Filovirus Entry: A Novelty in the Viral Fusion World

Department of Microbiology, University of Iowa, Iowa City, IA 52242, USA
*
Author to whom correspondence should be addressed.
Viruses 2012, 4(2), 258-275; https://doi.org/10.3390/v4020258
Submission received: 23 December 2011
/
Revised: 24 January 2012
/
Accepted: 30 January 2012
/
Published: 7 February 2012
(This article belongs to the Special Issue Virus-Induced Membrane Fusion)
{kind=link}
{kind=link}
Abstract
:Ebolavirus (EBOV) and Marburgvirus (MARV) that compose the filovirus family of negative strand RNA viruses infect a broad range of mammalian cells. Recent studies indicate that cellular entry of this family of viruses requires a series of cellular protein interactions and molecular mechanisms, some of which are unique to filoviruses and others are commonly used by all viral glycoproteins. Details of this entry pathway are highlighted here. Virus entry into cells is initiated by the interaction of the viral glycoprotein1 subunit (GP1) with both adherence factors and one or more receptors on the surface of host cells. On epithelial cells, we recently demonstrated that TIM-1 serves as a receptor for this family of viruses, but the cell surface receptors in other cell types remain unidentified. Upon receptor binding, the virus is internalized into endosomes primarily via macropinocytosis, but perhaps by other mechanisms as well. Within the acidified endosome, the heavily glycosylated GP1 is cleaved to a smaller form by the low pH-dependent cellular proteases Cathepsin L and B, exposing residues in the receptor binding site (RBS). Details of the molecular events following cathepsin-dependent trimming of GP1 are currently incomplete; however, the processed GP1 specifically interacts with endosomal/lysosomal membranes that contain the Niemann Pick C1 (NPC1) protein and expression of NPC1 is required for productive infection, suggesting that GP/NPC1 interactions may be an important late step in the entry process. Additional events such as further GP1 processing and/or reducing events may also be required to generate a fusion-ready form of the glycoprotein. Once this has been achieved, sequences in the filovirus GP2 subunit mediate viral/cellular membrane fusion via mechanisms similar to those previously described for other enveloped viruses. This multi-step entry pathway highlights the complex and highly orchestrated path of internalization and fusion that appears unique for filoviruses.
1. Introduction
Filoviruses (family Mononegavirales, genera Ebolavirus (EBOV) and Marburgvirus (MARV)) are single-stranded, negative-sense RNA viruses that exhibit a unique heterogeneous filamentous structure. Both EBOV and MARV infect a wide variety of mammals and this wide tropism has complicated the identification of cellular proteins required for viral entry. A hemorrhagic fever is caused by these viruses in humans, non-human primates and perhaps other mammals and is associated with high morbidity and mortality during outbreaks. No therapeutic drugs or vaccines are currently available to treat or prevent filoviral infection. Because of this and the high lethality associated with infection, filoviruses are considered Category A Priority Pathogens by NIAID and, in recent years, much research has focused on understanding how these viruses bind to and enter permissive cells.
3. Structure of Filovirus GPs
As a class I viral fusion protein, GP1,2 exists as a trimer of GP1 and GP2 heterodimeric subunits and is found in its full-length, heavily glycosylated, pre-fusion form on the surface of newly budded virions. This pre-fusion class I glycoprotein conformation is thought to be a metastable state, that upon the appropriate set of triggers, will convert to its post-fusion, low-energy form [6]. The conformational change from a pre-fusion to post-fusion structure provides the energy to permit the viral and cellular membranes to fuse and thereby release the viral core into the cytoplasm. The events that are required for filovirus/cellular membrane fusion to occur have yet to be completely elucidated, but current studies are unraveling these steps and are highlighted here.
Several EBOV GP crystal structures have been instrumental in understanding the conformational alterations that are required as the glycoprotein changes from the metastable, pre-fusion state to the low-energy, post-fusion form. Lee et al. elucidated the trimeric structure of pre-fusion, mucin domain-deleted EBOV GP1,2 ectodomain [7], whereas, two groups independently solved the trimeric GP2 six helix bundle structure that is formed during fusion events [8,9].
4. GP1
Mature GP1 is composed of three distinct domains: the RBD, the glycan cap and a heavily O-linked glycosylated mucin-like domain. The RBD in mature ZEBOV GP1 is located from amino acid ~54 to 201 and composed of a base region and a region that interacts with one or more receptors on the surface of cells (Figure 1) [10]. While RBDs of the different EBOV strains are relatively conserved, only 47% identity between EBOV and MARV RBDs exists. Nonetheless, EBOV and MARV GP pseudovirions compete with each other for filoviral GP1,2-dependent entry into permissive cells, indicating that a common receptor or receptors are used by both viruses [10,11].
Amino acids 33 through 69 and three additional short downstream regions interact with GP2, serving as the base of the RBD. Several linearly discontinuous regions from amino acids 70 to 190 sit above the base forming a series of beta sheets. Residues both in beta sheets and adjacent loops have been implicated in cell binding, leading to the conclusion that the receptor binding site (RBS) is located in this region of the RBD [12,13]. GP1 residues 227 to 313 encode for a “glycan cap” that is extensively N-linked glycosylated and sits distal to the RBD from the surface of the virion. The glycan cap may protect the RBS from antibodies [3]. This glycan cap also interacts with two regions of GP2, including the internal fusion loop of GP2 that is critical for GP2-mediated membrane fusion [7]. The glycan cap/GP2 interaction restricts the availability of the fusion peptide, preventing pre-mature fusion events. Finally, filovirus GP1 contains a mucin-like domain at the carboxy terminus from amino acids 310 to 511. This region is heavily glycosylated with both N- and O- linked glycans [7]. While the EBOV mucin domain is not required for virus entry [14,15], several roles for this domain have been suggested. Like the glycan cap, it may shield GP RBS residues from immune recognition on free virus [7]. In addition, the mucin domain causes cell rounding, masking of a number of cell surface markers and cytotoxicity that is not observed upon expression of mucin domain-deleted EBOV GP [15,16]. The shielding effect of the bulky mucin domain of the RBD of GP1 is thought to also obstruct RBS interactions with adherence factors and receptors since removal of this domain enhances EBOV titers. Similar attempts to delete the MARV mucin domain have proved unsuccessful [11].
