HIV-1 Infection of T Cells and Macrophages Are Differentially Modulated by Virion-Associated Hck: A Nef-Dependent Phenomenon


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
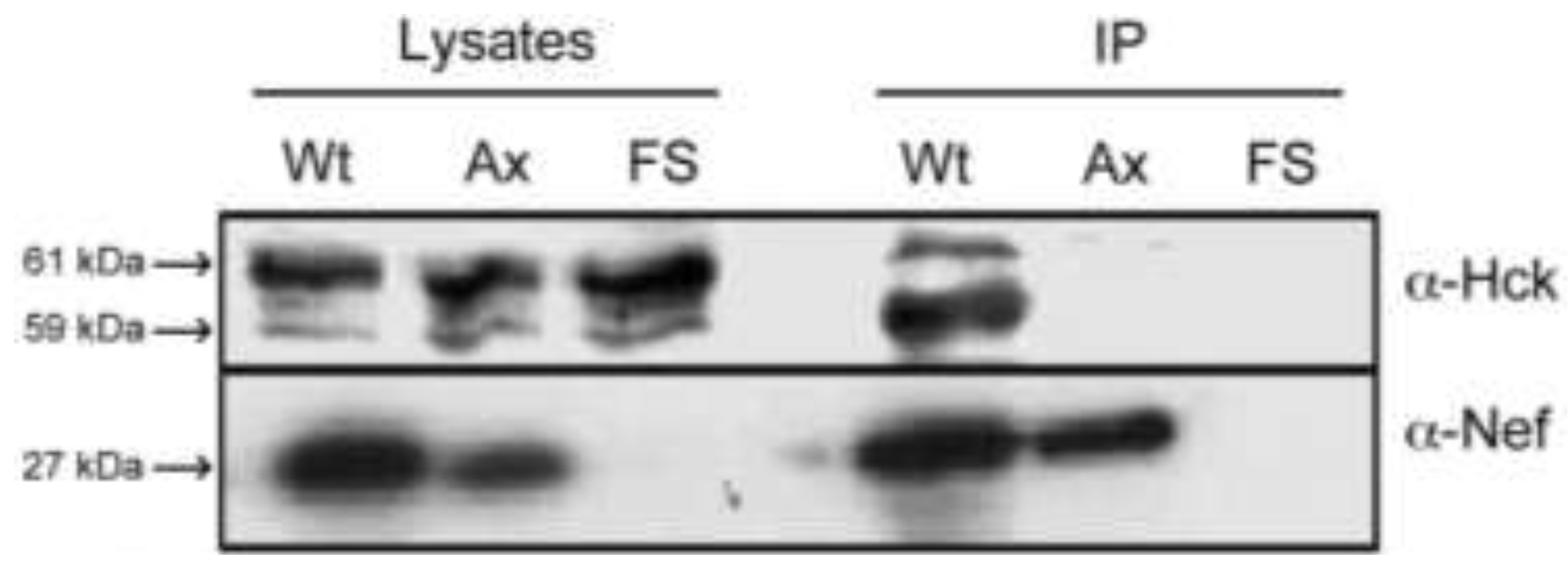
2.1. Nef Co-Precipitates Endogenous Hck from HIV-1 Infected Macrophages
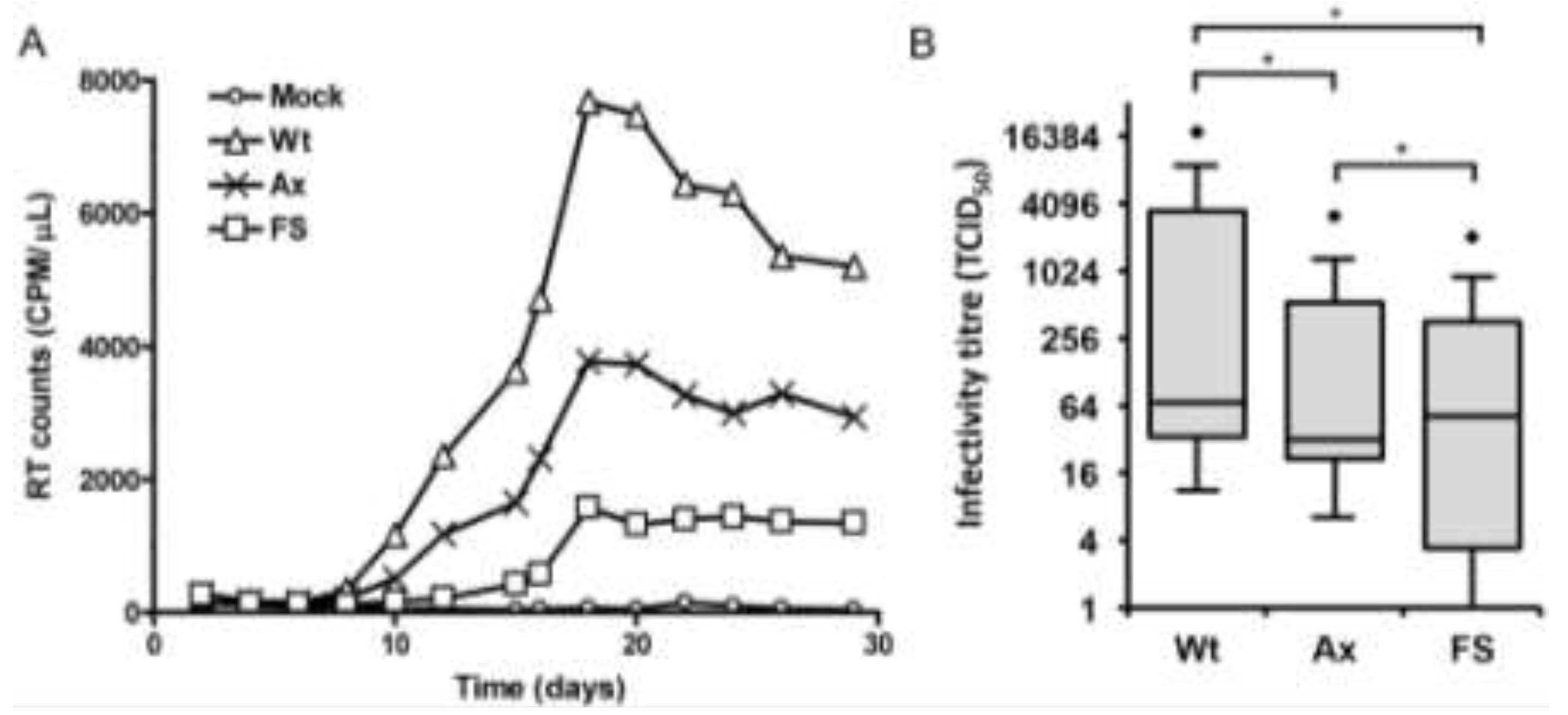
2.2. Nef and the Nef PxxP Motif Enhance HIV-1 Replication and Infectivity in Primary Monocyte-Derived Macrophages (MDMs)


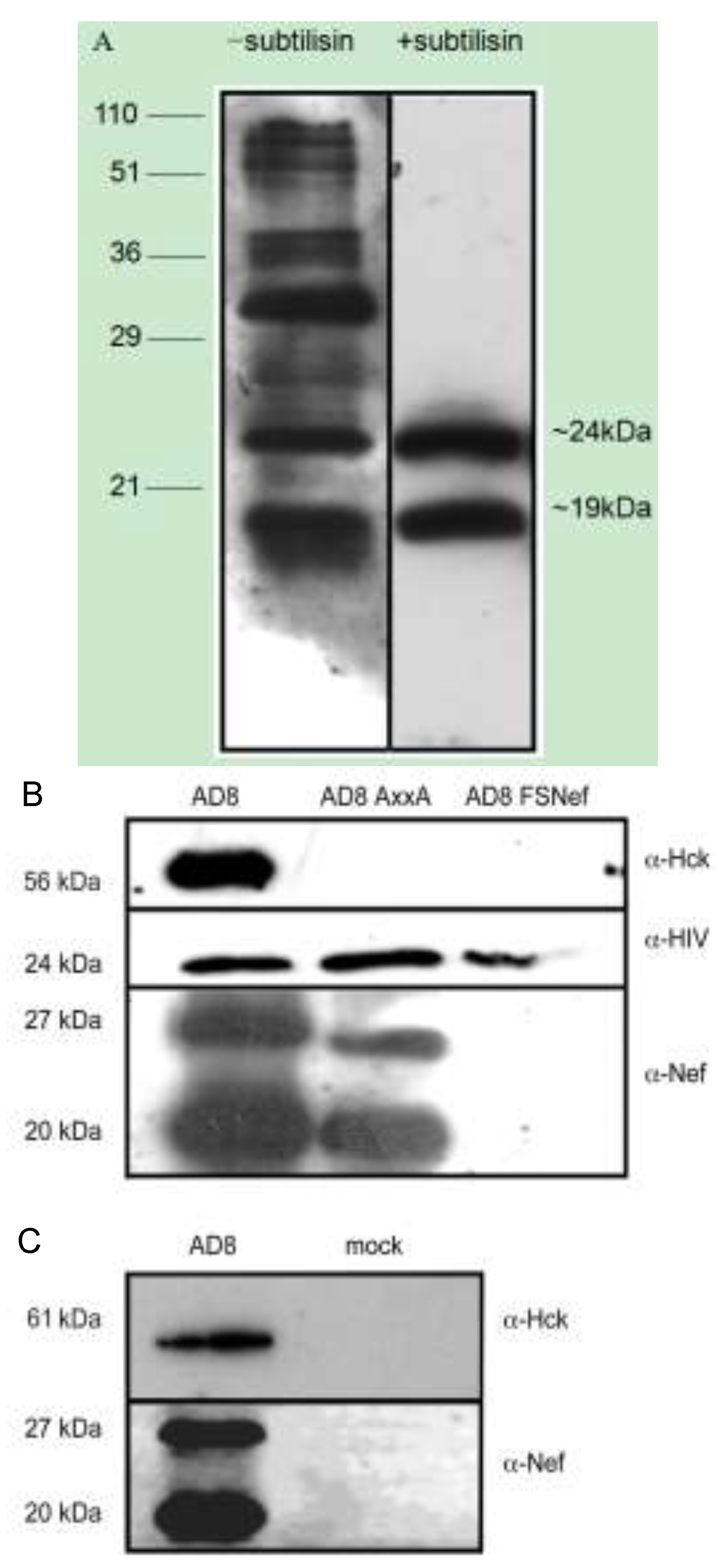
2.3. Nef and Hck Incorporation into HIV-1 AD8 Virions

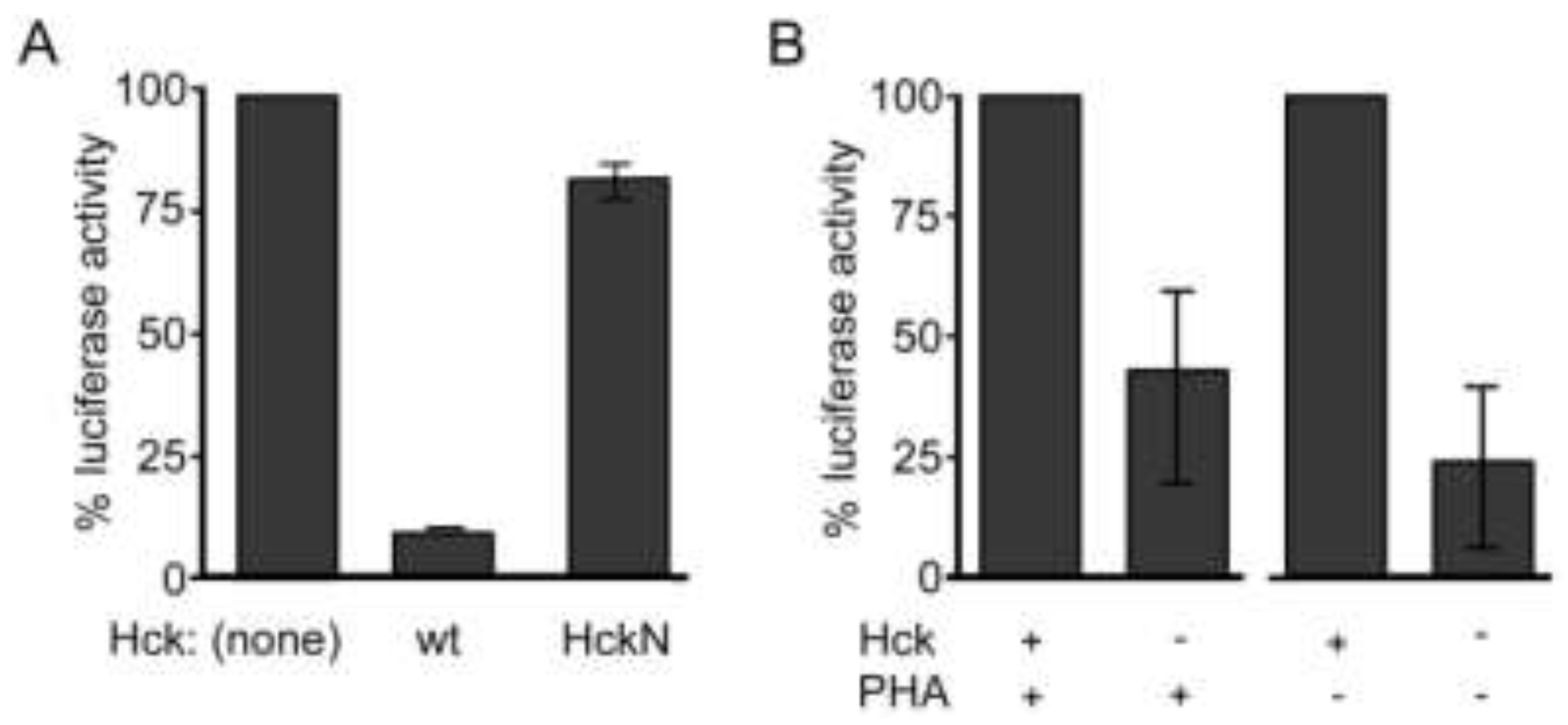
2.4. Hck Inhibits HIV-1 Infection of Monocyte-Derived Macrophages and Enhances Infection of Primary Lymphocytes

3. Experimental
3.1. HIV-1 Molecular Clones and Reporter Constructs
3.2. Antibodies and Cell Lines
3.3. Viral Stocks
3.4. Infection of T Lymphocytes and Macrophages
3.5. Infectivity Assays
3.6. Immunoprecipitation Assays
3.7. Detection of Virion-Associated Nef and Src Kinases
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Aiken, C.; Trono, D. Nef stimulates human immunodeficiency virus type 1 proviral DNA synthesis. J. Virol. 1995, 69, 5048–5056. [Google Scholar]
- Brown, A.; Moghaddam, S.; Kawano, T.; Cheng-Mayer, C. Multiple human immunodeficiency virus type 1 Nef functions contribute to efficient replication in primary human macrophages. J. Gen. Virol. 2004, 85, 1463–1469. [Google Scholar] [CrossRef]
- Carl, S.; Greenough, T.; Krumbiegel, M.; Greenberg, M.; Skowronski, J.; Sullivan, J.; Kirchhoff, F. Modulation of different human immunodeficiency virus type 1 Nef functions during progression to AIDS. J. Virol. 2001, 8, 3657–3665. [Google Scholar]
- Cheng, H.; Hoxie, J.; Parks, W. The conserved core of human immunodeficiency virus type 1 Nef is essential for association with Lck and for enhanced viral replication in T-lymphocytes. Virology 1999, 264, 5–15. [Google Scholar] [CrossRef]
- Chowers, M.; Pandori, M.; Spina, C.; Richman, D.; Guatelli, J. The growth advantage conferred by HIV-1 nef is determined at the level of viral DNA formation and is independent of CD4 downregulation. Virology 1995, 212, 451–457. [Google Scholar] [CrossRef]
- Goldsmith, M.; Wamerdam, M.; Atchison, R.; Miller, M.; Greene, W. Dissociation of the CD4 downregulation and viral infectivity enhancement functions of human immunodeficiency virus type 1 Nef. J. Virol. 1995, 69, 4112–4121. [Google Scholar]
- Kestler, H.I.; Ringler, D.; Mori, K.; Panicali, D.; Seghal, P.; Daniel, M.; Desrosiers, R. Importance of the nef gene for maintenance of high virus loads and for development of AIDS. Cell 1991, 65, 651–662. [Google Scholar]
