First Molecular Evidence for Puumala Hantavirus in Poland

Abstract
:1. Introduction
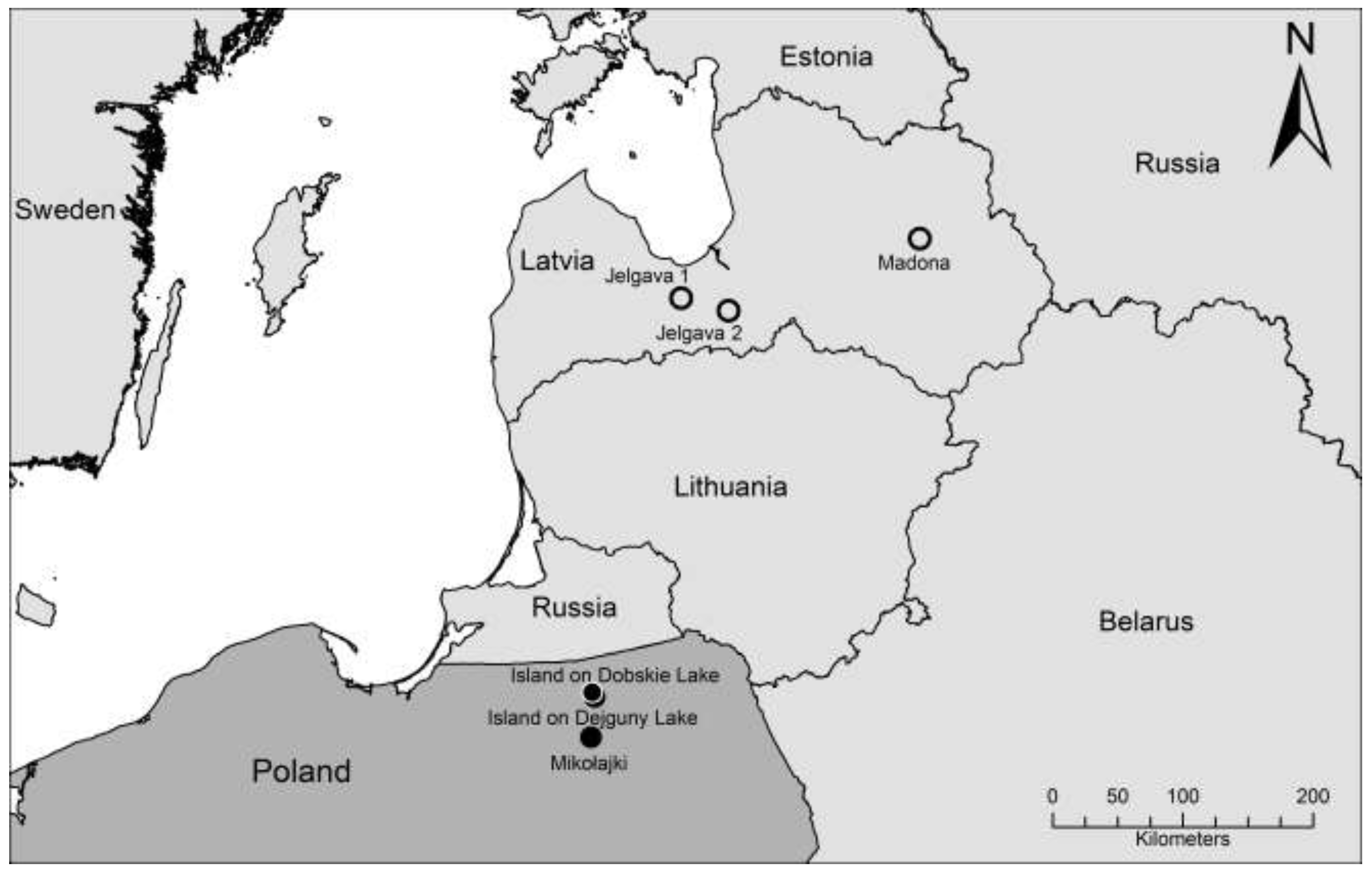
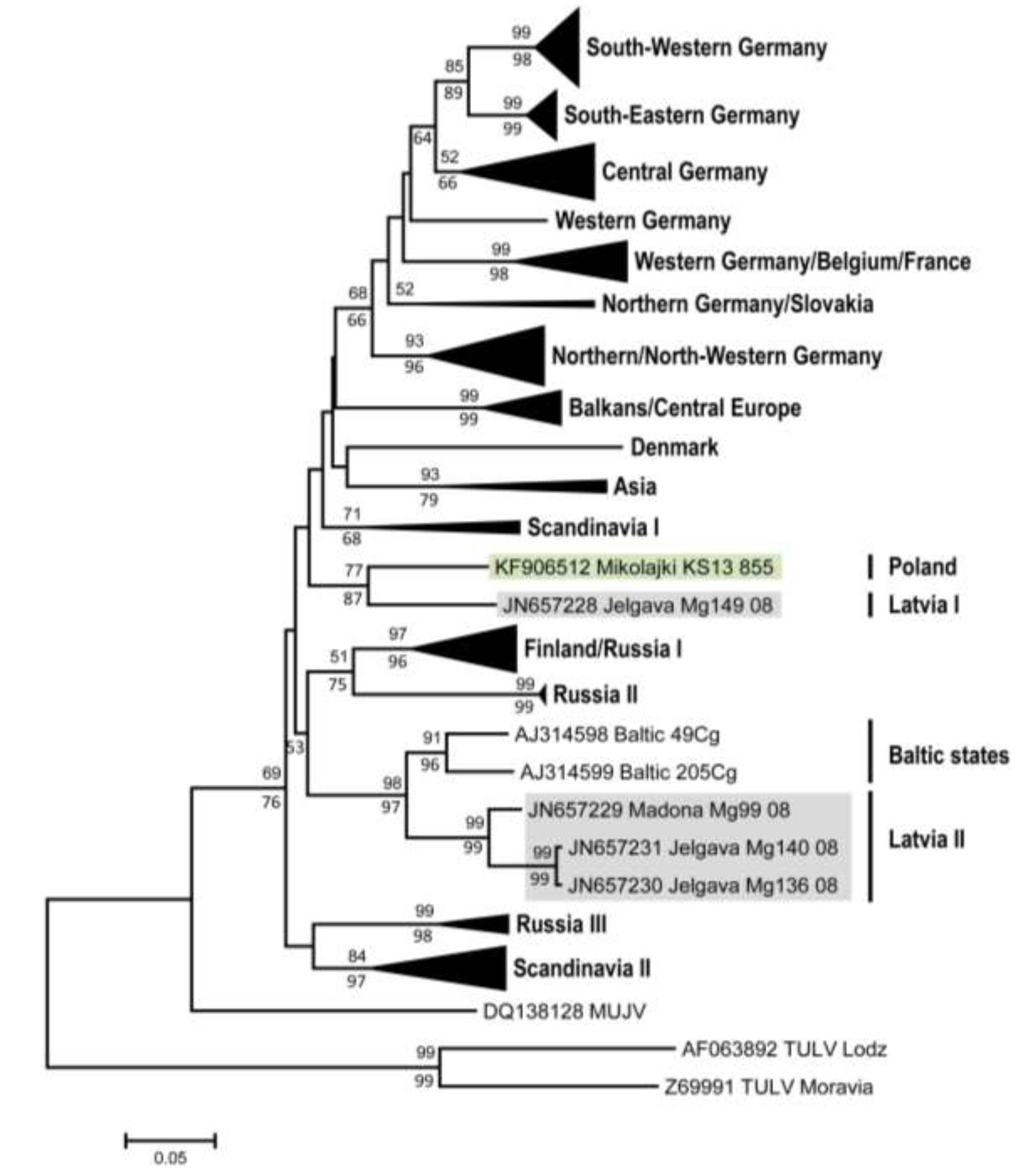
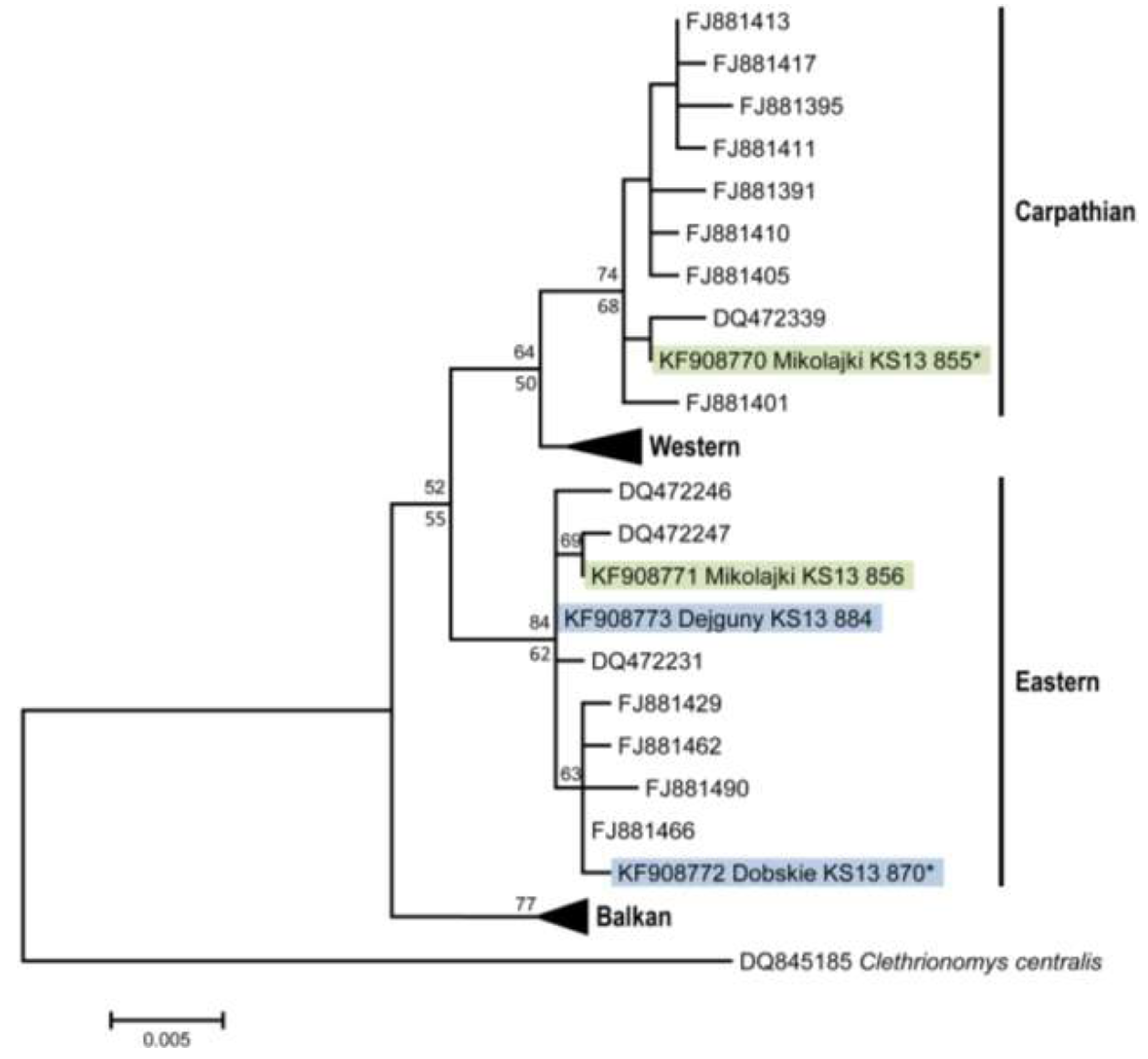
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
| Trapping site | Number of positive/total number of investigated animals | |
|---|---|---|
| Serology | S-RT-PCR | |
| Mikołajki forest | 2/15 * | 3/16 |
| Dobskie island | 0/16 | 0/16 |
| Dejguny island | 0/13 | 0/13 |


| PUUV strains | Mikołajki | Jelgava 1 | Jelgava 2 | Madona | Gerovo | Konnevesi | Vindeln | Bavaria | Fyn |
|---|---|---|---|---|---|---|---|---|---|
| KS13 855 | Mg149 08 | Mg136 08 | Mg99 08 | Mg979 08 | Mg M114B 05 | L20Cg 83 | Mu CG 9 04 | 19 | |
| Mikołajki KS13 855 | 0.888 | 0.808 | 0.825 | 0.827 | 0.860 | 0.819 | 0.819 | 0.787 | |
| Jelgava 1 Mg149 08 | 1 | 0.832 | 0.823 | 0.817 | 0.843 | 0.832 | 0.819 | 0.817 | |
| Jelgava 2 Mg136 08 | 0.948 | 0.948 | 0.948 | 0.789 | 0.806 | 0.806 | 0.780 | 0.782 | |
| Madona Mg99 08 | 0.954 | 0.954 | 0.980 | 0.780 | 0.812 | 0.812 | 0.797 | 0.776 | |
| Gerovo Mg979 08 | 0.954 | 0.954 | 0.922 | 0.935 | 0.821 | 0.810 | 0.825 | 0.800 | |
| Konnevesi Mg M114B 05 | 0.967 | 0.967 | 0.941 | 0.961 | 0.948 | 0.832 | 0.815 | 0.787 | |
| Vindeln L20Cg 83 | 0.967 | 0.967 | 0.935 | 0.954 | 0.935 | 0.948 | 0.817 | 0.819 | |
| Bavaria Mu CG 9 04 | 0.974 | 0.974 | 0.935 | 0.948 | 0.961 | 0.961 | 0.948 | 0.791 | |
| Fyn 19 | 0.961 | 0.961 | 0.922 | 0.935 | 0.935 | 0.935 | 0.941 | 0.954 |

3. Experimental Section
3.1. Rodent Trapping and Dissection
3.2. Serological Investigations
3.3. Nucleic Acid Isolation, Hantavirus RT-PCR, cytochrome b PCR and Sequence Determination
3.4. Sequence Comparison and Phylogenetic Analyses