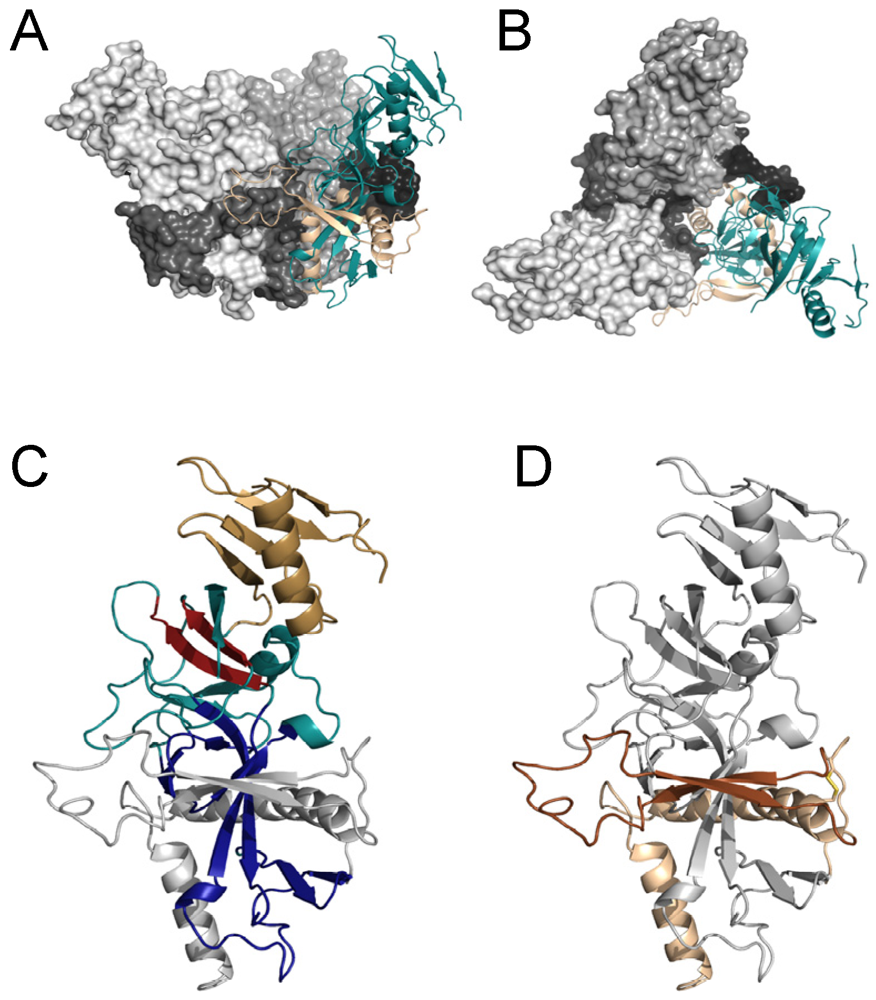
Figure 1.
Structure of the pre-fusion EBOV GP. (A,B) Structure of the trimer. Side view of the EBOV GP trimer is shown in A. Top-down view of the EBOV GP trimer is in B. Two of the three trimers are shown as space filling structures with GP1 in lighter grey and GP2 as dark grey/black. The third GP1,2 heterodimer of the trimer is depicted as a ribbon structure with GP1 shown in teal and the GP2 subunit shown in tan. (C,D) Ribbon diagrams of a single heterodimer of GP1,2. Domains in GP1 are highlighted in C, whereas domains in GP2 are highlighted in D. In panel C, the base domain of GP1 that interacts with GP2 is shown in royal blue, the head domain is shown in teal with the beta-strands and adjacent loop region containing the RBS highlighted in red and the glycan cap is shown in gold. GP2 is shown in grey. In panel D, the internal fusion loop (IFL) that is flanked by beta-strands is shown in dark brown and heptad repeat region 1 is shown in tan. The interaction of the IFL with GP1 residues from an adjacent subunit is evident in panel A. All EBOV GP graphics (PDB accession number 3CSY) were produced with PyMol.
Figure 1.
Structure of the pre-fusion EBOV GP. (A,B) Structure of the trimer. Side view of the EBOV GP trimer is shown in A. Top-down view of the EBOV GP trimer is in B. Two of the three trimers are shown as space filling structures with GP1 in lighter grey and GP2 as dark grey/black. The third GP1,2 heterodimer of the trimer is depicted as a ribbon structure with GP1 shown in teal and the GP2 subunit shown in tan. (C,D) Ribbon diagrams of a single heterodimer of GP1,2. Domains in GP1 are highlighted in C, whereas domains in GP2 are highlighted in D. In panel C, the base domain of GP1 that interacts with GP2 is shown in royal blue, the head domain is shown in teal with the beta-strands and adjacent loop region containing the RBS highlighted in red and the glycan cap is shown in gold. GP2 is shown in grey. In panel D, the internal fusion loop (IFL) that is flanked by beta-strands is shown in dark brown and heptad repeat region 1 is shown in tan. The interaction of the IFL with GP1 residues from an adjacent subunit is evident in panel A. All EBOV GP graphics (PDB accession number 3CSY) were produced with PyMol.

5. GP2
Filovirus GP2 is similar to other class I viral glycoproteins that mediate fusion events. GP2 is composed of a fusion loop found near the amino terminus followed by a N-terminal heptad repeat region, a C-terminal heptad repeat region, a transmembrane region and a short cytoplasmic tail [6,8,9]. In the pre-fusion form on the surface of newly budded virions, the fusion loop and N-terminal heptad region are integral regions of the GP1/GP2 trimeric structure, forming a platform or base upon which GP1 sits (Figure 1). In contrast, the C-terminal heptad repeat region in the pre-fusion form may contain little structure and was deleted in the crystallization studies and it is not thought to contribute to the metastable, pre-fusion form of GP [7]. Not surprisingly, both the fusion loop and N-terminal heptad repeat are conserved (>70% identity) between ZEBOV and the Musoke strain of MARV, but the remaining portions of GP2 have limited identity. Site directed mutagenesis studies of EBOV or MARV GP2 [14,17,18,19,20,21,22,23] suggest a similar chain of events to those previously reported for other class I viral glycoproteins leads to viral/cell membrane fusion [6,24,25]. These events will be discussed in detail below.
6. The Role of Cell Surface Proteins in Filovirus GP-Dependent Entry
While progress is being made to identify cell surface proteins that enhance filovirus transduction/infection, the advancement of this area of research has been slow. In part this is due to the broad tropism of filoviruses for a variety of different cell types as well as the ability of these viruses to infect cells from a wide range of species [26,27,28,29,30,31,32,33,34,35,36,37]. One classical approach to identifying virus receptors that has been used is the introduction of a cDNA library from a permissive cell into a cell that is not permissive for the virus [38,39,40]. However, for reasons that are not entirely clear, this type of study has not been successful in identifying cell surface proteins that directly interact with EBOV GP to mediate virus entry [41,42]. Instead, another screening approach that correlated gene expression in a large panel of human cells with EBOV GP-dependent transduction proved more productive and allowed us to identify a surface receptor for these viruses [43]. The cell surface protein we identified, TIM-1, as well as other cell surface proteins that enhance filovirus infectivity are discussed in detail below.
6.1. C-Type Lectins
C-type lectin family members L-SIGN, DC-SIGN and hMGL have been shown to enhance filovirus entry [19,44,45,46,47,48]. Studies have demonstrated that both the mucin domain and the glycan cap of GP1 interact with C-type lectins [47,49]. However, as both of these regions can be deleted from EBOV GP1 without loss of viral transduction efficiency [16,50,51,52], it is likely that C-type lectins increase filovirus attachment to cells rather than serving as cellular receptors that mediate internalization of the virus into endosomes [53]. A similar adherence function for C-type lectins has been identified for other enveloped viruses such as HIV [54].
6.2. β1 Integrins
Early studies found that surface expression of some integrins was down regulated upon transfection of full-length EBOV GP [15,55,56]. In addition, EBOV GP-mediated pseudovirion entry is reduced by either antibodies targeting β1 integrins or a soluble form of β1 integrins [56]. These characteristics suggested that one or more integrin subunits might serve as receptors for filoviruses. However, no direct interaction between any portion of EBOV GP and a member of the β1 integrin family has been identified. More recent studies implicate β1 integrin in stimulation of endosomal protease events that are required for productive EBOV transduction, thus reducing the likelihood of β1 integrins serving as bona fide filoviral receptors [57].