- Kuo, L.S.; Baugh, L.L.; Denial, S.J.; Watkins, R.L.; Liu, M.; Garcia, J.V.; Foster, J.L. Overlapping effector interfaces define the multiple functions of the HIV-1 Nef polyproline helix. Retrovirology 2012, 9. [Google Scholar] [CrossRef]
- Miller, M.D.; Warmerdam, M.T.; Gaston, I.; Greene, W.C.; Feinberg, M.B. The human immunodeficiency virus-1 nef gene product: A positive factor for viral infection and replication in primary lymphocytes and macrophages. J. Exp. Med. 1994, 179, 101–113. [Google Scholar] [CrossRef]
- Miller, M.D.; Warmerdam, M.T.; Page, K.A.; Feinberg, M.B.; Greene, W.C. Expression of the human immunodeficiency virus type 1 (HIV-1) nef gene during HIV-1 production increases progeny particle infectivity independently of gp160 or viral entry. J. Virol. 1995, 69, 579–584. [Google Scholar]
- De, S.K.; Venkateshan, C.N.S.; Seth, P.; Gajdusek, D.C.; Gibbs., C.J., Jr. Adenovirus-mediated human immunodeficiency virus-1 Nef expression in human monocytes/macrophages and effect of Nef on downmodulation of Fcγ receptors and expression of monokines. Blood 1998, 91, 2108–2117. [Google Scholar]
- Garcia, J.; Miller, A. Serine phosphorylation-independent downregulation of cell-surface CD4 by Nef. Nature 1991, 350, 508–511. [Google Scholar] [CrossRef]
- Scheppler, J.A.; Nicholson, J.K.; Swan, D.C.; Ahmed-Ansari, A.; McDougal, J.S. Down-modulation of MHC-I in a CD4+ T cell line, CEM-E5, after HIV-1 infection. J. Immunol. 1989, 143, 2858–2866. [Google Scholar]
- Swigut, T.; Shohdy, N.; Skowronski, J. Mechanism for down-regulation of CD28 by Nef. EMBO J. 2001, 20, 1593–2001. [Google Scholar] [CrossRef]
- Greenway, A.; Azad, A.; McPhee, D. Human immunodeficiency virus type 1 Nef protein inhibits activation pathways in peripheral blood mononuclear cells and T-cell lines. J. Virol. 1995, 69, 1842–1850. [Google Scholar]
- Simmons, A.; Aluvihare, V.; McMichael, A. Nef triggers a transcriptional program in T cells imitating single-signal T cell activation and inducing HIV virulence mediators. Immunity 2001, 14, 763–777. [Google Scholar] [CrossRef]
- Haller, C.; Rauch, S.; Fackler, O. HIV-1 Nef employs two distinct mechanisms to modulate Lck subcellular localization and TCR induced actin remodelling. PLoS One 2007, 2, e1212. [Google Scholar] [CrossRef]
- Wonderlich, E.; Williams, M.; Collins, K. The tyrosine binding pocket in the adaptor protein 1 (AP-1) µ1 subunit is necessary for Nef to recruit AP-1 to the major histocompatibility complex class I cytoplasmic tail. J. Biol. Chem. 2008, 283, 3011–3022. [Google Scholar] [CrossRef]
- Zheng, Y.-H.; Plemenitas, A.; Fielding, C.J.; Peterlin, B.M. Nef increases the synthesis of and transports cholesterol to lipid rafts and HIV-1 progeny virions. Proc. Natl. Acad. Sci. USA 2003, 100, 8460–8465. [Google Scholar] [CrossRef]