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Plyusnin, A.; Beaty, B.J.; Elliott, R.M.; Goldbach, R.; Kormelink, R.; Lundkvist, A.; Schmaljohn, C.S.; Tesh, R.B. Bunyaviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier Inc.: San Diego, CA, USA, 2011; pp. 725–741. [Google Scholar]
- Schlegel, M.; Jacob, J.; Krüger, D.H.; Rang, A.; Ulrich, R.G. Hantavirus Emergence in Rodents, Insectivores and Bats. In Role of Animals in Emerging Viral Diseases; Johnson, N., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 235–291. [Google Scholar]
- Krüger, D.H.; Schönrich, G.; Klempa, B. Human pathogenic hantaviruses and prevention of infection. Hum. Vaccin. 2011, 7, 685–693. [Google Scholar] [CrossRef]
- Bi, Z.; Formenty, P.B.; Roth, C.E. Hantavirus infection: A review and global update. J. Infect. Dev. Ctries. 2008, 2, 3–23. [Google Scholar]
- Heyman, P.; Ceianu, C.S.; Christova, I.; Tordo, N.; Beersma, M.; Joao Alves, M.; Lundkvist, A.; Hukic, M.; Papa, A.; Tenorio, A.; et al. A five-year perspective on the situation of haemorrhagic fever with renal syndrome and status of the hantavirus reservoirs in Europe, 2005–2010. Euro Surveill. 2011, 16, 977–986. [Google Scholar]
- Vaheri, A.; Henttonen, H.; Voutilainen, L.; Mustonen, J.; Sironen, T.; Vapalahti, O. Hantavirus infections in Europe and their impact on public health. Rev. Med. Virol. 2013, 23, 35–49. [Google Scholar] [CrossRef]
- Pilaski, J.; Feldmann, H.; Morzunov, S.; Rollin, P.E.; Ruo, S.L.; Lauer, B.; Peters, C.J.; Nichol, S.T. Genetic identification of a new Puumala virus strain causing severe hemorrhagic fever with renal syndrome in Germany. J. Infect. Dis. 1994, 170, 1456–1462. [Google Scholar] [CrossRef]
- Valtonen, M.; Kauppila, M.; Kotilainen, P.; Lahdevirta, J.; Svartback, C.M.; Kosunen, O.; Nurminen, J.; Sarkkinen, H.; Brummer-Korvenkontio, M. Four fatal cases of nephropathia epidemica. Scand. J. Infect. Dis. 1995, 27, 515–517. [Google Scholar] [CrossRef]