6.3. Tyro3 (TAM) Family Tyrosine Kinase Receptors
The TAM family member Axl was first implicated in filovirus entry through a cDNA screen that introduced Vero E6 cell cDNA into poorly permissive Jurkat T cells [41]. Axl is a tyrosine kinase receptor that is found on the plasma membrane in a variety of different cell types and enhances cell migration, division and viability upon activation [58]. Shimojima et al. demonstrated that anti-Axl antibodies blocked EBOV transduction of some cells, whereas these antibodies had no effect on transduction of other cells [41]. Mapping studies indicated that amino acid residues in both the ectodomain and the cytoplasmic tail of Axl were required for filovirus entry enhancement [59]. Subsequently, a screen performed in our laboratory also identified Axl as being important in EBOV GP-dependent entry [60]. Through the use of multiple biochemical inhibitors, siRNA and anti-Axl antibodies, we defined a role for Axl in EBOV uptake [61], demonstrating that Axl expression enhances macropinocytosis in some cells. As macropinocytosis is a principal uptake mechanism of filoviruses [62,63], increased Axl surface expression leads to greater virus internalization. However, Axl does not appear to interact directly with EBOV GP to promote viral internalization and therefore is unlikely to serve as a filoviral receptor [60].
6.4. TIM-1
Our lab performed a comparative genomic analysis (CGA) screen [64] to identify cellular genes whose expression highly correlated with EBOV pseudovirion transduction [43]. This screen showed a positive correlation between EBOV transduction and expression of a series of cellular proteins that were previously appreciated to enhance EBOV transduction (C-type lectins, integrins and Axl). Interestingly, expression of the T-cell immunoglobulin mucin domain-1 (TIM-1) gene proved to be one of the strongest positive correlations. Subsequent studies demonstrated that expression of TIM-1 in poorly permissive cells enhanced EBOV entry and loss of surface-expressed TIM-1 in highly permissive cells abrogated filovirus infection/transduction. Furthermore, TIM-1 and the mucin domain-deleted EBOV GP interacted, and removal of the glycan cap enhanced the specificity of GP interaction with TIM-1-expressing cells [43]. In total, these findings have led us to propose TIM-1 as a cell surface receptor for filoviruses. As epithelial cells are the only relevant cell type that expresses TIM-1, it is likely that other as of yet unidentified surface receptors will also prove to be important in mediating filovirus entry.
8. Proteolytic Processing of EBOV GP1,2 into a Fusion-Active Form
Unlike some other class I viral glycoproteins, no evidence exists that EBOV GP1 undergoes conformational changes upon binding to a cell surface receptor. Presumably receptor/virus interactions lead directly to virus internalization into endosomes; however, this has not been demonstrated directly to date. Characterization of the endosomal vesicles mediating EBOV uptake is limited. Saaed et al. have demonstrated that the Early Endosome Antigen 1 (EEA1) colocalizes with EBOV virus particles in endosomes early during infection of Vero cells [62], whereas Nanbo et al. have shown association of EBOV particles lacking VP30 associated with the sorting nexin 5 (SNX5) within 10 minutes of transduction [63]. Early and late endosomal proteins, Rab5 and Rab 7, respectively, have also been shown to be required for productive infection [62,63]. Several GTPases previously implicated in endocytosis, RhoB, Rac1 and CDC42, are also important for EBOV GP-dependent transduction, providing additional insights into trafficking pathways used by these viruses [63,76].
As the vesicles traffic into the cell, they become acidified. For some viral fusion proteins, the combination of receptor engagement and endosomal acidity is sufficient for conformational changes that lead to viral/cellular membrane fusion [77,78,79]. However, that is not the case for filoviruses. Instead, multiple low pH-dependent endosomal and lysosomal proteolytic proteins are involved in EBOV GP1 processing, priming a multi-step process that ultimately results in a small EBOV GP1,2 trimer that serves as the fusion-ready form of the glycoprotein. It is believed that this processed, fusion-ready form is then triggered by additional events to a conformationally stable state, resulting in fusion.
As endosomal vesicles mature into late endosomes and the vesicular pH drops, activation of endosomal cysteine proteases Cathepsin L and B occurs. These cathepsins sequentially process EBOV GP1 into smaller forms [50,51]. Cathepsin L proteolysis first removes a substantial portion of EBOV GP1, generating an approximate 50 kDa GP1,2 species that lacks both the glycan cap and mucin domain of GP1 [50,51]. Subsequently, GP1 is further trimmed by both Cathepsin L and B to generate a much smaller form of GP1. The exact size of this smaller GP1 remains controversial, but is between 17 and 19 kDa [50,51]. Irrespective of the exact size of the processed form, prevention of endosomal acidification or inhibition of Cathepsin B activity abolishes EBOV infectivity [50,51,52,70]. Interestingly, this processing may be insufficient for productive EBOV infection as studies have demonstrated that the smaller 17–19 kDa Cathepsin L and B-cleaved form cannot infect cells entirely lacking cathepsins [80]. Thus, it has been proposed that additional cathepsin-dependent GP1 processing is required to generate the fusion-ready form of the glycoprotein [80]. The size and composition of this smaller form is not known. In contrast to these studies, a recent study demonstrated that a thermolysin-trimmed GP1 that is believed to generate a GP1 that is similar to the Cathepsin L and B-cleaved form can be triggered to bind to liposomes at elevated temperatures under low pH and mildly reducing conditions [81]. This new study suggests that at least under these conditions this GP conformation is a fusion-ready form.
Given the apparent importance of cathepsin cleavage for the generation of a fusion-ready form of the filovirus glycoprotein, it is surprising that studies have demonstrated that Cathepsin L and B cleavage events can be sidestepped by the virus [80,82]. Martinez et al. have shown that in monocyte-derived dendritic cells, Cathepsin B is required for EBOV infection, but not Cathepsin L [82]. These observations were confirmed in Vero cells in a recent study by Wong et al. [80]. This group also extended the finding by selecting an infectious, recombinant VSV encoding EBOV GP over several passages to become resistant to the Cathepsin B inhibitor CA074 [80]. Six different mutants were identified that conferred Cathepsin B independence. Many of these mutations sit at the interface of GP1 and GP2 and the selected mutations were thought to alter the GP structure such that enhanced proteolysis by one or more currently undefined cysteine proteases was possible. Thus, while Cathepsin B and L independence can be achieved by filoviruses, processing by one or more additional cysteine proteases is still required for production of the fusion-ready form.
9. NPC1
Most recently, two groups independently identified a novel host protein essential for EBOV infection. Cote et al. screened a library of small chemical molecules to find those that inhibited EBOV GP pseudovirion entry into Vero cells [83], whereas Carette et al. utilized a genome-wide screen in mutagenized haploid human cells to look for those cells resistant to EBOV GP-dependent entry [84]. Each group was able to deduce that disruption of one endosomal/lysosomal membrane protein, Niemann-Pick C1 (NPC1), could significantly reduce EBOV entry into a variety of cell populations.
NPC1 is primarily a membrane bound, late endosomal/lysosomal protein that is critical for cholesterol absorption and homeostasis. Those individuals lacking a functional NPC1 exhibit an abnormally high accumulation of cholesterol in the lysosomes of their cells, leading to altered protein and lipid trafficking with most cases resulting in fatality by 18 months of age [85]. Cells where NPC1 function has been biochemically disrupted or cells lacking NPC1 showed a resistance to EBOV infection [83,84]. Clearance of cholesterol from NPC1 null cells by cultivation in lipoprotein-depleted media did not rescue EBOV infection, indicating that the NPC1 protein itself, and not aberrant cholesterol transport, was important for EBOV entry [84]. Cote et al. also showed that the expression of NPC1 and not its cholesterol transport activity were critical for EBOV entry [83].
