Peste Des Petits Ruminants Virus Infection of Small Ruminants: A Comprehensive Review

 and
and {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Epidemiology
3. Economic Consequences
4. Virion Structure
4.1. Genome Organization

4.1.1. N Protein
4.1.2. P Protein
4.1.3. M Protein
4.1.4. F Protein
4.1.5. Hemagglutinin (Hemagglutinin-Neuraminidase) Protein
4.1.6. L Protein
4.1.7. Nonstructural Proteins
5. Resistance to Physical and Chemical Action
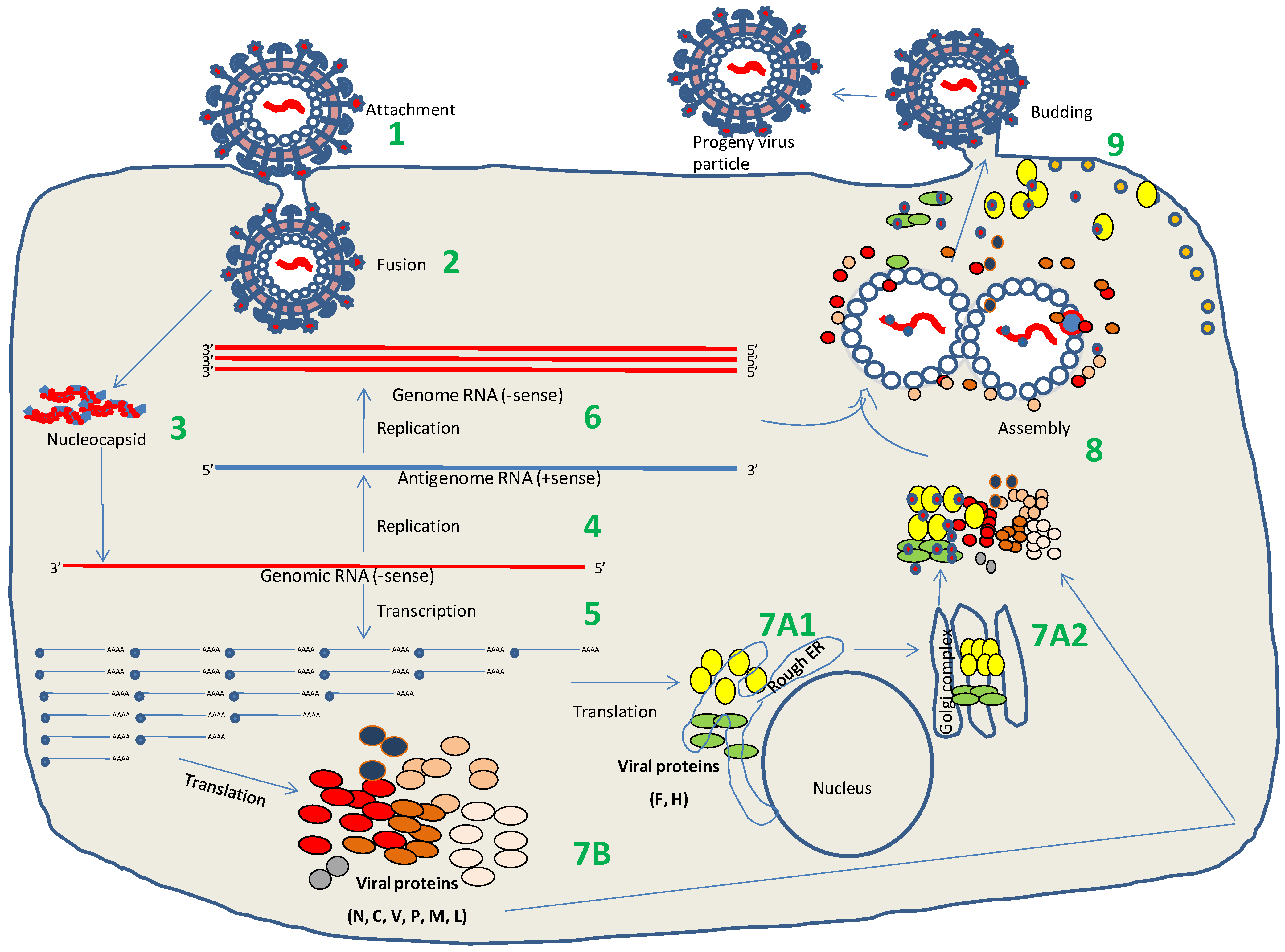
6. Virus Replication
6.1. Attachment and Entry
6.2. Transcription and Replication

