2.1. Transcriptional Analyses of GP129, 131 and 133
Initially, transcription from the GPCMV GP129, 131, and 133 locus was examined by reverse−transcriptase PCR (RT-PCR). Primer design was based upon published primer sequences reported by Inoue [
17]. Primers used for these studies are summarized in
Table 2. Previous reports indicated that these genes are encoded by single mRNAs, and the GP129 and GP131 RNAs, but not the GP133 RNA, are spliced (
Figure 1A). To confirm and extend these findings, RNA was purified from GPCMV-infected GPL cells at 8, 12, 24, and 48 hours post-inoculation. A 48-hour time point was also collected in the presence of phosphonoacetic acid (PAA), 300 µg/mL. The viral stock used was P1-passaged GPCMV Strain 22122 purchased directly from the American Type Culture Collection (ATCC; Manassas, VA, USA). As an additional control, cells were inoculated with vAM403, an eGFP-expressing recombinant GPCMV [
19] known to lack the 1.6 kb region containing the GP129-133 locus [
16]. The RT-PCR products predicted for each gene were: 540 base pairs (bp) for GP129 transcript; 515 bp for GP131; and 384 bp for GP133. Transcripts specific for GP131 and GP133 were apparent as early as 12 hours post-infection (
Figure 1B,C). A GP129-specific transcript could be faintly detected as soon as 12 hours post-infection, and was abundantly present at 24 hours post−infection (
Figure 1A). These data differ from those reported by Inoue and colleagues [
17], who did not note any GP129-specific transcript until ~48 hours post−infection, although this analysis was performed by northern blot, which may have been less sensitive than RT-PCR. Similar to Inoue’s findings, we noted that the spliced GP129 transcript was not identified in the presence of PAA, consistent with its classification as a late gene. However, we noted transcripts corresponding to the spliced GP131 mRNA as soon as 12 hours post-infection and continuing at high levels throughout 48 hours post-infection even in the presence of PAA (
Figure 1B), suggesting that GP131 is an early gene, in contrast to the previous report by Inoue that suggested (based on northern blot analysis) that both GP129 and GP131 were late genes [
17].
Table 2.
Primers used in GP129, 131 and 133 Cloning and Protein Expression Studies.
Table 2.
Primers used in GP129, 131 and 133 Cloning and Protein Expression Studies.
| ORF | Primer Sequences |
|---|
| GP129 (Baculovirus and Transcript Analysis) | 5' – TACGCTGCAGAATGCGTGTTATTGTT – 3' (F) |
| 5' – TACGAGATCTTTACTTCCCGTTACC – 3' (R) |
| GP129 GST (R1) | 5' – TACGGGATCCTATACCCGTCCCGGTATCTTTG – 3' (F) |
| 5' – TACGCTCGAGTTAAGTATTCCCACATCGTACTAATC – 3' (R) |
| GP129 GST (R2) | 5' – TACGGGATCCTCGCGGCAAGAACTCCAT – 3' (F) |
| 5' – TACGCTCGAGTTAACGGTAGGTCACCCCCAAG – 3' (R) |
| GP129 GST (R3) | 5' – TACGGGATCCAACGGTTTATTATGCACCTTTC – 3' (F) |
| 5' – TACGCTCGAGTTACTTCCCGTTACCATCGAC – 3' (R) |
| GP131 (Baculovirus) | 5' – TACGAATTCATGATGAAACGATAT – 3' (F) |
| 5' – TACGCTGCAGTTATCACGTCCAGTT – 3' (R) |
| GP131 (Transcript Analysis) | 5' – ATAATGATGAAACGATAT – 3' (F) |
| 5' – TTATCACGTCCAGTTCCA – 3' (R) |
| GP131 GST (R1) | 5' – GATAGGATCCTTTTACGCCTCGTTCGGA – 3' (F) |
| 5' – AATACTCGAGGTTCGTCAGGGTCAGGAC – 3' (R) |
| GP131 GST (R2) | 5' – GATAGGATCCCGCCGAATAGATTACGGA – 3' (F) |
| 5' – AATACTCGAGCCACAAGAAGGACGAATC – 3' (R) |
| GP131 GST (R3) | 5' – CGTGGGATCCTGGCATTATACGATACGG – 3' (F) |
| 5' – AATACTCGAGCAGGCAAGCGATAGAATC – 3' (R) |
| GP133 (Baculovirus) | 5' – TACGCTGCAGTATGTTTTGGCGTCTTGTA – 3' (F) |
| 5' – TACGAGATCTTTATGCTCTGTCTATGC – 3' (R) |
| GP133 (Transcript Analysis) | 5' – TATGTTTTGGCGTCTTGTA – 3' (F) |
| 5' – TTATGCTCTGTCTATGC – 3' (R) |
| GP133 GST (R1) | 5' – GATCGGATCCACAAGAGTTAAGAAAGAAAACCAACTG – 3' (F) |
| 5' – GATCCTCGAGTTAGGAGTCCGCTAACGTATG – 3' (R) |
| GP133 GST (R2) | 5' – GATCGGATCCAGGGGACGTTACAGGAAAGG – 3' (F) |
| 5' – GATCCTCGAGTTAAGTGCTTTGTTGAATAGAAATACG – 3' (R) |
| GAPDH | 5' – ATCTCATCGTATTTGGCCGGT – 3' (F) |
| 5' – AATGGGAAGCTCACAGGTATGG – 3' (R) |
Figure 1.
Transcription of GP129, 131 and 133 as assessed by reverse-transcriptase PCR (RT-PCR). RNA from ATCC GPCMV-infected cells (Lanes 2, 4, 6, 8, 10) and vAM403-infected cells (Lanes 3, 5, 7, 9 and 11) was purified at 8 hours (Lanes 2, 3), 12 hours (Lanes 4, 5), 24 hours (Lanes 6, 7) and 48 hours (Lanes 8, 9) post-infection. A 48-hour time point was also collected in the presence of phosphonoacetic acid (PAA) (Lanes 10, 11). Lane 1 represents RT-PCR of RNA from uninfected cells. Electrophoresis was performed on a 1.5% agarose gel. Molecular weights markers are the 1 Kb-Opti-DNA marker from ABM (Applied Biological Materials Inc., Richmond, BC, Canada). (a) GP129-specific transcripts. Predicted splice product of 540 nt is noted (arrow), but is not present in infected cells incubated with PAA. (b) GP131-specific transcripts. Predicted splice product of 515 nt is noted (arrow). (c) GP133-specific transcripts demonstrating no evidence of splicing. Predicted 384 nt product is noted (arrow). (d) GAPDH control.
Figure 1.