As the NPC1 protein is primarily located on the endosomal and lysosomal membranes, NPC1 has been proposed to serve as an entry factor downstream of EBOV GP engagement of attachment factors/receptor(s) at the cell surface. Consistent with a vesicular role for NPC1, Cote et al. showed that a soluble form of thermolysin-cleaved EBOV GP, but not uncleaved GP containing the glycan cap, bound to lysosomal membranes of NPC1-transfected CHO cells [83]. Thus, at least in this over expression system, proteolytically-processed EBOV GP appeared to interact with NPC1-containing membranes, suggesting that these interactions may be important for filovirus entry events that occur in late endosomes and/or in lysosomes. EBOV GP-mediated attachment and entry into early endosomes was unaffected in NPC1-defective cells; however, electron micrographs of NPC1 null cells infected with EBOV GP pseudotyped virus show the accumulation of perinuclear vesicles laden with EBOV GP pseudovirions that were positive for the lysosomal marker LAMP1 [84]. Therefore, Carette et al. have proposed that NCP1 is crucial for viral membrane fusion and escape from the lysosomal vesicle [84]. At present, the precise role of NPC1 during the EBOV entry process remains to be fully elucidated. In addition, investigations into the precise location of filovirus fusion events within endosomal compartments will provide important insights into these events.
Interestingly, an inhibitor of NPC1, U18666A, has recently been shown to block entry of several pathogens, including the flavivirus Dengue [86]. This inhibitor also inhibited entry of hepatitis C virus [87] as well as prions [88], suggesting that this cholesterol transporter may be critical for passage of a number of viruses, and perhaps other pathogens, through endosomes. Most recently, a more definitive role for NPC1 in hepatitis C virus entry has been determined through the use of additional biochemical agents [89]. Future studies exploring lipid accumulation and changes in lipid composition within the endosomal pathway could significantly enhance the understanding of the novel role of NPC1 specifically in filoviral entry and more generally in endosomal trafficking of a number of enveloped viruses.
10. Characterized Filovirus Fusion Events
As described above, the GP1 portion of EBOV GP1,2 allows delivery of the filovirion to an endosome where conditions become progressively more favorable for generating the fusion-ready form of GP1,2. Trimming of GP1,2 by host cathepsins (or artificially by thermolysin) enhances interaction of GP with TIM-1 [43] and permits NPC1 binding [83]. Whether EBOV GP interaction, with either of these molecules, directly mediates GP2 fusion events remains to be determined.
Filovirus GP2 contains an N-terminal internal fusion loop of 45 residues defined by a disulfide linkage between cysteines 511 and 556 [3,7]. A core hydrophobic sequence of 16 amino acids within this loop is thought to insert into host endosomal membranes, initiating membrane fusion events. Within the intact GP heterodimer, the hydrophobic, internal fusion loop is flanked by antiparallel β-strands composed of the most N-terminal portion of the internal fusion loop and the N-terminal region of first heptad repeat region. The fusion loop is restrained by GP1 residues from a neighboring subunit, preventing premature fusion events [3,7,90]. Cathepsin-dependent processing alone is insufficient to trigger insertion of the fusion loop into liposomes [81,91].
When the EBOV internal fusion loop interacts with liposomal membrane mimetics, lipid mixing is promoted with a parallel structural change in the loop [90]. In a neutral pH, lipid environment, the antiparallel β-strands that flank the fusion loop lose their structure, generating a more alpha helical content with a flattened extended loop structure. Under low pH conditions (<pH 5.5) in the presence of lipids, this flattened loop structure broadens out, forming a more hook-like structure [90]. It has been previously shown that specific proline residues contained within the central portion of the fusion loop facilitate this lipid membrane interaction [21,23,92,93]. The insertion of GP2 into host membranes causes an extension of the GP2 trimers into an energetically unfavorable state. This also causes the two heptad repeat regions (HRs) within GP2 to be fully exposed to the physiological conditions within the acidified host endosome, which may aid in further triggering of GP2 to promote final collapse of the two HRs [81].
The N-terminal GP2 heptad repeat region (HR1) is a highly ordered, alpha-helical structure that serves as a platform or base for GP1 and contains residues that are necessary for interactions with other GP2 HR1s to form the trimeric structure of the pre-fusion GP [3,7]. In the pre-fusion form, HR1 does not interact with the carboxy terminal heptad repeat region (HR2). Within the intact, pre-fusion trimer, HR2 appears to be disordered and was not included in the crystallized structure [7].
Following insertion of EBOV GP2 fusion loop into the host membrane, the EBOV GP2 trimeric heptad repeats collapse and form a six-helix bundle containing three HR1 and HR2 domains. The collapse into a coiled coil structure draws the two membranes into proximity allowing partial fusion (hemifusion) of the viral membrane and the host endosomal membrane [8,9]. Hemifusion eventually leads to complete fusion of the viral and host endosomal membranes, and an opening through which the viral RNA and its associated proteins can be released into the host cell cytoplasm, where the viral life cycle continues.
11. Potential Therapeutics against Filovirus Entry
This complex set of entry events summarized in Figure 2 provides numerous potential avenues for the development of antiviral therapies against filovirus infection. These possibilities include: (1) interfering with adherence to permissive cells by blocking C-type lectin (or other adherence factor) interactions with the filoviral GP; (2) blocking binding of GP to TIM-1 and/or other cellular receptors that are identified in the future; (3) preventing virus uptake by blocking macropinocytosis; (4) interfering with cathepsin activity; (5) inhibiting availability of NPC1 within the lysosomal compartment and (6) blocking HR1/HR2 (coiled coil) interactions. It is likely that some of these steps will be more amendable than others to the development of antivirals that have minimal or no cytotoxicity. Regardless, elucidation of these entry events provides clear targets for the development of drugs that may prevent both filovirus infection and disease.
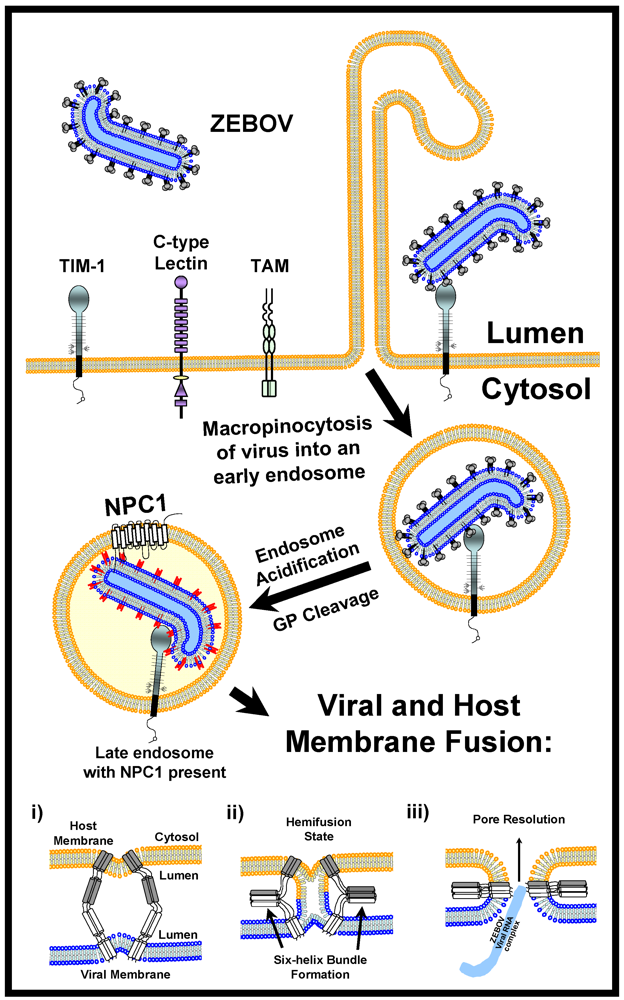
Figure 2.
