Intracellular Trafficking of the Human Cytomegalovirus-Encoded 7-trans-Membrane Protein Homologs pUS27 and pUL78 during Viral Infection: A Comparative Analysis

Abstract
:1. Introduction
2. Results
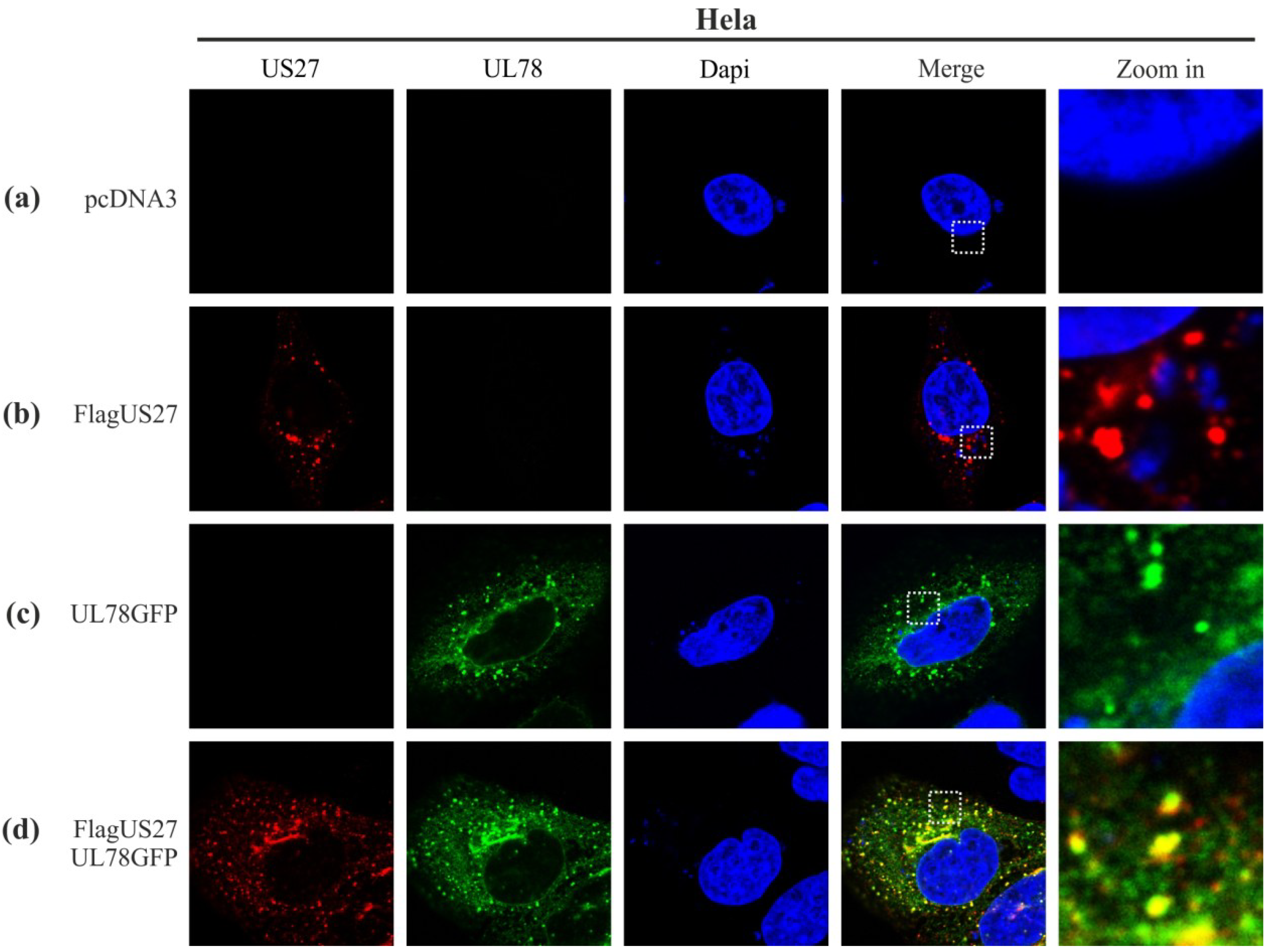
2.1. Colocalization of pUS27 and pUL78 in Transiently Transfected Cells

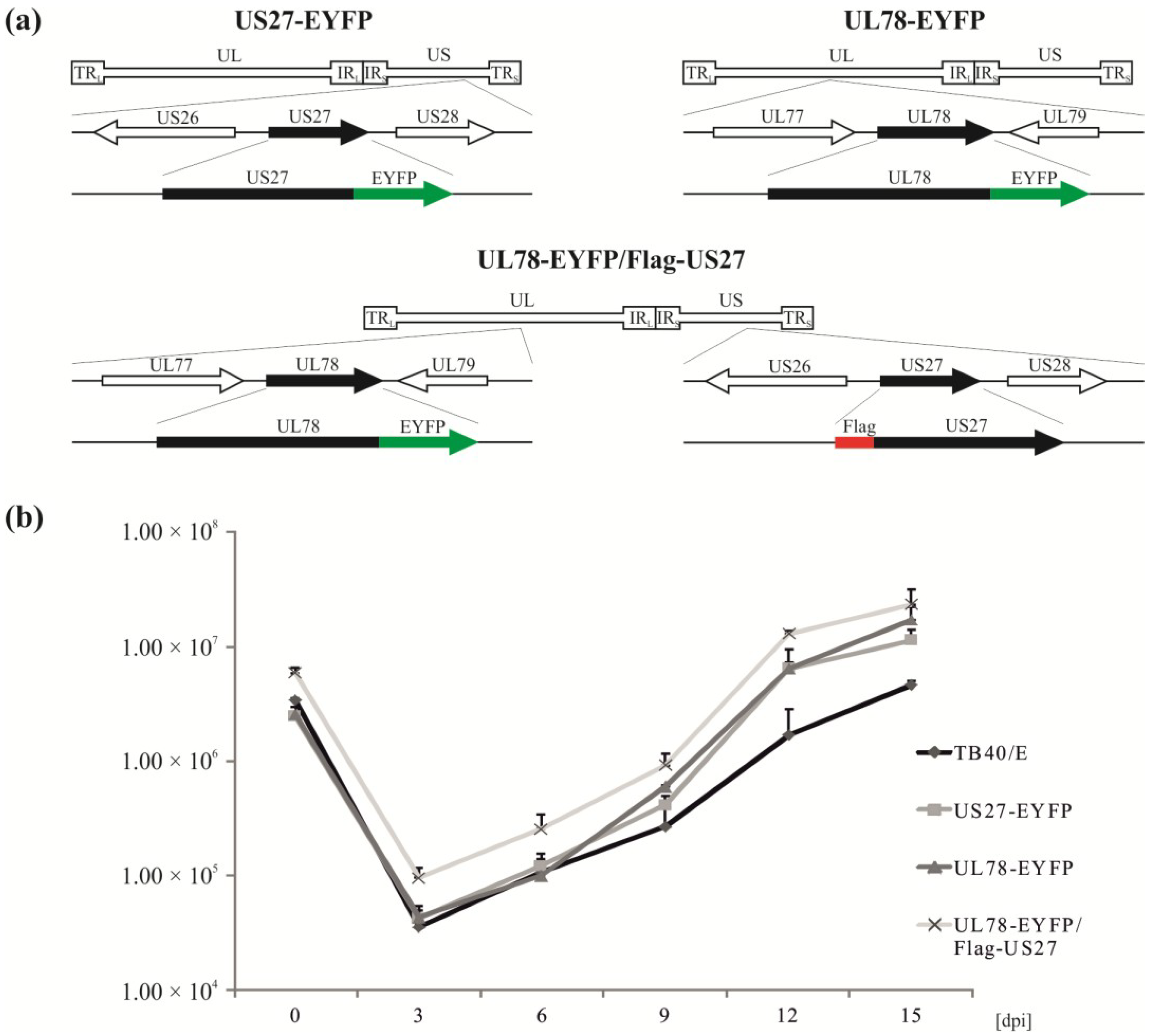
2.2. Recombinant Viruses Harboring Tagged Versions of pUS27 or pUL78 Show Similar Growth Kinetics Compared to Wildtype HCMV Strain TB40/E
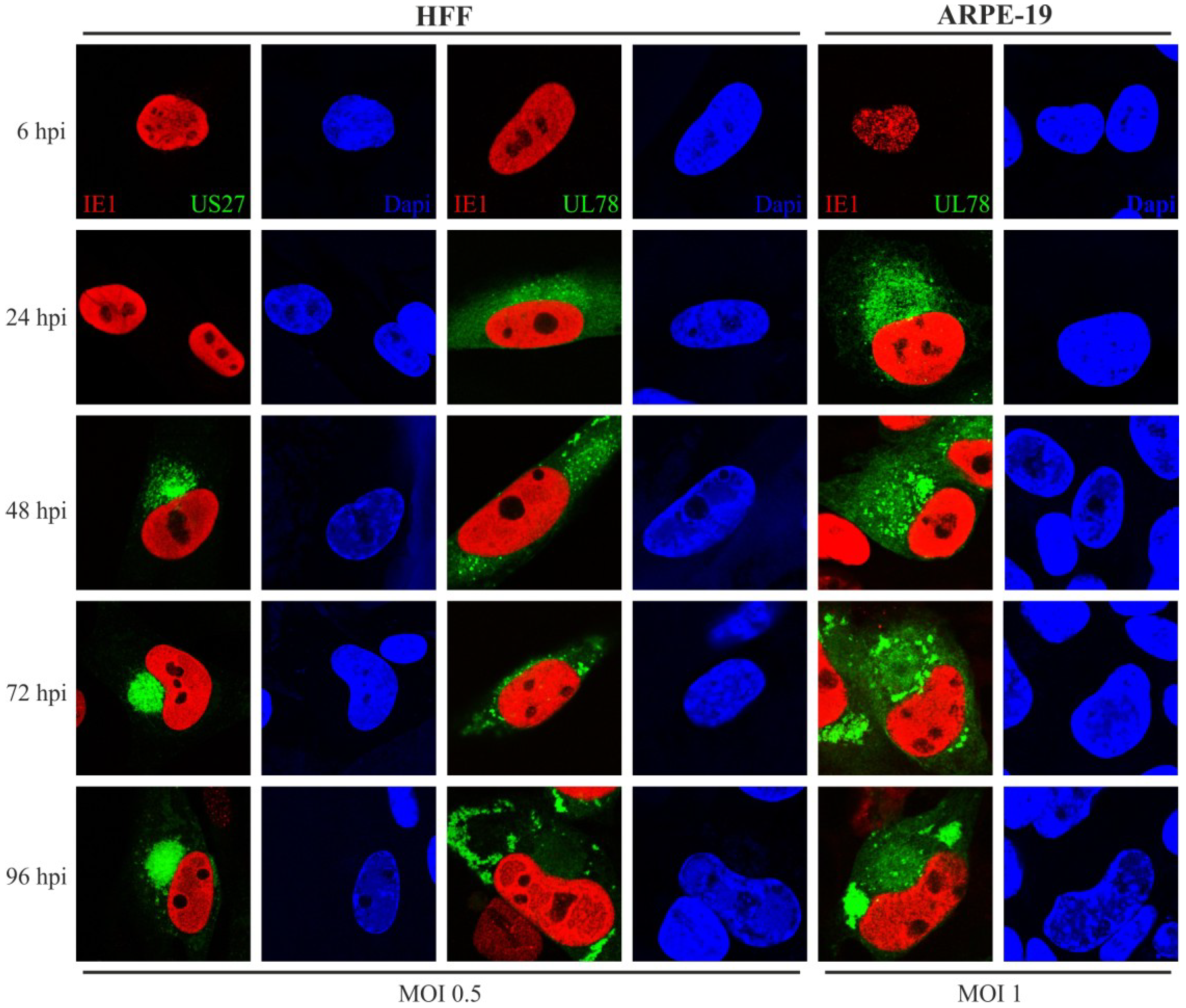
2.3. Expression and Localization Patterns of pUS27 and pUL78 Differ upon HCMV Infection of Fibroblasts and Epithelial Cells


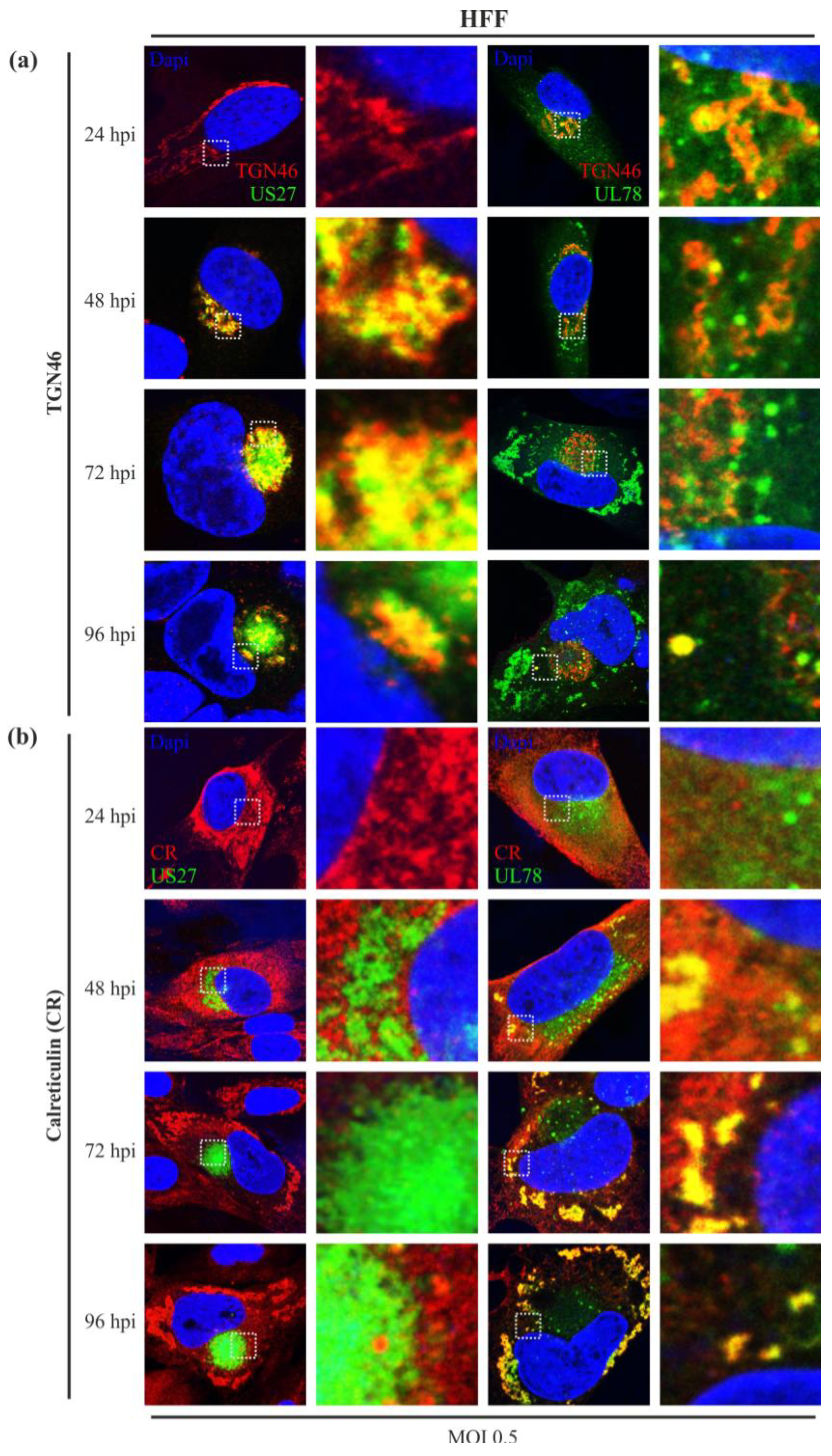
2.4. While Both Receptors Localize to the trans-Golgi Network in Virus Infected Cells, only pUL78 Colocalizes with Calreticulin

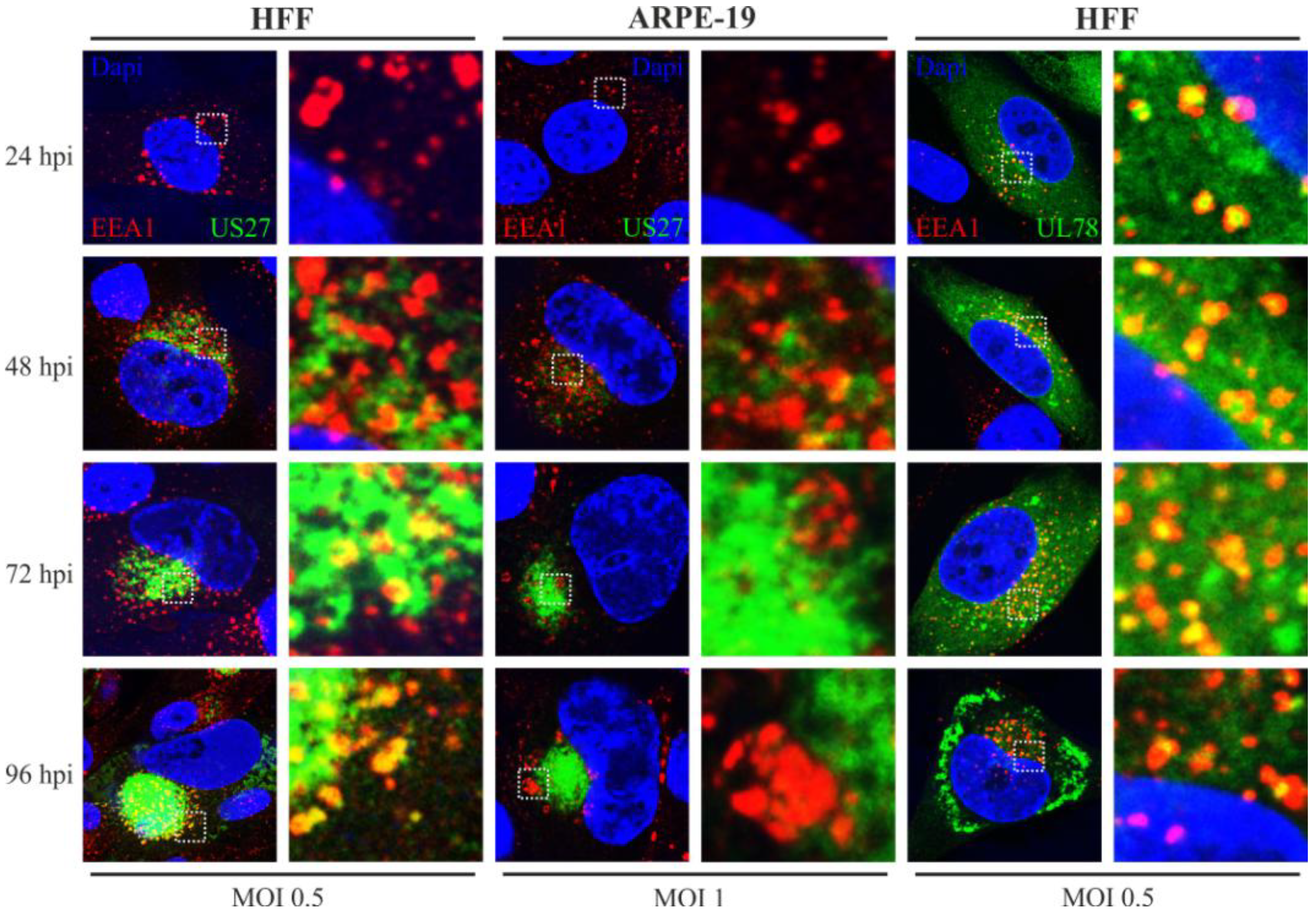
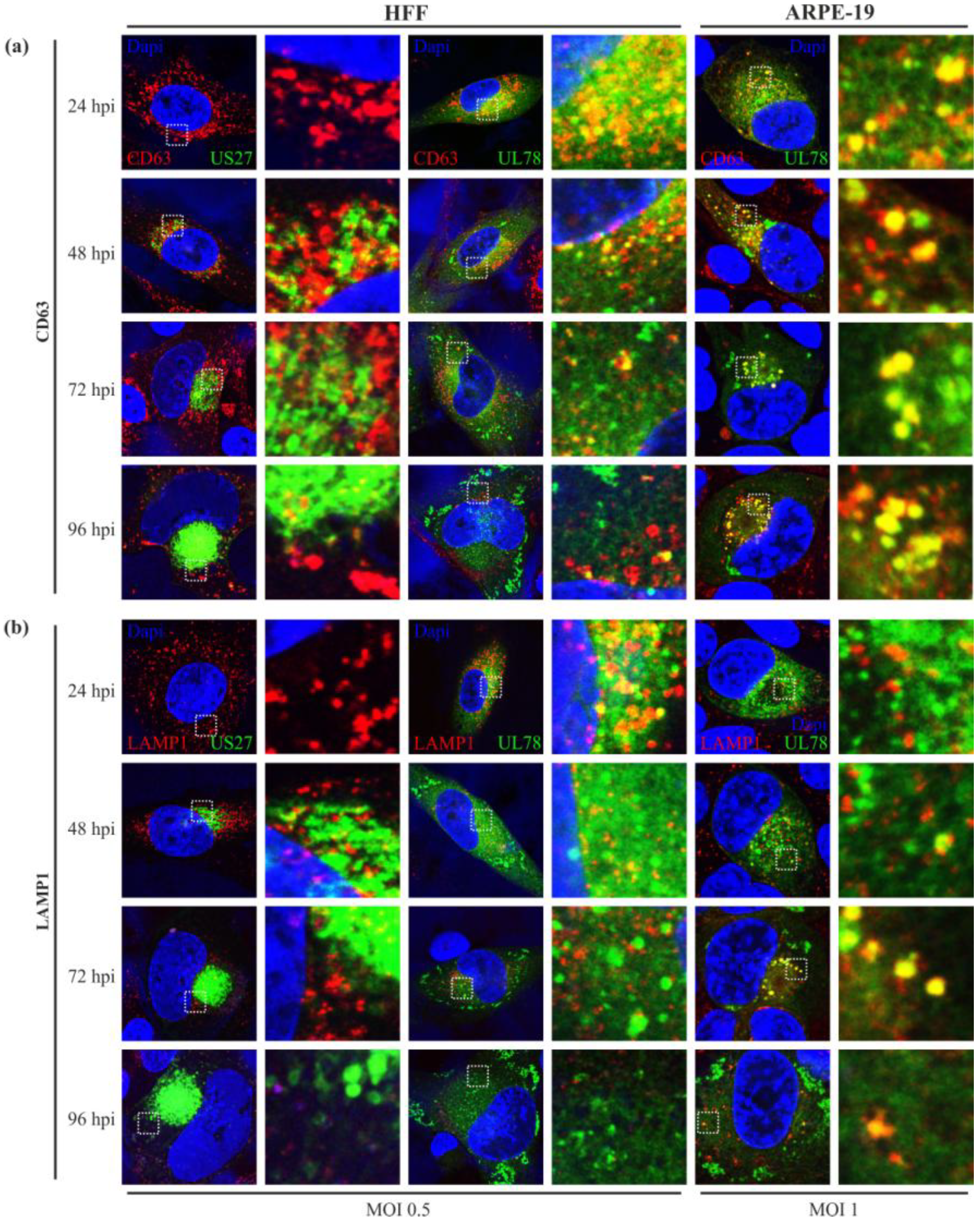
2.5. Endosomal Localization Patterns of pUS27 and pUL78 Diverge in Fibroblasts as Well as in Epithelial Cells during HCMV Replication


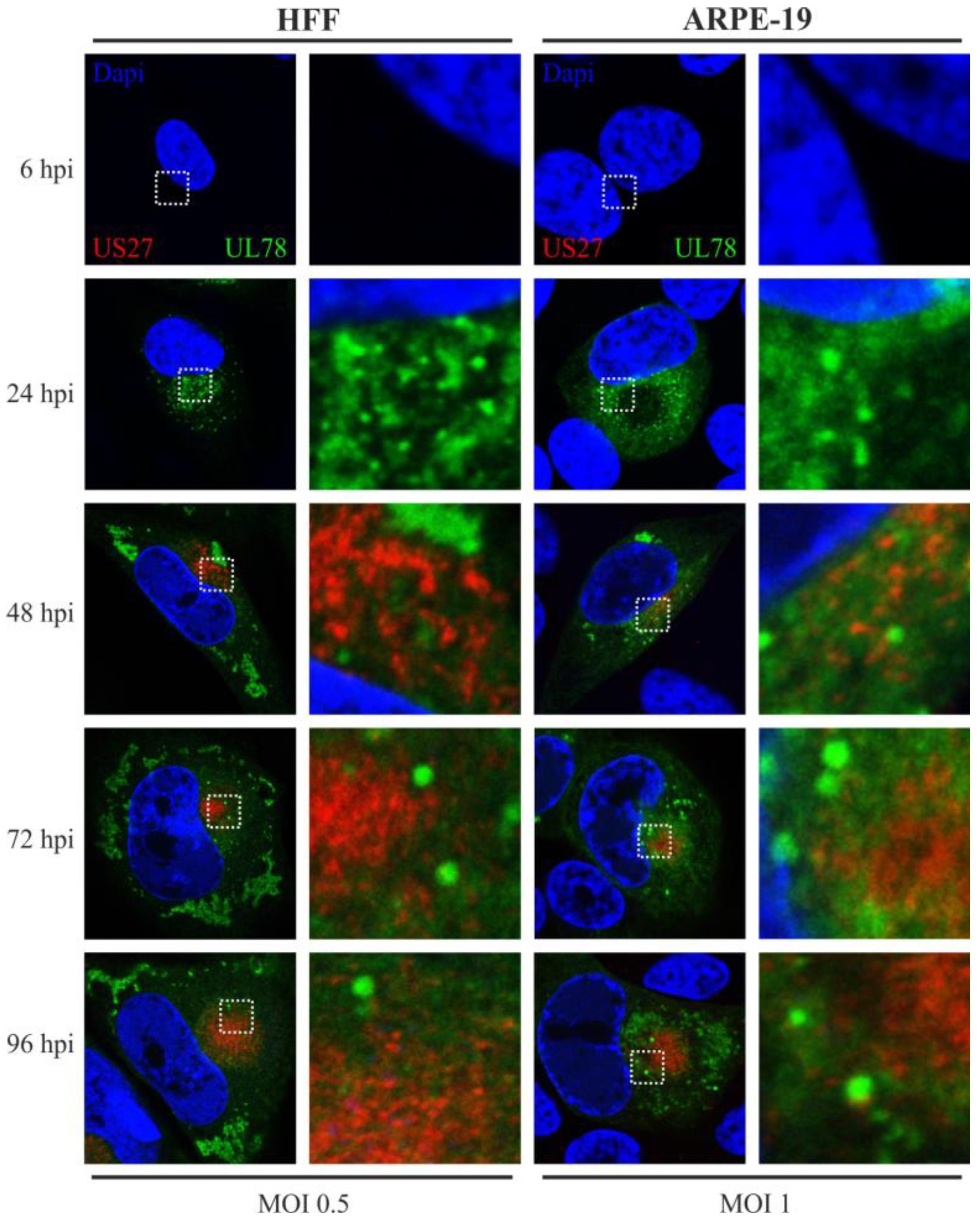
2.6. pUS27 and pUL78 Do Not Colocalize in Infected Fibroblasts or Epithelial Cells throughout the Replication Cycle

3. Discussion
4. Experimental Section
4.1. Cells
4.2. Construction of Vectors for Subcellular Localization Analyses
4.3. Transient Expression Analyses and Protein Localization
4.4. Viruses, Generation of Recombinant HCMVs, and Growth Curve Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Construct | Sequence in 5'-3' |
|---|---|---|
| Flag-kTischer-fw | Flag-US27 PCR1 | GAT TAC AAG GAT GAC GAC GAT AAG |
| Flag-US27-rev | CAT ATC aga att ggt taa ttg gtt ggt aac act acc tgt aag gtg atg gat tac aag gat gac gac gat aag atg acc acc tct aca aac caa acc tta aca cag gtg agc aac atg aca aa | |
| Flag-kUS27-rev | Flag-US27 PCR2 | ttt gtc atg ttg ctc acc tgt gtt |
| Flag-US27-fw | tgt tat gct ttt tac agg acc gtt cag cag gta aca cta cct gta agg tga tgg att aca agg atg acg acg ata ag | |
| US27-EYFP-fw | US27-EYFP | atg aca gaa aaa atg cac cta tgg agt ccg ggg agg agg aat ttc tat tga tgg tga gca agg gcg agg agc tg |
| US27-EYFP-rev | gtg caa tga gca aaa ata gat gtg cgg cgg acg cgt gaa aga gga tcg aat tac ttg tac agc tcg tcc atg ccg ag | |