- Lahdevirta, J. The minor problem of hemostatic impairment in nephropathia epidemica, the mild Scandinavian form of hemorrhagic fever with renal syndrome. Rev. Infect. Dis. 1989, 11, S860–S863. [Google Scholar] [CrossRef]
- Tesakov, A.S.; Lebedev, V.S.; Bannikova, A.A.; Abramson, N.I. Clethrionomys Tilesius, 1850 is the valid generic name for red-backed voles and Myodes Pallas, 1811 is a junior synonym of Lemmus Link, 1795. Russ. J. Theriol. 2010, 9, 83–86. [Google Scholar]
- Brummer-Korvenkontio, M.; Vaheri, A.; Hovi, T.; von Bonsdorff, C.H.; Vuorimies, J.; Manni, T.; Penttinen, K.; Oker-Blom, N.; Lahdevirta, J. Nephropathia epidemica: Detection of antigen in bank voles and serologic diagnosis of human infection. J. Infect. Dis. 1980, 141, 131–134. [Google Scholar] [CrossRef]
- Kariwa, H.; Yoshizumi, S.; Arikawa, J.; Yoshimatsu, K.; Takahashi, K.; Takashima, I.; Hashimoto, N. Evidence for the existence of Puumula-related virus among Clethrionomys rufocanus in Hokkaido, Japan. Am. J. Trop. Med. Hyg. 1995, 53, 222–227. [Google Scholar]
- Song, K.J.; Baek, L.J.; Moon, S.; Ha, S.J.; Kim, S.H.; Park, K.S.; Klein, T.A.; Sames, W.; Kim, H.C.; Lee, J.S.; et al. Muju virus, a novel hantavirus harboured by the arvicolid rodent Myodes regulus in Korea. J. Gen. Virol. 2007, 88, 3121–3129. [Google Scholar] [CrossRef]
- Aberle, S.W.; Lehner, P.; Ecker, M.; Aberle, J.H.; Arneitz, K.; Khanakah, G.; Radda, A.; Radda, I.; Popow-Kraupp, T.; Kunz, C.; et al. Nephropathia epidemica and Puumala virus in Austria. Eur. J. Clin. Microbiol. Infect. Dis. 1999, 18, 467–472. [Google Scholar] [CrossRef]
- Plyusnina, A.; Aberle, S.W.; Aberle, J.H.; Plyusnin, A. Genetic analysis of Puumala hantavirus strains from Austria. Scand. J. Infect. Dis. 2006, 38, 512–519. [Google Scholar] [CrossRef]