Model for filoviral entry. Trimers of filoviral GPs on virions interact with both attachment factors (C-type lectins) and receptors (TIM-1) on the surface of permissive cells. Attachment factors are likely to concentrate virions on cells before receptor engagement and virion internalization by macropinocytosis. Macropinocytosis is enhanced by tyrosine kinase receptors such as TAM family members. Following endosomal acidification, Cathepsins L and B trim the EBOV GP to a smaller form that needs at least one as yet undetermined factor to elicit GP fusion with host endosomal membranes. This smaller form of GP is able to interact with both TIM-1 and the endosomal portion of the NPC1 protein; however, whether GP and TIM-1 interact within endosomes is not known. The energetically unfavorable insertion of the EBOV GP2 fusion loop into host endosomal membranes (i) is followed by the energetically favorable collapse of EBOV GP into a six-helix bundle (ii) allowing for lipid mixing and hemifusion of host and viral membrane lipids (ii). Finally, the hemifused host and viral membranes resolve and a complete pore is formed (iii) through which the viral genomic complex passes into the cytoplasm, allowing the viral replication cycle to continue.
Figure 2.
Model for filoviral entry. Trimers of filoviral GPs on virions interact with both attachment factors (C-type lectins) and receptors (TIM-1) on the surface of permissive cells. Attachment factors are likely to concentrate virions on cells before receptor engagement and virion internalization by macropinocytosis. Macropinocytosis is enhanced by tyrosine kinase receptors such as TAM family members. Following endosomal acidification, Cathepsins L and B trim the EBOV GP to a smaller form that needs at least one as yet undetermined factor to elicit GP fusion with host endosomal membranes. This smaller form of GP is able to interact with both TIM-1 and the endosomal portion of the NPC1 protein; however, whether GP and TIM-1 interact within endosomes is not known. The energetically unfavorable insertion of the EBOV GP2 fusion loop into host endosomal membranes (i) is followed by the energetically favorable collapse of EBOV GP into a six-helix bundle (ii) allowing for lipid mixing and hemifusion of host and viral membrane lipids (ii). Finally, the hemifused host and viral membranes resolve and a complete pore is formed (iii) through which the viral genomic complex passes into the cytoplasm, allowing the viral replication cycle to continue.

Acknowledgments
We would like to thank Sven Moller-Tank, Bethany Rhein and Andrew Kondratowicz for critically reading the manuscript and their helpful suggestions and discussions.
Conflict of Interest
The authors declare no conflict of interest.
References and Notes
- Falzarano, D.; Krokhin, O.; Wahl-Jensen, V.; Seebach, J.; Wolf, K.; Schnittler, H.J.; Feldmann, H. Structure-function analysis of the soluble glycoprotein, sgp, of ebola virus. Chembiochem 2006, 7, 1605–1611. [Google Scholar] [CrossRef]
- Mehedi, M.; Falzarano, D.; Seebach, J.; Hu, X.; Carpenter, M.S.; Schnittler, H.J.; Feldmann, H. A new ebola virus nonstructural glycoprotein expressed through RNA editing. J. Virol. 2011, 85, 5406–5414. [Google Scholar]
- Lee, J.E.; Saphire, E.O. Ebolavirus glycoprotein structure and mechanism of entry. Future Virol. 2009, 4, 621–635. [Google Scholar]
- Neumann, G.; Feldmann, H.; Watanabe, S.; Lukashevich, I.; Kawaoka, Y. Reverse genetics demonstrates that proteolytic processing of the ebola virus glycoprotein is not essential for replication in cell culture. J. Virol. 2002, 76, 406–410. [Google Scholar]
- Sanchez, A.; Yang, Z.Y.; Xu, L.; Nabel, G.J.; Crews, T.; Peters, C.J. Biochemical analysis of the secreted and virion glycoproteins of ebola virus. J. Virol. 1998, 72, 6442–6447. [Google Scholar]
- White, J.M.; Delos, S.E.; Brecher, M.; Schornberg, K. Structures and mechanisms of viral membrane fusion proteins: Multiple variations on a common theme. Crit. Rev. Biochem. Mol. Biol. 2008, 43, 189–219. [Google Scholar]
- Lee, J.E.; Fusco, M.L.; Hessell, A.J.; Oswald, W.B.; Burton, D.R.; Saphire, E.O. Structure of the ebola virus glycoprotein bound to an antibody from a human survivor. Nature 2008, 454, 177–182. [Google Scholar]
- Malashkevich, V.N.; Schneider, B.J.; McNally, M.L.; Milhollen, M.A.; Pang, J.X.; Kim, P.S. Core structure of the envelope glycoprotein gp2 from ebola virus at 1.9-a resolution. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 2662–2667. [Google Scholar]
- Weissenhorn, W.; Carfi, A.; Lee, K.H.; Skehel, J.J.; Wiley, D.C. Crystal structure of the ebola virus membrane fusion subunit, gp2, from the envelope glycoprotein ectodomain. Mol. Cell 1998, 2, 605–616. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Radoshitzky, S.R.; Guth, A.C.; Warfield, K.L.; Li, W.; Vincent, M.J.; Towner, J.S.; Nichol, S.T.; Bavari, S.; Choe, H.; et al. Conserved receptor-binding domains of lake victoria marburgvirus and zaire ebolavirus bind a common receptor. J. Biol. Chem. 2006, 281, 15951–15958. [Google Scholar]
- Manicassamy, B.; Wang, J.; Rumschlag, E.; Tymen, S.; Volchkova, V.; Volchkov, V.; Rong, L. Characterization of marburg virus glycoprotein in viral entry. Virology 2007, 358, 79–88. [Google Scholar]
- Brindley, M.A.; Hughes, L.; Ruiz, A.; McCray, P.B., Jr.; Sanchez, A.; Sanders, D.A.; Maury, W. Ebola virus glycoprotein 1: Identification of residues important for binding and postbinding events. J. Virol. 2007, 81, 7702–7709. [Google Scholar]
- Dube, D.; Brecher, M.B.; Delos, S.E.; Rose, S.C.; Park, E.W.; Schornberg, K.L.; Kuhn, J.H.; White, J.M. The primed ebolavirus glycoprotein (19-kilodalton gp1,2): Sequence and residues critical for host cell binding. J. Virol. 2009, 83, 2883–2891. [Google Scholar]
- Jeffers, S.A.; Sanders, D.A.; Sanchez, A. Covalent modifications of the ebola virus glycoprotein. J. Virol. 2002, 76, 12463–12472. [Google Scholar]
- Yang, Z.Y.; Duckers, H.J.; Sullivan, N.J.; Sanchez, A.; Nabel, E.G.; Nabel, G.J. Identification of the ebola virus glycoprotein as the main viral determinant of vascular cell cytotoxicity and injury. Nat. Med. 2000, 6, 886–889. [Google Scholar]