| UL78-EYFP-fw | UL78-EYFP | gca ccg acg gcg aaa aca ccg tcg cgt ccg acg caa cgg tga cgg cat taa tgg tga gca agg gcg agg agc tg |
| UL78-EYFP-rev | gac gtg att tat ctg cca ctt ttc tcc ccg ctg ccg tac agc gcc gcc gct tac ttg tac agc tcg tcc atg ccg ag |
4.5. HCMV Infection and Protein Localization
| Target protein | Localization | Dilution | Nature of antibody (source) |
|---|---|---|---|
| IE1 | nucleus | 1:4 | MAb, clone p63–27 [57] |
| TGN46 | trans-Golgi network | 1:300 | Sheep polyclonal antibody (AbD Serotec catalog No. AHP500G) |
| calreticulin | endoplasmic reticulum | 1:100 | Rabbit polyclonal antibody (Thermo Fisher catalog No. PA3-900) |
| EEA1 | early endosomes | 1:200 | Rabbit polyclonal antibody, clone H-300 (SCBT catalog No. 33585) |
| CD63 | multivesicular bodies, late endosomes | 1:50 | MAb, clone 4X-49.129.5 (SCBT catalog No. 5275) |
| LAMP1 | lysosomes | 1:50 | MAb, clone H5G11 (SCBT catalog No. 18821) |
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Montaner, S.; Kufareva, I.; Abagyan, R.; Gutkind, J.S. Molecular mechanisms deployed by virally encoded G protein-coupled receptors in human diseases. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 331–354. [Google Scholar] [CrossRef]
- Couty, J.P.; Gershengorn, M.C. G-protein-coupled receptors encoded by human herpesviruses. Trends Pharmacol. Sci. 2005, 26, 405–411. [Google Scholar] [CrossRef]
- Sodhi, A.; Montaner, S.; Gutkind, J.S. Viral hijacking of G-protein-coupled-receptor signalling networks. Nat. Rev. Mol. Cell Biol. 2004, 5, 998–1012. [Google Scholar] [CrossRef]
- Chee, M.S.; Satchwell, S.C.; Preddie, E.; Weston, K.M.; Barrell, B.G. Human cytomegalovirus encodes three G protein-coupled receptor homologues. Nature 1990, 344, 774–777. [Google Scholar] [CrossRef]
- Attwood, T.K.; Findlay, J.B. Fingerprinting G-protein-coupled receptors. Protein Eng. 1994, 7, 195–203. [Google Scholar] [CrossRef]
- Margulies, B.J.; Browne, H.; Gibson, W. Identification of the human cytomegalovirus G protein-coupled receptor homologue encoded by UL33 in infected cells and enveloped virus particles. Virology 1996, 225, 111–125. [Google Scholar] [CrossRef]