Transcription of GP129, 131 and 133 as assessed by reverse-transcriptase PCR (RT-PCR). RNA from ATCC GPCMV-infected cells (Lanes 2, 4, 6, 8, 10) and vAM403-infected cells (Lanes 3, 5, 7, 9 and 11) was purified at 8 hours (Lanes 2, 3), 12 hours (Lanes 4, 5), 24 hours (Lanes 6, 7) and 48 hours (Lanes 8, 9) post-infection. A 48-hour time point was also collected in the presence of phosphonoacetic acid (PAA) (Lanes 10, 11). Lane 1 represents RT-PCR of RNA from uninfected cells. Electrophoresis was performed on a 1.5% agarose gel. Molecular weights markers are the 1 Kb-Opti-DNA marker from ABM (Applied Biological Materials Inc., Richmond, BC, Canada). (a) GP129-specific transcripts. Predicted splice product of 540 nt is noted (arrow), but is not present in infected cells incubated with PAA. (b) GP131-specific transcripts. Predicted splice product of 515 nt is noted (arrow). (c) GP133-specific transcripts demonstrating no evidence of splicing. Predicted 384 nt product is noted (arrow). (d) GAPDH control.
![Viruses 06 00727 g001]()
We saw no evidence of splicing of the GP133 transcript, and this RNA continued to be expressed to high levels through 48 hours post-infection. This transcript was also expressed in the presence of PAA. RNA species larger than we predicted were noted for the GP129 and GP131 transcripts. Absence of any signal in the absence of reverse transcriptase makes the possibility of DNA contamination unlikely. These were presumed to represent larger, unspliced RNAs driven by promoters upstream of the GP129-133 promoter region. RNA purified from vAM403-infected cells was negative for GP129, 131 and 133 RT-PCR products, as expected, since this virus lacks the region of the GPCMV genome encoding these ORFs. Guinea pig GAPDH cDNA was amplified to control for RNA recovery (1D). No bands were noted in the no-RT control (data not shown).
2.2. Mass Spectrometry Analysis
GPCMV virus particles were purified by sequential sucrose [
20] and glycerol-tartrate density gradient centrifugation [
21] as described in
Section 3.2. Particles were examined by coomassie blue stain (
Figure 2a) and transmission electron microscopy (EM;
Figure 2b) to confirm homogeneity. Particles were probed with a monoclonal antibody (moab) to glycoprotein B, IE321, generated in our laboratory (
Figure 2c), to further confirm their integrity. Particles were resolved by SDS PAGE followed by coomassie staining, and bands were excised from the gel for mass spectrometry. Three peptides corresponding to two unique sequences for GP129 were identified from a region of the gel with an approximate molecular weight of 25 kD. Additionally, six peptides corresponding to four unique sequences corresponding to GP131 were identified from a protein band excised from a region with a molecular weight spanning approximately 20–22 kD. Finally, seven peptides from four unique sequences of GP133 were identified from a protein band with an approximate molecular weight of 18 kD. Results are summarized in
Table 3. Mass spectra data are provided in
Appendix Figures A1–A3.
Figure 2.
Characterization of GPCMV viral particles. (a) Coomassie blue stain (5 µg) of purified virus particles by 12% gel SDS-PAGE. (b) Transmission electron microscope microphotograph of purified particles. Negative contrast preparation of purified particles demonstrates enveloped virions (arrows). Dense bodies are also noted. Magnification, 150,000×. (c) Western blot analysis of viral particles probed with IE3-21 monoclonal antibody recognizing GPCMV gB (1:2,000 dilution; arrow).
Figure 2.