- Ameisen, J. Apoptosis subversion:HIV-Nef provides both armor and sword. Nat. Med. 2001, 7, 1181–1182. [Google Scholar] [CrossRef]
- Geleziunas, R.; Xu, W.; Takeda, K.; Ichijo, H.; Greene, W. HIV-1 Nef inhibits ASK1-dependent death signalling providing a potential mechanism for protecting the infected host cell. Nature 2001, 410, 834–838. [Google Scholar] [CrossRef]
- Mahlknecht, U.; Dong, C.; Lu, M.C.; Greenough, T.C.; Sullivan, J.L.; O’brien, W.A.; Herbein, G. Resistance to apoptosis in HIV-infected CD4+ T lymphocytes is mediated by macrophages: Role for Nef and immune activation in viral persistance. J. Immunol. 2000, 165, 6437–6446. [Google Scholar]
- Pandori, M.W.; Fitch, N.J.S.; Craig, H.M.; Richman, D.D.; Spina, C.A.; Guatelli, J.C. Producer-cell modification of human immunodeficiency virus type 1: Nef is a virion protein. J. Virol. 1996, 70, 4283–4290. [Google Scholar]
- Welker, R.; Harris, M.; Cardel, B.; Krausslich, H.-G. Virion incorporation of human immunodeficiency virus type 1 Nef is mediated by a bipartite membrane-targeting signal: Analysis of its role in enhancement of viral infectivity. J. Virol. 1998, 72, 8833–8840. [Google Scholar]
- Laguette, N.; Benichou, S.; Basmaciogullari, S. Human immunodeficiency virus type 1 Nef incorporation into virions does not increase infectivity. J. Virol. 2009, 83, 1093–1104. [Google Scholar] [CrossRef]
- Qi, M.; Aiken, C. Nef enhances HIV-1 infectivity via association with the virus assembly complex. Virology 2008, 373, 287–297. [Google Scholar] [CrossRef]
- Jaque, J.-M.; Mann, A.; Enslen, H.; Sharova, N.; Brichacek, B.; Davis, R.J.; Stevenson, M. Modulation of HIV-1 infectivity by MAPK, a virion-associated kinase. EMBO J. 1998, 17, 2607–2618. [Google Scholar] [CrossRef]
- Ott, D.E.; Coren, L.V.; Johnson, D.G.; Kane, B.P.; Sowder, R.C., II; Kim, Y.D.; Fisher, R.J.; Zhou, X.Z.; Lu, K.P.; Hendersen, L.E. Actin-binding cellular proteins inside human immunodeficiency virus type 1. Virology 2000, 266, 42–51. [Google Scholar] [CrossRef]
- Geyer, M.; Fackler, O.T.; Peterlin, B.M. Structure-function relationships in HIV-1 Nef. EMBO Rep. 2001, 2, 580–585. [Google Scholar] [CrossRef]
- Roeth, J.; Collins, K. Human immunodeficiency virus type 1 Nef: Adapting to intracellular trafficking pathways. Microbiol. Mol. Biol. Rev. 2006, 70, 548–563. [Google Scholar] [CrossRef]
- Carl, S.; Iafrate, A.J.; Lang, S.M.; Stolte, N.; Stahl-Hennig, C.; Matz-Rensing, K.; Fuchs, D.; Skowronski, J.; Kirchhoff, F. Simian immunodeficiency virus containing mutations in N-terminal tyrosine residues and in the PxxP motif in Nef replicates efficicently in rhesus macaques. J. Virol. 2000, 74, 4155–4164. [Google Scholar] [CrossRef]