- Bodaghi, B.; Jones, T.R.; Zipeto, D.; Vita, C.; Sun, L.; Laurent, L.; renzana-Seisdedos, F.; Virelizier, J.L.; Michelson, S. Chemokine sequestration by viral chemoreceptors as a novel viral escape strategy: Withdrawal of chemokines from the environment of cytomegalovirus-infected cells. J. Exp. Med. 1998, 188, 855–866. [Google Scholar] [CrossRef]
- Cardin, R.D.; Schaefer, G.C.; Allen, J.R.; vis-Poynter, N.J.; Farrell, H.E. The M33 chemokine receptor homolog of murine cytomegalovirus exhibits a differential tissue-specific role during in vivo replication and latency. J. Virol. 2009, 83, 7590–7601. [Google Scholar] [CrossRef]
- Beisser, P.S.; Grauls, G.; Bruggeman, C.A.; Vink, C. Deletion of the R78 G protein-coupled receptor gene from rat cytomegalovirus results in an attenuated, syncytium-inducing mutant strain. J. Virol. 1999, 73, 7218–7230. [Google Scholar]
- Casarosa, P.; Bakker, R.A.; Verzijl, D.; Navis, M.; Timmerman, H.; Leurs, R.; Smit, M.J. Constitutive signaling of the human cytomegalovirus-encoded chemokine receptor US28. J. Biol. Chem. 2001, 276, 1133–1137. [Google Scholar]
- Waldhoer, M.; Kledal, T.N.; Farrell, H.; Schwartz, T.W. Murine cytomegalovirus (CMV) M33 and human CMV US28 receptors exhibit similar constitutive signaling activities. J. Virol. 2002, 76, 8161–8168. [Google Scholar] [CrossRef]
- Gao, J.L.; Murphy, P.M. Human cytomegalovirus open reading frame US28 encodes a functional beta chemokine receptor. J. Biol. Chem. 1994, 269, 28539–28542. [Google Scholar]
- Waldhoer, M.; Casarosa, P.; Rosenkilde, M.M.; Smit, M.J.; Leurs, R.; Whistler, J.L.; Schwartz, T.W. The carboxyl terminus of human cytomegalovirus-encoded 7 transmembrane receptor US28 camouflages agonism by mediating constitutive endocytosis. J. Biol. Chem. 2003, 278, 19473–19482. [Google Scholar]
- Miller, W.E.; Houtz, D.A.; Nelson, C.D.; Kolattukudy, P.E.; Lefkowitz, R.J. G-protein-coupled receptor (GPCR) kinase phosphorylation and beta-arrestin recruitment regulate the constitutive signaling activity of the human cytomegalovirus US28 GPCR. J. Biol. Chem. 2003, 278, 21663–21671. [Google Scholar]
- Fraile-Ramos, A.; Kledal, T.N.; Pelchen-Matthews, A.; Bowers, K.; Schwartz, T.W.; Marsh, M. The human cytomegalovirus US28 protein is located in endocytic vesicles and undergoes constitutive endocytosis and recycling. Mol. Biol. Cell 2001, 12, 1737–1749. [Google Scholar] [CrossRef]