Characterization of GPCMV viral particles. (a) Coomassie blue stain (5 µg) of purified virus particles by 12% gel SDS-PAGE. (b) Transmission electron microscope microphotograph of purified particles. Negative contrast preparation of purified particles demonstrates enveloped virions (arrows). Dense bodies are also noted. Magnification, 150,000×. (c) Western blot analysis of viral particles probed with IE3-21 monoclonal antibody recognizing GPCMV gB (1:2,000 dilution; arrow).
Table 3.
Mass spectrometry identification of peptides corresponding to GP129, 131 and 133. Other major virion structural proteins included as controls for mass spectrometry analyses (protein probability calculations performed as described in materials and methods).
Table 3.
Mass spectrometry identification of peptides corresponding to GP129, 131 and 133. Other major virion structural proteins included as controls for mass spectrometry analyses (protein probability calculations performed as described in materials and methods).
| ORF | Unique Spectra | Unique Peptide Sequences | % Coverage | Max Xcorr | Probability |
|---|
| GP 129 | 2 | MPSVQSKPEKPSILGVTYR VDYTVMIPTPHFPR | 18 | 3.37 | 99% |
| GP131 | 6 | RIDYGSTGTAASTLPSLTSLR TYFGDRDSSFLWHYTIRT DCDVYVTSR HPADSIACLL | 29 | 5.01 | 99.9% |
| GP133 | 7 | EDEGIDTWWLGGVTDNTR VKKENQLAHYILK KGLVSIDGIR TIVLTHHR | 39 | 4.51 | 99.9% |
| GP83 | 74 | 61 | 59 | 62 | 100% |
| GP25 | 78 | 55 | 77 | 5.9 | 100% |
| gB | 51 | 37 | 31 | 5.1 | 100% |
2.3. Characterization of GP129 and 131 Proteins
Initially, an anti-peptide rabbit antiserum was generated targeting a GP131-specific peptide sequence, NH
2-CYYPSTPIPKSFVKHVDTTRSLPE-COOH, conjugated with keyhole limpet hemocyanin (see
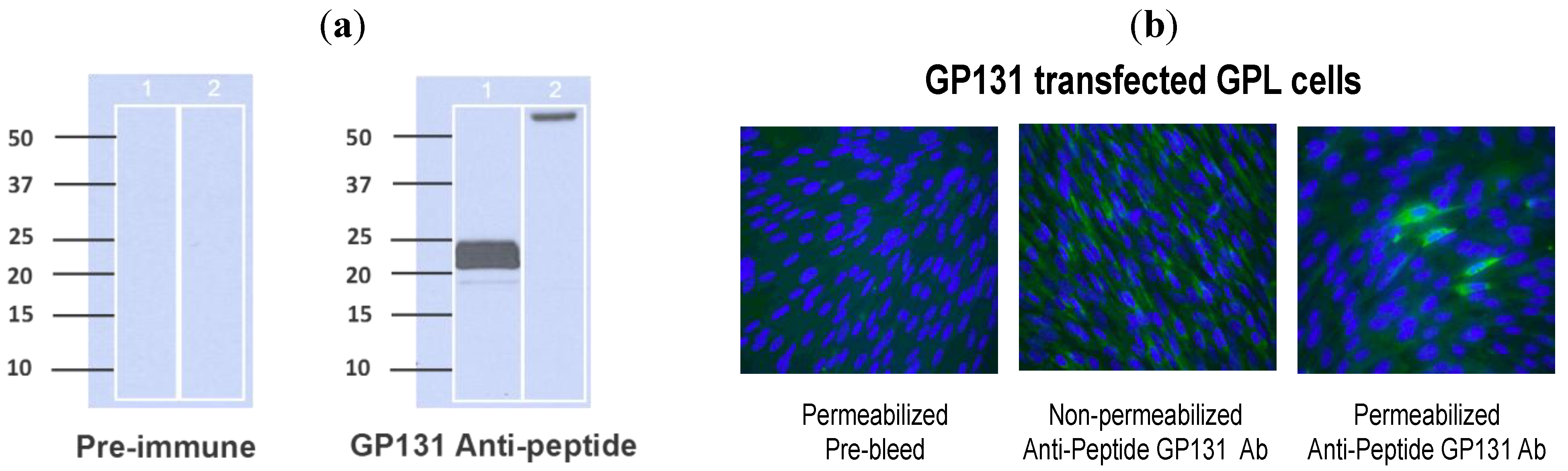
Section 3 for details on generation of antibody). This antibody was used in western blot assay to examine purified virions (purified from passage 1 ATCC Strain 22122 GPCMV), as well as infected cell lysates, for expression of the GP131 protein. Immune sera consistently demonstrated the appearance of a broadly migrating band at approximately 22 kDa. In some experiments, the band appeared to separate into two discrete bands at ~20 and ~22 kDa (
Figure 3A). To confirm the specificity of this response for GP131, westerns were also performed using a recombinant, eGFP-expressing GPCMV, vAM403, known to lack the 1.6 kb region spanning the GP129-133 locus. Western assay targeting vAM403 proteins using the GP131 rabbit polyclonal antibody did not demonstrate presence of this band in purified lysates from cells infected with this virus, as expected (
Figure 3A, Lane 2).
The GP131-specific antibody was next examined in an immunofluorescence assay using cells transfected with a GP131 expression plasmid, pKTS 789. Immune, (but not pre-immune) sera identified a protein in GP131-transfected cells, but only under conditions of permeabilization (
Figure 3B). This result is compatible with the known requirement of GP131 to be co-expressed with gH in order for cell surface localization.
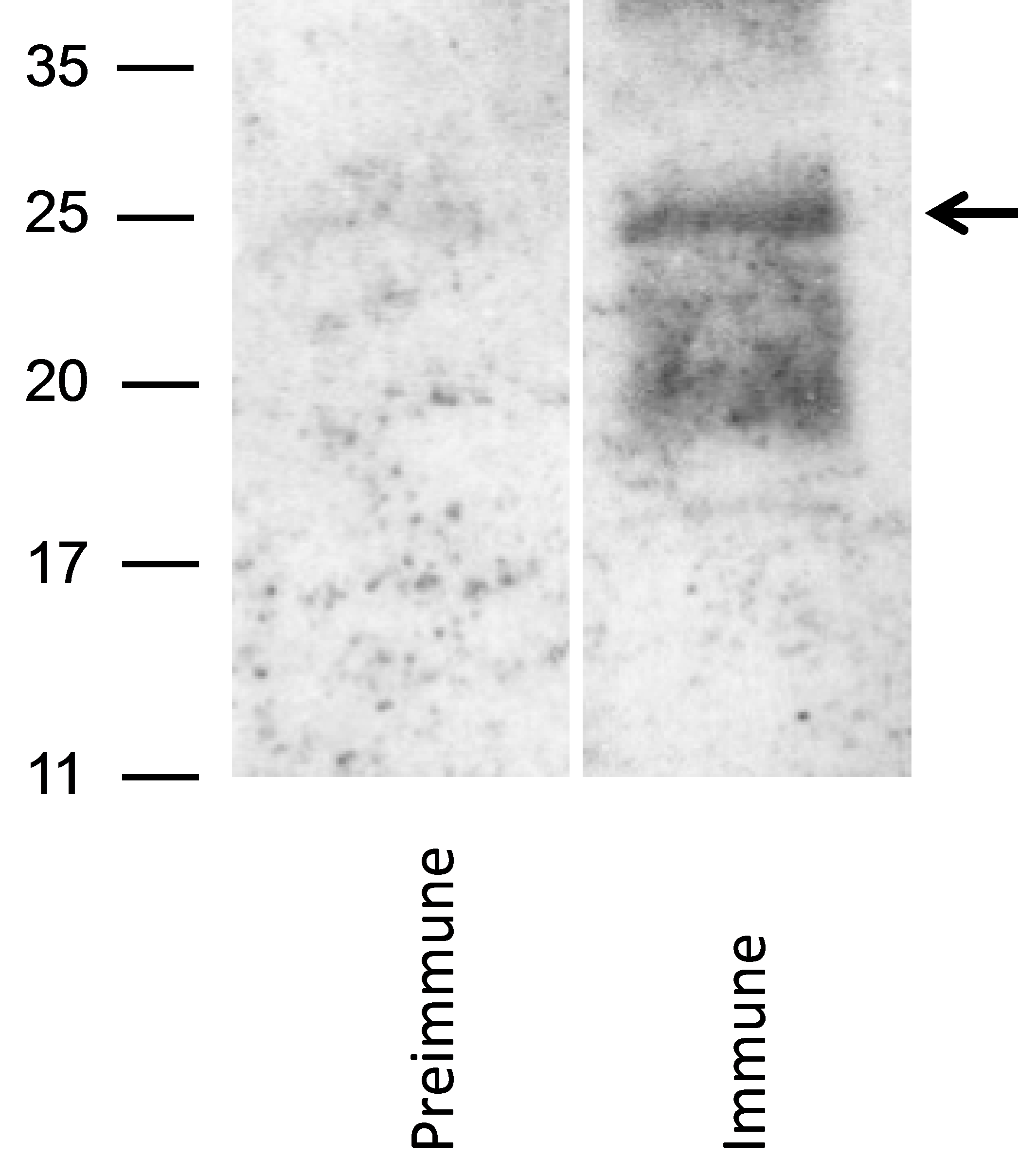
To identify GP129 in virions, a GP129-specific antibody was next generated following vaccination of naïve guinea pigs with a GST fusion protein, designated GST_R3_GP129 (described in
Section 3). Three guinea pigs were immunized with fusion protein. All animals generated ELISA responses to GST_R3_129 (data not shown). The GST_R3_GP129 immune, but not pre-immune, sera identified a broadly migrating band in purified GPCMV virions (
Figure 4). This result suggested that the GP129 protein is expressed in virions, and migrates broadly in the ~25 kDa range by SDS-PAGE. These data confirmed the identification of GP129-specific peptides present in the mass spectrometry analyses of virus particles.