6.3. Virus Assembly and Release
7. Immune Response
7.1. Innate Immune Response
7.2. Adaptive Immune Response
8. Pathogenesis
9. Immunosuppression
10. Host Susceptibility and Resistance to PPR
11. Disease Control
11.1. Vaccination
11.2. New Generation Vaccines
11.3. Progressive Control of PPR in SAARC Countries
11.4. Cross Protection within PPRV Strains
11.5. Should Cattle Be Included under the Mass Vaccination Campaign for Progressive Control of PPR?
11.6. Did Vaccination against Rinderpest Suppress the Upsurge of PPRV?
11.7. Should PPR Be Eradicated or a Live-With Option Be Adopted?
13. Conclusions
Acknowledgments
Author Contributions
Conflicts of interest
References and Notes
- Gargadennec, L.; Lalanne, A. La peste des petits ruminants. Bull. Serv. Zoot. Epiz. Afr. Occ. Fr. 1942, 5, 16–21. (in French). [Google Scholar]
- Pastoret, P. Rinderpest: A general introduction. In Rinderpest and Peste Des Petits Ruminants Virus: Plague of Large and Small Ruminants; Biology of Animal Infections; Academic Press: London, UK, 2006; pp. 1–12. [Google Scholar]
- Gibbs, E.P.; Taylor, W.P.; Lawman, M.J.; Bryant, J. Classification of peste des petits ruminants virus as the fourth member of the genus morbillivirus. Intervirology 1979, 11, 268–274. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Wong, B.H.; Fan, R.Y.; Wong, A.Y.; Zhang, A.J.; Wu, Y.; Choi, G.K.; Li, K.S.; Hui, J. Feline Morbillivirus, a previously undescribed Paramyxovirus associated with tubulointerstitial nephritis in domestic cats. Proc. Natl. Acad. Sci. USA 2012, 109, 5435–5440. [Google Scholar] [CrossRef]
- Gilbert, Y.; Monnier, J. Adaptation of the virus of peste des petits ruminants virus to tissue cultures. Revue Elev. Med. Vet. Pays Trop. 1962, 15, 321–335. [Google Scholar]
- Bourdin, P.; Laurent-Vautier, A. Note sur la structure du virus de la peste des petits ruminants. Rev. Elev. Med. Vet. Pays Trop. 1967, 20, 383–386. (in French). [Google Scholar]
- Wohlsein, P.; Saliki, J. Rinderpest and Peste Des Petits Ruminants Virus: Plague of Large and Small Ruminants; Biology of Animal Infections; Academic Press: London, UK, 2006; pp. 69–85. [Google Scholar]
- Singh, R.K.; Balamurugan, V.; Bhanuprakash, V.; Sen, A.; Saravanan, P.; Pal Yadav, M. Possible control and eradication of peste des petits ruminants from India: Technical aspects. Vet. Ital. 2009, 45, 449–462. [Google Scholar]
- Khan, H.A.; Siddique, M.; Sajjad ur, R.; Abubakar, M.; Ashraf, M. The detection of antibody against peste des petits ruminants virus in sheep, goats, cattle and buffaloes. Trop. Anim. Health Prod. 2008, 40, 521–527. [Google Scholar] [CrossRef]
- Taylor, W.P. Protection of goats against peste des Petits Ruminants with attenuated rinderpest virus. Res. Vet. Sci. 1979, 27, 321–324. [Google Scholar]
- Anderson, J.; McKay, J.A. The detection of antibodies against peste des petits ruminants virus in cattle, sheep and goats and the possible implications to rinderpest control programmes. Epidemiol. Infect. 1994, 112, 225–231. [Google Scholar] [CrossRef]
- Abubakar, M.; Jamal, S.M.; Arshed, M.J.; Hussain, M.; Ali, Q. Peste des petits ruminants virus (PPRV) infection; its association with species, seasonal variations and geography. Trop. Anim. Health Prod. 2009, 41, 1197–1202. [Google Scholar] [CrossRef]
- Felix, N. Current scenario and control initiatives for PPR at global, regional and country level according to the risk factors and socioeconomic impact. In Proceedings of the Second Regional Conference on Progressive Control of Peste Des Petits Ruminants in South Asia, Kathmandu, Nepal, 19–20 December 2013.