- Mokros, T.; Rehm, A.; Droese, J.; Oppermann, M.; Lipp, M.; Hopken, U.E. Surface expression and endocytosis of the human cytomegalovirus-encoded chemokine receptor US28 is regulated by agonist-independent phosphorylation. J. Biol. Chem. 2002, 277, 45122–45128. [Google Scholar]
- Billstrom, M.A.; Lehman, L.A.; Scott, W.G. Depletion of extracellular RANTES during human cytomegalovirus infection of endothelial cells. Am. J. Respir. Cell Mol. Biol. 1999, 21, 163–167. [Google Scholar] [CrossRef]
- Randolph-Habecker, J.R.; Rahill, B.; Torok-Storb, B.; Vieira, J.; Kolattukudy, P.E.; Rovin, B.H.; Sedmak, D.D. The expression of the cytomegalovirus chemokine receptor homolog US28 sequesters biologically active CC chemokines and alters IL-8 production. Cytokine 2002, 19, 37–46. [Google Scholar] [CrossRef]
- Vieira, J.; Schall, T.J.; Corey, L.; Geballe, A.P. Functional analysis of the human cytomegalovirus US28 gene by insertion mutagenesis with the green fluorescent protein gene. J. Virol. 1998, 72, 8158–8165. [Google Scholar]
- Minisini, R.; Tulone, C.; Luske, A.; Michel, D.; Mertens, T.; Gierschik, P.; Moepps, B. Constitutive inositol phosphate formation in cytomegalovirus-infected human fibroblasts is due to expression of the chemokine receptor homologue pUS28. J. Virol. 2003, 77, 4489–4501. [Google Scholar] [CrossRef]
- Vischer, H.F.; Leurs, R.; Smit, M.J. HCMV-encoded G-protein-coupled receptors as constitutively active modulators of cellular signaling networks. Trends Pharmacol. Sci. 2006, 27, 56–63. [Google Scholar] [CrossRef]
- McLean, K.A.; Holst, P.J.; Martini, L.; Schwartz, T.W.; Rosenkilde, M.M. Similar activation of signal transduction pathways by the herpesvirus-encoded chemokine receptors US28 and ORF74. Virology 2004, 325, 241–251. [Google Scholar] [CrossRef]
- Farrell, H.E.; Abraham, A.M.; Cardin, R.D.; Sparre-Ulrich, A.H.; Rosenkilde, M.M.; Spiess, K.; Jensen, T.H.; Kledal, T.N.; vis-Poynter, N. Partial functional complementation between human and mouse cytomegalovirus chemokine receptor homologues. J. Virol. 2011, 85, 6091–6095. [Google Scholar] [CrossRef]