- Plyusnina, A.; Ferenczi, E.; Racz, G.R.; Nemirov, K.; Lundkvist, A.; Vaheri, A.; Vapalahti, O.; Plyusnin, A. Co-circulation of three pathogenic hantaviruses: Puumala, Dobrava, and Saaremaa in Hungary. J. Med. Virol. 2009, 81, 2045–2052. [Google Scholar] [CrossRef]
- Ahlm, C.; Linderholm, M.; Juto, P.; Stegmayr, B.; Settergren, B. Prevalence of serum IgG antibodies to Puumala virus (haemorrhagic fever with renal syndrome) in northern Sweden. Epidemiol. Infect. 1994, 113, 129–136. [Google Scholar] [CrossRef]
- Heyman, P.; Cochez, C.; Ducoffre, G.; Mailles, A.; Zeller, H.; Abu Sin, M.; Koch, J.; van Doornum, G.; Koopmans, M.; Mossong, J.; et al. Haemorrhagic Fever with Renal Syndrome: An analysis of the outbreaks in Belgium, France, Germany, the Netherlands and Luxembourg in 2005. Euro Surveill. 2007, 12, E15–E16. [Google Scholar]
- Groen, J.; Gerding, M.N.; Jordans, J.G.; Clement, J.P.; Nieuwenhuijs, J.H.; Osterhaus, A.D. Hantavirus infections in The Netherlands: Epidemiology and disease. Epidemiol. Infect. 1995, 114, 373–383. [Google Scholar] [CrossRef]
- Nurgaleeva, R.G.; Tkachenko, E.A.; Stepanenko, A.G.; Mustafin, I.M.; Kireev, S.G.; Dzagurova, T.K.; Dekonenko, A.E.; Klimchuk, L.A.; Minin, G.D. An epidemiological analysis of hemorrhagic fever with renal syndrome morbidity in the Republic of Bashkortostan in 1997. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 1999, 6, 45–49. [Google Scholar]
- Sandmann, S.; Meisel, H.; Razanskiene, A.; Wolbert, A.; Pohl, B.; Krüger, D.H.; Sasnauskas, K.; Ulrich, R. Detection of human hantavirus infections in Lithuania. Infection 2005, 33, 66–72. [Google Scholar] [CrossRef]
- Golovljova, I.; Sjolander, K.B.; Lindegren, G.; Vene, S.; Vasilenko, V.; Plyusnin, A.; Lundkvist, A. Hantaviruses in Estonia. J. Med. Virol. 2002, 68, 589–598. [Google Scholar] [CrossRef]