- Craig, H.M.; Pandori, M.W.; Riggs, N.L.; Richman, D.D.; Guatelli, J.C. Analysis of the SH3-binding region of HIV-1 Nef: Partial functional defects introduced by mutations in the polyproline helix and hydrophobic pocket. Virology 1999, 262, 55–63. [Google Scholar] [CrossRef]
- Fackler, O.; Wolf, D.; Weber, H.; Laffert, B.; D’Aloja, P.; Schuler-Thurner, B.; Geffin, R.; Saksela, K.; Geyer, M.; Peterlin, B.; et al. A natural variability in the proline-rich motif of Nef modulates HIV-1 replication in primary T cells. Curr. Biol. 2001, 11, 1294–1299. [Google Scholar] [CrossRef]
- Saksela, K.; Cheng, G.; Baltimore, D. Proline-rich (PxxP) motifs in HIV-1 Nef bind to SH3 domains of a subset of Src kinases and are required for the enhanced growth of Nef+ viruses but not for down-regulation of CD4. EMBO J. 1995, 14, 484–491. [Google Scholar]
- Arold, S.; Franken, P.; Strub, M.-P.; Hoh, F.; Benichou, S.; Benarous, R.; Dumas, C. The crystal structure of HIV-1 Nef protein bound to the Fyn kinase SH3 domain suggests a role for this complex in altered T cell receptor signalling. Structure 1997, 5, 1361–1372. [Google Scholar] [CrossRef]
- Arold, S.; O’Brien, R.; Franken, P.; Strub, M.-P.; Hoh, F.; Dumas, C.; Ladbury, J. RT loop flexibility enhances the specificity of Src family SH3 domains for HIV-1 Nef. Biochemistry 1998, 37, 14683–14691. [Google Scholar]
- Choi, H.; Smithgall, T. Conserved residues in the HIV-1 Nef hydrophobic pocket are essential for recruitment and activation of the Hck tyrosine kinase. J. Mol. Biol. 1998, 343, 1255–1268. [Google Scholar] [CrossRef]
- Greenway, A.; Azad, A.; Mills, J.; McPhee, D. Human immunodeficiency virus type 1 Nef binds directly to lck and mitogen-activated protein kinase, inhibiting kinase activity. J. Virol. 1996, 70, 6701–6708. [Google Scholar]
- Jung, J.; Byeon, I.-J.L.; Ahn, J.; Gronenborn, A.M. Structure, dynamics and Hck interaction of full-length HIV-1 Nef. Proteins 2011, 79, 1609–1622. [Google Scholar] [CrossRef]
- Lee, C.-H.; Saksela, K.; Mirza, U.; Chait, B.; Kuriyan, J. Crystal structure of the conserved core of HIV-1 Nef complexed with a Src family SH3 domain. Cell 1996, 85, 931–942. [Google Scholar] [CrossRef]
- Briggs, S.; Sharkey, M.; Stevenson, M.; Smithgall, T. SH3-mediated Hck tyrosine kinase activation and FIbroblast transformation by the Nef protein of HIV-1. J. Biol. Chem. 1997, 272, 17899–17902. [Google Scholar]
- Briggs, S.D.; Lerner, E.C.; Smithgall, T.E. Affinity of Src family kinase SH3 domains for HIV Nef in vitro does not predict kinase activation by Nef in vivo. Biochemistry 2000, 39, 489–495. [Google Scholar] [CrossRef]
- Grzesiek, S.; Bax, A.; Clore, G.; Gronenborn, A.; Hu, J.-S.; Kaufman, J.; Palmer, I.; Stahl, S.; Wingfield, P. The solution structure of HIV-1 Nef reveals an unexpected fold and permits delineation of the binding surface for the SH3 domain of Hck tyrosine protein kinase. Nat. Struct. Biol. 1996, 3, 340–345. [Google Scholar] [CrossRef]