- Shaila, M.S.; Purushothaman, V.; Bhavasar, D.; Venugopal, K.; Venkatesan, R.A. Peste des petits ruminants of sheep in India. Vet. Rec. 1989, 125, 602. [Google Scholar]
- Taylor, W.P.; Diallo, A.; Gopalakrishna, S.; Sreeramalu, P.; Wilsmore, A.J.; Nanda, Y.P.; Libeau, G.; Rajasekhar, M.; Mukhopadhyay, A.K. Peste des petits ruminants has been widely present in southern India since, if not before, the late 1980s. Prev. Vet.Med. 2002, 52, 305–312. [Google Scholar] [CrossRef]
- Diallo, A. Control of peste des petits ruminants and poverty alleviation? J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 11–13. [Google Scholar] [CrossRef]
- Diallo, A.; Barrett, T.; Barbron, M.; Subbarao, S.M.; Taylor, W.P. Differentiation of rinderpest and peste des petits ruminants viruses using specific cDNA clones. J. Virol. Methods 1989, 23, 127–136. [Google Scholar] [CrossRef]
- Roger, F.; Diallo, A.; Yigezu, L.M.; Hurard, C.; Libeau, G.; Mebratu, G.Y.; Faye, B. Investigation of a new pathological condition of camels in Ethiopia. J. Camel Pract. Res. 2000, 2, 163–166. [Google Scholar]
- Govindarajan, R.; Koteeswaran, A.; Venugopalan, A.T.; Shyam, G.; Shaouna, S.; Shaila, M.S.; Ramachandran, S. Isolation of peste des petits ruminants virus from an outbreak in Indian buffalo (bubalus bubalis). Vet. Rec. 1997, 141, 573–574. [Google Scholar] [CrossRef]
- OIE. OIE-Listed diseases, infections and infestations in force in 2014. Available online: http://www.oie.int/animal-health-in-the-world/oie-listed-diseases-2014/ (accessed on 5 May 2014).
- Staal, S.; Poole, J.; Baltenweck, I.; Mwacharo, J.; Notenbaert, A.; Randolph, T.; Thorpe, W.; Nzuma, J.; Herrero, M. Strategic investment in livestock development as a vehicle for rural livelihoods; International Livestock Research Institute: Nairobi, Kenya, 15 October 2009; pp. 1–78. [Google Scholar]
- Perry, B.D.; Randolph, T.F.; McDermott, J.J.; Sones, K.R.; Thornton, P.K. Investing in animal health research to alleviate poverty; International Livestock Research Institute: Nairobi, Kenya, 2002; p. 148. [Google Scholar]
- Bailey, D.; Chard, L.S.; Dash, P.; Barrett, T.; Banyard, A.C. Reverse genetics for peste des petits ruminants virus (PPRV): Promoter and protein specificities. Virus Res. 2007, 126, 250–255. [Google Scholar] [CrossRef]
- Bailey, D.; Banyard, A.; Dash, P.; Ozkul, A.; Barrett, T. Full genome sequence of peste des petits ruminants virus, a member of the Morbillivirus genus. Virus Res. 2005, 110, 119–124. [Google Scholar] [CrossRef]
- Mahapatra, M.; Parida, S.; Baron, M.D.; Barrett, T. Matrix protein and glycoproteins F and H of peste des petits ruminants virus function better as a homologous complex. J. Gen. Virol. 2006, 87, 2021–2029. [Google Scholar] [CrossRef]
- Rahaman, A.; Srinivasan, N.; Shamala, N.; Shaila, M.S. The fusion core complex of the Peste des petits ruminants virus is a six-helix bundle assembly. Biochemistry 2003, 42, 922–931. [Google Scholar] [CrossRef]
- Ismail, T.M.; Yamanaka, M.K.; Saliki, J.T.; el-Kholy, A.; Mebus, C.; Yilma, T. Cloning and expression of the nucleoprotein of Peste des petits ruminants virus in baculovirus for use in serological diagnosis. Virology 1995, 208, 776–778. [Google Scholar] [CrossRef]