- Sullivan, N.J.; Peterson, M.; Yang, Z.Y.; Kong, W.P.; Duckers, H.; Nabel, E.; Nabel, G.J. Ebola virus glycoprotein toxicity is mediated by a dynamin-dependent protein-trafficking pathway. J. Virol. 2005, 79, 547–553. [Google Scholar]
- Jaskierny, A.J.; Panahi, A.; Feig, M. Effect of flanking residues on the conformational sampling of the internal fusion peptide from ebola virus. Proteins 2011, 79, 1109–1117. [Google Scholar]
- Watanabe, S.; Takada, A.; Watanabe, T.; Ito, H.; Kida, H.; Kawaoka, Y. Functional importance of the coiled-coil of the ebola virus glycoprotein. J. Virol. 2000, 74, 10194–10201. [Google Scholar]
- Usami, K.; Matsuno, K.; Igarashi, M.; Denda-Nagai, K.; Takada, A.; Irimura, T. Involvement of viral envelope gp2 in ebola virus entry into cells expressing the macrophage galactose-type c-type lectin. Biochem. Biophys. Res. Commun. 2011, 407, 74–78. [Google Scholar]
- Han, Z.; Licata, J.M.; Paragas, J.; Harty, R.N. Permeabilization of the plasma membrane by ebola virus gp2. Virus Genes 2007, 34, 273–281. [Google Scholar]
- Adam, B.; Lins, L.; Stroobant, V.; Thomas, A.; Brasseur, R. Distribution of hydrophobic residues is crucial for the fusogenic properties of the ebola virus gp2 fusion peptide. J. Virol. 2004, 78, 2131–2136. [Google Scholar]
- Ito, H.; Watanabe, S.; Sanchez, A.; Whitt, M.A.; Kawaoka, Y. Mutational analysis of the putative fusion domain of ebola virus glycoprotein. J. Virol. 1999, 73, 8907–8912. [Google Scholar]
- Ruiz-Arguello, M.B.; Goni, F.M.; Pereira, F.B.; Nieva, J.L. Phosphatidylinositol-dependent membrane fusion induced by a putative fusogenic sequence of ebola virus. J. Virol. 1998, 72, 1775–1781. [Google Scholar]
- Plemper, R.K. Cell entry of enveloped viruses. Curr. Opin. Virol. 2011, 1, 92–100. [Google Scholar]
- Schroth-Diez, B.; Ludwig, K.; Baljinnyam, B.; Kozerski, C.; Huang, Q.; Herrmann, A. The role of the transmembrane and of the intraviral domain of glycoproteins in membrane fusion of enveloped viruses. Biosci. Rep. 2000, 20, 571–595. [Google Scholar]
- Ito, H.; Watanabe, S.; Takada, A.; Kawaoka, Y. Ebola virus glycoprotein: Proteolytic processing, acylation, cell tropism, and detection of neutralizing antibodies. J. Virol. 2001, 75, 1576–1580. [Google Scholar] [CrossRef]
- Feldmann, H.; Volchkov, V.E.; Volchkova, V.A.; Klenk, H.D. The glycoproteins of marburg and ebola virus and their potential roles in pathogenesis. Arch. Virol. Suppl. 1999, 15, 159–169. [Google Scholar]
- Geisbert, T.W.; Hensley, L.E. Ebola virus: New insights into disease aetiopathology and possible therapeutic interventions. Expert Rev. Mol. Med. 2004, 6, 1–24. [Google Scholar]
- Geisbert, T.W.; Hensley, L.E.; Larsen, T.; Young, H.A.; Reed, D.S.; Geisbert, J.B.; Scott, D.P.; Kagan, E.; Jahrling, P.B.; Davis, K.J. Pathogenesis of ebola hemorrhagic fever in cynomolgus macaques: Evidence that dendritic cells are early and sustained targets of infection. Am. J. Pathol. 2003, 163, 2347–2370. [Google Scholar]
- Wool-Lewis, R.J.; Bates, P. Characterization of ebola virus entry by using pseudotyped viruses: Identification of receptor-deficient cell lines. J. Virol. 1998, 72, 3155–3160. [Google Scholar]
- Yang, Z.; Delgado, R.; Xu, L.; Todd, R.F.; Nabel, E.G.; Sanchez, A.; Nabel, G.J. Distinct cellular interactions of secreted and transmembrane ebola virus glycoproteins. Science 1998, 279, 1034–1037. [Google Scholar]
- Breman, J.G.; Johnson, K.M.; van der Groen, G.; Robbins, C.B.; Szczeniowski, M.V.; Ruti, K.; Webb, P.A.; Meier, F.; Heymann, D.L. A search for ebola virus in animals in the democratic republic of the Congo and Cameroon: Ecologic, virologic, and serologic surveys, 1979–1980. Ebola virus study teams. J. Infect. Dis. 1999, 179, S139–S147. [Google Scholar]
- Chan, S.Y.; Speck, R.F.; Ma, M.C.; Goldsmith, M.A. Distinct mechanisms of entry by envelope glycoproteins of marburg and ebola (zaire) viruses. J. Virol. 2000, 74, 4933–4937. [Google Scholar]
- Geisbert, T.W.; Hensley, L.E.; Gibb, T.R.; Steele, K.E.; Jaax, N.K.; Jahrling, P.B. Apoptosis induced in vitro and in vivo during infection by ebola and marburg viruses. Lab. Invest. 2000, 80, 171–186. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Jahrling, P.B.; Hanes, M.A.; Zack, P.M. Association of ebola-related reston virus particles and antigen with tissue lesions of monkeys imported to the united states. J. Comp. Pathol. 1992, 106, 137–152. [Google Scholar]
- Simmons, G.; Rennekamp, A.J.; Chai, N.; Vandenberghe, L.H.; Riley, J.L.; Bates, P. Folate receptor alpha and caveolae are not required for ebola virus glycoprotein-mediated viral infection. J. Virol. 2003, 77, 13433–13438. [Google Scholar]
- Simmons, G.; Reeves, J.D.; Grogan, C.C.; Vandenberghe, L.H.; Baribaud, F.; Whitbeck, J.C.; Burke, E.; Buchmeier, M.J.; Soilleux, E.J.; Riley, J.L.; et al. Dc-sign and dc-signr bind ebola glycoproteins and enhance infection of macrophages and endothelial cells. Virology 2003, 305, 115–123. [Google Scholar]
- Albritton, L.M.; Tseng, L.; Scadden, D.; Cunningham, J.M. A putative murine ecotropic retrovirus receptor gene encodes a multiple membrane-spanning protein and confers susceptibility to virus infection. Cell 1989, 57, 659–666. [Google Scholar]
- O'Hara, B.; Johann, S.V.; Klinger, H.P.; Blair, D.G.; Rubinson, H.; Dunn, K.J.; Sass, P.; Vitek, S.M.; Robins, T. Characterization of a human gene conferring sensitivity to infection by gibbon ape leukemia virus. Cell Growth Differ. 1990, 1, 119–127. [Google Scholar]
- Bates, P.; Young, J.A.; Varmus, H.E. A receptor for subgroup a rous sarcoma virus is related to the low density lipoprotein receptor. Cell 1993, 74, 1043–1051. [Google Scholar]
- Shimojima, M.; Takada, A.; Ebihara, H.; Neumann, G.; Fujioka, K.; Irimura, T.; Jones, S.; Feldmann, H.; Kawaoka, Y. Tyro3 family-mediated cell entry of ebola and marburg viruses. J. Virol. 2006, 80, 10109–10116. [Google Scholar]
- Chan, S.Y.; Empig, C.J.; Welte, F.J.; Speck, R.F.; Schmaljohn, A.; Kreisberg, J.F.; Goldsmith, M.A. Folate receptor-alpha is a cofactor for cellular entry by marburg and ebola viruses. Cell 2001, 106, 117–126. [Google Scholar]