Figure 3.
Characterization of GP131 protein. (a) Western blot analysis using anti-peptide GP131 antibody. Virion particles were probed with either pre-immune sera (left panel) or immune sera (right panel) following purification of virus particles from cells infected either with ATCC GPCMV (Lane 1 in each panel) or vAM403 (Lane 2 in each panel). A band of approximately 20–22 kDa was noted with immune, but not pre-immune sera. On shorter exposure, this band appeared as a doublet. (b) Effect of cell permeabilization on detection of GP131. GPL cells were transfected with a GP131 expression plasmid and immunofluorescence performed with anti-GP131 antibody. Little to no protein expression was detected in non−permeabilized cells (middle panel), but protein expression was readily detected following permeabilization (right panel). Left panel demonstrates pre-immune antibody control.
Figure 3.
Characterization of GP131 protein. (a) Western blot analysis using anti-peptide GP131 antibody. Virion particles were probed with either pre-immune sera (left panel) or immune sera (right panel) following purification of virus particles from cells infected either with ATCC GPCMV (Lane 1 in each panel) or vAM403 (Lane 2 in each panel). A band of approximately 20–22 kDa was noted with immune, but not pre-immune sera. On shorter exposure, this band appeared as a doublet. (b) Effect of cell permeabilization on detection of GP131. GPL cells were transfected with a GP131 expression plasmid and immunofluorescence performed with anti-GP131 antibody. Little to no protein expression was detected in non−permeabilized cells (middle panel), but protein expression was readily detected following permeabilization (right panel). Left panel demonstrates pre-immune antibody control.
Figure 4.
Identification of GP129 in virus particles. Western blot analysis using an anti-GST/GP129 fusion protein derived antibody. Following electrophoresis with 12% Bis-Tris gel, western transfer was performed and virion particles were probed (1:200 dilution) with either pre-immune sera (left panel) or immune sera (right panel). GST_R3_129 immune (but not pre-immune) antisera identified a broadly migrating band with MW of the predominate species at ~25 kDa (arrow). Band was not identified in blots of vAM403 (lacking GP129-133 locus) virus particles probed with identical antibodies (data not shown).
Figure 4.
Identification of GP129 in virus particles. Western blot analysis using an anti-GST/GP129 fusion protein derived antibody. Following electrophoresis with 12% Bis-Tris gel, western transfer was performed and virion particles were probed (1:200 dilution) with either pre-immune sera (left panel) or immune sera (right panel). GST_R3_129 immune (but not pre-immune) antisera identified a broadly migrating band with MW of the predominate species at ~25 kDa (arrow). Band was not identified in blots of vAM403 (lacking GP129-133 locus) virus particles probed with identical antibodies (data not shown).
2.4. Characterization of Guinea Pig Antibody Reactivity to Recombinant-Expressed GP129, 131, and 133
To examine the guinea pig immune response to GP129, 131, and 133, a series of GST fusion proteins were generated, corresponding to predicted immunogenic domains in the respective proteins. Primers used in the generation of these fusion proteins are indicated in
Table 2. Immune responses in guinea pig convalescent sera from infected animals were readily demonstrable to GP129 fusion proteins. Three domains of GP129 (GP129 R1, R2, and R3;
Figure 5) were cloned as GST fusion proteins and western blots probed with anti-GPCMV antisera obtained from experimentally infected animals. The R3 domain (
Figure 6A) was the domain most highly immunoreactive with convalescent anti-GPCMV sera from infected animals. This region of the GP129 contains a cysteine residue highly conserved with HCMV and RhCMV UL128 (the homolog of GP129). Although interactions between UL128 and the other constituents of the pentameric complex are not believe to involve disulfide bond formation, this region of the UL128 protein in HCMV is known to be the target of neutralizing monoclonal antibodies [
22,
23]. When GP129 was expressed in recombinant baculovirus, purified recombinant protein was also immunoreactive with immune sera from animals challenged with salivary gland-adapted GPCMV (
Figure 6B). Pre-immune sera failed to recognize the recombinant protein. These analyses strongly suggest that the GP129 protein is a target of the guinea pig antibody response following GPCMV infection.
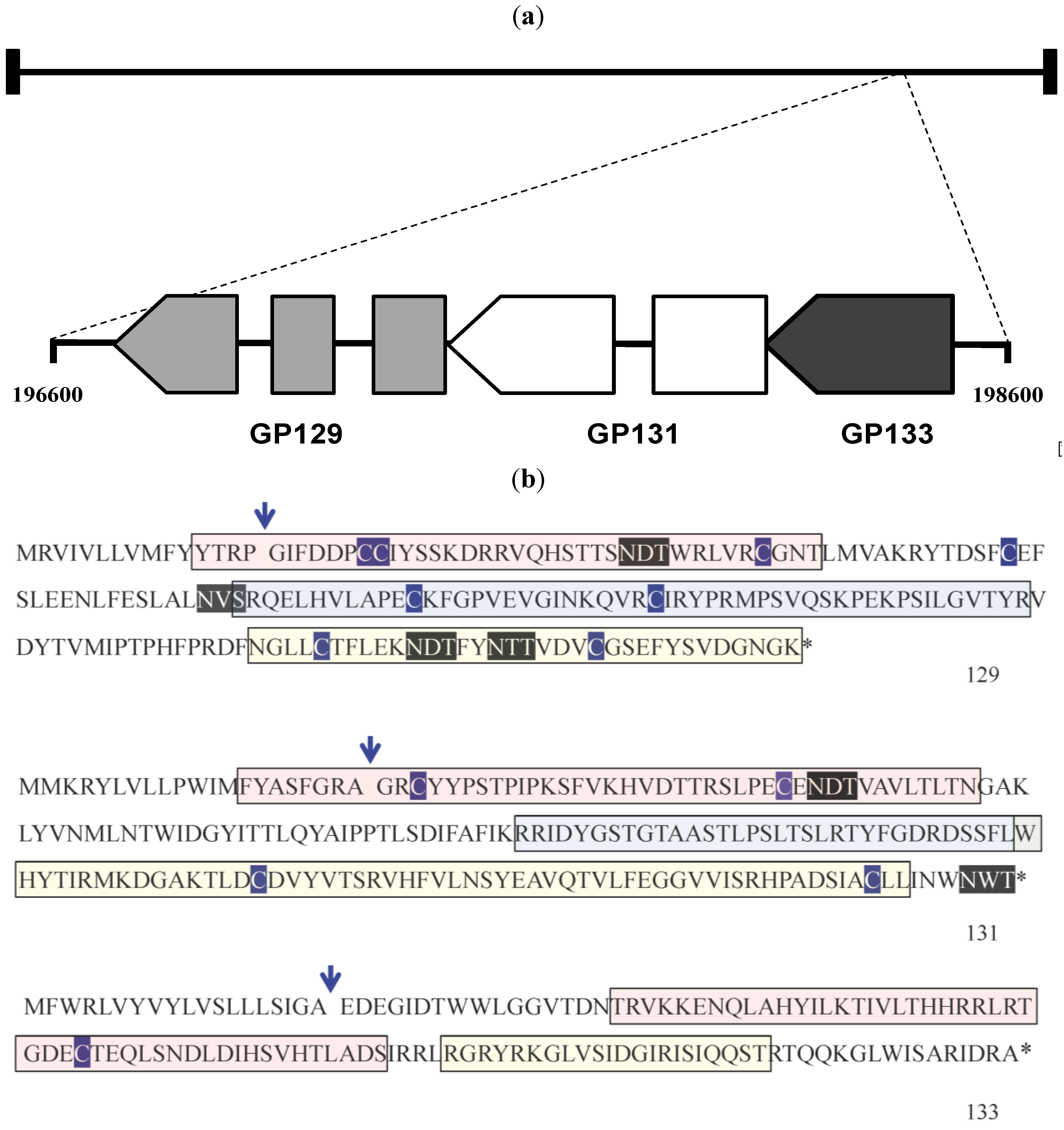
Figure 5.