- Lundkvist, A.; Lindegren, G.; Brus Sjolander, K.; Mavtchoutko, V.; Vene, S.; Plyusnin, A.; Kalnina, V. Hantavirus infections in Latvia. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 626–629. [Google Scholar] [CrossRef]
- Bowen, M.D.; Kariwa, H.; Rollin, P.E.; Peters, C.J.; Nichol, S.T. Genetic characterization of a human isolate of Puumala hantavirus from France. Virus Res. 1995, 38, 279–289. [Google Scholar] [CrossRef]
- Cebalo, L.; Dusek, T.; Kuzman, I.; Markotic, A. Grading the severity of disease in patients with Puumala or Dobrava virus infections from 1995 to 2000 in Croatia. Acta Med. Croat. 2003, 57, 355–359. [Google Scholar]
- Leitmeyer, K.; Sibold, C.; Meisel, H.; Ulrich, R.; Labuda, M.; Kruger, D.H. First molecular evidence for Puumala hantavirus in Slovakia. Virus Genes 2001, 23, 165–169. [Google Scholar] [CrossRef]
- Korva, M.; Saksida, A.; Kejzar, N.; Schmaljohn, C.; Avsic-Zupanc, T. Viral load and immune response dynamics in patients with haemorrhagic fever with renal syndrome. Clin. Microbiol. Infect. 2013, 19, E358–E366. [Google Scholar] [CrossRef]
- Papa, A.; Antoniadis, A. Hantavirus infections in Greece—An update. Eur. J. Epidemiol. 2001, 17, 189–194. [Google Scholar] [CrossRef]
- Hukic, M.; Tulumovic, D.; Calkic, L. The renal failure and capillary leak during the acute stage of (Dobrava) DOB and PUU (Puumala) infection. Med. Arh. 2005, 59, 227–230. [Google Scholar]
- Lundkvist, A.; Wiger, D.; Horling, J.; Sjolander, K.B.; Plyusnina, A.; Mehl, R.; Vaheri, A.; Plyusnin, A. Isolation and characterization of Puumala hantavirus from Norway: Evidence for a distinct phylogenetic sublineage. J. Gen. Virol. 1998, 79, 2603–2614. [Google Scholar]
- Clement, J.; Maes, P.; van Ypersele de Strihou, C.; van der Groen, G.; Barrios, J.M.; Verstraeten, W.W.; van Ranst, M. Beechnuts and outbreaks of nephropathia epidemica (NE): Of mast, mice and men. Nephrol. Dial. Transplant. 2010, 25, 1740–1746. [Google Scholar] [CrossRef]