- Kondratowicz, A.S.; Lennemann, N.J.; Sinn, P.L.; Davey, R.A.; Hunt, C.L.; Moller-Tank, S.; Meyerholz, D.K.; Rennert, P.; Mullins, R.F.; Brindley, M.; et al. T-cell immunoglobulin and mucin domain 1 (tim-1) is a receptor for zaire ebolavirus and lake victoria marburgvirus. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 8426–8431. [Google Scholar]
- Alvarez, C.P.; Lasala, F.; Carrillo, J.; Muniz, O.; Corbi, A.L.; Delgado, R. C-type lectins dc-sign and l-sign mediate cellular entry by ebola virus in cis and in trans. J. Virol. 2002, 76, 6841–6844. [Google Scholar]
- Becker, S.; Spiess, M.; Klenk, H.D. The asialoglycoprotein receptor is a potential liver-specific receptor for marburg virus. J. Gen. Virol. 1995, 76, 393–399. [Google Scholar]
- Gramberg, T.; Hofmann, H.; Moller, P.; Lalor, P.F.; Marzi, A.; Geier, M.; Krumbiegel, M.; Winkler, T.; Kirchhoff, F.; Adams, D.H.; et al. Lsectin interacts with filovirus glycoproteins and the spike protein of sars coronavirus. Virology 2005, 340, 224–236. [Google Scholar]
- Takada, A.; Fujioka, K.; Tsuiji, M.; Morikawa, A.; Higashi, N.; Ebihara, H.; Kobasa, D.; Feldmann, H.; Irimura, T.; Kawaoka, Y. Human macrophage c-type lectin specific for galactose and n-acetylgalactosamine promotes filovirus entry. J. Virol. 2004, 78, 2943–2947. [Google Scholar]
- Marzi, A.; Gramberg, T.; Simmons, G.; Moller, P.; Rennekamp, A.J.; Krumbiegel, M.; Geier, M.; Eisemann, J.; Turza, N.; Saunier, B.; et al. Dc-sign and dc-signr interact with the glycoprotein of marburg virus and the s protein of severe acute respiratory syndrome coronavirus. J. Virol. 2004, 78, 12090–12095. [Google Scholar]
- Marzi, A.; Wegele, A.; Pohlmann, S. Modulation of virion incorporation of ebolavirus glycoprotein: Effects on attachment, cellular entry and neutralization. Virology 2006, 352, 345–356. [Google Scholar]
- Chandran, K.; Sullivan, N.J.; Felbor, U.; Whelan, S.P.; Cunningham, J.M. Endosomal proteolysis of the ebola virus glycoprotein is necessary for infection. Science 2005, 308, 1643–1645. [Google Scholar]
- Schornberg, K.; Matsuyama, S.; Kabsch, K.; Delos, S.; Bouton, A.; White, J. Role of endosomal cathepsins in entry mediated by the ebola virus glycoprotein. J. Virol. 2006, 80, 4174–4178. [Google Scholar]
- Kaletsky, R.L.; Simmons, G.; Bates, P. Proteolysis of the ebola virus glycoproteins enhances virus binding and infectivity. J. Virol. 2007, 81, 13378–13384. [Google Scholar]
- Matsuno, K.; Nakayama, E.; Noyori, O.; Marzi, A.; Ebihara, H.; Irimura, T.; Feldmann, H.; Takada, A. C-type lectins do not act as functional receptors for filovirus entry into cells. Biochem. Biophys. Res. Commun. 2010, 403, 144–148. [Google Scholar]
- Tsegaye, T.S.; Pohlmann, S. The multiple facets of HIV attachment to dendritic cell lectins. Cell Microbiol. 2010, 12, 1553–1561. [Google Scholar]
- Simmons, G.; Wool-Lewis, R.J.; Baribaud, F.; Netter, R.C.; Bates, P. Ebola virus glycoproteins induce global surface protein down-modulation and loss of cell adherence. J. Virol. 2002, 76, 2518–2528. [Google Scholar]
- Takada, A.; Watanabe, S.; Ito, H.; Okazaki, K.; Kida, H.; Kawaoka, Y. Downregulation of beta1 integrins by ebola virus glycoprotein: Implication for virus entry. Virology 2000, 278, 20–26. [Google Scholar]
- Schornberg, K.L.; Shoemaker, C.J.; Dube, D.; Abshire, M.Y.; Delos, S.E.; Bouton, A.H.; White, J.M. Alpha5beta1-integrin controls ebolavirus entry by regulating endosomal cathepsins. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 8003–8008. [Google Scholar]
- Linger, R.M.; Keating, A.K.; Earp, H.S.; Graham, D.K. Tam receptor tyrosine kinases: Biologic functions, signaling, and potential therapeutic targeting in human cancer. Adv. Cancer Res. 2008, 100, 35–83. [Google Scholar] [CrossRef]
- Shimojima, M.; Ikeda, Y.; Kawaoka, Y. The mechanism of axl-mediated ebola virus infection. J. Infect. Dis. 2007, 196, S259–S263. [Google Scholar]
- Brindley, M.A.; Hunt, C.L.; Kondratowicz, A.S.; Bowman, J.; Sinn, P.L.; McCray, P.B., Jr.; Quinn, K.; Weller, M.L.; Chiorini, J.A.; Maury, W. Tyrosine kinase receptor axl enhances entry of zaire ebolavirus without direct interactions with the viral glycoprotein. Virology 2011, 415, 83–94. [Google Scholar]
- Hunt, C.L.; Kolokoltsov, A.A.; Davey, R.A.; Maury, W. The tyro3 receptor kinase axl enhances macropinocytosis of zaire ebolavirus. J. Virol. 2011, 85, 334–347. [Google Scholar]
- Saeed, M.F.; Kolokoltsov, A.A.; Albrecht, T.; Davey, R.A. Cellular entry of ebola virus involves uptake by a macropinocytosis-like mechanism and subsequent trafficking through early and late endosomes. PLoS Pathog. 2010, 6. [Google Scholar]
- Nanbo, A.; Imai, M.; Watanabe, S.; Noda, T.; Takahashi, K.; Neumann, G.; Halfmann, P.; Kawaoka, Y. Ebolavirus is internalized into host cells via macropinocytosis in a viral glycoprotein-dependent manner. PLoS Pathog. 2010, 6. [Google Scholar]
- Weller, M.L.; Amornphimoltham, P.; Schmidt, M.; Wilson, P.A.; Gutkind, J.S.; Chiorini, J.A. Epidermal growth factor receptor is a co-receptor for adeno-associated virus serotype 6. Nat. Med. 2010, 16, 662–664. [Google Scholar]
- Yonezawa, A.; Cavrois, M.; Greene, W.C. Studies of ebola virus glycoprotein-mediated entry and fusion by using pseudotyped human immunodeficiency virus type 1 virions: Involvement of cytoskeletal proteins and enhancement by tumor necrosis factor alpha. J. Virol. 2005, 79, 918–926. [Google Scholar]
- Bavari, S.; Bosio, C.M.; Wiegand, E.; Ruthel, G.; Will, A.B.; Geisbert, T.W.; Hevey, M.; Schmaljohn, C.; Schmaljohn, A.; Aman, M.J. Lipid raft microdomains: A gateway for compartmentalized trafficking of ebola and marburg viruses. J. Exp. Med. 2002, 195, 593–602. [Google Scholar]
- Empig, C.J.; Goldsmith, M.A. Association of the caveola vesicular system with cellular entry by filoviruses. J. Virol. 2002, 76, 5266–5270. [Google Scholar]
- Bhattacharyya, S.; Hope, T.J.; Young, J.A. Differential requirements for clathrin endocytic pathway components in cellular entry by ebola and marburg glycoprotein pseudovirions. Virology 2011, 419, 1–9. [Google Scholar]
