Association of an Alphasatellite with Tomato Yellow Leaf Curl Virus and Ageratum Yellow Vein Virus in Japan Is Suggestive of a Recent Introduction


Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
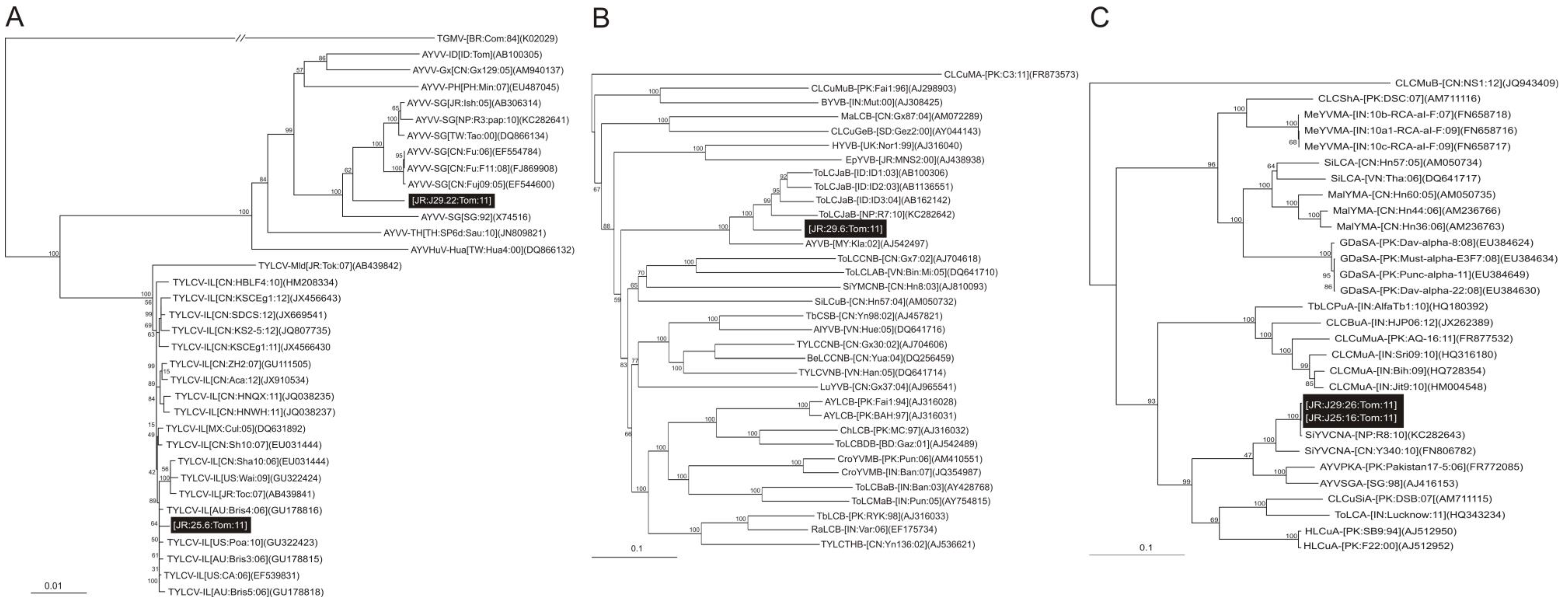
2.1.1. Cloning, Sequencing and Sequence Analysis of Begomoviruses


{kind=link}
{kind=link}
| Isolate | Virus Species | Size (nt) | Genes (coordinates/coding capacity [no. of amino acids]/predicted size of product [kDa]) | |||||
|---|---|---|---|---|---|---|---|---|
| CP | V2 | Rep | TrAP | REn | C4 | |||
| J29.22 | AYVV | 2,751 | 291-1064 | 131-481/ | 2601-1513 | 1613-1206 | 1465-1061 | 2444-2154 |
| /257/29.9 | 116/13.5 | /362/40.8 | /135/15.5 | /134/16.1 | /96/10.8 | |||
| J25.6 | TYLCV | 2,784 | 308-1084 | 148-498 | 2618-1542 | 1633-1226 | 1485-1081 | 2467-2171 |
| /258/30.1 | /116/13.5 | /358/41.0 | /135/15.6 | /134/15.9 | /98/11.2 | |||
2.1.2. Analysis of Betasatellite
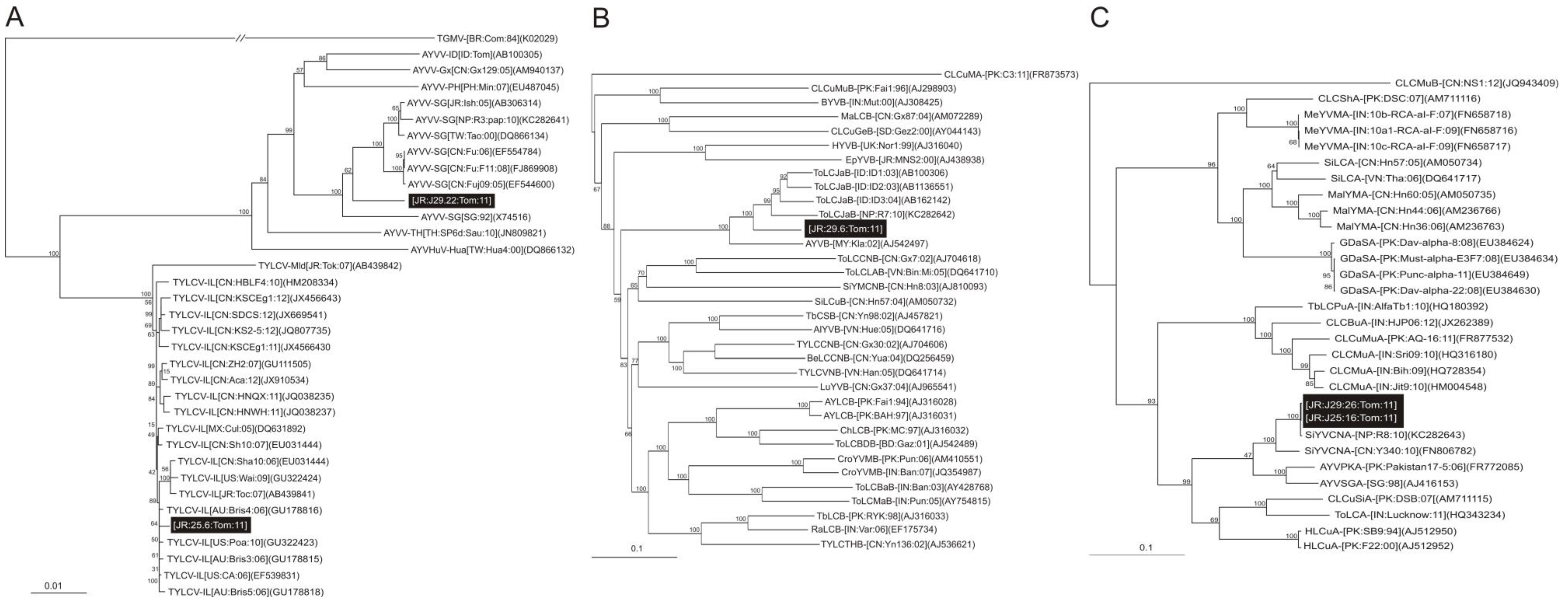
2.1.3. Analysis of Alphasatellites
2.2. Discussion
3. Experimental Section
3.1. Sample Collection, Cloning and Sequencing
3.2. Sequence Analysis
4. Conclusions
Acknowledgments
Author’s Contributions
Conflicts of Interest
References and Notes
- Brown, J.K.; Fauquet, C.M.; Briddon, R.W.; Zerbini, M.; Moriones, E.; Navas-Castillo, J. Virus Taxonomy—Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Lefkowitz, E., Adams, M.J., Carstens, E.B., Eds.; Associated Press: London, UK, 2012; pp. 351–373. [Google Scholar]
- Adams, M.J.; King, A.M.; Carstens, E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2013). Arch. Virol. 2013, 158, 2023–2030. [Google Scholar] [CrossRef]
- Melgarejo, T.A.; Kon, T.; Rojas, M.R.; Paz-Carrasco, L.; Zerbini, F.M.; Gilbertson, R.L. Characterization of a new world monopartite begomovirus causing leaf curl disease of tomato in Ecuador and Peru reveals a new direction in geminivirus evolution. J. Virol. 2013, 87, 5397–5413. [Google Scholar] [CrossRef]
- Sánchez-Campos, S.; Martínez-Ayala, A.; Márquez-Martín, B.; Aragón-Caballero, L.; Navas-Castillo, J.; Moriones, E. Fulfilling Koch’s postulates confirms the monopartite nature of tomato leaf deformation virus: A begomovirus native to the New World. Virus Res. 2013, 173, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Dry, J.; Rigden, L.; Krake, P.; Mullineaux, M.; Rezaian, A. Nucleotide sequence and genome organization of tomato leaf curl geminivirus. J. Gen. Virol. 1993, 74, 147–151. [Google Scholar] [CrossRef]