- Lee, C.; Leung, B.; Lemmon, M.; Zheng, J.; Cowburn, D.; Kuriyan, J.; Saksela, K. A single amino acid in the SH3 domain of Hck determines its high affinity and specificity in binding to HIV-1 Nef protein. EMBO J. 1995, 14, 5006–5015. [Google Scholar]
- Tokunaga, K.; Kiyokawa, E.; Nakaya, M.; Otsuka, N.; Kojima, A.; Kurata, T.; Matsuda, M. Inhibition of human immunodeficiency virus type 1 virion entry by dominant-negative Hck. J. Virol. 1998, 72, 6257–6259. [Google Scholar]
- Williams, K.C.; Hickley, W.F. Central nervous system damage, monocytes and macrophages, and neurological disorders in AIDS. Ann. Rev. Neurosci. 2002, 25, 537–562. [Google Scholar] [CrossRef]
- Kameoka, M.; Kimura, T.; Ikuta, K. Superoxide enhances the spread of HIV-l infection by cell-to-cell transmission. FEBS 1993, 331, 182–186. [Google Scholar] [CrossRef]
- Pomerantz, R.J.; Horn, D.L. Twenty years of therapy for HIV-1 infection. Nat. Med. 2003, 9, 867–873. [Google Scholar] [CrossRef]
- Swingler, S.; Brichacek, B.; Jacque, J.-M.; Ulich, C.; Zhou, J.; Stevenson, M. HIV-1 Nef intersects the macrophage CD40L signalling pathway to promote resting-cell infection. Nature 2003, 424, 213–219. [Google Scholar] [CrossRef]
- Koppensteiner, H.; Brack-Werner, R.; Schindler, M. Macrophages and their relevance in human immunodeficiency virus type I infection. Retrovirology 2012, 9. [Google Scholar] [CrossRef]
- Le Douce, V.; Herbein, G.; Rohr, O.; Schwartz, C. Molecular mechanisms of HIV-1 persistence in the monocyte-macrophage lineage. Retrovirology 2010, 7. [Google Scholar] [CrossRef] [Green Version]
- Briggs, S.; Scholtz, B.; Jacque, J.; Swingler, S.; Stevenson, M.; Smithgall, T. HIV-1 Nef promotes survival of myeloid cells by a Stat3-dependent pathway. J. Biol. Chem. 2001, 276, 25605–25611. [Google Scholar]
- Kedzierska, K.; Mak, J.; Jaworowski, A.; Greenway, A.; Violo, A.; Chan, H.; Hocking, J.; Purcell, D.; Sullivan, J.; Mills, J.; et al. nef-deleted HIV-1 inhibits phagocytosis by monocyte-derived macrophages in vitro but not by peripheral blood monocytes in vivo. AIDS 2001, 15, 945–955. [Google Scholar] [CrossRef]
- Olivetta, E.; Percario, Z.; Fiorucci, G.; Mattia, G.; Schiavoni, I.; Dennis, C.; Jager, J.; Harris, M.; Romeo, G.; Affabris, E.; et al. HIV-1 Nef induces the release of inflammatory factors from human monocyte/macrophages: Involvement of Nef endocytic SIgnals and NF-κB activation. J. Immunol. 2003, 170, 1716–1727. [Google Scholar]
- Verollet, C.; Zhang, Y.M.; le Cabec, V.; Mazzolini, J.; Charriere, G.; Labrousse, A.; Bouchet, J.; Medina, I.; Biessen, E.; Niedergang, F.; et al. HIV-1 Nef triggers macrophage fusion in a p61Hck- and protease-dependent manner. J. Immunol. 2010, 184, 7030–7039. [Google Scholar] [CrossRef]
