Photodynamic Inactivation of Mammalian Viruses and Bacteriophages

 ,
, 
Abstract
:Nomenclature
| AlPcS4 | Aluminum phthalocyanine tetrasulfonate |
| AZT | Azidothymidine |
| BVDV | Bovine viral diarrhea virus |
| DMTU | Dimethylthiourea |
| EMCV | Encephalomyocarditis virus |
| HAV | Hepatitis A virus |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| HIV | Human immunodeficiency virus |
| HPV | Human papillomatosis virus |
| HSV | Herpes simplex virus |
| LED | Light emitting diode |
| MB | Methylene blue |
| NM | Not mentioned |
| NQ | Not quantified |
| Pc4 | Silicon phthalocyanine |
| PDI | Photodynamic inactivation |
| PS | Photosensitizer |
| ROS | Reactive oxygen species |
| SFV | Semliki Forest virus |
| SHV | Suid herpes virus |
| SOD | Superoxide dismutase |
| SSB | Singlet strand breaks |
| Tri-Py+-Me-PF | 5-(pentafluorophenyl)-10,15,20-tris(1-methylpyridinium-4-yl)porphyrin tri-iodide |
| VSV | Vesicular stomatitis virus |
| VZV | Varicella zoster virus |
| 1O2 | Singlet oxygen |
| 3O2 | Molecular oxygen |
| 1PS | Ground state photosensitizer |
| 3PS* | Triplet excited state photosensitizer |
1. Introduction
2. Antimicrobial PDI
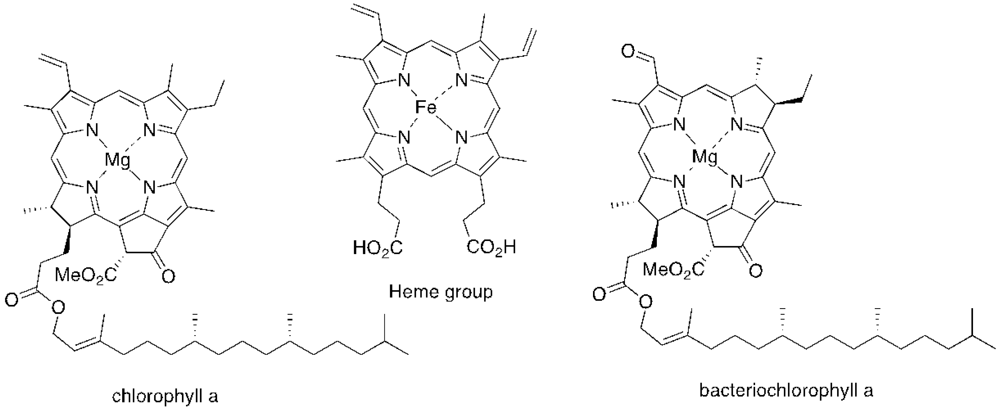
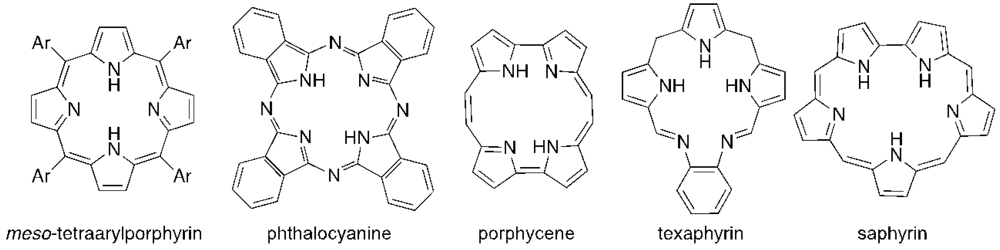
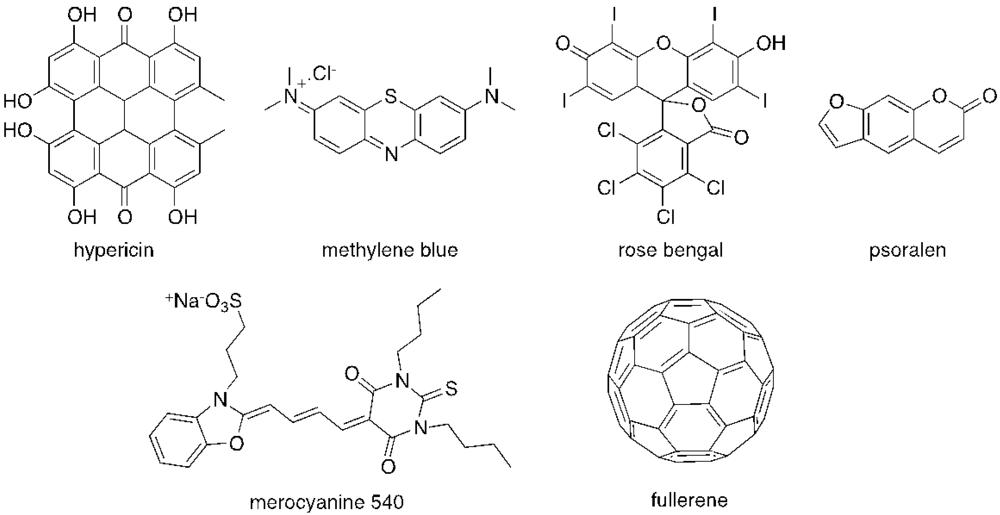

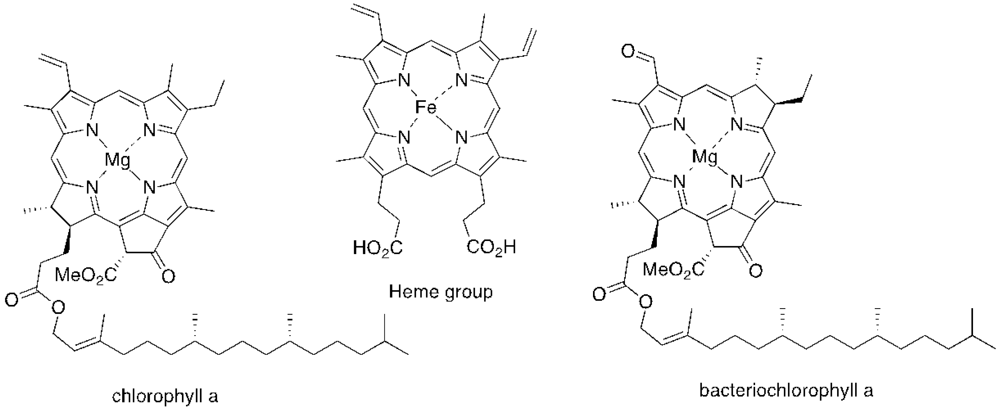{kind=link}
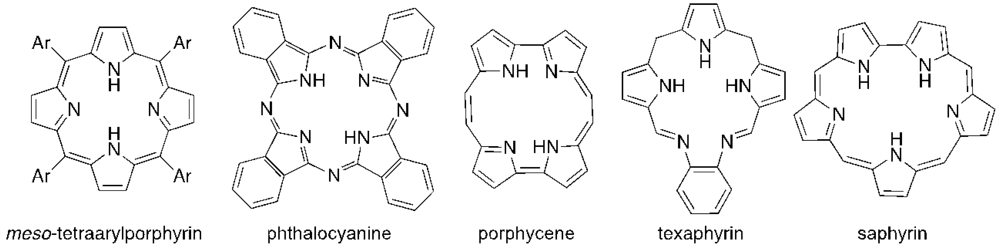{kind=link}
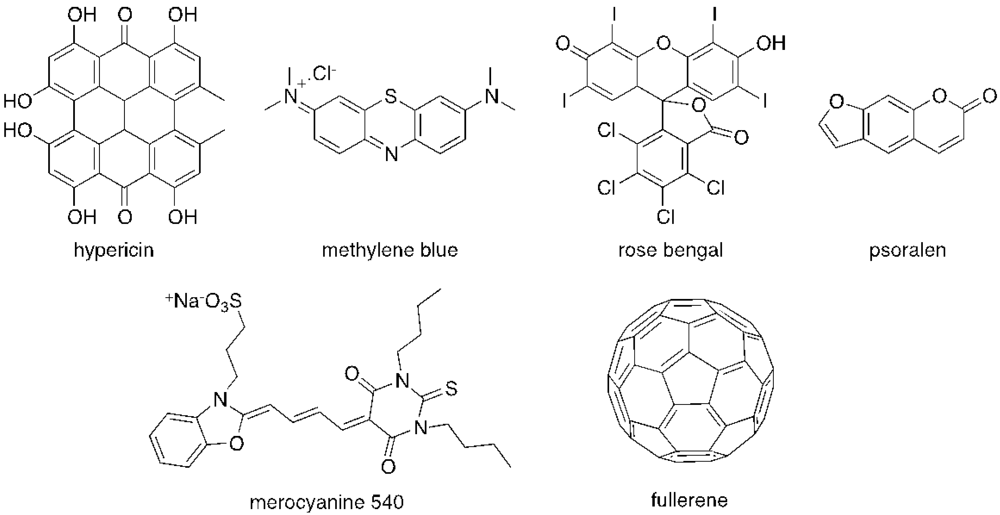{kind=link}
{kind=link}
| Photosensitizer | Microorganism | PDI | Reference |
|---|---|---|---|
| Mammalian viruses | |||
| Hematoporphyrin derivative | HSV-1 | 7 log | [75] |
| HSV-1 | <0.8 log | [36] | |
| Uroporphyrin | Adenovirus | 7 log | [76] |
| Natural metalloporphyrin derivatives | HIV-1 | <0.8 log | [36] |
| Chlorophyll derivatives | VSV | ~6 log | [77] |
| 7-despropionate-7-hydroxypropylmesopyropheophorbide a | BVDV | ~5 log | [78] |
| Benzoporphyrin derivative monoacid ring A | HIV-1 | >4 log | [33] |
| Glycoconjugated meso-tetraarylporphyrin derivatives | HSV-1 | 6 log | [79] |
| Metallo tetrasulfonated meso-tetraarylporphyrin derivatives | HIV-1 | ≤2 log | [36] |
| Tetrasulfonated meso-tetraarylporphyrin derivatives | HIV-1 | ≤2 log | [36] |
| HAV | ~4 log | [44] | |
| meso-Tetrakis(1-methylpyridinium-4-yl)porphyrin | HAV | ~4 log | [44] |
| meso-Tetrakis(1-butylpyridinium-4-yl)porphyrin | HAV | >3.8 log | [44] |
| meso-Tetrakis(1-octylpyridinium-4-yl)porphyrin | HAV | >3.9 log | [44] |
| Cationic β-vinyl substituted meso-tetraphenylporphyrin derivatives | HSV-1 | <3 log | [80] |
| Aluminum dibenzodisulfophthalocyanine | HIV-1 | 3.7 log | [49] |
| Aluminum phthalocyanine tetrasulfonate | HIV-1 | >5 log | [49] |
| VSV | 4.2 log | [82] | |
| Adenovirus | 4 log | [76] | |
| Silicon phthalocyanine derivative | VSV | 4 log | [82] |
| Cationic phthalocyanines | HIV-1 | >5 log | [49] |
| HSV-1 | ≥5 log | [83] | |
| Hypericin | HIV-1 | NQ | [30] |
| VSV | 4-5 log | ||
| Influenza virus | NQ | ||
| Sendai virus | NQ | ||
| Methylene blue | VSV | 4.7 log | [81] |
| HSV-1 | 5 log | [84] | |
| SHV-1 | 2.5 log | [84] | |
| HCV | <2 log | [41] | |
| HIV-1 | <2 log | [41] | |
| Adenovirus | 7 log | [76] | |
| Dengue virus | 5–6.4 log | [74] | |
| Enterovirus 71 | ~8 log | [85] | |
| Vaccinia virus | 5 log | [86] | |
| Phenothiazine derivatives | VSV | >4.4 log | [60] |
| Rose bengal | Vaccinia virus | 5 log | [86] |
| HIV-1 | NQ | [30] | |
| VSV | 4–5 log | ||
| Influenza virus | NQ | ||
| Sendai virus | NQ | ||
| Adenovirus | 7 log | [76] | |
| Buckminsterfullerene | SFV | 7 log | [50] |
| Merocyanine 540 | HSV-1 | 5–6 log | [45] |
| Bacteriophages | |||
| Glycoconjugated meso-tetraarylporphyrins | T7 phage | <3 log | [64] |
| T7 phage | <3.5 log | [87] | |
| Tetrasulfonated meso-tetraarylporphyrin derivatives | MS2 phage | >3.8 log | [44] |
| meso-Tetrakis(1-methylpyridinium-4-yl)porphyrin | λ phage | <7 log | [58] |
| MS2 phage | >4.1 log | [44] | |
| T4 phage | 7 log | [66,67] | |
| T7 phage | <4 log | [88] | |
| 5-(pentafluorophenyl)-10,15,20-tris(1-methylpyridinium-4-yl)porphyrin | T4 phage | 7 log | [66,67,68] |
| 5-(4-methoxicarbonylphenyl)-10,15,20-tris(1-methylpyridinium-4-yl)porphyrin | T4 phage | 7 log | [66] |
| 5-(4-carboxyphenyl)-10,15,20-tris(1-methylpyridinium-4-yl)porphyrin | T4 phage | 3.9 log | [66] |
| 5,10-bis(4-carboxyphenyl)-15,20-bis(1-methylpyridinium-4-yl)porphyrin | T4 phage | 1.4 log | [66] |
| 5,15-bis(4-carboxyphenyl)-10,20-bis(1-methylpyridinium-4-yl)porphyrin | T4 phage | 1.2 log | [66] |
| 5,10,15-tris(1-methylpyridinium-4-yl)-20-phenylporphyrin | T7 phage | 1.7 log | [88] |
| Methylene blue | Serratia phage kappa | >4 log | [61] |
| M13 phage | 2.2 log | [52,81] | |
| f2 phage | 5 log | [56] | |
| Qβ phage | 7–8 log | [56] | |
| Qβ phage | 7–8 log | [89] | |
| Phenothiazine derivatives | R17 phage | 4–7 log | [60] |
| Rose bengal | PRD1 phage | ~3.5 log* | [57] |
| Riboflavin | λ phage | <4 log | [59] |
| Proflavine | Serratia phage kappa | 4 log | [61] |
| T3 phage | 7–11 log | [63] | |
| Polyhydroxylated fullerene | MS2 phage | ~4 log | [90] |
| PRD1 phage | ~2.5 log* | [57] | |
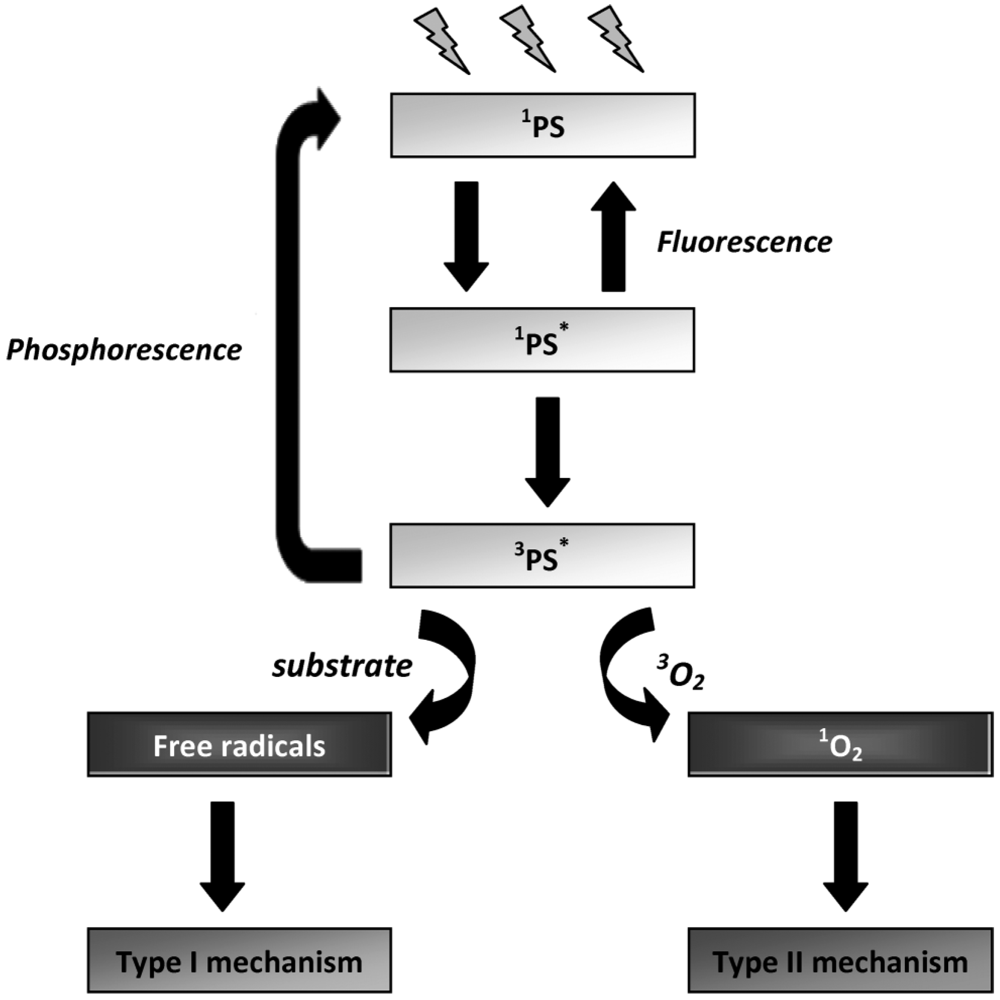
3. Mechanisms of Photodynamic Inactivation
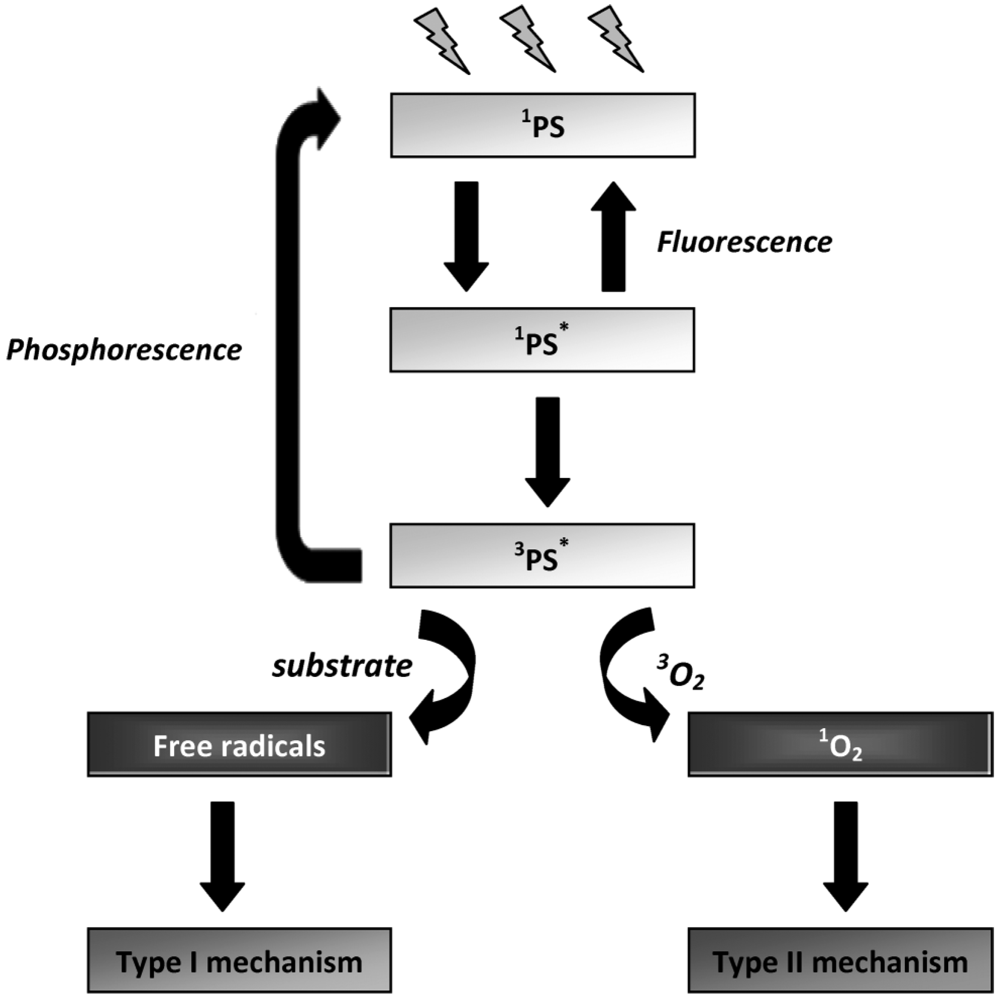
3.1. Type I and Type II Mechanisms

3.2. Evaluation of the Specific Involvement of Type I and Type II Mechanisms
3.2.1. Type I Mechanism Scavengers
| PS | Scavenger | Microorganism | Scavenger protection | Reference |
|---|---|---|---|---|
| Mammalian viruses | ||||
| Aluminum phthalocyanine tetrasulfonate | Reduced glutathione | VSV | Little/no effect | [106] |
| Polyhydroxylated fullerene | Glutathione (2.0 mM) | SFV | no effect | [50] |
| Hydroquinone (2.0 mM) | SFV | no effect | [50] | |
| Merocyanine 540 | Glutathione (10 and 30 mmol L−1) | HSV-1 | 30-50% | [45] |
| Methylene blue | Mannitol (100 mM) | HSV-1 | 24% | [84] |
| Bacteriophages | ||||
| 5,10,15-(4-β- D-glucosylphenyl)-20-phenylporphyrin | DMTU (0.1–5.0 mM) | T7 phage | 44% | [64] |
| 5,10.15,20-Tetrakis(4-β- D-glucosylphenyl) porphyrin | DMTU (0.1–5.0 mM) | T7 phage | 79% | [64] |