- Farrell, H.E.; Abraham, A.M.; Cardin, R.D.; Molleskov-Jensen, A.S.; Rosenkilde, M.M.; vis-Poynter, N. Identification of common mechanisms by which human and mouse cytomegalovirus seven-transmembrane receptor homologues contribute to in vivo phenotypes in a mouse model. J. Virol. 2013, 87, 4112–4117. [Google Scholar] [CrossRef]
- Gruijthuijsen, Y.K.; Casarosa, P.; Kaptein, S.J.; Broers, J.L.; Leurs, R.; Bruggeman, C.A.; Smit, M.J.; Vink, C. The rat cytomegalovirus R33-encoded G protein-coupled receptor signals in a constitutive fashion. J. Virol. 2002, 76, 1328–1338. [Google Scholar] [CrossRef]
- Casarosa, P.; Gruijthuijsen, Y.K.; Michel, D.; Beisser, P.S.; Holl, J.; Fitzsimons, C.P.; Verzijl, D.; Bruggeman, C.A.; Mertens, T.; Leurs, R.; et al. Constitutive signaling of the human cytomegalovirus- encoded receptor UL33 differs from that of its rat cytomegalovirus homolog R33 by promiscuous activation of G proteins of the Gq, Gi, and Gs classes. J. Biol. Chem. 2003, 278, 50010–50023. [Google Scholar] [CrossRef]
- Beisser, P.S.; Vink, C.; van Dam, J.G.; Grauls, G.; Vanherle, S.J.; Bruggeman, C.A. The R33 G protein-coupled receptor gene of rat cytomegalovirus plays an essential role in the pathogenesis of viral infection. J. Virol. 1998, 72, 2352–2363. [Google Scholar]
- Case, R.; Sharp, E.; ned-Jensen, T.; Rosenkilde, M.M.; vis-Poynter, N.; Farrell, H.E. Functional analysis of the murine cytomegalovirus chemokine receptor homologue M33: Ablation of constitutive signaling is associated with an attenuated phenotype in vivo. J. Virol. 2008, 82, 1884–1898. [Google Scholar] [CrossRef]
- Stapleton, L.K.; Arnolds, K.L.; Lares, A.P.; Devito, T.M.; Spencer, J.V. Receptor chimeras demonstrate that the C-terminal domain of the human cytomegalovirus US27 gene product is necessary and sufficient for intracellular receptor localization. Virol. J. 2012, 9. [Google Scholar] [CrossRef]
- Fraile-Ramos, A.; Pelchen-Matthews, A.; Kledal, T.N.; Browne, H.; Schwartz, T.W.; Marsh, M. Localization of HCMV UL33 and US27 in endocytic compartments and viral membranes. Traffic 2002, 3, 218–232. [Google Scholar] [CrossRef]