- Sadkowska-Todys, M.; Gut, W.; Baumann, A.; Siennicka, J.; Litwinska, B.; Zielinski, A. Occurrence of human hantavirus infections in Poland. Prz. Epidemiol. 2007, 61, 497–503. [Google Scholar]
- Czarkowski, M.P.; Cielebak, E.; Kondej, E.; Staszewska, E. Infectious Diseases and Poisonings in Poland in 2012; Bulletin of the National Institute of Public Health and Chief Sanitary Inspectorate: Warszawa, Poland, 2013. [Google Scholar]
- Knap, J.P.; Brzostek, T.; Raczka, A.; Burzynski, W.; Litarska, U. A case of haemorrhagic fever with renal syndrome (HFRS). Pol. Merkur Lek. 2006, 21, 474–476. [Google Scholar]
- Nowakowska, A.; Heyman, P.; Knap, J.P.; Burzynski, W.; Witas, M. The first established focus of hantavirus infection in Poland, 2007. Ann. Agric. Environ. Med. 2009, 16, 79–85. [Google Scholar]
- Panasiak, W.; Wleklik, M.; Oraczewska, A.; Luczak, M. Serological studies of haemorrhagic fever with renal syndrome (HFRS) in Poland. Preliminary report. Acta Microbiol. Pol. 1989, 38, 63–67. [Google Scholar]
- Gut, W.; Siennicka, J.; Sadkowska-Todys, M.; Gozdowska, J.; Litwinska, B. The cross and unspecific reactions in serological examination for antibodies against hantavirus Puumala. Prz. Epidemiol. 2007, 61, 489–495. [Google Scholar]
- Grygorczuk, S.; Pancewicz, S.; Zajkowska, J.; Kondrusik, M.; Swierzbinska, R.; Moniuszko, A.; Pawlak-Zalewska, W. Detection of anti-hantavirus antibodies in forest workers in the north-east of Poland. Prz. Epidemiol. 2008, 62, 531–537. [Google Scholar]
- Song, J.W.; Baek, L.J.; Song, K.J.; Skrok, A.; Markowski, J.; Bratosiewicz-Wasik, J.; Kordek, R.; Liberski, P.P.; Yanagihara, R. Characterization of Tula virus from common voles (Microtus arvalis) in Poland: Evidence for geographic-specific phylogenetic clustering. Virus Genes 2004, 29, 239–247. [Google Scholar] [CrossRef]