| 5,10,15-(4-β- D-galactosylphenyl)-20-(pentafluorophenyl)-porphyrin | DMTU (0.1–5.0 mM) | T7 phage | 89% | [87] |
| 5-(pentafluorophenyl)-10,15,20-tris(1-methylpyridinium-4- yl)porphyrin | D-mannitol (100 mM) | T4 phage | 20% | [107] |
| L-cysteine (100 mM) | T4 phage | 9% | [107] | |
| 5,10,15,20-tetrakis(1-methylpyridinium-4-yl)porphyrin | D-mannitol (100 mM) | T4 phage | no effect | [107] |
| Proflavine | L-cysteine (0.025 M) | T3 phage | 75–80% | [63] |
| Polyhydroxylated fullerene | SOD | MS2 phage | no effect | [90] |
3.2.1.1. Free Radicals in PDI of Mammalian Viruses
3.2.1.2. Free Radicals in PDI of Bacteriophages
3.2.2. Type II Mechanism Quenchers
| PS | Quencher | Microorganism | Quencher protection | Reference |
|---|---|---|---|---|
| Mammalian viruses | ||||
| Aluminum phthalocyanine tetrasulfonate | Sodium azide | VSV | significant effect | [106] |
| Rose bengal | β-carotene | Influenza virus | Significant effect | [108] |
| Hypericin | Sodium azide | HIV | Significant effect | [111] |
| Methylene blue | Imidazole (5.0 and 10 mM) | HSV-1 | 55–75% | [84] |
| Bacteriophages | ||||
| 5,10,15-(4-β- D-galactosylphenyl)-20-(pentafluorophenyl)porphyrin | Sodium azide (0.1–5.0 mM) | T7 phage | 38% | [87] |
| 5-(pentafluorophenyl)-10,15,20-tris(1-methylpyridinium-4-yl)porphyrin | Sodium azide (100 mM) | T4 phage | 80% | [107] |
| L-histidine (50 mM) | T4 phage | 74% | ||
| meso-tetrakis(1-methylpyridinium-4-yl)porphyrin | Sodium azide (100 mM) | T4 phage | 90% | [107] |
| L-histidine (100 mM) | T4 phage | 78% | ||
| 5,10,15,20-Tetrakis(4-β- D-glucosylphenyl)porphyrin | 1,3-diphenylisobenzofuran (0.1-5.0 mM) | T7 phage | 42% | [64] |
| 5,10,15-(4-β- D-glucosylphenyl)-20-phenylporphyrin | 1,3-diphenylisobenzofuran (0.1-5.0 mM) | T7 phage | 74% | [64] |
| Polyhydroxylated fullerene | β-carotene | T7 phage | 69% | [57] |
| β-carotene (26 μM) | MS2 phage | 50–60% | [90] | |
| Rose bengal | Sodium azide (3.5–35 mM) | M13 phage | 31% | [52] |
3.2.2.1. Singlet Oxygen in PDI of Mammalian Viruses
3.2.2.2. Singlet Oxygen in PDI of Bacteriophages
4. Molecular Targets of Antiviral PDI
4.1. Nucleic Acids
4.1.1. DNA Damage
4.1.1.1. Damages in the DNA of Mammalian Viruses
4.1.1.2. Damage in the DNA of Bacteriophages
4.1.2. RNA Damage
4.1.2.1. Damage in the RNA of Mammalian Viruses
4.1.2.2. Damage in the RNA of Bacteriophages
4.2. Outer Structures
| Virus | Type of damage | PS | Reference |
|---|---|---|---|
| Enveloped-mammalian viruses | |||
| HSV-1 | Viral envelope (reduced ability to adhere to and penetrate host cells) | Merocyanine 540 | [45] |
| Viral envelope (prevention of viral adsorption and host penetration) | Phthalocyanine derivatives | [131] | |
| Glycoprotein D; loss of proteins; dimerization; protein cross-links; alterations in protein molecular mass and charge | Phthalocyanine derivatives | [83] | |
| HSV-2 | Viral envelope (prevention of viral adsorption and host penetration) | Phthalocyanine derivatives | [131] |
| HSV | Protein cross-links | Phthalocyanine derivatives | [132] |
| VZV | Viral envelope (prevention of viral adsorption and host penetration) | Phthalocyanine derivatives | [131] |
| HIV | Major capsid protein p24 | Hypericin | [111] |
| HIV-1 | Loss of infectivity; loss of fusion function; membrane proteins cross-links | Hypericin | [30] |
| Loss of infectivity; loss of fusion function; membrane proteins cross-links | Rose bengal | [30] | |
| p24 and gp120 proteins; protein cross-links | MB | [34] | |
| Inhibition of cell fusion activity of Env proteins | Natural and sulfonated tetraarylporphyrins | [36] | |
| VSV | Loss of infectivity; loss of fusion function; cross-linking of G and M proteins | Hypericin | [30] |
| Loss of infectivity; loss of fusion function; cross-linking of G and M proteins | Rose bengal | [30] | |
| Inhibition of fusion of the envelope to Vero cells; G protein | MB | [81] | |
| Inhibition of fusion of the envelope to Vero cells; G protein | Aluminum phthalocyanine tetrasulfonate | [81] | |
| G and M proteins; protein cross-links | Phthalocyanine derivatives | [82] | |
| G, M, L and N proteins; protein cross-links | Chlorophyll derivatives | [77] | |
| Influenza virus | Loss of infectivity; loss of fusion function; cross-linking of G and M proteins | Hypericin | [30] |
| Loss of infectivity; loss of fusion function; cross-linking of G and M proteins | Rose bengal | [30] | |
| Loss of infectivity; HA fusion protein; protein cross-links | Rose bengal | [108] | |
| Sendai virus | Loss of infectivity; loss of fusion function; cross-linking of G and M proteins | Hypericin | [30] |
| Loss of infectivity; loss of fusion function; cross-linking of G and M proteins | Rose bengal | [30] | |
| Vaccinia virus | Histidine residues in virus proteins | Rose bengal | [86] |
| Human cytomegalovirus | Viral envelope (reduced ability to adhere to and penetrate host cells) | Merocyanine 540 | [45] |
| Sindbis virus | Viral envelope (reduced ability to adhere to and penetrate host cells) | Merocyanine 540 | [47] |
| Viral capsid protein | Hypericin | [133] | |
| Friend erythroleukemia virus | Viral envelope (reduced ability to adhere to and penetrate host cells) | Merocyanine 540 | [134] |
| Non-enveloped mammalian viruses | |||
| Adenovirus | Not damaged | Phthalocyanine derivatives | [131] |
| Enterovirus 71 | Appearance/disappearance of protein bands; increase of the protein band intensity | Methylene blue | [85] |
| T7 phage | Protein capsid; loosening of the protein-DNA interaction | Glycoconjugated meso-tetraarylporphyrins | [64] |
| Capsid and core proteins; loosening of protein-DNA interaction | Glycoconjugated meso-tetraarylporphyrins | [87] | |
| Capsid proteins; protein cross-links | meso-Tetrakis(1-methylpyridinium-4-yl)porphyrin | [88] | |
| Capsid proteins; protein cross-links | Polyhydroxylated fullerene | [57] | |
| M13 phage | Coat protein | Methylene blueAluminum phthalocyanine tetrasulfunate | [81] |
| PRD1 phage | Capsid proteins; protein cross-links; phospholipids (less affected) | Polyhydroxylated fullerene | [57] |