- Margulies, B.J.; Gibson, W. The chemokine receptor homologue encoded by US27 of human cytomegalovirus is heavily glycosylated and is present in infected human foreskin fibroblasts and enveloped virus particles. Virus Res. 2007, 123, 57–71. [Google Scholar] [CrossRef]
- O’Connor, C.M.; Shenk, T. Human cytomegalovirus pUS27 G protein-coupled receptor homologue is required for efficient spread by the extracellular route but not for direct cell-to-cell spread. J. Virol. 2011, 85, 3700–3707. [Google Scholar] [CrossRef]
- O’Connor, C.M.; Shenk, T. Human cytomegalovirus pUL78 G protein-coupled receptor homologue is required for timely cell entry in epithelial cells but not fibroblasts. J. Virol. 2012, 86, 11425–11433. [Google Scholar] [CrossRef]
- Kaptein, S.J.; Beisser, P.S.; Gruijthuijsen, Y.K.; Savelkouls, K.G.; van Cleef, K.W.; Beuken, E.; Grauls, G.E.; Bruggeman, C.A.; Vink, C. The rat cytomegalovirus R78 G protein-coupled receptor gene is required for production of infectious virus in the spleen. J. Gen. Virol. 2003, 84, 2517–2530. [Google Scholar] [CrossRef]
- Oliveira, S.A.; Shenk, T.E. Murine cytomegalovirus M78 protein, a G protein-coupled receptor homologue, is a constituent of the virion and facilitates accumulation of immediate-early viral mRNA. Proc. Natl. Acad. Sci. USA 2001, 98, 3237–3242. [Google Scholar] [CrossRef]
- Wagner, S.; Arnold, F.; Wu, Z.; Schubert, A.; Walliser, C.; Tadagaki, K.; Jockers, R.; Mertens, T.; Michel, D. The 7-transmembrane protein homologue UL78 of the human cytomegalovirus forms oligomers and traffics between the plasma membrane and different intracellular compartments. Arch. Virol. 2012, 157, 935–949. [Google Scholar] [CrossRef]
- Tschische, P.; Tadagaki, K.; Kamal, M.; Jockers, R.; Waldhoer, M. Heteromerization of human cytomegalovirus encoded chemokine receptors. Biochem. Pharmacol. 2011, 82, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Tischer, B.K.; von, E.J.; Kaufer, B.; Osterrieder, N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. Biotechniques 2006, 40, 191–197. [Google Scholar]
- Sinzger, C.; Hahn, G.; Digel, M.; Katona, R.; Sampaio, K.L.; Messerle, M.; Hengel, H.; Koszinowski, U.; Brune, W.; Adler, B. Cloning and sequencing of a highly productive, endotheliotropic virus strain derived from human cytomegalovirus TB40/E. J. Gen. Virol. 2008, 89, 359–368. [Google Scholar] [CrossRef]