- Wojcik-Fatla, A.; Zajac, V.; Knap, J.P.; Sroka, J.; Cisak, E.; Sawczyn, A.; Dutkiewicz, J. A small-scale survey of hantavirus in mammals from eastern Poland. Ann. Agric. Environ. Med. 2013, 20, 283–286. [Google Scholar]
- Gu, S.H.; Markowski, J.; Kang, H.J.; Hejduk, J.; Sikorska, B.; Liberski, P.P.; Yanagihara, R. Boginia virus, a newfound hantavirus harbored by the Eurasian water shrew (Neomys fodiens) in Poland. Virol. J. 2013, 10, 160. [Google Scholar] [CrossRef]
- Razzauti, M.; Plyusnina, A.; Niemimaa, J.; Henttonen, H.; Plyusnin, A. Co-circulation of two Puumala hantavirus lineages in Latvia: A Russian lineage described previously and a novel Latvian lineage. J. Med. Virol. 2012, 84, 314–318. [Google Scholar] [CrossRef]
- Wojcik, J.M.; Kawalko, A.; Markova, S.; Searle, J.B.; Kotlik, P. Phylogeographic signatures of northward post-glacial colonization from high-latitude refugia: A case study of bank voles using museum specimens. J. Zool. 2010, 281, 249–262. [Google Scholar]
- Mertens, M.; Kindler, E.; Emmerich, P.; Esser, J.; Wagner-Wiening, C.; Wolfel, R.; Petraityte-Burneikiene, R.; Schmidt-Chanasit, J.; Zvirbliene, A.; Groschup, M.H.; et al. Phylogenetic analysis of Puumala virus subtype Bavaria, characterization and diagnostic use of its recombinant nucleocapsid protein. Virus Genes 2011, 43, 177–191. [Google Scholar] [CrossRef]
- Essbauer, S.; Schmidt, J.; Conraths, F.J.; Friedrich, R.; Koch, J.; Hautmann, W.; Pfeffer, M.; Wolfel, R.; Finke, J.; Dobler, G.; et al. A new Puumala hantavirus subtype in rodents associated with an outbreak of Nephropathia epidemica in South-East Germany in 2004. Epidemiol. Infect. 2006, 134, 1333–1344. [Google Scholar] [CrossRef]