| Qβ phage | Coat and maturation (A) proteins; formation of protein carbonyls; RNA-protein cross-links | Methylene blue | [89] |
| RNA-protein cross-links | Methylene blue | [92] | |
| MS2 phage | A protein | Polyhydroxylated fullerene | [57] |
4.2.1. Damage on Mammalian Viral Outer Structures
4.2.2. Damages on Bacteriophage Outer Structures
5. Resistance to PDI and Recovery of Viability
5.1. Resistance of Mammalian Viruses and Recovery of Viability after Photosensitization
5.2. Bacteriophage Resistance and Viability Recovery after Photosensitization
6. Factors Affecting Viral PDI
6.1. Effect of the Number of Charges, Symmetry, Size of Meso Substituent Groups and Photosensitizer Concentration
6.1.1. Mammalian Viruses PDI
6.1.2. Bacteriophage PDI
6.2. Effect of Different Light Sources and Fluence Rate on Antimicrobial PDT
6.2.1. Effect of Light on Mammalian Viruses PDI
6.2.2. Effect of Light on Bacteriophage PDI
7. Conclusion
Acknowledgments
Conflict of Interest
References and Notes
- Jori, G.; Brown, S.B. Photosensitized inactivation of microorganisms. Photochem. Photobio. Sci. 2004, 3, 403–405. [Google Scholar] [CrossRef]
- van Der Poel, W.H.; Vinjé, J.; van Der Heide, R.; Herrera, M.I.; Vivo, A.; Koopmans, M.P. Norwalk-like calicivirus genes in farm animals. Emerg. Infect. Dis. 2000, 6, 36–41. [Google Scholar]
- Blerkom, L.V.L. Role of viruses in human evolution. Yearbk. Phys. Anthropol. 2009, 46, 14–46. [Google Scholar]
- Pulitzer, M.P.; Amin, B.D.; Busam, K.J. Merkel cell carcinoma: Review. Adv. Anat. Pathol. 2009, 16, 135–44. [Google Scholar] [CrossRef]
- Sullivan, V.; Biron, K.K.; Talarico, C.; Stanat, S.C.; Davis, M.; Pozzi, L.M.; Coen, D.M. A point mutation in the human cytomegalovirus DNA polymerase gene confers resistance to ganciclovir and phosphonylmethoxyalkyl derivatives. Antimicrob. Agents Ch. 1993, 37, 19–25. [Google Scholar]
- Smee, D.F.; Barnett, B.B.; Sidwell, R.W.; Reist, E.J.; Holy, A. Antiviral activities of nucleosides and nucleotides against wild-type and drug-resistant strains of murine cytomegalovirus. Antivir. Res. 1995, 26, 1–9. [Google Scholar] [CrossRef]
- Kimberlin, D.W.; Whitley, R.J. Antiviral resistance: Mechanisms, clinical significance, and future implications. J. Antimicrob. Chemother. 1996, 37, 403–421. [Google Scholar] [CrossRef]
- Jabs, D.A.; Enger, C.; Forman, M.; Dunn, J.P. for The cytomegalovirus retinitis and viral resistance study group. Incidence of foscarnet resistance and cidofovir resistance in patients treated for cytomegalovirus retinitis. Antimicrob. Agents Chemother. 1998, 42, 2240–2244. [Google Scholar]
- Pillay, D.; Zambon, M. Antiviral drug resistance. Br. Med. J. 1998, 317, 660–662. [Google Scholar] [CrossRef]
- Smee, D.F.; Sidwell, R.W.; Kefauver, D.; Bray, M.; Huggins, J.W. Characterization of wild-type and cidofovir-resistant strains of camelpox, cowpox, monkeypox, and vaccinia viruses. Antimicrob. Agents Ch. 2002, 46, 1329–1335. [Google Scholar] [CrossRef]
- Schultz, E.W.; Krueger, A.P. Inactivation of Staphylococcus bacteriophage by methylene blue. P. Soc. Exp. Biol. Med. 1928, 26, 100–101. [Google Scholar]
- Perdrau, J.R.; Todd, C. The photodynamic action of methylene blue on certain viruses. Proc. Roy. Soc. Lond. B Biol. Sci. 1933, 112, 288–298. [Google Scholar] [CrossRef]
- Käsermann, F.; Kempf, C. Buckminsterfullerene and photodynamic inactivation of viruses. Rev. Med. Virol. 1998, 8, 143–151. [Google Scholar] [CrossRef]
- Hamblin, M.R.; Hasan, T. Photodynamic therapy: A new antimicrobial approach to infectious disease? Photoch. Photobio. Sci. 2004, 5, 436–450. [Google Scholar] [CrossRef]
- Felber, T.D.; Smith, E.B.; Knox, J.M.; Wallis, C.; Melnick, J.L. Photodynamic inactivation of herpes simplex: Report of a clinical trial. J. Am. Med. Assoc. 1973, 92, 223–289. [Google Scholar]
- Almeida, A.; Cunha, A.; Faustino, M.A.F.; Tomé, A.C.; Neves, M.G.P.M.S. Porphyrins as antimicrobial photosensitizing agents. In Photodynamic Inactivation of Microbial Pathogens: Medical and Environmental Applications; Royal Society of Chemistry: Cambridge, UK, 2011; pp. 83–160. [Google Scholar]
- Mullooly, V.M.; Abramson, A.L.; Shikowitz, M.J. Dihemato-porphyrin ether-induced photosensitivity in laryngeal papilloma patients. Laser. Surg. Med. 1990, 10, 349–356. [Google Scholar] [CrossRef]
- Karrer, S.; Szeimies, R.M.; Abels, C.; Wlotzke, U.; Stolz, W.; Landthaler, M. Epidermodysplasia verruciformis treated using topical 5-aminolaevulinic acid photodynamic therapy. Br. J. Dermatol. 1999, 140, 935–938. [Google Scholar] [CrossRef]
- Lavie, G.; Mazur, Y.; Lavie, D.; Meruelo, D. The chemical and biological properties of hypericin—A compound with a broad spectrum of biological activities. Med. Res. Rev. 1995, 15, 111–119. [Google Scholar] [CrossRef]
- Smetana, Z.; Malik, Z.; Orenstein, A.; Mendelson, E.; Ben-Hur, E. Treatment of viral infections with 5-aminolevulinic acid and light. Laser. Surg. Med. 1997, 21, 351–358. [Google Scholar] [CrossRef]
- Sloand, E.M.; Pitt, E.; Klein, H.G. Safety of the blood supply. J. Am. Med. Assoc. 1995, 274, 1368–1373. [Google Scholar]
- Mannucci, P.M. Outbreak of hepatitis A among Italian patients with haemophilia. Lancet 1992, 339, 819. [Google Scholar] [CrossRef]
- Klein, H.G. Oxygen carriers and transfusion medicine. Artif. Cell. Blood Substit. Biotechnol. 1994, 22, 123–135. [Google Scholar] [CrossRef]
- Azzi, A.; Fanci, R.; Ciappi, S.; Zakrzewska, K.; Bosi, A. Human parvovirus B19 infection in bone marrow transplantation patients. Am. J. Hematol. 1993, 44, 207–209. [Google Scholar] [CrossRef]
- Asanaka, M.; Kurimura, T.; Toya, H.; Ogaki, J. Anti-HIV activity of protoporphyrin. AIDS 1989, 3, 403–404. [Google Scholar] [CrossRef]
- Dixon, D.W.; Marzilli, L.G.; Schinazi, R.F. Porphyrins as agents against the human immunodeficiency virus. Ann. N. Y. Acad. Sci. 1990, 616, 511–513. [Google Scholar] [CrossRef]
- Lambrecht, B.; Mohr, H.; Knuver-Hopf, J.; Schmitt, H. Photoinactivation of viruses in human fresh plasma by phenothiazine dyes in combination with visible light. Vox Sang. 1991, 60, 207–213. [Google Scholar] [CrossRef]
