- U.S. Department of Agriculture. Animals Used in Research: Animal Care Annual Report of Activities, Fiscal Year 2010. Available online: http://www.aphis.usda.gov/animal_welfare/efoia/downloads/2010_Animals_Used_In_Research.pdf (accessed 30 June 2012).
- Hankenson, F.C.; Van Hoosier, G.L. Biology and Diseases of Hamsters. In Laboratory Animal Medicine; Fox, J.G., Anderson, L.C., Loew, F.L., Quimby, F.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 167–202. [Google Scholar]
- Zlotnik, I. "Marburg disease". The Pathology of Experimentally Infected Hamsters. In Marburg Virus Disease; Martini, G.A., Siegert, R., Eds.; Springer-Verlag: Berlin, Germany, 1971; pp. 129–135. [Google Scholar]
- Hammerbeck, C.D.; Hooper, J.W. T cells are not required for pathogenesis in the Syrian hamster model of hantavirus pulmonary syndrome. J. Virol. 2011, 85, 9929–9944. [Google Scholar] [CrossRef]
- Wahl-Jensen, V.; Chapman, J.; Asher, L.; Fisher, R.; Zimmerman, M.; Larsen, T.; Hooper, J.W. Temporal analysis of Andes virus and Sin Nombre virus infections of Syrian hamsters. J. Virol. 2007, 81, 7449–7462. [Google Scholar]
- Safronetz, D.; Zivcec, M.; Lacasse, R.; Feldmann, F.; Rosenke, R.; Long, D.; Haddock, E.; Brining, D.; Gardner, D.; Feldmann, H.; et al. Pathogenesis and host response in Syrian hamsters following intranasal infection with Andes virus. PLoS Pathogens 2011, 7, e1002426. [Google Scholar]
- Campen, M.J.; Milazzo, M.L.; Fulhorst, C.F.; Obot Akata, C.J.; Koster, F. Characterization of shock in a hamster model of hantavirus infection. Virology 2006, 356, 45–49. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Eyzaguirre, E.J.; Molina, C.P.; Fulhorst, C.F. Maporal viral infection in the Syrian golden hamster: A model of hantavirus pulmonary syndrome. J. Infect. Dis. 2002, 186, 1390–1395. [Google Scholar] [CrossRef]
- Hooper, J.W.; Larsen, T.; Custer, D.M.; Schmaljohn, C.S. A lethal disease model for hantavirus pulmonary syndrome. Virology 2001, 289, 6–14. [Google Scholar] [CrossRef]
- Gowen, B.B.; Julander, J.G.; London, N.R.; Wong, M.H.; Larson, D.; Morrey, J.D.; Li, D.Y.; Bray, M. Assessing changes in vascular permeability in a hamster model of viral hemorrhagic fever. Virol. J. 2010, 7, 240. [Google Scholar] [CrossRef]
- Sbrana, E.; Xiao, S.Y.; Popov, V.L.; Newman, P.C.; Tesh, R.B. Experimental yellow fever virus infection in the golden hamster (Mesocricetus auratus) III. Clinical laboratory values. Am. J. Trop. Med. Hyg. 2006, 74, 1084–1089. [Google Scholar]
- Xiao, S.Y.; Zhang, H.; Guzman, H.; Tesh, R.B. Experimental yellow fever virus infection in the Golden hamster (Mesocricetus auratus). II. Pathology. J. Infect. Dis. 2001, 183, 1437–1444. [Google Scholar] [CrossRef]
- Fisher, A.F.; Tesh, R.B.; Tonry, J.; Guzman, H.; Liu, D.; Xiao, S.Y. Induction of severe disease in hamsters by two sandfly fever group viruses, Punta toro and Gabek Forest (Phlebovirus, Bunyaviridae), similar to that caused by Rift Valley fever virus. Am. J. Trop. Med. Hyg. 2003, 69, 269–276. [Google Scholar]
- Vela, E.M.; Knostman, K.A.; Mott, J.M.; Warren, R.L.; Garver, J.N.; Vela, L.J.; Stammen, R.L. Genistein, a general kinase inhibitor, as a potential antiviral for arenaviral hemorrhagic fever as described in the Pirital virus-Syrian golden hamster model. Antivir. Res. 2010, 87, 318–328. [Google Scholar] [CrossRef]
- Li, G.; Duan, T.; Wu, X.; Tesh, R.B.; Soong, L.; Xiao, S.Y. Yellow fever virus infection in Syrian golden hamsters: Relationship between cytokine expression and pathologic changes. Int. J. Clin. Exp. Pathol. 2008, 1, 169–179. [Google Scholar]
