In Vitro Evaluation of the Antiviral Activity of the Synthetic Epigallocatechin Gallate Analog-Epigallocatechin Gallate (EGCG) Palmitate against Porcine Reproductive and Respiratory Syndrome Virus

 , ,
, ,
Abstract
:1. Introduction
2. Results
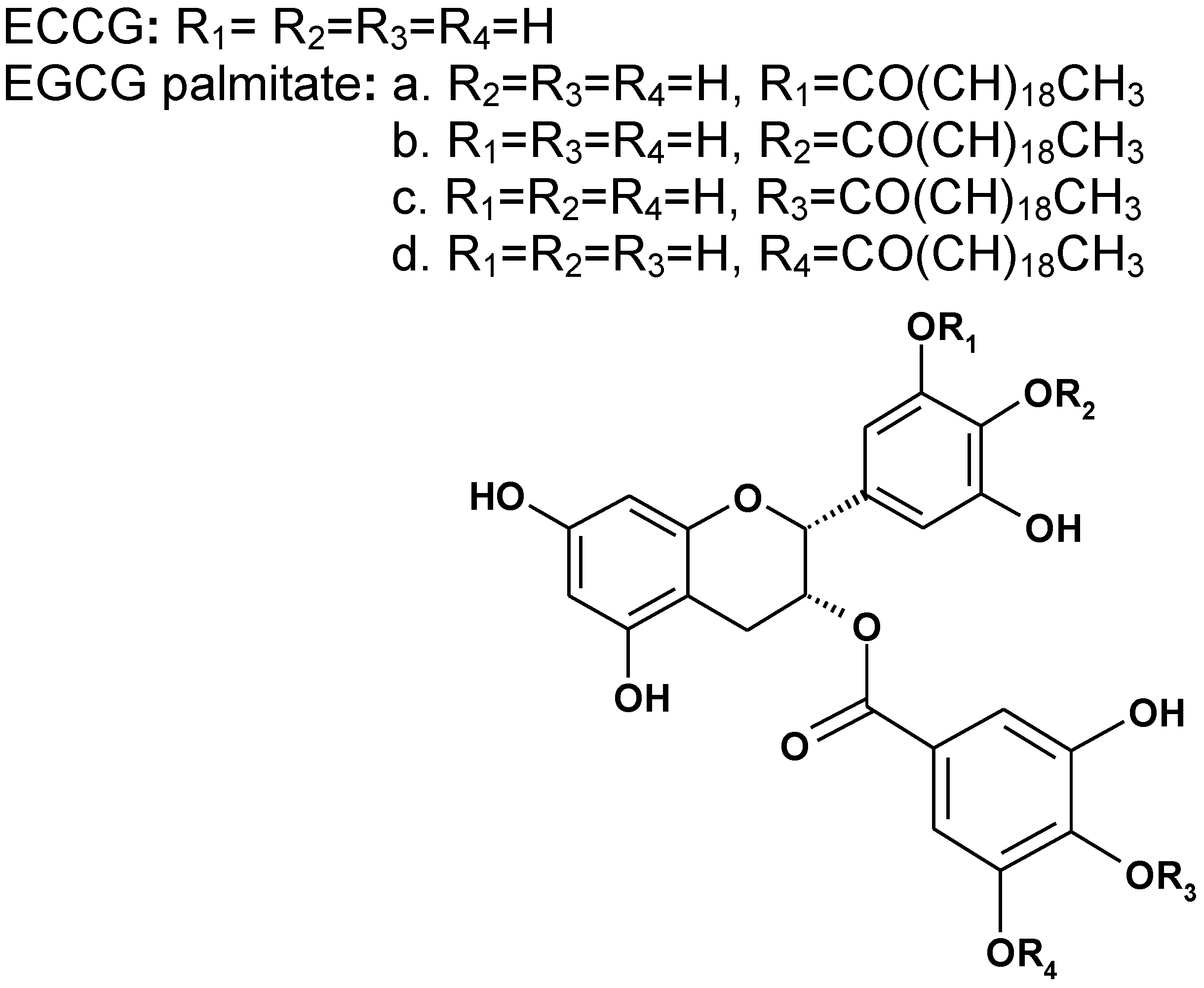
2.1. Chemical Structure of EGCG Palmitate

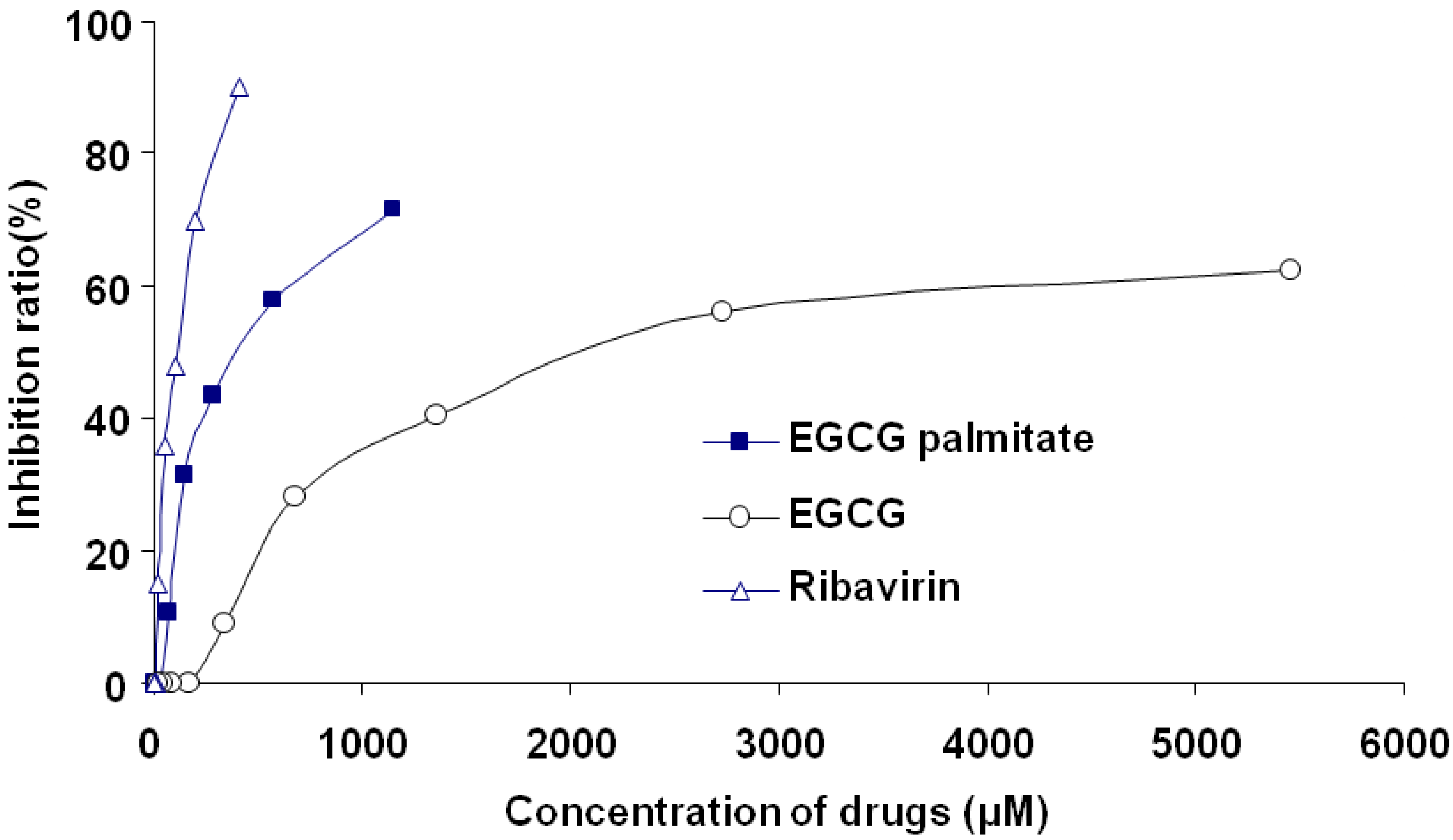
2.2. Cytotoxicity of EGCG, EGCG Palmitate, and Ribavirin


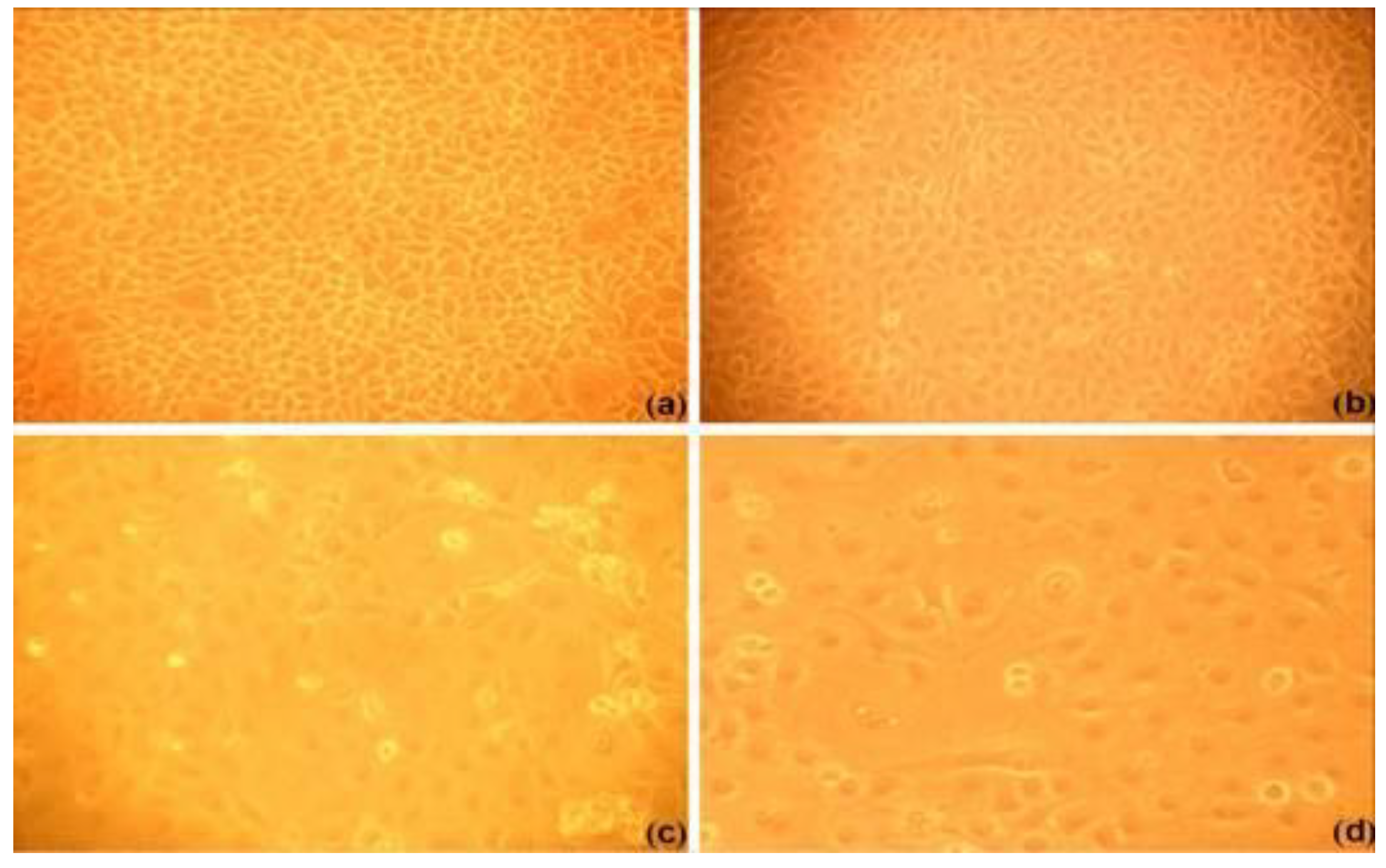
2.3. The TCID50 of PRRSV

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dilution | Infected well | Uninfected well | Cumulative infected well | Cumulative uninfected well | Infected (%) |
|---|---|---|---|---|---|
| 10−1 | 6 | 0 | 20 | 0 | 100 (20/20) |
| 10−2 | 6 | 0 | 14 | 0 | 100 (14/14) |
| 10−3 | 5 | 1 | 9 | 1 | 90 (9/10) |
| 10−4 | 3 | 3 | 3 | 4 | 42.8 (3/7) |
| 10−5 | 0 | 6 | 0 | 10 | 0 (0/10) |
| 10−6 | 0 | 6 | 0 | 16 | 0 (0/16) |
| 10−7 | 0 | 6 | 0 | 22 | 0 (0/22) |
| 10−8 | 0 | 6 | 0 | 28 | 0 (0/28) |
| 10−9 | 0 | 6 | 0 | 28 | 0 (0/28) |
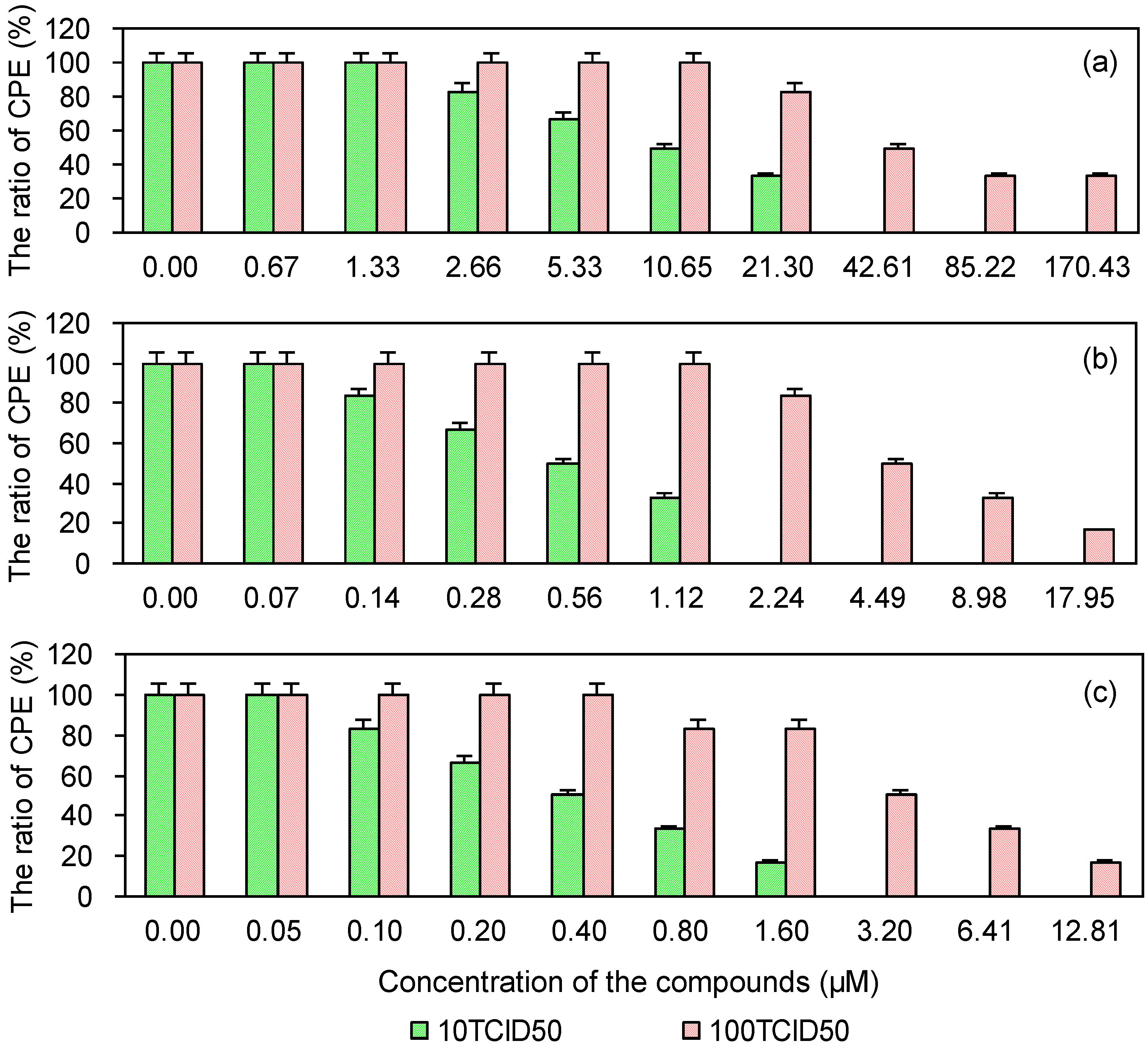
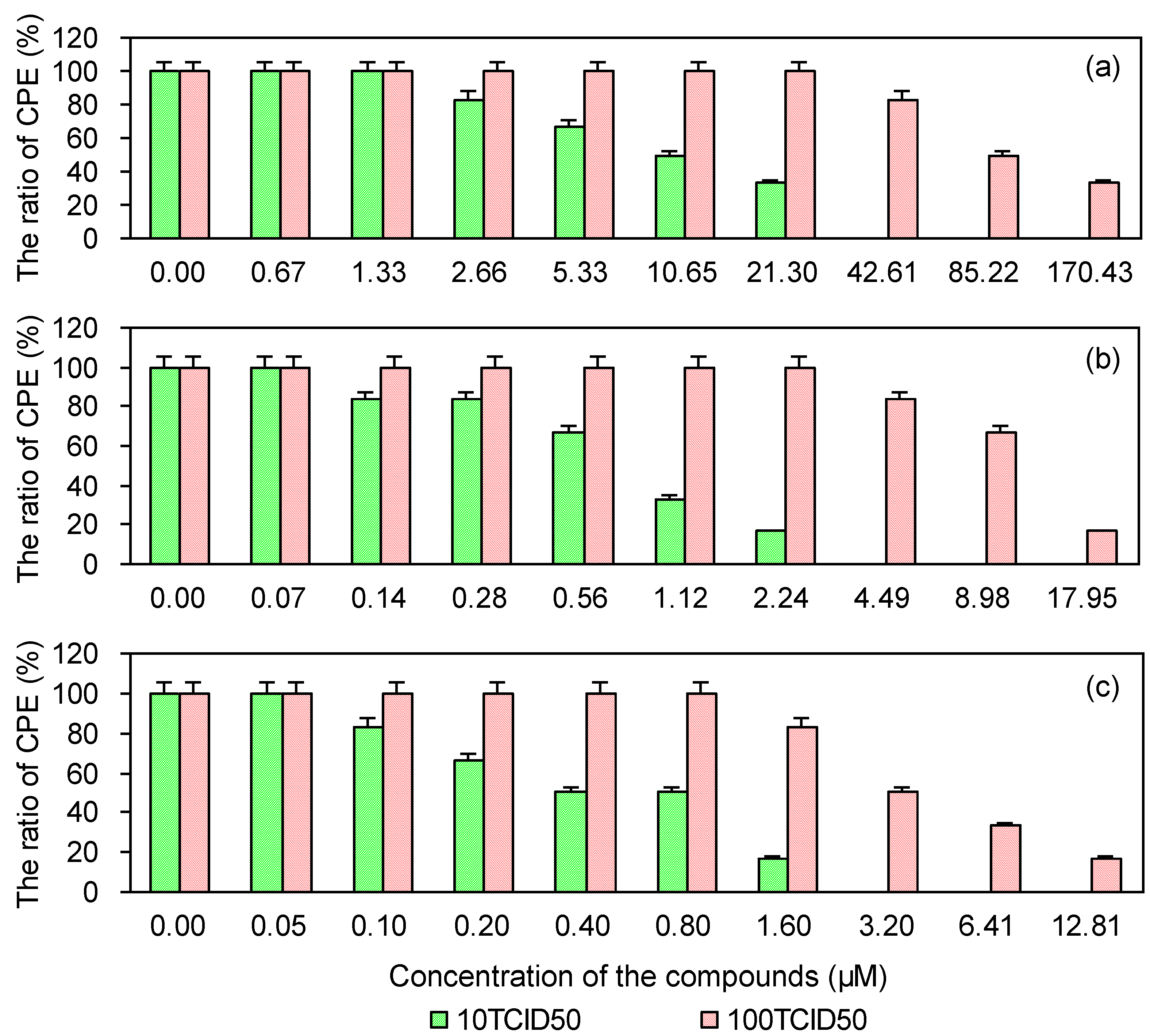
2.4. Cytopathic Effect of PRRSV on MARC-145 Cells Pre-Treated with the Test and Control Compounds

| Compounds | 10 TCID50 | 100 TCID50 | ||
|---|---|---|---|---|