Subcloning of GP129, 131, 133 proteins. (
a) Schematic map of the GPCMV GP129, 131 and 133 ORFs, patterned after [
18]. These ORFs map to the rightward end of the GPCMV genome. Genome coordinates of this region are indicated (based on sequence Genbank KC503762 [
24]). In contrast to HCMV, the GP131 (UL130) homolog is spliced, but the GP133 (UL131 homolog) is not. (
b) GPCMV GP129, GP131, and GP133 ORFs. Predicted van Heinje signal sequence cleavage site is indicated by arrow. Cysteine residues are outlined in dark blue. Potential glycosylation sites (N-X-T and N-X-S) are shown in black outline. Regions of ORFs subcloned as GST fusion proteins are indicated. For GP129 and GP131, pink region represents R1 region; blue, R2 region; yellow, R3. For GP133, pink region is R1 and yellow region R2 (see
Table 2 and text for details).
Figure 5.
Subcloning of GP129, 131, 133 proteins. (
a) Schematic map of the GPCMV GP129, 131 and 133 ORFs, patterned after [
18]. These ORFs map to the rightward end of the GPCMV genome. Genome coordinates of this region are indicated (based on sequence Genbank KC503762 [
24]). In contrast to HCMV, the GP131 (UL130) homolog is spliced, but the GP133 (UL131 homolog) is not. (
b) GPCMV GP129, GP131, and GP133 ORFs. Predicted van Heinje signal sequence cleavage site is indicated by arrow. Cysteine residues are outlined in dark blue. Potential glycosylation sites (N-X-T and N-X-S) are shown in black outline. Regions of ORFs subcloned as GST fusion proteins are indicated. For GP129 and GP131, pink region represents R1 region; blue, R2 region; yellow, R3. For GP133, pink region is R1 and yellow region R2 (see
Table 2 and text for details).
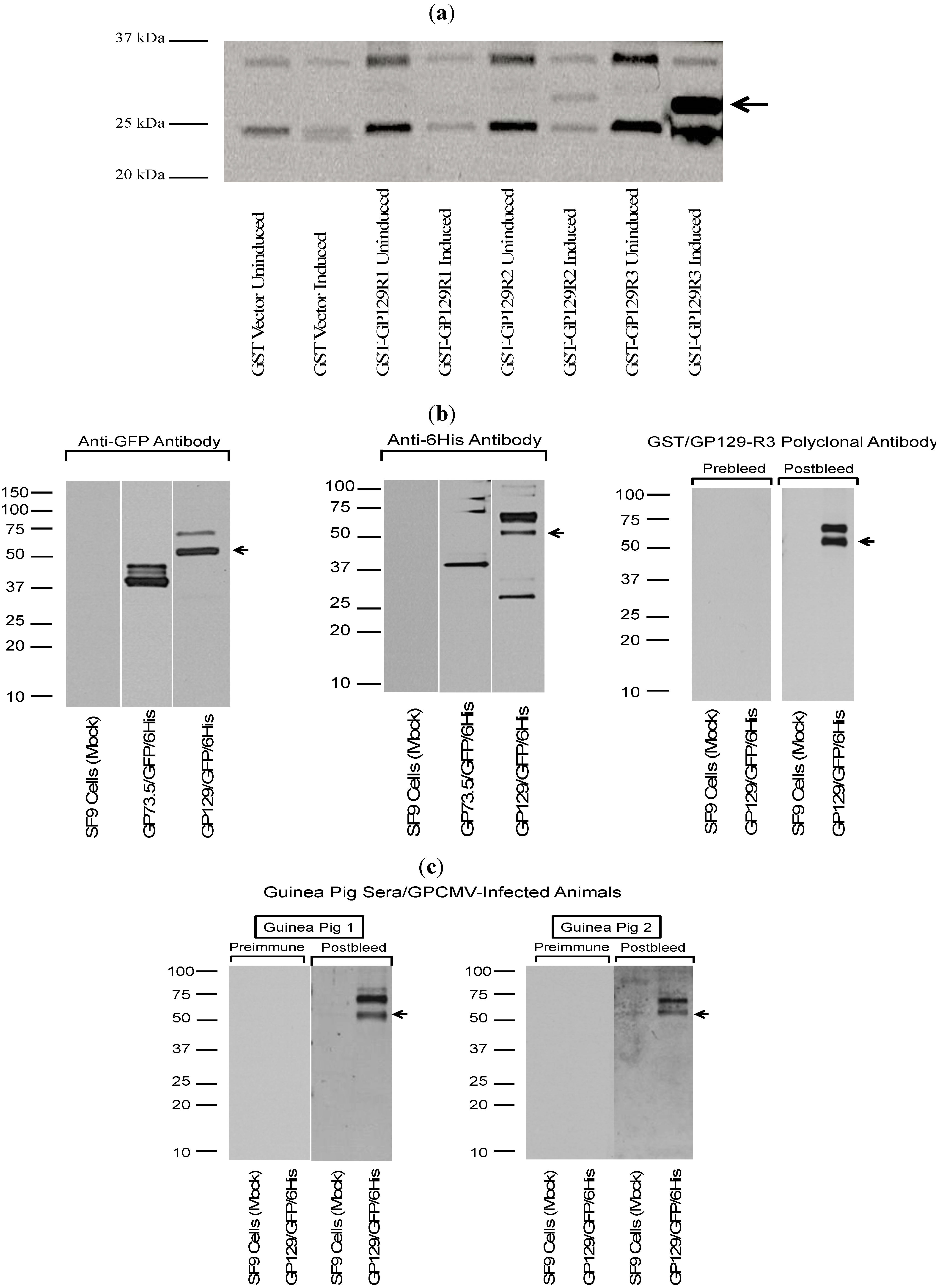
Figure 6.