- Diallo, A.; Barrett, T.; Lefevre, P.C.; Taylor, W.P. Comparison of proteins induced in cells infected with rinderpest and peste des petits ruminants viruses. J. Gen. Virol. 1987, 68, 2033–2038. [Google Scholar] [CrossRef]
- Barrett, T.; Banyard, A.C.; Diallo, A. Molecular biology of the Morbilliviruses. In Rinderpest and Peste Des Petits Ruminants Virus: Plague of Large and Small Ruminants; Biology of Animal Infections; Academic Press: London, UK, 2006; pp. 31–67. [Google Scholar]
- Mitra-Kaushik, S.; Nayak, R.; Shaila, M.S. Identification of a cytotoxic T-cell epitope on the recombinant nucleocapsid proteins of rinderpest and peste des petits ruminants viruses presented as assembled nucleocapsids. Virology 2001, 279, 210–220. [Google Scholar] [CrossRef]
- Bankamp, B.; Horikami, S.M.; Thompson, P.D.; Huber, M.; Billeter, M.; Moyer, S.A. Domains of the Measles virus N protein required for binding to P protein and self-assembly. Virology 1996, 216, 272–277. [Google Scholar] [CrossRef]
- Rahaman, A.; Srinivasan, N.; Shamala, N.; Shaila, M.S. Phosphoprotein of the rinderpest virus forms a tetramer through a coiled coil region important for biological function. A structural insight. J. Biol. Chem. 2004, 279, 23606–23614. [Google Scholar] [CrossRef]
- Bao, J.Y.; Zhao, W.J.; Li, L.; Wang, Z.L.; Wu, G.Z.; Wu, X.D.; Liu, C.J.; Wang, Q.H.; Wang, J.W.; Liu, Y.T. Sequence analysis of the phosphoprotein gene of peste des petits ruminants virus of Chinese origin. Bing Du Xue Bao 2011, 27, 26–33. [Google Scholar]
- Mahapatra, M.; Parida, S.; Egziabher, B.G.; Diallo, A.; Barrett, T. Sequence analysis of the phosphoprotein gene of peste des petits ruminants (PPRV) virus: Editing of the gene transcript. Virus Res. 2003, 96, 85–98. [Google Scholar] [CrossRef]
- Muthuchelvan, D.; Sanyal, A.; Sarkar, J.; Sreenivasa, B.P.; Bandyopadhyay, S.K. Comparative nucleotide sequence analysis of the phosphoprotein gene of peste des petits ruminants vaccine virus of Indian origin. Res. Vet. Sci. 2006, 81, 158–164. [Google Scholar] [CrossRef]
- Kaushik, R.; Shaila, M.S. Cellular casein kinase II-mediated phosphorylation of rinderpest virus P protein is a prerequisite for its role in replication/transcription of the genome. J. Gen. Virol. 2004, 85, 687–691. [Google Scholar] [CrossRef]
- Johansson, K.; Bourhis, J.M.; Campanacci, V.; Cambillau, C.; Canard, B.; Longhi, S. Crystal structure of the Measles virus phosphoprotein domain responsible for the induced folding of the C-terminal domain of the nucleoprotein. J. Biol. Chem. 2003, 278, 44567–44573. [Google Scholar]
- Horikami, S.M.; Curran, J.; Kolakofsky, D.; Moyer, S.A. Complexes of sendai virus NP-P and P-L proteins are required for defective interfering particle genome replication in vitro. J. Virol. 1992, 66, 4901–4908. [Google Scholar]
- Liu, F.; Wu, X.; Li, L.; Liu, Z.; Wang, Z. Expression, purification and characterization of two truncated peste des petits ruminants virus matrix proteins in Escherichia coli, and production of polyclonal antibodies against this protein. Protein Expr. Purif. 2013, 91, 1–9. [Google Scholar] [CrossRef]
- Haffar, A.; Libeau, G.; Moussa, A.; Cecile, M.; Diallo, A. The matrix protein gene sequence analysis reveals close relationship between peste des petits ruminants virus (PPRV) and dolphin Morbillivirus. Virus Res. 1999, 64, 69–75. [Google Scholar] [CrossRef]