| EC50 (μM) a | SI b | EC50 (μM) | SI | |
| EGCG | 8.53 | 276.62 | 60.25 | 39.16 |
| EGCG palmitate | 0.48 | 892.29 | 5.53 | 77.96 |
| Ribavirin | 0.40 | 234.98 | 3.48 | 27.02 |
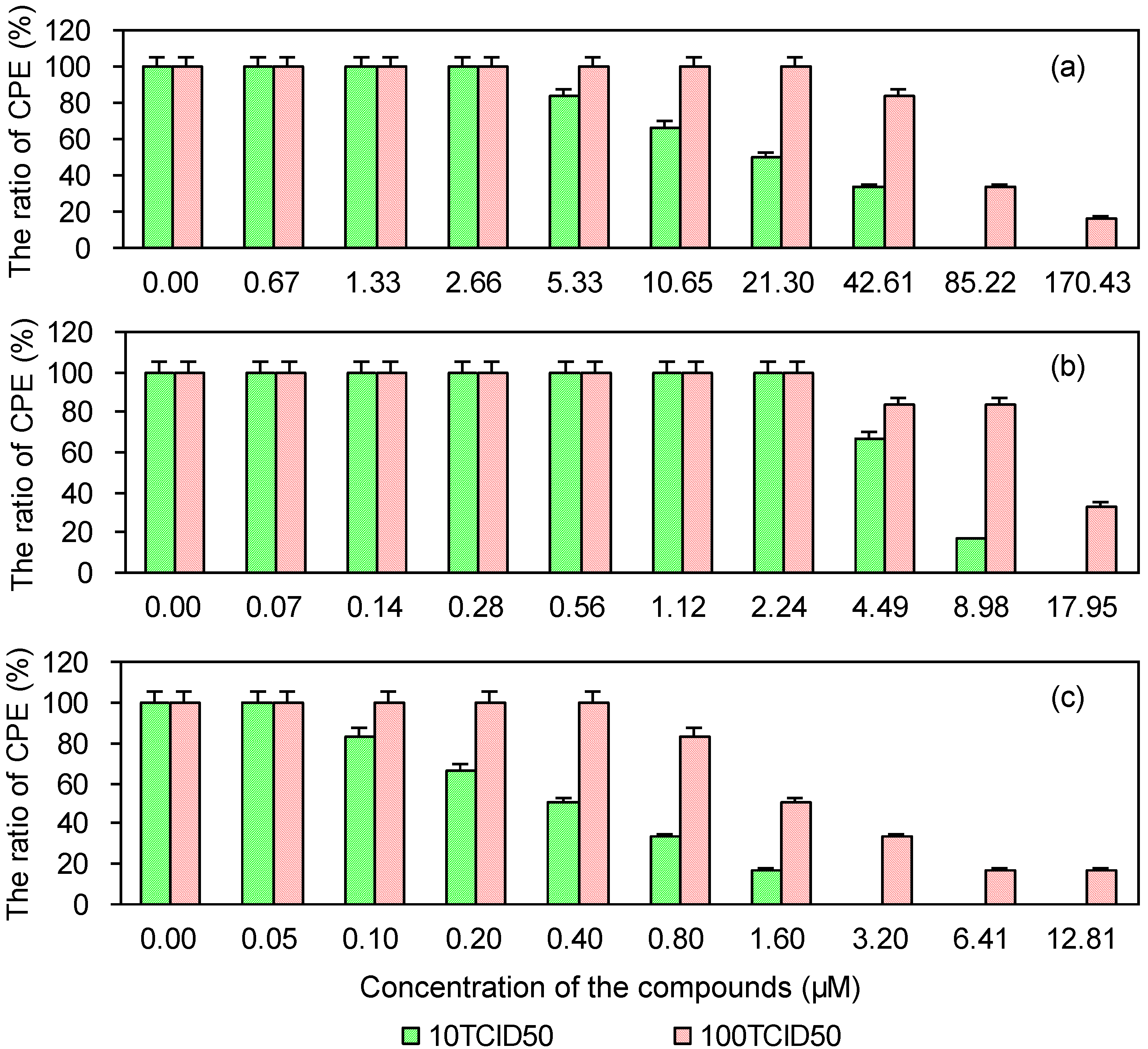
2.5. Cytopathic Effect of PRRSV in MARC-145 Cells Post-Treated with the Test and Control Compounds

| Compounds | 10 TCID50 | 100 TCID50 | ||
|---|---|---|---|---|
| EC50 (μM) a | SI b | EC50 (μM) | SI | |
| EGCG | 9.18 | 257.01 | 97.88 | 24.11 |
| EGCG palmitate | 0.68 | 635.93 | 9.43 | 45.75 |
| Ribavirin | 0.44 | 212.71 | 3.95 | 23.82 |
2.6. Cytopathic Effect of PRRSV on MARC-145 Cells Co-Treated with the Test and Control Compounds

| Compounds | 10TCID50 | 100TCID50 | ||
|---|---|---|---|---|
| EC50 (μM) a | SI b | EC50 (μM) | SI | |
| EGCG | 18.36 | 128.50 | 66.23 | 35.63 |
| EGCG palmitate | 5.86 | 73.64 | 12.69 | 33.99 |
| Ribavirin | 0.40 | 234.98 | 2.21 | 42.58 |
3. Discussion
4. Experimental
4.1. Materials and Chemicals
4.2. The Synthesis, Preparation, and Identification of EGCG Palmitate
4.3. Cell Culture
4.4. Cytotoxicity Testing of EGCG, EGCG Palmitate, and Ribavirin
4.5. PRRSV Titration
4.6. The Effect of EGCG Palmitate on PRRSV in Vitro