- Emert-Sedlak, L.; Kodama, T.; Lerner, E.C.; Dai, W.; Foster, C.; Day, B.W.; Lazo, J.S.; Smithgall, T.E. Chemical library screens targeting an HIV-1 accessory factor/host cell kinase complex identify novel anti-retroviral compounds. ACS Chem. Biol. 2009, 4, 939–947. [Google Scholar] [CrossRef]
- Komuro, I.; Sunazuka, T.; Akagawa, K.S.; Yokota, Y.; Iwamoto, A.; Omura, S. Erythromycin derivatives inhibit HIV-1 replication in macrophages through modulation of MAPK activity to induce small isoforms of C/EBPβ. Proc. Natl. Acad. Sci. USA 2008, 105, 12509–12514. [Google Scholar]
- Komuro, I.; Yokota, Y.; Yasuda, S.; Iwamoto, A.; Kagawa, K.S. CSF-induced and HIV-1—Mediated distinct regulation of Hck and C/EBPβ represent a heterogeneous susceptibility of monocyte-derived macrophages to M-tropic HIV-1 infection. J. Exp. Med. 2003, 198, 443–453. [Google Scholar] [CrossRef]
- Narute, P.S.; Smithgall, T.E. Nef alleles from all major HIV-1 clades activate Src-family kinases and enhance HIV-1 replication in an inhibitor-sensitive manner. PLoS One 2012, 7, e32561. [Google Scholar] [CrossRef] [Green Version]
- Carreno, S.; Gouze, M.-E.; Schaak, S.; Emorine, L.J.; Maridonneau-Parini, I. Lack of palmitoylation redirects p59Hck from the plasma membrane to p61Hck-positive lysosomes. J. Biol. Chem. 2000, 275, 36223–36229. [Google Scholar]
- Lock, P.; Ralph, S.; Stanley, E.; Boulet, I.; Ramsay, R.; Dunn, A.R. Two isoforms of murine hck, generated by utilization of alternative translational initiation codons, exhibit different patterns of subcellular localization. Mol. Cell Biol. 1991, 11, 4363–4370. [Google Scholar]
- Mohn, H.; le Cabec, V.; Fischer, S.; Maridonneau-Parini, I. The src-family protein-tyrosine kinase p59hck is located on the secretory granules in human neutrophils and translocates towards the phagosome during cell activation. Biochem. J. 1995, 309, 657–665. [Google Scholar]
- Robbins, S.M.; Quintrell, N.A.; Bishop, J.M. Myristoylation and differential palmitoylation of the HCK protein-tyrosine kinases govern their attachment to membranes and association with caveolae. Mol. Cell Biol. 1995, 15, 3507–3515. [Google Scholar]
- Wahl, S.M.; Greenwell-Wild, T.; Peng, G.; Ma, G.; Orenstein, J.M.; Vazquez, N. Viral and host cofactors facilitate HIV-1 replication in macrophages. J. Leukoc. Biol. 2003, 74, 726–735. [Google Scholar] [CrossRef]
- Tobiume, M.; Lineberger, J.E.; Lundquist, C.A.; Miller, M.D.; Aiken, C. Nef does not affect the efficiency of human immunodeficiency virus type 1 fusion with target cells. J. Virol. 2003, 77, 10645–10650. [Google Scholar] [CrossRef]