Antibodies from GPCMV-seropositive guinea pigs react with recombinant forms of GP129. (
a) Analysis of GP129/GST fusion proteins. Three GP129 domains (
Figure 5b) were cloned in-frame with GST using vector pGEX 6-P as described in materials and methods. Constructs evaluated by western blot included GST only; GST-GP129R1 fusion; GST-GP129R2 fusion; and GST-GP129R3 fusion, without IPTG induction (uninduced) and following IPTG induction (induced). Anti-GPCMV antisera from immune, experimentally infected guinea pigs demonstrated strongest immunoreactivity with GST−GP129R3 fusion protein. In some experiments, some signal could be observed with GP129R2 fusion protein. Data shown is a representative result from an experimentally infected animal. All fusion proteins expressed to high levels following IPTG induction as demonstrated by western analysis with anti-GST monoclonal antibody (data not shown). No background reactivity was noted with serum from GPCMV-seronegative animals or with pre-immune sera from experimentally infected guinea pigs (data not shown). (
b) Western blot analysis of recombinant baculovirus-expressed GP129. The GP129 was cloned in-frame with GFP and a 6-His tag. This resulted in a fusion protein of ~55 kDa, made up of ~6 kDa from the 6-His tag and vector sequences; ~28 kDa from the GFP sequence; and ~21 kDa from the GP129 sequences. Purified recombinant protein was subjected to SDS-PAGE and western assays performed as described in materials and methods section. For some western blots, a control baculovirus expressing a recombinant form of the GPCMV GP73.5 glycoprotein was also evaluated. Top panel, both the anti−GFP and anti-6 His antibodies were immunoreactive with GP73.5 (~37 kDa) and GP129 (~55 kDa) fusion proteins, as expected. A higher MW band was noted at ~65–70 kDa, possibly representing a glycosylated variant of the baculovirus-expressed GP129 fusion protein. Sera, raised against the GST-GP129-R3 fusion protein (
Figure 5a), was also used to probe baculovirus-expressed GP129 as a control. Immune, but not preimmune, sera (1:200 dilution) from a guinea pig immunized with the GST fusion protein were reactive with the baculovirus expressed protein, but not proteins in uninfected SF9 cells. Again, both the 55 kDa fusion protein as well as a larger species of 65–70 kDa, were identified. (
c) Sera from experimentally infected animals were immunoreactive with the GP129−baulovirus fusion protein. Results shown are representative pre- and post-infection sera (1:200 dilution) from two GPCMV-infected guinea pigs (GP1; GP2) demonstrating immunoreactivity with bacolovirus-GP129 fusion protein (arrow), but not a pre-immune guinea pig serum (NEG).
Figure 6.
Antibodies from GPCMV-seropositive guinea pigs react with recombinant forms of GP129. (
a) Analysis of GP129/GST fusion proteins. Three GP129 domains (
Figure 5b) were cloned in-frame with GST using vector pGEX 6-P as described in materials and methods. Constructs evaluated by western blot included GST only; GST-GP129R1 fusion; GST-GP129R2 fusion; and GST-GP129R3 fusion, without IPTG induction (uninduced) and following IPTG induction (induced). Anti-GPCMV antisera from immune, experimentally infected guinea pigs demonstrated strongest immunoreactivity with GST−GP129R3 fusion protein. In some experiments, some signal could be observed with GP129R2 fusion protein. Data shown is a representative result from an experimentally infected animal. All fusion proteins expressed to high levels following IPTG induction as demonstrated by western analysis with anti-GST monoclonal antibody (data not shown). No background reactivity was noted with serum from GPCMV-seronegative animals or with pre-immune sera from experimentally infected guinea pigs (data not shown). (
b) Western blot analysis of recombinant baculovirus-expressed GP129. The GP129 was cloned in-frame with GFP and a 6-His tag. This resulted in a fusion protein of ~55 kDa, made up of ~6 kDa from the 6-His tag and vector sequences; ~28 kDa from the GFP sequence; and ~21 kDa from the GP129 sequences. Purified recombinant protein was subjected to SDS-PAGE and western assays performed as described in materials and methods section. For some western blots, a control baculovirus expressing a recombinant form of the GPCMV GP73.5 glycoprotein was also evaluated. Top panel, both the anti−GFP and anti-6 His antibodies were immunoreactive with GP73.5 (~37 kDa) and GP129 (~55 kDa) fusion proteins, as expected. A higher MW band was noted at ~65–70 kDa, possibly representing a glycosylated variant of the baculovirus-expressed GP129 fusion protein. Sera, raised against the GST-GP129-R3 fusion protein (
Figure 5a), was also used to probe baculovirus-expressed GP129 as a control. Immune, but not preimmune, sera (1:200 dilution) from a guinea pig immunized with the GST fusion protein were reactive with the baculovirus expressed protein, but not proteins in uninfected SF9 cells. Again, both the 55 kDa fusion protein as well as a larger species of 65–70 kDa, were identified. (
c) Sera from experimentally infected animals were immunoreactive with the GP129−baulovirus fusion protein. Results shown are representative pre- and post-infection sera (1:200 dilution) from two GPCMV-infected guinea pigs (GP1; GP2) demonstrating immunoreactivity with bacolovirus-GP129 fusion protein (arrow), but not a pre-immune guinea pig serum (NEG).
![Viruses 06 00727 g006]()
The guinea pig antibody response to GP131 was next evaluated, using recombinant GP131 in western blot assay along with sera from GPCMV-seropositive guinea pigs. As was the approach for GP129, the GP131 coding sequence was first subcloned as three GST fusion proteins, spanning potentially immunogenic domains of the GP131 protein (GP131 R1, R2 and R3). The GST fusion proteins were purified and, following western transfer, blots were probed with sera from experimentally infected GPCMV-seropositive animals (1:200 dilution; details as noted in
Figure 6). A representative result from a GPCMV-infected animal is shown. These analyses demonstrated (
Figure 7) that convalescent, but not pre-immune, sera from GPCMV-infected guinea pigs recognized the GST fusion protein corresponding to domains R1, R2 and R3 of the GP131 ORF.