- Levere, R.D.; Gong, Y.F.; Kappas, A.; Bucher, D.J.; Wormser, G.; Abraham, N.G. Heme inhibits human immunodeficiency virus 1 replication in cell cultures and enhances the antiviral effect of zidovudine. Proc. Natl. Acad. Sci. U. S. A. 1991, 88, 1756–1759. [Google Scholar]
- Matthews, J.L.; Sogandares-Bernal, F.; Judy, M.; Gulliya, K.; Newman, J.; Chanh, T.; Marengo-Rowe, A.J. Inactivation of viruses with photoactive compounds. Blood Cell. 1992, 18, 75–88. [Google Scholar]
- Lenard, J.; Rabson, A.; Vanderoef, R. Photodynamic inactivation of infectivity of humam immunodeficiency virus and other enveloped viruses using hypericin and rose bengal: Inhibition of fusion and syncytia formation. Proc. Natl. Acad. Sci. U. S. A. 1993, 90, 158–162. [Google Scholar]
- Neurath, A.R.; Strick, N.; Jiang, S. Rapid prescreening for antiviral agents against HIV-1 based on their inhibitory activity in site-directed immunoassays—Approaches applicable to epidemic HIV-1 strains. Antivir. Chem. Chemother. 1993, 4, 207–214. [Google Scholar]
- Debnath, A.K.; Jiang, S.; Strick, N.; Lin, K.; Haberfield, P.; Neurath, A.R. 3-Dimensional structure-activity analysis of a series of porphyrin derivatives withanti-HIV-1 activity targeted to the v3 loop of the gp120 envelope glycoprotein of the human-immunodeficiency-virus type 1. J. Med. Chem. 1994, 37, 1099–1108. [Google Scholar] [CrossRef]
- North, J.; Coombs, R.; Levy, J. Photodynamic inactivation of free and cell-associated HIV-1 using the photosensitizer, benzoporphyrin derivative. J. Acquir. Immune Defic. Syndr. 1994, 7, 891–898. [Google Scholar]
- Bachmann, B.K-H.J.B.; Lambrecht, B.; Mohr, H. Target structures for HIV-1 inactivation by methylene blue and light. J. Med. Virol. 1995, 47, 172–178. [Google Scholar] [CrossRef]
- Song, R.; Witvrouw, M.; Schols, D.; Robert, A.; Balzarini, J.; De Clercq, E.; Bernadou, J.; Meunier, B. Anti-HIV activities of anionic metalloporphyrins and related compounds. Antivir. Chem. Chemother. 1997, 8, 85–97. [Google Scholar]
- Vzorov, A.N.; Dixon, D.W.; Trommel, J.S.; Marzilli, L.G.; Compans, R.W. Inactivation of human immunodeficiency virus type 1 by porphyrins. Antimicrob. Agents Ch. 2002, 46, 3917–3925. [Google Scholar] [CrossRef]
- Vanyur, R.; Heberger, K.; Jakus, J. Prediction of anti-HIV-1 activity of a series of tetrapyrrole molecules. J. Chem. Inform. Comput. Sci. 2003, 43, 1829–1836. [Google Scholar] [CrossRef]
- Dairou, J.; Vever-Bizet, C.; D. Brault, D. Interaction of sulfonated anionic porphyrins with HIV glycoprotein gp120: photodamages revealed by inhibition of antibody binding to V3 and C5 domains. Antivir. Res. 2004, 61, 37–47. [Google Scholar]
- Marchesan, S.; Da Ros, T.; Spalluto, G.; Balzarini, J.; Prato, M. Anti-HIV properties of cationic fullerene derivatives. Bioorg. Med. Chem. Lett. 2005, 15, 3615–3618. [Google Scholar]
- North, J.; Freeman, S.; Overbaugh, J.; Levy, J.; Lansman, R. Photodynamic inactivation of retrovirus by benzoporphyrin derivative: A feline leukemia virus model. Transfusion 1992, 32, 121–128. [Google Scholar]
- Müller-Breitkreutz, K.; Mohr, H. Hepatitis C and human immunodeficiency virus RNA degradation by methylene blue/light treatment of human plasma. J. Med. Virol. 1998, 56, 239–245. [Google Scholar] [CrossRef]
- Cheng, Y.; Tsou, L.K.; Cai, J.; Aya, T.; Dutschman, G.E.; Gullen, E.A.; Grill, S.P.; Chen, A.P-C.; Lindenbach, B.D.; Hamilton, A.D.; Cheng, Y-C. A novel class of meso-tetrakis-porphyrin derivatives exhibits potent activities against hepatitis C virus genotype 1b replicons in vitro. Antimicrob. Agents Ch. 2010, 54, 197–206. [Google Scholar]
- Lin, L.; Hu, J. Inhibition of hepadnavirus reverse transcriptase RNA interaction by porphyrin compounds. J. Virol. 2008, 82, 2305–2312. [Google Scholar] [CrossRef]
- Casteel, B.M.J.; Jayaraj, K.; Avram, G.; Bail, L.M.; Sobsey, M.D. Photoinactivation of hepatitis A virus by synthetic porphyrins. Photochem. Photobiol. 2004, 80, 294–300. [Google Scholar] [CrossRef]
- O’Brien, J.M.; Gaffney, D.K.; Wang, T.P.; Sieber, F. Merocyanine 540 sensitized photoinactivation of enveloped viruses in blood products: Site and mechanism of phototoxicity. Blood 1992, 80, 277–285. [Google Scholar]
- Mohr, H.; Bachmann, B.; Klein-Struckmeier, A.; Lambrecht, B. Virus inactivation of blood products by phenothiazine dyes and light. Photochem. Photobiol. 1997, 65, 441–445. [Google Scholar] [CrossRef]
- Sieber, F.; O’Brien, J.M.; Krueger, G.J.; Schober, S.L.; Burns, W.H.; Sharkis, S.J.; Sensenbrenner, L.L. Antiviral activity of merocyanine 540. Photochem. Photobiol. 1987, 46, 707–711. [Google Scholar] [CrossRef]
- Leclerc, H.; Edberg, S.; Pierzo, V.; Delattre, J.M. Bacteriophages as indicators of enteric viruses and public health risk in groundwaters. A review. J. Appl. Microbiol. 2000, 88, 5–21. [Google Scholar]
- Rywkin, S.; Ben-Hur, E.; Malik, Z.; Prince, A.M.; Li, Y.S.; Kenney, M.E.; Oleinick, N.L.; Horowitz, B. New phthalocynanines for photodynamic virus inactivation in red blood cell concentrates. Photochem. Photobiol. 1994, 60, 165–170. [Google Scholar] [CrossRef]
- Käsermann, F.; Kempf, C. Photodynamic inactivation of enveloped viruses by buckminsterfullerene. Antivir. Res. 1997, 34, 65–70. [Google Scholar]
- DiMascio, P.; Wefers, H.; Do-Thi, H-P.; Lafleur, M.V.M.; Sies, H. Singlet molecular oxygen causes loss of biological activity in plasmid and bacteriophage DNA and induces single strand breaks. Biochim. Biophys. Acta 1989, 1007, 151–157. [Google Scholar]
- Abe, H.; Ikebuchi, K.; Wagner, S.J.; Kuwabara, M.; Kamo, N.; Sekiguchi, S. Potential involvement of both type I and type II mechanisms in M13 virus inactivation by methylene blue photosensitization. Photochem. Photobiol. 1997, 66, 204–208. [Google Scholar] [CrossRef]
- Specht, K.G. The role of DNA damage in PM2 viral inactivation by methylene blue photosensitization. Photochem. Photobiol. 1994, 59, 506–514. [Google Scholar] [CrossRef]