- Sonza, S.; Mutimer, H.P.; O’Brien, K.; Ellery, P.; Howard, J.L.; Axelrod, J.H.; Deacon, N.J.; Crowe, S.M.; Purcell, D.F.J. Selectively reduced Tat mRNA heralds the decline in productive human immunodeficiency virus type 1 infection in monocyte-derived macrophages. J. Virol. 2002, 76, 12611–12621. [Google Scholar] [CrossRef]
- Pene-Dumitrescu, T.; Shu, S.T.; Wales, T.E.; Alvarado, J.J.; Shi, H.; Narute, P.; Moroco, J.A.; Yeh, J.I.; Engen, J.R.; Smithgall, T.E. HIV-1 Nef interaction influences the ATP-binding site of the Src-family kinase, Hck. BMC Chem. Biol. 2012, 12. [Google Scholar] [CrossRef]
- Trible, R.P.; Emert-Sedlak, L.; Smithgall, T.E. HIV-1 Nef selectively activates Src family kinases Hck, Lyn, and c-Src through direct SH3 domain interaction. J. Biol. Chem. 2006, 281, 27029–27038. [Google Scholar] [CrossRef]
- Bukovsky, A.A.; Dorfman, T.; Weimann, A.; Gottlinger, H.G. Nef Association with human immunodeficiency virus type 1 virions and cleavage by the viral protease. J. Virol. 1997, 71, 1013–1018. [Google Scholar]
- Jouvenet, N.; Neil, S.; Bess, C.; Johnson, M.; Virgen, C.; Simon, S.; Bieniasz, P. Plasma membrane is the site of productive HIV-1 particle assembly. PLoS Biol. 2006, 4, 2296–2310. [Google Scholar]
- Frankel, A.D.; Young, J.A.T. HIV-1: Fifteen proteins and an RNA. Annu. Rev. Biochem. 1998, 67, 1–25. [Google Scholar] [CrossRef]
- Goto, T.; Nakai, M.; Ikuta, K. The life-cycle of human immunodeficiency virus type 1. Micron 1998, 29, 123–138. [Google Scholar] [CrossRef]
- Warrilow, D.; Tachedjian, G.; Harrich, D. Maturation of the HIV reverse transcription complex: Putting the jigsaw together. Rev. Med. Virol. 2009, 19, 324–337. [Google Scholar] [CrossRef]
- Carter, C.A.; Ehrlich, L.S. Cell biology of HIV-1 infection of macrophages. Annu. Rev. Microbiol. 2008, 62, 425–443. [Google Scholar] [CrossRef]
- Douaisi, M.; Dussart, S.; Courcoul, M.; Bessou, G.; Lerner, E.C.; Decroly, E.; Vigne, R. The tyrosine kinases Fyn and Hck favor the recruitment of tyrosine-phosphorylated APOBEC3G into vif-defective HIV-1 particles. Biochem. Biophys. Res. Commun. 2005, 329, 917–924. [Google Scholar] [CrossRef]
- Bukrinskaya, A.G.; Ghorpade, A.; Heinzinger, N.K.; Smithgall, T.E.; Lewis, R.E.; Stevenson, M. Phosphorylation-dependent human immunodeficiency virus type 1 infection and nuclear targeting of viral DNA. Proc. Natl. Acad. Sci. USA 1996, 93, 367–371. [Google Scholar]
- Swingler, S.; Gallay, P.; Camaur, D.; Song, J.; Abo, A.; Trono, D. The Nef protein of human immunodeficiency virus type1 enhances serine phosphorylation of the viral matrix. J. Virol. 1997, 71, 4372–4377. [Google Scholar]
- Theodore, T.; Englund, G.; Buckler-White, A.; Buckler, C.; Martin, M.; Peden, K. Construction and characterization of a stable full-length macrophage-tropic HIV type 1 molecular clone that directs the production of high titers of progeny virions. AIDS Res. Hum. Retroviruses 1996, 10, 191–194. [Google Scholar]