- Schlegel, M.; Ali, H.S.; Stieger, N.; Groschup, M.H.; Wolf, R.; Ulrich, R.G. Molecular identification of small mammal species using novel cytochrome B gene-derived degenerated primers. Biochem. Genet. 2012, 50, 440–447. [Google Scholar] [CrossRef]
- Basic Local Alignment Search Tool (BLAST). Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi/ (accessed on 14 October 2013).
- Braaker, S.; Heckel, G. Transalpine colonisation and partial phylogeographic erosion by dispersal in the common vole (Microtus arvalis). Mol. Ecol. 2009, 18, 2518–2531. [Google Scholar] [CrossRef]
- Ettinger, J.; Hofmann, J.; Enders, M.; Tewald, F.; Oehme, R.M.; Rosenfeld, U.M.; Ali, H.S.; Schlegel, M.; Essbauer, S.; Osterberg, A.; et al. Multiple synchronous outbreaks of Puumala virus, Germany, 2010. Emerg. Infect. Dis. 2012, 18, 1461–1464. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Fink, S.; Fischer, M.C.; Excoffier, L.; Heckel, G. Genomic scans support repetitive continental colonization events during the rapid radiation of voles (Rodentia: Microtus): The utility of AFLPs versus mitochondrial and nuclear sequence markers. Syst. Biol. 2010, 59, 548–572. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ali, H.S.; Drewes, S.; Sadowska, E.T.; Mikowska, M.; Groschup, M.H.; Heckel, G.; Koteja, P.; Ulrich, R.G. First Molecular Evidence for Puumala Hantavirus in Poland. Viruses 2014, 6, 340-353. https://doi.org/10.3390/v6010340
Ali HS, Drewes S, Sadowska ET, Mikowska M, Groschup MH, Heckel G, Koteja P, Ulrich RG. First Molecular Evidence for Puumala Hantavirus in Poland. Viruses. 2014; 6(1):340-353. https://doi.org/10.3390/v6010340
Chicago/Turabian StyleAli, Hanan Sheikh, Stephan Drewes, Edyta T. Sadowska, Magdalena Mikowska, Martin H. Groschup, Gerald Heckel, Pawel Koteja, and Rainer G. Ulrich. 2014. "First Molecular Evidence for Puumala Hantavirus in Poland" Viruses 6, no. 1: 340-353. https://doi.org/10.3390/v6010340