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Prieto, C.; Castro, J.M. Porcine reproductive and respiratory syndrome virus infection in the boar: A review. Theriogenology 2005, 63, 1–16. [Google Scholar] [CrossRef]
- Bao, Y.H.; Guo, Y.C.; Zhang, L.Y.; Zhao, Z.H.; Li, N. Inhibition of porcine reproductive and respiratory syndrome virus replication by RNA interference in MARC-145 cells. Mol. Biol. Rep. 2012, 39, 2515–2522. [Google Scholar] [CrossRef]
- Snijder, E.J.; Meulenberg, J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998, 79, 961–979. [Google Scholar]
- Xiao, S.A.; Wang, Q.W.; Gao, J.T.; Wang, L.L.; He, Z.Y.; Mo, D.L.; Liu, X.H.; Chen, Y.H. Inhibition of highly pathogenic PRRSV replication in MARC-145 cells by artificial microRNAs. Virol. J. 2011, 8, e491. [Google Scholar] [CrossRef]
- William, A.C.; Richard, G.D.; Grace, H.W.; Suleman, S.; Pam, W.D.; Raymond, R.R.; Eric, A.N. Porcine reproductive and respiratory syndrome virus (PRRSV) infection spreads by cell-to-cell transfer in cultured MARC-145 cells, is dependent on an intact cytoskeleton, and is suppressed by drug-targeting of cell permissiveness to virus infection. Virol. J. 2006, 3, e90. [Google Scholar] [CrossRef]