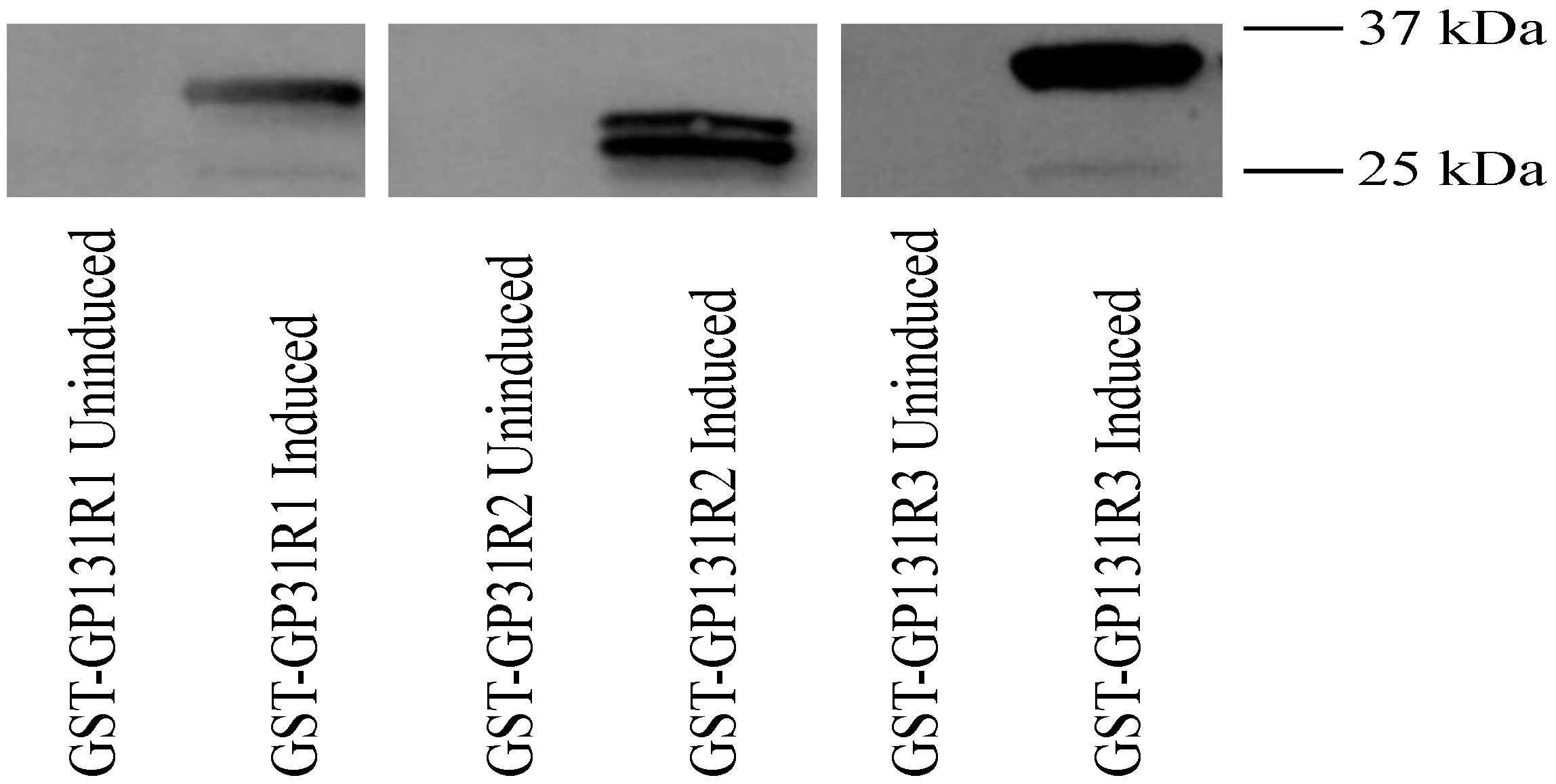
Figure 7.
Characterization of antibody responses of GPCMV-infected guinea pigs to recombinant GP131 proteins. Western blots examining reactivity of anti-GPCMV antisera with uninduced and IPTG-induced GST-GP131R1, R2, and R3 fusion proteins. GP133R1 and GP131R3 were most consistently reactive with post-infection antisera (1:200 dilution), but not preimmune sera (data not shown). Representative result from an infected animal is shown. 37 and 25 kDa markers are indicated in right side of figure.
Figure 7.
Characterization of antibody responses of GPCMV-infected guinea pigs to recombinant GP131 proteins. Western blots examining reactivity of anti-GPCMV antisera with uninduced and IPTG-induced GST-GP131R1, R2, and R3 fusion proteins. GP133R1 and GP131R3 were most consistently reactive with post-infection antisera (1:200 dilution), but not preimmune sera (data not shown). Representative result from an infected animal is shown. 37 and 25 kDa markers are indicated in right side of figure.
Finally, antibody responses to recombinant GP133 proteins were assessed. GP133 coding sequences were subcloned as depicted in
Figure 5b. GST fusion constructs corresponding to Region 1 (predicted MW of 31.8 kDa) and Region 2 (predicted MW of 28.5 kDa) were evaluated, as well as a full-length GST fusion construct spanning both domains (R1/R2 construct; predicted MW of 34.8). These fusion protein clones were generated using primers as depicted in
Table 2. The GST fusion proteins were purified from insoluble inclusion bodies and, following western transfer, blots were probed with convalescent sera from GPCMV seropositive animals that had been infected subcutaneously with SG virus. Results were confirmed with three independently GPCMV-infected guinea pigs;
Figure 8 demonstrates representative results from one of these animals. These analyses demonstrated that convalescent, but not pre-immune (data not shown), sera from GPCMV-infected guinea pigs recognized the proteins corresponding to both the R1 and the R1/R2 regions of GP133 expressed as GST fusions, but not the R2 region of the GP133 protein. Reactivity of sera from infected animals, in contrast, was not noted with recombinant GP133 expressed as a GFP/6-His fusion protein in recombinant baculovirus (data not shown). This may have been due to loss of reactive epitopes engendered by fusion of the GFP and 6-His tags on the amino-terminal coding sequences of the recombinant GP133 protein. Further experiments will be necessary to resolve this question.