- Connor, R.; Chen, B.; Choe, S.; Landau, N. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology 1995, 206, 935–944. [Google Scholar]
- Strebel, K.; Daugherty, D.; Clouse, K.; Cohen, D.; Folks, T.; Martin, M. The HIV A (sor) gene product is essential for virus infectivity. Nature 1987, 328, 728–730. [Google Scholar] [CrossRef]
- Jones, K.; Sonza, S.; Mak, J. Primary T-lymphocytes rescue the replication of HIV-1 DIS RNA mutants in part by facilitating reverse transcription. Nucleic Acids Res. 2008, 36, 1578–1588. [Google Scholar] [CrossRef]
- Pleiman, C.; Abrams, C.; Gauen, L.; Bedzyk, W.; Jongstra, J.; Shaw, A.; Cambier, J. Distinct p53/56lyn and p59fyn domains associate with nonphosphorylated and phosphorylated Ig-α. Proc. Natl. Acad. Sci.USA 1994, 91, 4268–4272. [Google Scholar] [CrossRef]
- Inoue, M.; Koga, Y.; Djordjijevic, D.; Fukuma, T.; Reddy, E.; Yokoyama, M.; Sagawa, K. Down-regulation of CD4 molecules by the expression of Nef: a quantitative analysis of CD4 antigens on the cell surfaces. Int. Immunol. 1993, 5, 1067–1073. [Google Scholar] [CrossRef]
- Chatila, T.; Wong, R.; Young, M.; Miller, R.; Terhorst, C.; Geha, R. An immunodeficiency characterized by defective signal transduction in T lymphocytes. N. Engl. J. Med. 1989, 320, 696–702. [Google Scholar] [CrossRef]
- Goff, S.; Traktman, P.; Baltimore, D. Isolation and properties of moloney murine leukemia virus mutants: Use of a rapid assay for release of virion reverse transcriptase. J. Virol. 1981, 38, 239–248. [Google Scholar]
- Reed, L.; Muench, H. A simple method for estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Sova, P.; Volsky, D.J.; Wang, L.; Chao, W. Vif is largely absent from human immunodeficiency virus type 1 mature virions and associates mainly with viral particles containing unprocessed gag. J. Virol. 2001, 75, 5504–5517. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cornall, A.; Mak, J.; Greenway, A.; Tachedjian, G. HIV-1 Infection of T Cells and Macrophages Are Differentially Modulated by Virion-Associated Hck: A Nef-Dependent Phenomenon. Viruses 2013, 5, 2235-2252. https://doi.org/10.3390/v5092235
Cornall A, Mak J, Greenway A, Tachedjian G. HIV-1 Infection of T Cells and Macrophages Are Differentially Modulated by Virion-Associated Hck: A Nef-Dependent Phenomenon. Viruses. 2013; 5(9):2235-2252. https://doi.org/10.3390/v5092235
Chicago/Turabian StyleCornall, Alyssa, Johnson Mak, Alison Greenway, and Gilda Tachedjian. 2013. "HIV-1 Infection of T Cells and Macrophages Are Differentially Modulated by Virion-Associated Hck: A Nef-Dependent Phenomenon" Viruses 5, no. 9: 2235-2252. https://doi.org/10.3390/v5092235