- Hu, J.; Ni, Y.; Dryman, B.A.; Meng, X.J.; Zhang, C. Immunogenicity study of plant-made oral subunit vaccine against porcine reproductive and respiratory syndrome virus (PRRSV). Vaccine 2012, 30, 2068–2074. [Google Scholar] [CrossRef]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; Green, A.L.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef]
- Song, J.M.; Lee, K.H.; Seong, B.L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Xu, J.; Wang, J.; Deng, F.; Hu, Z.; Wang, H. Green tea extract and its major component epigallocatechin gallate inhibits hepatitis B virus in vitro. Antivir. Res. 2008, 78, 242–249. [Google Scholar] [CrossRef]
- Kawai, K.; Tsuno, N.H.; Kitayama, J.; Okaji, Y.; Yazawa, K.; Asakage, M.; Hori, N.; Watanabe, T.; Takahashi, K.; Nagawa, H. Epigallocatechin gallate, the main component of tea polyphenol, binds to CD4 and interferes with gp120 binding. J. Allergy Clin. Immunol. 2003, 112, 951–957. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Calland, N.; Albecka, A.; Belouzard, S.; Wychowski, C.; Duverlie, G.; Descamps, V.; Hober, D.; Dubuisson, J.; Rouillé, Y.; Séron, K. (−)-Epigallocatechin-3-gallate is a new inhibitor of hepatitis C virus entry. Hepatology 2012, 55, 720–729. [Google Scholar] [CrossRef]
- Chen, C.; Qiu, H.; Gong, J.; Liu, Q.; Xiao, H.; Chen, X.W.; Su, B.L.; Yang, R.G. (−)-Epigallocatechin-3-gallate inhibits the replication cycle of hepatitis C virus. Arch. Virol. 2012, 157, 1301–1312. [Google Scholar] [CrossRef]