- Muthuchelvan, D.; Sanyal, A.; Sreenivasa, B.P.; Saravanan, P.; Dhar, P.; Singh, R.P.; Singh, R.K.; Bandyopadhyay, S.K. Analysis of the matrix protein gene sequence of the Asian lineage of peste des petits ruminants vaccine virus. Vet. Microbiol. 2006, 113, 83–87. [Google Scholar] [CrossRef]
- Coronel, E.C.; Takimoto, T.; Murti, K.G.; Varich, N.; Portner, A. Nucleocapsid incorporation into parainfluenza virus is regulated by specific interaction with matrix protein. J. Virol. 2001, 75, 1117–1123. [Google Scholar] [CrossRef]
- Bodjo, S.C.; Lelenta, M.; Couacy-Hymann, E.; Kwiatek, O.; Albina, E.; Gargani, D.; Libeau, G.; Diallo, A. Mapping the peste des petits ruminants virus nucleoprotein: Identification of two domains involved in protein self-association. Virus Res. 2008, 131, 23–32. [Google Scholar] [CrossRef]
- Moll, M.; Pfeuffer, J.; Klenk, H.D.; Niewiesk, S.; Maisner, A. Polarized glycoprotein targeting affects the spread of Measles virus in vitro and in vivo. J. Gen. Virol. 2004, 85, 1019–1027. [Google Scholar] [CrossRef]
- Vincent, S.; Spehner, D.; Manie, S.; Delorme, R.; Drillien, R.; Gerlier, D. Inefficient Measles virus budding in murine CD46 fibroblasts. Virology 1999, 265, 185–195. [Google Scholar] [CrossRef]
- Maisner, A.; Klenk, H.; Herrler, G. Polarized budding of Measles virus is not determined by viral surface glycoproteins. J. Virol. 1998, 72, 5276–5278. [Google Scholar]
- Moll, M.; Klenk, H.D.; Maisner, A. Importance of the cytoplasmic tails of the Measles virus glycoproteins for fusogenic activity and the generation of recombinant Measles viruses. J. Virol. 2002, 76, 7174–7186. [Google Scholar] [CrossRef]
- Seth, S.; Shaila, M.S. The fusion protein of Peste des petits ruminants virus mediates biological fusion in the absence of hemagglutinin-neuraminidase protein. Virology 2001, 289, 86–94. [Google Scholar] [CrossRef]
- Scheid, A.; Choppin, P.W. Identification of biological activities of paramyxovirus glycoproteins. Activation of cell fusion, hemolysis, and infectivity of proteolytic cleavage of an inactive precursor protein of sendai virus. Virology 1974, 57, 475–490. [Google Scholar] [CrossRef]
- Morrison, T.; Ward, L.J.; Semerjian, A. Intracellular processing of the Newcastle disease virus fusion glycoprotein. J. Virol. 1985, 53, 851–857. [Google Scholar]
- Paal, T.; Brindley, M.A.; St Clair, C.; Prussia, A.; Gaus, D.; Krumm, S.A.; Snyder, J.P.; Plemper, R.K. Probing the spatial organization of Measles virus fusion complexes. J. Virol. 2009, 83, 10480–10493. [Google Scholar] [CrossRef]
- Watanabe, M.; Hirano, A.; Stenglein, S.; Nelson, J.; Thomas, G.; Wong, T.C. Engineered serine protease inhibitor prevents furin-catalyzed activation of the fusion glycoprotein and production of infectious Measles virus. J. Virol. 1995, 69, 3206–3210. [Google Scholar]
- von Messling, V.; Milosevic, D.; Devaux, P.; Cattaneo, R. Canine distemper virus and Measles virus fusion glycoprotein trimers: Partial membrane-proximal ectodomain cleavage enhances function. J. Virol. 2004, 78, 7894–7903. [Google Scholar] [CrossRef]
- Novick, S.L.; Hoekstra, D. Membrane penetration of sendai virus glycoproteins during the early stages of fusion with liposomes as determined by hydrophobic photoaffinity labeling. Proc. Natl. Acad. Sci. U S A 1988, 85, 7433–7437. [Google Scholar] [CrossRef]