- Schneider, J.E.; Philips, J.R.; Pye, Q.; Maidt, M.L.; Price, S.; Floyd, R.A. Methylene blue and rose bengal photoinactivation of RNA bacteriophages: Comparative studies of 8-oxoguanine formation in isolated RNA. Arch. Biochem. Biophys. 1993, 301, 91–97. [Google Scholar] [CrossRef]
- Jockush, S.; Lee, D.; Turro, N.J.; Leonard, E.F. Photoinduced inactivation of viruses: Adsorption of methylene blue, thionine and thiopyronine on Qβ bacteriophage. Proc. Natl. Acad. Sci. U. S. A. 1996, 93, 7446–7451. [Google Scholar]
- Lee, D.; Foux, M.; Leonard, E.F. The effects of methylene blue and oxygen concentration on the photoinactivation of Qβ bacteriophage. Photochem. Photobiol. 1997, 65, 161–165. [Google Scholar] [CrossRef]
- Hotze, E.M.; Badireddy, A.R.; Chellam, S.; Wiesner, M.R. Mechanisms of bacteriophage inactivation via singlet oxygen generation in UV illuminated fullerol suspensions. Environ. Sci. Tech. 2009, 43, 6639–6645. [Google Scholar] [CrossRef]
- Kasturi, C.; Platz, M.S. Inactivation of lambda phage with 658 nm light using a DNA binding porphyrin sensitizer. Photochem. Photobiol. 1992, 56, 427–429. [Google Scholar] [CrossRef]
- Martin, C.B.; Wilfong, E.; Ruane, P.; Goodrich, R.; Platz, M. An action spectrum of the riboflavin-photosensitized inactivation of lambda phage. Photochem. Photobiol. 2005, 81, 474–480. [Google Scholar] [CrossRef]
- Wagner, S.J.; Skripchenkol, A.; Robinenel, D.; Foley, J.W.; Cincotta, L. Factors affecting virus photoinactivation by a series of phenothiazine dyes. Photochem. Photobiol. 1998, 67, 343–349. [Google Scholar] [CrossRef]
- Brendel, M. Different photodynamic action of proflavine and methylene blue on bacteriophage. I. Host cell reactivation of Serratiaphage kappa. Mol. Gen. Genet. 1970, 108, 303–311. [Google Scholar] [CrossRef]
- Yamamoto, N. Photodynamic inactivation of bacteriophage and its inhibition. J. Bacteriol. 1957, 6, 510–521. [Google Scholar]
- Witmer, H.; Fraser, D. Photodynamic action of proflavine on coliphage T3 II. Protection by L-cysteine. J. Virol. 1971, 7, 319–322. [Google Scholar]
- Gábor, F.; Szolnoki, J.; Tóth, K.; Fekete, A.; Maillard, P.; Csík, G. Photoinduced inactivation of T7 phage sensitized by symmetrically and asymmetrically substituted tetraphenyl porphyrin: comparison of efficiency and mechanism of action. Photochem. Photobiol. 2001, 73, 304–311. [Google Scholar] [CrossRef]
- Kadish, L.L.; Fisher, D.B.; Pardee, A.B. Photodynamic inactivation of free and vegetative bacteriophage T4. Biochim. Biophys. Acta 1967, 138, 57–65. [Google Scholar]
- Costa, L.; Alves, E.; Carvalho, C.M.B.; Tomé, J.P.C.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, A.; Almeida, A. Sewage bacteriophage photoinactivation by cationic porphyrins: a study of charge effect. Photochem. Photobio. Sci. 2008, 7, 415–422. [Google Scholar]
- Costa, L.; Carvalho, C.M.B.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Tomé, J.P.C.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, Â.; Almeida, A. Sewage bacteriophage inactivation by cationic porphyrins: influence of light parameters. Photochem. Photobio. Sci. 2010, 9, 1126–1133. [Google Scholar]
- Costa, L.; Tomé, J.P.C.; Neves, M.G.P.M.S.; Tomé, A.C.; Cavaleiro, J.A.S.; Faustino, M.A.F.; Cunha, Â.; Gomes, N.C.M.; Almeida, A. Evaluation of resistance development and viability recovery by a non-enveloped virus after repeated cycles of aPDT. Antivir. Res. 2011, 91, 278–282. [Google Scholar] [CrossRef]
- DeRosa, M.C.; Crutchley, R.J. Photosensitized singlet oxygen and its applications. Coord. Chem. Rev. 2002, 233–234, 351–371. [Google Scholar]
- Capella, M.A.M.; Capella, L.S. A light in multidrug resistance: Photodynamic treatment of multidrug-resistant tumors. J. Biomed. Sci. 2003, 10, 361–366. [Google Scholar] [CrossRef]
- Castano, A.P.; Demidova, T.N.; Hamblin, M.R. Mechanisms in photodynamic therapy: Part one-photosensitizers, photochemistry and cellular localization. Photodiagn. Photodyn. 2004, 1, 279–293. [Google Scholar] [CrossRef]
- Prates, R.A.; da Silva, E.G.; Yomada, A.M., Jr.; Suzuki, L.C.; Paula, C.R.; Ribeiro, M.S. Light parameters influence cell viability in antifungal photodynamic therapy in a fluence and rate fluence dependent manner. Laser Phys. 2009, 19, 1038–1044. [Google Scholar]
- Welsh, J.N.; Adams, M.H. Photodynamic inactivation of bacteriophage. J. Bacteriol. 1954, 1, 122–127. [Google Scholar]
- Huang, Q.; Fu, W-L.; Chen, B.; Huang, J-F.; Zhang, X.; Xue, Q. Inactivation of dengue virus by methylene blue/narrow bandwidth light system. J. Photochem. Photobiol. B Biol. 2004, 77, 39–43. [Google Scholar]
- Schnipper, L.E.; Lewin, A.A.; Swartz, M.; Crumpacker, C.S. Mechanisms of photodynamic inactivation of herpes simplex viruses; comparison between methylene blue, light plus electricity, and hematoporphyrin plus light. J. Clin. Investig. 1980, 65, 432–438. [Google Scholar] [CrossRef]
- Schagen, F.H.E.; Moor, A.C.E.; Cheong, S.C.; Cramer, S.J.; van Ormondt, H.; van der Eb, A.J.; Dubbelman, T.M.A.R.; Hoeben, R.C. Photodynamic treatment of adenoviral vectors with visible light: An easy and convenient method for viral inactivation. Gene Ther. 1999, 6, 873–881. [Google Scholar]
- Lim, D-S.; Ko, S-H.; Kim, S-J.; Park, Y-J.; Park, J-H.; Lee, W-Y. Photoinactivation of vesicular stomatitis virus by a photodynamic agent, chlorophyll derivatives from silkworm excreta. J. Photochem. Photobiol. B Biol. 2002, 67, 149–156. [Google Scholar] [CrossRef]
- Sagristá, M.L.; Postigo, F.; De Madariaga, M.A.; Pinto, R.M.; Caballero, S.; Bosch, A.; Vallés, M.A.; Mora, M. Photodynamic inactivation of viruses by immobilized chlorine-containing liposomes. J. Porphyrin Phthalocyanines. 2009, 13, 578–588. [Google Scholar] [CrossRef]
- Tomé, J.P.C.; Neves, M.G.P.M.S.; Tomé, A.C.; Cavaleiro, J.A.S.; Mendonça, A.F.; Pegado, I.N.; Duarte, R.; Valdeira, M.L. Synthesis of glycoporphyrin derivatives and their antiviral activity against herpes simplex virus types 1 and 2. Bioorg. Med. Chem. 2005, 13, 3878–3888. [Google Scholar]