- Steinmann, J.; Buer, J.; Pietschmann, T.; Steinmann, E. Anti-infective properties of epigallocatechin-3-gallate (EGCG), a component of green tea. Br. J. Pharmacol. 2013, 168, 1059–1073. [Google Scholar] [CrossRef]
- Hong, J.; Lu, H.; Meng, X.; Ryu, J.H.; Hara, Y.; Yang, C.S. Stability, cellular uptake, biotransformation, and efflux of tea polyphenol (−)-epigallocatechin-3-gallate in HT-29 human colon adenocarcinoma cells. Cancer Res. 2002, 62, 7241–7246. [Google Scholar]
- Ying, Z.; Fereidoon, S. Lipophilised epigallocatechin gallate (EGCG) derivatives and their antioxidant potential in food and biological systems. Food Chem. 2012, 131, 22–30. [Google Scholar] [CrossRef]
- Ogbomo, H.; Michaelis, M.; Altenbrandt, B.; Doerr, H.W.; Cinatl, J. A novel immunomodulatory mechanismof ribavirin in suppressing natural killer cell function. Biochem. Pharmacol. 2010, 79, 188–197. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, X.; Zhang, H.; Wen, G.; Tong, C.; Li, X.; Fang, W. Molecular characterization of three porcine reproductive and respiratory syndrome virus isolates and their susceptibility to antiviral drugs. Pak. Vet. J. 2013, in press. [Google Scholar]