- Briddon, R.W.; Stanley, J. Subviral agents associated with plant single-stranded DNA viruses. Virology 2006, 344, 198–210. [Google Scholar] [CrossRef]
- Briddon, R.W.; Brown, J.K.; Moriones, E.; Stanley, J.; Zerbini, M.; Zhou, X.; Fauquet, C.M. Recommendations for the classification and nomenclature of the DNA-β satellites of begomoviruses. Arch. Virol. 2008, 153, 763–781. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.; Amin, I.; Idris, A.; Mansoor, S.; Bedford, I.; Dhawan, P.; Rishi, N.; Siwatch, S.; Abdel-Salam, A. Diversity of DNA β: A satellite molecule associated with some monopartite begomoviruses. Virology 2003, 312, 106–121. [Google Scholar] [CrossRef]
- Kon, T.; Sharma, P.; Ikegami, M. Suppressor of RNA silencing encoded by the monopartite tomato leaf curl Java begomovirus. Arch. Virol. 2007, 152, 1273–1282. [Google Scholar] [CrossRef]
- Saeed, S.; Zafar, Y.; Randles, J.; Rezaian, M.A. A monopartite begomovirus-associated DNA β satellite substitutes for the DNA B of a bipartite begomovirus to permit systemic infection. J. Gen. Virol. 2007, 88, 2881–2889. [Google Scholar] [CrossRef]
- Gronenborn, B. Nanoviruses: Genome organisation and protein function. Vet. Microbiol. 2004, 98, 103–110. [Google Scholar] [CrossRef]
- Mansoor, S.; Khan, S.H.; Bashir, A.; Saeed, M.; Zafar, Y.; Malik, K.; Briddon, R.W.; Stanley, J.; Markham, P.G. Identification of a novel circular single-stranded DNA associated with cotton leaf curl disease in Pakistan. Virology 1999, 259, 190–199. [Google Scholar] [CrossRef]
- Saunders, K.; Stanley, J. A nanovirus like DNA component associated with yellow vein disease of Ageratum conyzoides: Evidence for interfamilial recombination between plant DNA viruses. Virology 1999, 264, 142–152. [Google Scholar] [CrossRef]
- Paprotka, T.; Metzler, V.; Jeske, H. The first DNA 1-like alpha-satellites in association with New World begomoviruses in natural infections. Virology 2010, 404, 148–157. [Google Scholar] [CrossRef]
- Romay, G.; Chirinos, D.; Geraud-Pouey, F.; Desbiez, C. Association of an atypical alphasatellite with a bipartite New World begomovirus. Arch. Virol. 2010, 155, 1843–1847. [Google Scholar] [CrossRef]
- Shimizu, S.; Ikegami, M. Complete nucleotide sequence and the genome organization of tobacco leaf curl geminivirus from Japan. Microbiol. Immunol. 1999, 43, 989–992. [Google Scholar] [CrossRef]
- Ogawa, T.; Sharma, P.; Ikegami, M. The begomoviruses, Honeysuckle yellow vein mosaic virus and Tobacco leaf curl Japan virus with DNAβ satellites cause yellow dwarf disease of tomato. Virus Res. 2008, 137, 235–244. [Google Scholar] [CrossRef]