- Bhattacharyya, S.; Warfield, K.L.; Ruthel, G.; Bavari, S.; Aman, M.J.; Hope, T.J. Ebola virus uses clathrin-mediated endocytosis as an entry pathway. Virology 2010, 401, 18–28. [Google Scholar]
- Sanchez, A. Analysis of filovirus entry into vero e6 cells, using inhibitors of endocytosis, endosomal acidification, structural integrity, and cathepsin (b and l) activity. J. Infect. Dis. 2007, 196, S251–S258. [Google Scholar] [CrossRef]
- Aleksandrowicz, P.; Marzi, A.; Biedenkopf, N.; Beimforde, N.; Becker, S.; Hoenen, T.; Feldmann, H.; Schnittler, H.J. Ebola virus enters host cells by macropinocytosis and clathrin-mediated endocytosis. J. Infect. Dis. 2011, 204, S957–S967. [Google Scholar]
- Dolnik, O.; Kolesnikova, L.; Becker, S. Filoviruses: Interactions with the host cell. Cell Mol. Life Sci. 2008, 65, 756–776. [Google Scholar]
- Lim, J.P.; Gleeson, P.A. Macropinocytosis: An endocytic pathway for internalising large gulps. Immunol. Cell Biol. 2011, 89, 836–843. [Google Scholar]
- Cureton, D.K.; Massol, R.H.; Saffarian, S.; Kirchhausen, T.L.; Whelan, S.P. Vesicular stomatitis virus enters cells through vesicles incompletely coated with clathrin that depend upon actin for internalization. PLoS Pathog. 2009, 5. [Google Scholar]
- Rodriguez, N.E.; Gaur, U.; Wilson, M.E. Role of caveolae in leishmania chagasi phagocytosis and intracellular survival in macrophages. Cell Microbiol. 2006, 8, 1106–1120. [Google Scholar]
- Quinn, K.; Brindley, M.A.; Weller, M.L.; Kaludov, N.; Kondratowicz, A.; Hunt, C.L.; Sinn, P.L.; McCray, P.B., Jr.; Stein, C.S.; Davidson, B.L.; et al. Rho gtpases modulate entry of ebola virus and vesicular stomatitis virus pseudotyped vectors. J. Virol. 2009, 83, 10176–10186. [Google Scholar]
- Cote, M.; Zheng, Y.M.; Liu, S.L. Receptor binding and low ph coactivate oncogenic retrovirus envelope-mediated fusion. J. Virol. 2009, 83, 11447–11455. [Google Scholar]
- Brindley, M.A.; Maury, W. Endocytosis and a low-ph step are required for productive entry of equine infectious anemia virus. J. Virol. 2005, 79, 14482–14488. [Google Scholar]
- Barnard, R.J.; Young, J.A. Alpharetrovirus envelope-receptor interactions. Curr. Top. Microbiol. Immunol. 2003, 281, 107–136. [Google Scholar]
- Wong, A.C.; Sandesara, R.G.; Mulherkar, N.; Whelan, S.P.; Chandran, K. A forward genetic strategy reveals destabilizing mutations in the ebolavirus glycoprotein that alter its protease dependence during cell entry. J. Virol. 2010, 84, 163–175. [Google Scholar]
- Brecher, M.; Schornberg, K.L.; Delos, S.E.; Fusco, M.L.; Saphire, E.O.; White, J.M. Cathepsin cleavage potentiates the ebola virus glycoprotein to undergo a subsequent fusion relevant conformational change. J. Virol. 2011. [Google Scholar]
- Martinez, O.; Johnson, J.; Manicassamy, B.; Rong, L.; Olinger, G.G.; Hensley, L.E.; Basler, C.F. Zaire ebola virus entry into human dendritic cells is insensitive to cathepsin l inhibition. Cell Microbiol. 2010, 12, 148–157. [Google Scholar]
- Cote, M.; Misasi, J.; Ren, T.; Bruchez, A.; Lee, K.; Filone, C.M.; Hensley, L.; Li, Q.; Ory, D.; Chandran, K.; et al. Small molecule inhibitors reveal niemann-pick c1 is essential for ebola virus infection. Nature 2011, 477, 344–348. [Google Scholar]
- Carette, J.E.; Raaben, M.; Wong, A.C.; Herbert, A.S.; Obernosterer, G.; Mulherkar, N.; Kuehne, A.I.; Kranzusch, P.J.; Griffin, A.M.; Ruthel, G.; et al. Ebola virus entry requires the cholesterol transporter niemann-pick c1. Nature 2011, 477, 340–343. [Google Scholar]
- Rosenbaum, A.I.; Maxfield, F.R. Niemann-pick type c disease: Molecular mechanisms and potential therapeutic approaches. J. Neurochem. 2011, 116, 789–795. [Google Scholar]
- Poh, M.K.; Shui, G.; Xie, X.; Shi, P.Y.; Wenk, M.R.; Gu, F. U18666a, an intra-cellular cholesterol transport inhibitor, inhibits dengue virus entry and replication. Antivir. Res. 2012, 93, 191–198. [Google Scholar]
- Lai, C.K.; Jeng, K.S.; Machida, K.; Lai, M.M. Hepatitis c virus egress and release depend on endosomal trafficking of core protein. J. Virol. 2010, 84, 11590–11598. [Google Scholar]
- Hagiwara, K.; Nakamura, Y.; Nishijima, M.; Yamakawa, Y. Prevention of prion propagation by dehydrocholesterol reductase inhibitors in cultured cells and a therapeutic trial in mice. Biol. Pharm. Bull. 2007, 30, 835–838. [Google Scholar]
- Sainz, B., Jr.; Barretto, N.; Martin, D.N.; Hiraga, N.; Imamura, M.; Hussain, S.; Marsh, K.A.; Yu, X.; Chayama, K.; Alrefai, W.A.; et al. Identification of the niemann-pick c1-like 1 cholesterol absorption receptor as a new hepatitis c virus entry factor. Nat. Med. 2012. [Google Scholar]
- Gregory, S.M.; Harada, E.; Liang, B.; Delos, S.E.; White, J.M.; Tamm, L.K. Structure and function of the complete internal fusion loop from ebolavirus glycoprotein 2. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 11211–11216. [Google Scholar]
- Bale, S.; Liu, T.; Li, S.; Wang, Y.; Abelson, D.; Fusco, M.; Woods, V.L., Jr.; Ollmann Saphire, E. Ebola virus glycoprotein needs an additional trigger, beyond proteolytic priming for membrane fusion. PLoS Negl. Trop. Dis. 2011, 5. [Google Scholar]
- Freitas, M.S.; Gaspar, L.P.; Lorenzoni, M.; Almeida, F.C.; Tinoco, L.W.; Almeida, M.S.; Maia, L.F.; Degreve, L.; Valente, A.P.; Silva, J.L. Structure of the ebola fusion peptide in a membrane-mimetic environment and the interaction with lipid rafts. J. Biol. Chem. 2007, 282, 27306–27314. [Google Scholar]
- Gomara, M.J.; Mora, P.; Mingarro, I.; Nieva, J.L. Roles of a conserved proline in the internal fusion peptide of ebola glycoprotein. FEBS Lett. 2004, 569, 261–266. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Hunt, C.L.; Lennemann, N.J.; Maury, W. Filovirus Entry: A Novelty in the Viral Fusion World. Viruses 2012, 4, 258-275. https://doi.org/10.3390/v4020258
AMA Style
Hunt CL, Lennemann NJ, Maury W. Filovirus Entry: A Novelty in the Viral Fusion World. Viruses. 2012; 4(2):258-275. https://doi.org/10.3390/v4020258
Chicago/Turabian StyleHunt, Catherine L., Nicholas J. Lennemann, and Wendy Maury. 2012. "Filovirus Entry: A Novelty in the Viral Fusion World" Viruses 4, no. 2: 258-275. https://doi.org/10.3390/v4020258