- Kim, Y.; Lee, C. Ribavirin efficiently suppresses porcine nidovirus replication. Virus Res. 2013, 171, 44–53. [Google Scholar] [CrossRef]
- Matsumura, K.; Kaihatsu, K.; Mori, S.; Cho, H.H.; Kato, N.; Hyon, S.H. Enhanced antitumor activities of (−)-epigallocatechin-3-O-gallate fatty acid monoester derivatives in vitro and in vivo. Biochem. Biophys. Res. Commun. 2008, 3, 1118–1122. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar]
- Nakayama, M.; Suzuki, K.; Toda, M.; Okubo, S.; Hara, Y.; Shimamura, T. Inhibition of the infectivity of influenza virus by tea polyphenols. Antivir. Res. 1993, 21, 289–299. [Google Scholar] [CrossRef]
- Nance, C.L.; Siwak, E.B.; Shearer, W.T. Preclinical development of the green tea catechin, epigallocatechin gallate, as an HIV-1 therapy. J. Allergy. Clin. Immunol. 2009, 123, 459–465. [Google Scholar] [CrossRef]
- Mori, S.; Miyake, S.; Kobe, T.; Nakaya, T.; Fuller, S.D.; Kato, N.; Kaihatsu, K. Enhanced anti-influenza A virus activity of (−)-epigallocatechin-3-O-gallate fatty acid monoester derivatives: Effect of alkyl chain length. Bioorg. Med. Chem. Lett. 2008, 18, 4249–4252. [Google Scholar] [CrossRef]
- Kaihatsu, K.; Mori, S.; Matsumura, H.; Daidoji, T.; Kawakami, C.; Kurata, H.; Nakaya, T.; Kato, N. Broad and potent anti-influenza virus spectrum of epigallocatechin-3-O-gallate-monopalmitate. J. Mol. Genet. Med. 2009, 3, 195–197. [Google Scholar]