- Sbrana, E.; Mateo, R.I.; Xiao, S.Y.; Popov, V.L.; Newman, P.C.; Tesh, R.B. Clinical laboratory, virologic, and pathologic changes in hamsters experimentally infected with Pirital virus (Arenaviridae): A rodent model of Lassa fever. Am. J. Trop. Med. Hyg. 2006, 74, 1096–1102. [Google Scholar]
- Xiao, S.Y.; Zhang, H.; Yang, Y.; Tesh, R.B. Pirital virus (Arenaviridae) infection in the syrian golden hamster, Mesocricetus auratus: a new animal model for arenaviral hemorrhagic fever. Am. J. Trop. Med. Hyg. 2001, 64, 111–118. [Google Scholar]
- Lima, E.Q.; Nogueira, M.L. Viral hemorrhagic fever-induced acute kidney injury. Semin. Nephrol. 2008, 28, 409–415. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Young, H.A.; Jahrling, P.B.; Davis, K.J.; Larsen, T.; Kagan, E.; Hensley, L.E. Pathogenesis of Ebola hemorrhagic fever in primate models: Evidence that hemorrhage is not a direct effect of virus-induced cytolysis of endothelial cells. Am. J. Pathol. 2003, 163, 2371–2382. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Young, H.A.; Jahrling, P.B.; Davis, K.J.; Kagan, E.; Hensley, L.E. Mechanisms underlying coagulation abnormalities in ebola hemorrhagic fever: Overexpression of tissue factor in primate monocytes/macrophages is a key event. J. Infect. Dis. 2003, 188, 1618–1629. [Google Scholar] [CrossRef]
- Ryabchikova, E.I.; Kolesnikova, L.V.; Netesov, S.V. Animal pathology of filoviral infections. Curr. Top Microbiol. Immunol. 1999, 235, 145–173. [Google Scholar]
- Feldmann, H.; Geisbert, T.W. Ebola haemorrhagic fever. Lancet 2011, 377, 849–862. [Google Scholar] [CrossRef]
- Aleksandrowicz, P.; Wolf, K.; Falzarano, D.; Feldmann, H.; Seebach, J.; Schnittler, H. Viral haemorrhagic fever and vascular alterations. Hamostaseologie 2008, 28, 77–84. [Google Scholar]
- Zivcec, M.; Safronetz, D.; Haddock, E.; Feldmann, H.; Ebihara, H. Validation of assays to monitor immune responses in the Syrian golden hamster (Mesocricetus auratus). J. Immunol. Meth. 2011, 368, 24–35. [Google Scholar] [CrossRef]
- McNees, A.L.; Vilchez, R.A.; Heard, T.C.; Sroller, V.; Wong, C.; Herron, A.J.; Hamilton, M.J.; Davis, W.C.; Butel, J.S. SV40 lymphomagenesis in Syrian golden hamsters. Virology 2009, 384, 114–124. [Google Scholar] [CrossRef]
- Dondji, B.; Sun, T.; Bungiro, R.D.; Vermeire, J.J.; Harrison, L.M.; Bifulco, C.; Cappello, M. CD4 T cells mediate mucosal and systemic immune responses to experimental hookworm infection. Parasite Immunol. 2010, 32, 406–413. [Google Scholar] [CrossRef]
- Dondji, B.; Bungiro, R.D.; Harrison, L.M.; Vermeire, J.J.; Bifulco, C.; McMahon-Pratt, D.; Cappello, M. Role for nitric oxide in hookworm-associated immune suppression. Infect. Immun. 2008, 76, 2560–2567. [Google Scholar] [CrossRef]
- Yang, K.; Zhang, G.; Mei, J.; Chen, D.; Wu, M. Screening and analysis of pathogenic genes during DMBA-induced buccal mucosa carcinogenesis in golden hamsters. Oncol. Rep. 2010, 23, 1619–1624. [Google Scholar]
- Yu, T.; Wang, X.Y.; Gong, R.G.; Li, A.; Yang, S.; Cao, Y.T.; Wen, Y.M.; Wang, C.M.; Yi, X.Z. The expression profile of microRNAs in a model of 7,12-dimethyl-benz[a]anthrance-induced oral carcinogenesis in Syrian hamster. J. Exp. Clin. Canc. Res. 2009, 28, 64. [Google Scholar] [CrossRef]
- Hohlweg, U.; Hosel, M.; Dorn, A.; Webb, D.; Hilger-Eversheim, K.; Remus, R.; Schmitz, B.; Buettner, R.; Schramme, A.; Corzilius, L.; et al. Intraperitoneal dissemination of Ad12-induced undifferentiated neuroectodermal hamster tumors: De novo methylation and transcription patterns of integrated viral and of cellular genes. Virus Res. 2003, 98, 45–56. [Google Scholar] [CrossRef]