- Silva, E.M.P.; Giuntini, F.; Faustino, M.A.F.; Tomé, J.P.C.; Neves, M.G.P.M.S.; Tomé, A.C.; Silva, A.M.S.; Santana-Marques, M.G.; Ferrer-Correia, A.J.; Cavaleiro, J.A.S.; et al. Synthesis of cationic β-vinyl substituted meso-tetraphenylporphyrins and their in vitro activity against herpes simplex virus type 1. Bioorg. Med. Chem. Lett. 2005, 15, 3333–3337. [Google Scholar]
- Abe, H.; Wagner, S.J. Analysis of viral DNA, protein and envelope damage after methylene blue, phthalocyanine derivative or merocyanine 540 photosensitization. Photochem. Photobiol. 1995, 61, 402–409. [Google Scholar] [CrossRef]
- Moor, A.C.E.; Wagenaars-van Gompel, A.E.; Brand, A.; Dubbelman, T.M.A.R.; Van Steveninck, J. Primary targets for photoinactivation of vesicular stomatitis virus by AlPcS4 or Pc4 and red light. Photochem. Photobiol. 1997, 65, 465–470. [Google Scholar] [CrossRef]
- Smetana, Z.; Ben-Hur, E.; Mendelson, E.; Salzberg, S.; Wagner, P.; Malik, Z. Herpes simplex virus proteins are damaged following photodynamic inactivation with phthalocyanines. J. Photochem. Photobiol. B Biol. 1998, 44, 77–83. [Google Scholar] [CrossRef]
- Müller-Breitkreutz, K.; Mohr, H.; Briviba, K.; Sies, H. Inactivation of viruses by chemically and photochemically generated singlet molecular oxygen. J. Photochem. Photobiol. B Biol. 1995, 30, 63–70. [Google Scholar] [CrossRef]
- Wong, T-W.; Huang, H-J.; Wang, Y-F. ; Lee, Y-P. ; Huang, C-C. ; Yu, C-K. Methylene blue-mediated photodynamic inactivation as a novel disinfectant of enterovirus 71. J. Antimicrob. Chemother. 2010, 65, 2176–2182. [Google Scholar] [CrossRef]
- Turner, G.S.; Kaplan, C. Photoinactivation of vaccinia virus with rose bengal. J. Gen. Virol. 1968, 3, 433–443. [Google Scholar] [CrossRef]
- Egyeki, M.; Turóczy, G.; Majer, Zs.; Tóth, K.; Fekete, A.; Maillard, Ph.; Csík, G. Photosensitized inactivation of T7 phage as surrogate of non-enveloped DNA viruses: Efficiency and mechanism of action. Biochim. Biophys. Acta 2003, 1624, 115–124. [Google Scholar] [CrossRef]
- Zupán, K.; Egyeki, M.; Tóth, K.; Fekete, A.; Herényi, L.; Módos, K.; Csík, G. Comparison of the efficiency and the specificity of DNA-bound and free cationic porphyrin in photodynamic virus inactivation. J. Photochem. Photobiol. B Biol. 2008, 90, 105–112. [Google Scholar] [CrossRef]
- Schneider, J.E., Jr.; Tabatabale, T.; Maidt, L.; Smith, R.H.; Nguyen, X.; Pye, Q.; Floyd, R.A. Potential mechanisms of photodynamic inactivation of virus by methylene blue I. RNA-protein crosslinks and other oxidative lesions in Qβ bacteriophage. Photochem. Photobiol. 1998, 67, 350–357. [Google Scholar] [CrossRef]
- Badireddy, A.R.; Hotze, E.M.; Chellam, S.; Alvarez, P.J.J.; Wiesner, M.R. Inactivation of bacteriophages via photosensitization of fullerol nanoparticles. Environ. Sci. Tech. 2007, 41, 6627–6632. [Google Scholar] [CrossRef]
- Marotti, J.; Aranha, A.C.C.; Eduardo, C.D.P.; Ribeiro, M.S. Photodynamic therapy can be effective as a treatment for herpes simplex labialis. Photomed. Laser Surg. 2009, 27, 357–363. [Google Scholar] [CrossRef]
- Floyd, R.A.; Schneider, J.E.; Dittmer, D.P. Methylene blue photoinactivation of RNA viruses. Antivir. Res. 2004, 61, 141–151. [Google Scholar]
- Wallis, C.; Melnick, J.L. Photodynamic inactivation of animal viruses: A review. Photochem. Photobiol. 1965, 4, 159–170. [Google Scholar] [CrossRef]
- Lytle, C.D.; Budacz, A.P.; Keville, E.; Miller, S.A.; Prodouz, K.N. Differential inactivation of surrogate viruses with merocyanine 540. Photochem. Photobiol. 1991, 54, 489–493. [Google Scholar] [CrossRef]
- Via, L.D.; Magno, S.M. Photochemotherapy in the treatment of cancer. Curr. Med. Chem. 2001, 8, 1405–1418. [Google Scholar]
- Schmidt-Erfurth, U.; Hasan, T. Mechanisms of action of photodynamic therapy with verteporfin for the treatment of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 195–214. [Google Scholar] [CrossRef]
- Wainwright, M. Photodynamic antimicrobial chemotherapy (PACT). J. Antimicrob. Chemother. 1998, 42, 13–28. [Google Scholar] [CrossRef]
- Bonnett, R. Chemical Aspects of Photodynamic Therapy; Gordon and Breach Science Publishers: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Girotti, A.W. Photosensitized oxidation of membrane lipids: Reaction pathways, cytotoxic effects, and cytoprotective mechanisms. J. Photochem. Photobiol. B Biol. 2001, 63, 103–113. [Google Scholar] [CrossRef]
- Calin, M.A.; Parasca, S.V. Light sources for photodynamic inactivation of bacteria. Laser Med. Sci. 2009, 24, 453–460. [Google Scholar] [CrossRef]
- Min, D.B.; Boff, J.M. Chemistry and reaction of singlet oxygen in foods. Compr. Rev. Food Sci. Food Saf. 2002, 1, 58–72. [Google Scholar] [CrossRef]
- Maisch, T.; Bosl, C.; Szeimies, R.M.; Lehn, N.; Abels, C. Photodynamic effects of novel XF porphyrin derivativeson prokaryotic and eukaryotic cells. Antimicrob. Agents Ch. 2005, 49, 1542–1552. [Google Scholar]
- Ochsner, M. Photophysical and photobiological processes in the photodynamic therapy of tumours. J. Photochem. Photobiol. B Biol. 1997, 39, 1–18. [Google Scholar] [CrossRef]
- Wondrak, G.T.; Jacobson, M.K.; Jacobson, E.L. Identification of quenchers of photoexcited states as novel agents for skin photoprotection. J. Pharmacol. Exp. Therapeut. 2005, 312, 482–491. [Google Scholar]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Exp. Physiol. 1997, 82, 291–295. [Google Scholar]
- Rywkin, S.; Lenny, L.; Goldstein, J.; Geacintov, N.E.; Margolis-Nunno, H.; Horowitz, B. Importance of type I and type II mechanisms in the photodynamic inactivation of viruses in blood with aluminum phthalocyanine derivatives. Photochem. Photobiol. 1992, 56, 463–469. [Google Scholar] [CrossRef]
- Costa, L.; Tomé, J.P.C.; Faustino, M.A.F.; Neves, M.G.P.S.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, A.; Almeida, A. Involvement of type I and type II mechanisms on the photoinactivation of non-enveloped DNA and RNA bacteriophages. Environ. Sci. Tech. 2012. submitted for publication. [Google Scholar]
- Lenard, J.; Vanderoef, R. Photoinactivation of influenza virus fusion and infectivity by rose bengal. Photochem. Photobiol. 1993, 58, 527–531. [Google Scholar] [CrossRef]