- Zhou, H.R.; Tang, Q.W.; Yu, X.L.; Li, R.C.; Luo, W.; Jiang, D.L.; Ge, M. Antiviral effect of aqueous extracts of four kinds of tea on porcine reproductive and respiratory syndrome virus. J. Hunan Agric. Univ. 2012, 38, 305–309. [Google Scholar]
- Chen, P.; Sun, D.; Zheng, X.M. Preparation, structure and antioxidant activity of EGCG palmitate. J. Zhejiang Univ. (Sci. Edition) 2003, 30, 422–425. [Google Scholar]
- Ran, Z.G.; Chen, X.Y.; Guo, X.; Ge, X.N.; Yoon, K.J.; Yang, H.C. Recovery of viable porcine reproductive and respiratory syndrome virus from an infectious clone containing a partial deletion within the Nsp2-encoding region. Arch. Virol. 2008, 153, 899–907. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Mu, L.; Ding, Z. Immune responses in pigs induced by recombinant DNA vaccine co-expressing swine IL-18 and membrane protein of porcine reproductive and respiratory syndrome virus. Int. J. Mol. Sci. 2012, 13, 5715–5728. [Google Scholar] [CrossRef]
- Xu, J.M.; Song, S.T.; Tang, Z.M.; Jiang, Z.F.; Liu, X.Q.; Zhou, L.; Zhang, J.; Liu, X.W. Predictive chemotherapy of advanced breast cancer directed by MTT assay in vitro. Breast Canc. Res. Treat. 1999, 53, 77–85. [Google Scholar] [CrossRef]