- Maywood, E.S.; Chahad-Ehlers, S.; Garabette, M.L.; Pritchard, C.; Underhill, P.; Greenfield, A.; Ebling, F.J.; Akhtar, R.A.; Kyriacou, C.P.; Hastings, M.H.; et al. Differential testicular gene expression in seasonal fertility. J. Biol. Rhythm. 2009, 24, 114–125. [Google Scholar] [CrossRef]
- Bradley, K.C.; Boulware, M.B.; Jiang, H.; Doerge, R.W.; Meisel, R.L.; Mermelstein, P.G. Changes in gene expression within the nucleus accumbens and striatum following sexual experience. Gene Brain Behav. 2005, 4, 31–44. [Google Scholar]
- Matsui, M.; Rouleau, V.; Bruyere-Ostells, L.; Goarant, C. Gene expression profiles of immune mediators and histopathological findings in animal models of leptospirosis: comparison between susceptible hamsters and resistant mice. Infect Immun. 2011, 79, 4480–4492. [Google Scholar] [CrossRef]
- Gowen, B.B.; Fairman, J.; Dow, S.; Troyer, R.; Wong, M.H.; Jung, K.H.; Melby, P.C.; Morrey, J.D. Prophylaxis with cationic liposome-DNA complexes protects hamsters from phleboviral disease: importance of liposomal delivery and CpG motifs. Antivir. Res. 2009, 81, 37–46. [Google Scholar]
- Gowen, B.B.; Judge, J.W.; Wong, M.H.; Jung, K.H.; Aylsworth, C.F.; Melby, P.C.; Rosenberg, B.; Morrey, J.D. Immunoprophylaxis of Punta Toro virus (Phlebovirus, Bunyaviridae) infection in hamsters with recombinant Eimeria profilin-like antigen. Int. Immunopharm. 2008, 8, 1089–1094. [Google Scholar] [CrossRef]
- Mendez, S.; Valenzuela, J.G.; Wu, W.; Hotez, P.J. Host cytokine production, lymphoproliferation, and antibody responses during the course of Ancylostoma ceylanicum infection in the Golden Syrian hamster. Infect. Immun. 2005, 73, 3402–3407. [Google Scholar] [CrossRef]
- Rockx, B.; Brining, D.; Kramer, J.; Callison, J.; Ebihara, H.; Mansfield, K.; Feldmann, H. Clinical outcome of henipavirus infection in hamsters is determined by the route and dose of infection. J. Virol. 2011, 85, 7658–7671. [Google Scholar] [CrossRef]
- Rama Iniguez, S.; Dea-Ayuela, M.A.; Sanchez-Brunete, J.A.; Torrado, J.J.; Alunda, J.M.; Bolas-Fernandez, F. Real-time reverse transcription-PCR quantification of cytokine mRNA expression in golden Syrian hamster infected with Leishmania infantum and treated with a new amphotericin B formulation. Antimicrob. Agents Chemother. 2006, 50, 1195–1201. [Google Scholar] [CrossRef]
- Brown, K.S.; Safronetz, D.; Marzi, A.; Ebihara, H.; Feldmann, H. Vesicular stomatitis virus-based vaccine protects hamsters against lethal challenge with Andes virus. J. Virol. 2011, 85, 12781–12791. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wahl-Jensen, V.; Bollinger, L.; Safronetz, D.; De Kok-Mercado, F.; Scott, D.P.; Ebihara, H. Use of the Syrian Hamster as a New Model of Ebola Virus Disease and Other Viral Hemorrhagic Fevers. Viruses 2012, 4, 3754-3784. https://doi.org/10.3390/v4123754
Wahl-Jensen V, Bollinger L, Safronetz D, De Kok-Mercado F, Scott DP, Ebihara H. Use of the Syrian Hamster as a New Model of Ebola Virus Disease and Other Viral Hemorrhagic Fevers. Viruses. 2012; 4(12):3754-3784. https://doi.org/10.3390/v4123754
Chicago/Turabian StyleWahl-Jensen, Victoria, Laura Bollinger, David Safronetz, Fabian De Kok-Mercado, Dana P. Scott, and Hideki Ebihara. 2012. "Use of the Syrian Hamster as a New Model of Ebola Virus Disease and Other Viral Hemorrhagic Fevers" Viruses 4, no. 12: 3754-3784. https://doi.org/10.3390/v4123754