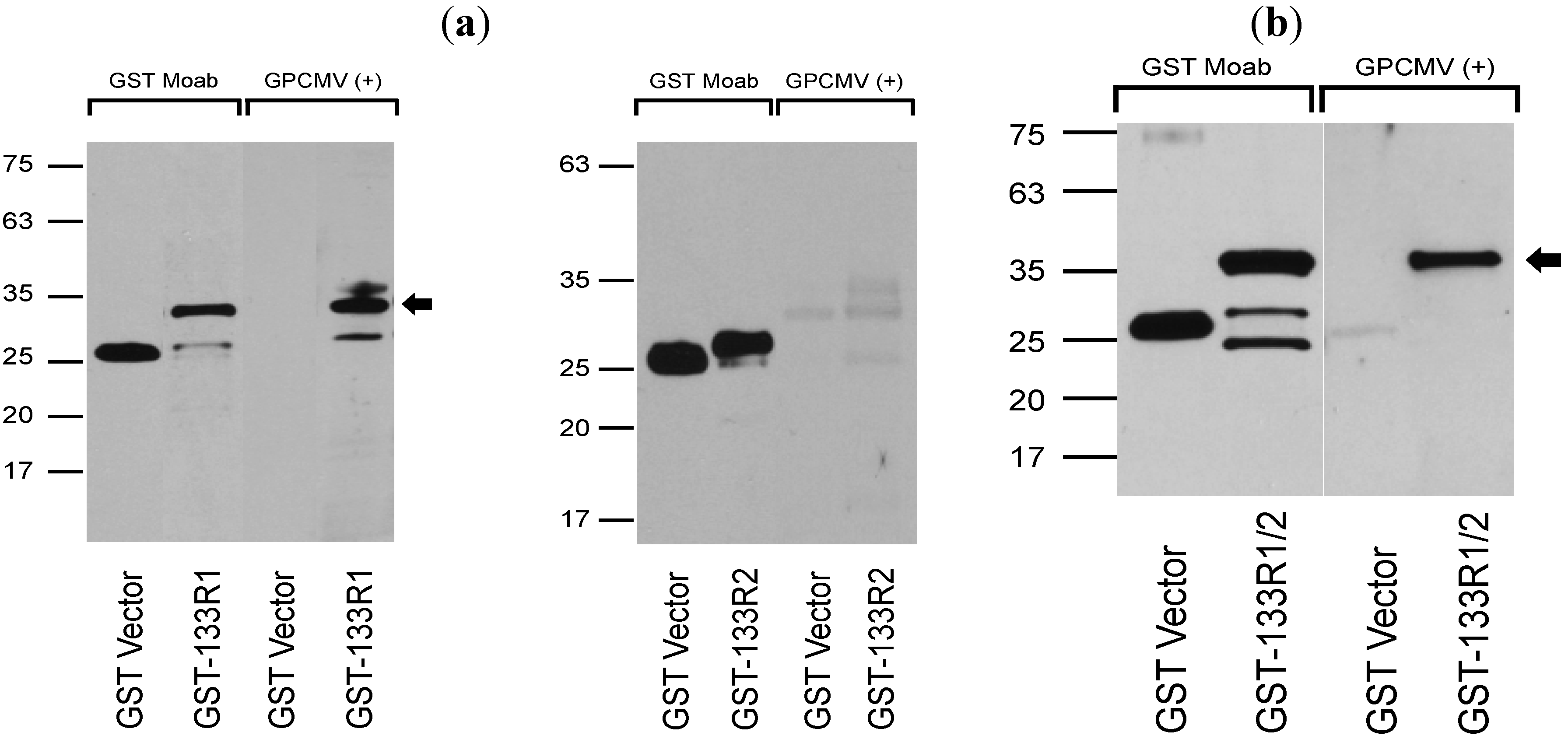
Figure 8.
Characterization of antibody responses from GPCMV-immune guinea pigs to recombinant GP133 protein. (a) Antisera from a GPCMV-infected animal [GPCMV (+)] demonstrates immunoreactivity with the GST-GP133_R1_fusion protein (arrow) but not with purified GST protein. Sera is not reactive with GST-R2 fusion protein (right panel). Anti−GST moab (1:5,000 dilution) demonstrated reactivity both with the GST-GP133R1, R2, and R1/2 fusion proteins, as well as the purified GST protein. (b) Western analysis of GST−GP133R1/2 fusion protein. Both GST (GST vector) and GST-GP133R1/2 fusion protein are reactive with anti-GST moab; however, only sera from GPCMV-infected animal [GPCMV(+), 1:200 dilution] was reactive with the GST-R1/2 fusion protein. For both experiments, pre-immune sera from GPCMV-infected guinea pig was unreactive with all GP133/GST fusion proteins and with the GST parent protein itself (data not shown).
Figure 8.
Characterization of antibody responses from GPCMV-immune guinea pigs to recombinant GP133 protein. (a) Antisera from a GPCMV-infected animal [GPCMV (+)] demonstrates immunoreactivity with the GST-GP133_R1_fusion protein (arrow) but not with purified GST protein. Sera is not reactive with GST-R2 fusion protein (right panel). Anti−GST moab (1:5,000 dilution) demonstrated reactivity both with the GST-GP133R1, R2, and R1/2 fusion proteins, as well as the purified GST protein. (b) Western analysis of GST−GP133R1/2 fusion protein. Both GST (GST vector) and GST-GP133R1/2 fusion protein are reactive with anti-GST moab; however, only sera from GPCMV-infected animal [GPCMV(+), 1:200 dilution] was reactive with the GST-R1/2 fusion protein. For both experiments, pre-immune sera from GPCMV-infected guinea pig was unreactive with all GP133/GST fusion proteins and with the GST parent protein itself (data not shown).
Table 4 summarizes the immunoreactivity of the various GST fusion constructs (depicted in
Figure 5b) with immune sera from infected guinea pigs post-infection with SG virus. Designations: (−), no immunoreactivity; (+), minimal immunoreactivity; (++), moderate immunoreactivity; (+++) consistent, high-level immunoreactivity.
Table 4.
Summary of reactivity of sera from infected guinea pigs with GST-GP129, 131 and 133 fusion proteins.
Table 4.
Summary of reactivity of sera from infected guinea pigs with GST-GP129, 131 and 133 fusion proteins.
| GST Fusion Protein Domain | Immunoreactivity with Guinea Pig Sera Post-Experimental Infection with GPCMV |
|---|
| GP129-R1 | − |
| GP129-R2 | + |
| GP129-R3 | +++ |
| GP131-R1 | ++ |
| GP131-R2 | + |
| GP131-R3 | +++ |
| GP133-R1 | ++ |
| GP133-R2 | − |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