- Erica, S.; Carol, B. Cytopathic effects of viruses protocols. Available online: http://www.microbelibrary.org/component/resource/laboratory-test/2875-cytopathic-effects-of-viruses-protocols/ (accessed on 22 July 2012).
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, C.; Liu, S.; Li, C.; Yang, L.; Zu, Y. In Vitro Evaluation of the Antiviral Activity of the Synthetic Epigallocatechin Gallate Analog-Epigallocatechin Gallate (EGCG) Palmitate against Porcine Reproductive and Respiratory Syndrome Virus. Viruses 2014, 6, 938-950. https://doi.org/10.3390/v6020938
Zhao C, Liu S, Li C, Yang L, Zu Y. In Vitro Evaluation of the Antiviral Activity of the Synthetic Epigallocatechin Gallate Analog-Epigallocatechin Gallate (EGCG) Palmitate against Porcine Reproductive and Respiratory Syndrome Virus. Viruses. 2014; 6(2):938-950. https://doi.org/10.3390/v6020938
Chicago/Turabian StyleZhao, Chunjian, Shuaihua Liu, Chunying Li, Lei Yang, and Yuangang Zu. 2014. "In Vitro Evaluation of the Antiviral Activity of the Synthetic Epigallocatechin Gallate Analog-Epigallocatechin Gallate (EGCG) Palmitate against Porcine Reproductive and Respiratory Syndrome Virus" Viruses 6, no. 2: 938-950. https://doi.org/10.3390/v6020938