- Andou, T.; Yamaguchi, A.; Kawano, S.; Kawabe, K.; Ueda, S.; Onuki, M. Ageratum yellow vein virus isolated from tomato plants with leaf curl on Ishigaki Island, Okinawa, Japan. J. Gen. Plant Pathol. 2010, 76, 287–291. [Google Scholar] [CrossRef]
- Lefeuvre, P.; Martin, D.P.; Harkins, G.; Lemey, P.; Gray, A.J.; Meredith, S.; Lakay, F.; Monjane, A.; Lett, J.M.; Varsani, A.; et al. The Spread of tomato yellow leaf curl virus from the Middle East to the World. PLoS Pathog. 2010, 6, e1001164. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Onuki, M.; Fuji, S.; Hanada, K. The first occurrence of tomato yellow leaf curl virus in tomato (Lycopersicon esculentum Mill.) in Japan. Ann. Phytopathol. Soc. Jpn. 1998, 64, 552–55. [Google Scholar] [CrossRef]
- Idris, A.M.; Shahid, M.S.; Briddon, R.W.; Khan, A.J.; Zhu, J.K.; Brown, J.K. An unusual alphasatellite associated with monopartite begomoviruses attenuates symptoms and reduces betasatellite accumulation. J. Gen. Virol. 2011, 92, 706–717. [Google Scholar] [CrossRef]
- Orozco, B.M.; Hanley-Bowdoin, L. A DNA structure is required for geminivirus replication origin function. J. Virol. 1996, 7, 148–158. [Google Scholar]
- Shahid, M.S.; Yoshida, S.; Khatri-Chhetri, G.B.; Briddon, R.W.; Natsuaki, K.T. Complete nucleotide sequence of a monopartite begomovirus and associated satellites infecting Carica papaya in Nepal. Virus Genes 2013, 46, 581–584. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.E.; Amin, I.; Mansoor, S.; Bedford, I.D.; Rishi, N.; Siwatch, S.S.; Zafar, Y.; Abdel-Salam, A.M.; Markham, P.G. Diversity of DNA 1: A satellite-like molecule associated with monopartite begomovirus-DNA β complexes. Virology 2004, 324, 462–474. [Google Scholar] [CrossRef]
- Ueda, S.; Kimura, T.; Onuki, M.; Hanada, K.; Iwanami, T. Three distinct groups of isolates of tomato yellow leaf curl virus in Japan and construction of an infectious clone. J. Gen. Plant Pathol. 2004, 70, 232–238. [Google Scholar]
- Ueda, S.; Takeuchi, S.; Okabayashi, M.; Hanada, K.; Tomimura, K.; Iwanami, T. Evidence of a new tomato yellow leaf curl virus in Japan and its detection using PCR. J. Gen. Plant Pathol. 2005, 71, 319–325. [Google Scholar] [CrossRef]
- Kon, T.; Hidayat, S.H.; Hase, S.; Takahashi, H.; Ikegami, M. The natural occurrence of two distinct begomoviruses associated with DNA β and a recombinant DNA in a tomato plant from Indonesia. Phytopathol 2006, 96, 517–525. [Google Scholar] [CrossRef]