- Resnik, K.S.; DiLeonardo, M.; Maillet, M. Histopathologic findings in cutaneous cytomegalovirus infection. Am. J. Dermatopathol. 2000, 22, 397–407. [Google Scholar] [CrossRef]
- Das, S.; Vasanji, A.; Pellett, P.E. Three-dimensional structure of the human cytomegalovirus cytoplasmic virion assembly complex includes a reoriented secretory apparatus. J. Virol. 2007, 81, 11861–11869. [Google Scholar] [CrossRef]
- Das, S.; Pellett, P.E. Spatial relationships between markers for secretory and endosomal machinery in human cytomegalovirus-infected cells versus those in uninfected cells. J. Virol. 2011, 85, 5864–5879. [Google Scholar] [CrossRef]
- Cepeda, V.; Esteban, M.; Fraile-Ramos, A. Human cytomegalovirus final envelopment on membranes containing both trans-Golgi network and endosomal markers. Cell Microbiol. 2010, 12, 386–404. [Google Scholar] [CrossRef]
- Sinzger, C.; Digel, M.; Jahn, G. Cytomegalovirus cell tropism. Curr. Top. Microbiol. Immunol. 2008, 325, 63–83. [Google Scholar]
- Sharp, E.L.; vis-Poynter, N.J.; Farrell, H.E. Analysis of the subcellular trafficking properties of murine cytomegalovirus M78, a 7 transmembrane receptor homologue. J. Gen. Virol. 2009, 90, 59–68. [Google Scholar] [CrossRef]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Hanyaloglu, A.C.; Von, Z.M. Regulation of GPCRs by endocytic membrane trafficking and its potential implications. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 537–568. [Google Scholar] [CrossRef]
- Michel, D.; Milotic, I.; Wagner, M.; Vaida, B.; Holl, J.; Ansorge, R.; Mertens, T. The human cytomegalovirus UL78 gene is highly conserved among clinical isolates, but is dispensable for replication in fibroblasts and a renal artery organ-culture system. J. Gen. Virol. 2005, 86, 297–306. [Google Scholar] [CrossRef]
- Taguchi, T. Emerging roles of recycling endosomes. J. Biochem. 2013, 153, 505–510. [Google Scholar] [CrossRef]
- Cheng, S.B.; Filardo, E.J. trans-Golgi Network (TGN) as a regulatory node for beta1-adrenergic receptor (beta1AR) down-modulation and recycling. J. Biol. Chem. 2012, 287, 14178–14191. [Google Scholar] [CrossRef]