- Bisby, R.H.; Morgan, C.G.; Hamblett, I.; Gorman, A.A. 1999. Quenching of singlet oxygen by trolox c, ascorbate, and amino acids: effects on pH and temperature. J. Phys. Chem. A 1999, 103, 7454–7459. [Google Scholar]
- Baker, A.; Kanofsky, J.R. Quenching of singlet oxygen bybiomolecules from Ll210 leukemia cells. Photochem. Photobiol. 1992, 55, 523–528. [Google Scholar] [CrossRef]
- Degar, S.; Prince, A.M.; Pascual, D.; Lavie, G.; Levin, B.; Mazur, Y.; Lavie, D.; Ehrlich, L.S.; Carter, C.; Meruelo, D. Inactivation of the human immunodeficiency virus by hypericin: Evidence for photochemical alterations of p24 and a block in uncoating. AIDS Res. Hum. Retrovir. 1992, 8, 1929–1936. [Google Scholar] [CrossRef]
- Wainwright, M. Local treatment of viral disease using photodynamic therapy. Int. J. Antimicrob. Agents 2003, 21, 510–520. [Google Scholar] [CrossRef]
- Garcia, G.; Sarrazy, V.; Sol, V.; Morvan, C.L.; Granet, R.; Alves, S.; Krausz, P. DNA photocleavage by porphyrin–polyamine conjugates. Bioorg. Med. Chem. 2009, 17, 767–776. [Google Scholar]
- Miranda, M.A. Photosensitization by drugs. Pure Appl. Chem. 2001, 73, 481–486. [Google Scholar] [CrossRef]
- Wainwright, M. The use of methylene blue derivatives in blood product disinfection. Int. J. Antimicrob. Agents 2000, 16, 381–394. [Google Scholar] [CrossRef]
- McBride, T.J.; Schneider, J.E.; Floyd, R.E.; Loeb, L.A. Mutations induced by methylene blue plus light in single stranded M13mp2. Proc. Natl. Acad. Sci. U. S. A. 1992, 89, 6866–6870. [Google Scholar]
- OhUigin, C.; McConnell, D.J.; Kelly, J.M.; van der Putten, W.J.M. Methylene blue photosensitised strand cleavage of DNA: Effects of dye binding and oxygen. Nucleic Acids Res. 1987, 15, 7411–7427. [Google Scholar] [CrossRef]
- Mettath, S.; Munson, B.R.; Pandey, R.K. DNA interaction and photocleavage properties of porphyrins containing cationic substituents at the peripheral position. Bioconjugate Chem. 1999, 10, 94–102. [Google Scholar] [CrossRef]
- Kubát, P.; Lang, K.; Anzenbacher, P., Jr.; Jursíkova, K.; Král, V.; Ehrenberg, B. Interaction of novel cationic meso-tetraphenylporphyrins in the ground and excited states with DNA and nucleotides. J. Chem. Soc., Perkin Trans. 1 2000, 1, 933–941. [Google Scholar]
- Caminos, D.A.; Durantini, E.N. Interaction and photodynamic activity of cationic porphyrin derivativesbearing different patterns of charge distribution with GMP and DNA. J. Photochem. Photobiol. A: Chem. 2008, 198, 274–281. [Google Scholar] [CrossRef]
- Müller-Breitkreutz, K.; Mohr, H. Infection cycle of herpes viruses after photodynamic treatment with methylene blue and light. Transfusions Medizin 1997, 34, 37–42. [Google Scholar]
- Schneider, J.E., Jr.; Pye, Q.; Floyd, R.A. Qβ bacteriophage photoinactivated by methylene blue plus light involves inactivation of its genomic RNA. Photochem. Photobiol. 1999, 70, 902–909. [Google Scholar] [CrossRef]
- Smetana, Z.; Mendelson, E.; Manor, J.; Van Lier, J.E.; Ben-Hur, E.; Salzberg, S.; Malik, Z. Photodynamic inactivation of herpes simplex viruses with phthalocyanine derivatives. J. Photochem. Photobiol. B: Biol. 1994, 22, 37–43. [Google Scholar] [CrossRef]
- Ben-Hur, E.; Horowitz, B. Virus inactivation in blood. AIDS 1996, 11, 1183–1190. [Google Scholar] [CrossRef]
- Girotti, A.W. Photodynamic action of protoporphyrin IX on human erythrocytes: Cross-linking of membrane proteins. Biochem. Biophys. Res. Comm. 1976, 72, 1367–1374. [Google Scholar] [CrossRef]
- Verweij, H.; van Steveninck, J. Model studies on photodynamic cross-linking. Photochem. Photobiol. 1982, 35, 265–267. [Google Scholar] [CrossRef]
- Macdonald, I.J.; Dougherty, T.J. Basic principles of photodynamic therapy. J. Porphyrin Phthalocyanines 2001, 5, 105–129. [Google Scholar] [CrossRef]
- Davies, M.J. Singlet oxygen-mediated damage to proteins and its consequences. Biochem. Biophys. Res. Comm. 2003, 305, 761–770. [Google Scholar] [CrossRef]
- Afonso, S.G.; Enriquez, S.R.; Batlle, C.A.M. The photodynamic and non photodynamic actions of porphyrins. Braz. J. Med. Biol. Res. 1999, 32, 255–266. [Google Scholar]
- Jori, G.; Galiazzo, G.; Tamburro, A.M.; Scoffone, E. Dye-sensitized photooxidation as a tool for determining the degree of exposure of amino acid residues in proteins. J. Biol. Chem. 1970, 245, 3375–3383. [Google Scholar]
- Malik, Z.; Ladan, H.; Nitzan, Y.; Smetana, Z. Antimicrobial and antiviral activity of porphyrin photosensitization. Proc. SPIE 1993, 2078, 305–312. [Google Scholar]
- Malik, Z.; Smetana, Z.; Mendelson, E.; Wagner, P.; Salzberg, S.; Ben-Hur, E. Alteration in herpes simplex virus proteins following photodynamic treatment with phthalocyanines. Photochem. Photobiol. 1996, 63, 59S. [Google Scholar]
- Yip, L.; Hudson, J.B.; Gruszecka-Kowalik, E.; Zalkow, L.H.; Neil Towers, G.H.N. Antiviral activity of a derivative of the photosensitive compound hypericin. Phytomedicine 1996, 2, 185–190. [Google Scholar]
- Sieber, F.; Krueger, G.J.; O’Brien, J.M.; Schober, S.L.; Sensenbrenner, L.L.; Sharkis, S.J. Inactivation of Friend erythroleukemia virus and Friend virus-transformed cells by merocyanine 540-mediated photosensitization. Blood 1989, 73, 345–350. [Google Scholar]
- Melki, R.; Gaudin, Y.; Blondel, D. Interaction between tubulin and the viral matrix protein of vesicular stomatitis virus: possible implications in the viral cytopathic effect. Virology 1994, 202, 339–347. [Google Scholar] [CrossRef]
- Zupán, K.; Herényi, L.; Tóth, K.; Majer, Z.; Csík, G. Binding of cationic porphyrin to isolated and encapsidated viral DNA analyzed by comprehensive spectroscopic methods. Biochemistry 2004, 43, 9151–9159. [Google Scholar]
- Yoshikawa, T.T. Antimicrobial resistance and aging: beginning of the end of the antibiotic era? J. Am. Geriatr. Soc. 2002, 50, S226–S229. [Google Scholar] [CrossRef]
- Malik, Z.; Gozhansky, S.; Nitzan, Y. Effects of photoactivated HPD on bacteria and antibiotic resistance. Microbios Lett. 1982, 21, 103–112. [Google Scholar]