- Ueda, S.; Onuki, M.; Yamashita, M.; Yamato, Y. Pathogenicity and insect transmission of a begomovirus complex between tomato yellow leaf curl virus and Ageratum yellow vein betasatellite. Virus Genes 2012, 44, 338–344. [Google Scholar] [CrossRef]
- Nawaz-Ul-Rehman, M.S.; Nahid, N.; Mansoor, S.; Briddon, R.W.; Fauquet, C.M. Post-transcriptional gene silencing suppressor activity of two non-pathogenic alphasatellites associated with a begomovirus. Virology 2010, 405, 300–308. [Google Scholar] [CrossRef]
- Shahid, M.S.; Ito, T.; Kimbara, J.; Onozato, A.; Natsuaki, K.T.; Ikegami, M. Evaluation of tomato hybrids carrying Ty-1 and Ty-2 loci to Japanese monopartite begomovirus species. J. Phytopathol. 2013, 161, 205–209. [Google Scholar] [CrossRef]
- Shahid, M.S.; Mansoor, S.; Briddon, R.W. Complete nucleotide sequences of cotton leaf curl Rajasthan virus and its associated DNA β molecule infecting tomato. Arch. Virol. 2007, 152, 2131–2134. [Google Scholar] [CrossRef]
- Briddon, R.W.; Bull, S.; Mansoor, S.; Amin, I.; Markham, P. Universal primers for the PCR-mediated amplification of DNA β: A molecule associated with some monopartite begomoviruses. Mol. Biotechnol. 2002, 20, 315–318. [Google Scholar] [CrossRef]
- Bull, S.; Briddon, R.W.; Markham, P. Universal primers for the PCR-mediated amplification of DNA1: A satellite-like molecule associated with begomovirus-DNA β complexes. Mol. Biotechnol. 2003, 23, 83–86. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshield, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Page, R.D. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shahid, M.S.; Ikegami, M.; Waheed, A.; Briddon, R.W.; Natsuaki, K.T. Association of an Alphasatellite with Tomato Yellow Leaf Curl Virus and Ageratum Yellow Vein Virus in Japan Is Suggestive of a Recent Introduction. Viruses 2014, 6, 189-200. https://doi.org/10.3390/v6010189
Shahid MS, Ikegami M, Waheed A, Briddon RW, Natsuaki KT. Association of an Alphasatellite with Tomato Yellow Leaf Curl Virus and Ageratum Yellow Vein Virus in Japan Is Suggestive of a Recent Introduction. Viruses. 2014; 6(1):189-200. https://doi.org/10.3390/v6010189
Chicago/Turabian StyleShahid, Muhammad Shafiq, Masato Ikegami, Abdul Waheed, Rob W. Briddon, and Keiko T. Natsuaki. 2014. "Association of an Alphasatellite with Tomato Yellow Leaf Curl Virus and Ageratum Yellow Vein Virus in Japan Is Suggestive of a Recent Introduction" Viruses 6, no. 1: 189-200. https://doi.org/10.3390/v6010189