- Terrillon, S.; Durroux, T.; Mouillac, B.; Breit, A.; Ayoub, M.A.; Taulan, M.; Jockers, C.; Barberis, R.; Bouvier, M. Oxytocin and vasopressin V1a and V2 receptors form constitutive homo- and heterodimers during biosynthesis. Mol. Endocrinol. 2003, 17, 677–691. [Google Scholar] [CrossRef]
- Tadagaki, K.; Tudor, D.; Gbahou, F.; Tschische, P.; Waldhoer, M.; Bomsel, M.; Jockers, R.; Kamal, M. Human cytomegalovirus-encoded UL33 and UL78 heteromerize with host CCR5 and CXCR4 impairing their HIV coreceptor activity. Blood 2012, 119, 4908–4918. [Google Scholar] [CrossRef]
- Zoncu, R.; Perera, R.M.; Balkin, D.M.; Pirruccello, M.; Toomre, D.; De, C.P. A phosphoinositide switch controls the maturation and signaling properties of APPL endosomes. Cell 2009, 136, 1110–1121. [Google Scholar] [CrossRef]
- Adobe Photoshop CS2; Adobe Systems Incorporated: San Jose, CA, USA, 2003.
- Tischer, B.K.; Smith, G.A.; Osterrieder, N. En passant mutagenesis: A two step markerless red recombination system Methods. Mol. Biol. 2010, 634, 421–430. [Google Scholar]
- ABI PRISM® 7500 Sequence Detection System; SDS 1.4 Software Package; Applied Biosystems: Foster City, CA, USA, 2001.
- Andreoni, M.; Faircloth, M.; Vugler, L.; Britt, W.J. A rapid microneutralization assay for the measurement of neutralizing antibody reactive with human cytomegalovirus. J. Virol. Methods 1989, 23, 157–167. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Niemann, I.; Reichel, A.; Stamminger, T. Intracellular Trafficking of the Human Cytomegalovirus-Encoded 7-trans-Membrane Protein Homologs pUS27 and pUL78 during Viral Infection: A Comparative Analysis. Viruses 2014, 6, 661-682. https://doi.org/10.3390/v6020661
Niemann I, Reichel A, Stamminger T. Intracellular Trafficking of the Human Cytomegalovirus-Encoded 7-trans-Membrane Protein Homologs pUS27 and pUL78 during Viral Infection: A Comparative Analysis. Viruses. 2014; 6(2):661-682. https://doi.org/10.3390/v6020661
Chicago/Turabian StyleNiemann, Ina, Anna Reichel, and Thomas Stamminger. 2014. "Intracellular Trafficking of the Human Cytomegalovirus-Encoded 7-trans-Membrane Protein Homologs pUS27 and pUL78 during Viral Infection: A Comparative Analysis" Viruses 6, no. 2: 661-682. https://doi.org/10.3390/v6020661