- Maisch, T.; Szeimies, R-M.; Jori, G.; Abels, C. Antibacterial photodynamic therapy in dermatology. Photochem. Photobio. Sci. 2004, 3, 907–917. [Google Scholar] [CrossRef]
- Pillay, D. Emergence and control of resistance to antiviral drugs in resistance in herpes viruses, hepatitis B virus, and HIV. Commun. Dis. Public Health 1998, 1, 5–13. [Google Scholar]
- Reddi, E.; Ceccon, M.; Valduga, G.; Jori, G.; Bommer, J.C.; Elisei, F.; Latterini, L.; Mazzucato, U. Photophysical properties and antibacterial activity of meso-substituted cationic porphyrins. Photochem. Photobiol. 2002, 75, 462–470. [Google Scholar] [CrossRef]
- Jori, G.; Coppellotti, O. Inactivation of pathogenic microorganisms by photodynamic techniques: Mechanistic aspects and perspective applications. Anti-Infect. Agents Med. Chem. 2007, 6, 119–131. [Google Scholar]
- Minnock, A.; Vernon, D.I.; Schofield, J.; Griffiths, J.; Parish, J.H.; Brown, S.B. Mechanism of uptake of a cationic water-soluble pyridinium zinc phthalocyanine across the outer membrane of Escherichia coli. Antimicrob. Agents Ch. 2000, 44, 522–527. [Google Scholar] [CrossRef]
- Demidova, T.; Hamblin, M. Effects of cell-photosensitizer binding and cell density on microbial photoinactivation. Antimicrob. Agents Ch. 2005, 6, 2329–2335. [Google Scholar] [CrossRef]
- Jori, G.; Fabris, C.; Soncin, M.; Ferro, S.; Coppellotti, O.; Dei, D.; Fantetti, L.; Chiti, G.; Roncucci, G. Photodynamic therapy in the treatment of microbial infections: Basic principles and perspective applications. Laser. Surg. Med. 2006, 38, 468–481. [Google Scholar] [CrossRef]
- Wainwright, M. Photoantimicrobials—So what’s stopping us? Photodiagn. Photodyn. 2009, 6, 167–169. [Google Scholar] [CrossRef]
- Merchat, M.; Bertolini, G.; Giacomini, P.; Villanueva, A.; Jori, G. Meso-substituted cationic porphyrins as efficient photosensitizers of Gram-positive and Gram-negative bacteria. J. Photochem. Photobiol. 1996, 32, 153–157. [Google Scholar] [CrossRef]
- Dowd, S.E.; Pillai, S.D.; Wang, S.; Corapcioglu, M.Y. Delineating the specific influence of virus isoelectric point and size on virus adsorption and transport through sandy soils. Appl. Environ. Microbiol. 1998, 64, 405–410. [Google Scholar]
- Wainwright, W. Photoantimicrobials—A PACT against resistance and infection. Drugs Future 2004, 29, 85–93. [Google Scholar] [CrossRef]
- Kessel, D.; Raymund, L.; Vicente, M.G.H. Localization and photodynamic efficacy of two cationic porphyrins varying in charge distribution. Photochem. Photobiol. 2003, 78, 431–435. [Google Scholar] [CrossRef]
- Banfi, S.; Caruso, E.; Buccafurni, L.; Battini, V.; Zazzaron, S.; Barbieri, P.; Orlandi, V. Antibacterial activity of tetraaryl-porphyrin photosensitizers: An in vivo study on Gram negative and Gram positive bacteria. J. Photochem. Photobiol. B Biol. 2006, 85, 28–38. [Google Scholar] [CrossRef]
- Konopka, K.; Goslinski, T. Photodynamic therapy in dentistry. Crit. Rev. Oral Biol. Med. 2007, 8, 694–707. [Google Scholar]
- Robertson, C.A.; Evans, D.H.; Abrahamse, H. Photodynamic therapy (PDT): A short review on cellular mechanisms and cancer research applications for PDT. J. Photochem. Photobiol. B Biol. 2009, 96, 1–8. [Google Scholar] [CrossRef]
- Brancaleon, L.; Moseley, H. Laser and non-laser light sources for photodynamic therapy. Laser Med. Sci. 2002, 17, 173–186. [Google Scholar] [CrossRef]
- Veenhuizen, R.B.; Stewart, F.A. The importance of fluence rate in photodynamic therapy: Is there a parallel with ionizing radiation dose-rate effects? Radiother. Oncol. 1995, 37, 131–135. [Google Scholar] [CrossRef]
- Allison, R.R.; Mota, H.C.; Sibata, C.H. Clinical PD/PDT in North America: an historical review. Photodiagn. Photodyn. 2004, 1, 263–277. [Google Scholar] [CrossRef]
- Juzeniene, A.; Juzena, P.; Ma, L-W.; Iani, V.; Moan, J. Effectiveness of different light sources for 5-aminolevulinic acid photodynamic therapy. Laser Med. Sci. 2004, 19, 139–149. [Google Scholar] [CrossRef]
- Kübler, A.C. Photodynamic therapy. Med. Laser Appl. 2005, 20, 37–45. [Google Scholar] [CrossRef]
- Lukšiene, Z. New approach to inactivation of harmful and pathogenic microorganisms by photosensitization. Food Tech. Biotechnol. 2005, 43, 411–418. [Google Scholar]
- Qin, Y.; Luan, X.; Bi, L.; He, G.; Bai, X.; Zhou, C.; Zhang, Z. Toluidine blue-mediated photoinactivation of periodontal pathogens from supragingival plaques. Laser Med. Sci. 2008, 23, 49–54. [Google Scholar] [CrossRef]
- Schindl, A.; Rosado-Sholosser, B.; Trautinger, F. Reciprocity regulation in photobiology: An overview (in German). Hautarzt 2001, 52, 779–785. [Google Scholar] [CrossRef]
- Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Woolsey, G.A. The role of oxygen in the visible-light inactivation of Staphylococcus aureus. J. Photochem. Photobiol. B Biol. 2008, 92, 180–184. [Google Scholar] [CrossRef]
- Langmack, K.; Mehta, R.; Twyman, P.; Norris, P. Topical photodynamic therapy at low fluence rates—theory and practice. J. Photochem. Photobiol. B Biol. 2001, 60, 37–43. [Google Scholar] [CrossRef]
- Wagner, S.J.; Storry, J.R.; Mallory, D.A.; Stromberg, R.R.; Benade, L.E.; Friedman, L.I. Red cell alterations associated with virucidal methylene blue phototreatment. Transfusion 1993, 33, 30–36. [Google Scholar]
- Wagner, S.J. Virus inactivation in blood components by photoactive phenothiazine dyes. Transfus. Med. Rev. 2002, 16, 61–66. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Costa, L.; Faustino, M.A.F.; Neves, M.G.P.M.S.; Cunha, Â.; Almeida, A. Photodynamic Inactivation of Mammalian Viruses and Bacteriophages. Viruses 2012, 4, 1034-1074. https://doi.org/10.3390/v4071034
Costa L, Faustino MAF, Neves MGPMS, Cunha Â, Almeida A. Photodynamic Inactivation of Mammalian Viruses and Bacteriophages. Viruses. 2012; 4(7):1034-1074. https://doi.org/10.3390/v4071034
Chicago/Turabian StyleCosta, Liliana, Maria Amparo F. Faustino, Maria Graça P. M. S. Neves, Ângela Cunha, and Adelaide Almeida. 2012. "Photodynamic Inactivation of Mammalian Viruses and Bacteriophages" Viruses 4, no. 7: 1034-1074. https://doi.org/10.3390/v4071034