- Horvath, C.M.; Lamb, R.A. Studies on the fusion peptide of a Paramyxovirus fusion glycoprotein: Roles of conserved residues in cell fusion. J. Virol. 1992, 66, 2443–2455. [Google Scholar]
- Buckland, R.; Malvoisin, E.; Beauverger, P.; Wild, F. A leucine zipper structure present in the Measles virus fusion protein is not required for its tetramerization but is essential for fusion. J. Gen. Virol. 1992, 73, 1703–1707. [Google Scholar] [CrossRef]
- Sergel-Germano, T.; McQuain, C.; Morrison, T. Mutations in the fusion peptide and heptad repeat regions of the Newcastle disease virus fusion protein block fusion. J. Virol. 1994, 68, 7654–7658. [Google Scholar]
- Rapaport, D.; Shai, Y. Interaction of fluorescently labeled analogues of the amino-terminal fusion peptide of Sendai virus with phospholipid membranes. J. Biol. Chem. 1994, 269, 15124–15131. [Google Scholar]
- Reitter, J.N.; Sergel, T.; Morrison, T.G. Mutational analysis of the leucine zipper motif in the Newcastle disease virus fusion protein. J. Virol. 1995, 69, 5995–6004. [Google Scholar]
- Muhlebach, M.D.; Leonard, V.H.; Cattaneo, R. The Measles virus fusion protein transmembrane region modulates availability of an active glycoprotein complex and fusion efficiency. J. Virol. 2008, 82, 11437–11445. [Google Scholar] [CrossRef]
- Lee, J.K.; Prussia, A.; Snyder, J.P.; Plemper, R.K. Reversible inhibition of the fusion activity of Measles virus F protein by an engineered intersubunit disulfide bridge. J. Virol. 2007, 81, 8821–8826. [Google Scholar] [CrossRef]
- Dutch, R.E.; Jardetzky, T.S.; Lamb, R.A. Virus membrane fusion proteins: Biological machines that undergo a metamorphosis. Biosci. Rep. 2000, 20, 597–612. [Google Scholar] [CrossRef]
- Hernandez, L.D.; Hoffman, L.R.; Wolfsberg, T.G.; White, J.M. Virus-cell and cell-cell fusion. Annu. Rev. Cell Dev. Biol. 1996, 12, 627–661. [Google Scholar] [CrossRef]
- Lawless-Delmedico, M.K.; Sista, P.; Sen, R.; Moore, N.C.; Antczak, J.B.; White, J.M.; Greene, R.J.; Leanza, K.C.; Matthews, T.J.; Lambert, D.M. Heptad-repeat regions of respiratory syncytial virus F1 protein form a six-membered coiled-coil complex. Biochemistry 2000, 39, 11684–11695. [Google Scholar] [CrossRef]
- Singethan, K.; Hiltensperger, G.; Kendl, S.; Wohlfahrt, J.; Plattet, P.; Holzgrabe, U.; Schneider-Schaulies, J. N-(3-cyanophenyl)-2-phenylacetamide, an effective inhibitor of Morbillivirus-induced membrane fusion with low cytotoxicity. J. Gen. Virol. 2010, 91, 2762–2772. [Google Scholar] [CrossRef]
- Meyer, G.; Diallo, A. The nucleotide sequence of the fusion protein gene of the peste des petits ruminants virus: The long untranslated region in the 5'-end of the F-protein gene of Morbilliviruses seems to be specific to each virus. Virus Res. 1995, 37, 23–35. [Google Scholar] [CrossRef]
- Alkhatib, G.; Roder, J.; Richardson, C.; Briedis, D.; Weinberg, R.; Smith, D.; Taylor, J.; Paoletti, E.; Shen, S.H. Characterization of a cleavage mutant of the Measles virus fusion protein defective in syncytium formation. J. Virol. 1994, 68, 6770–6774. [Google Scholar]
- Heidmeier, S.; Hanauer, J.R.; Friedrich, K.; Prufer, S.; Schneider, I.C.; Buchholz, C.J.; Cichutek, K.; Muhlebach, M.D. A single amino acid substitution in the Measles virus F protein reciprocally modulates membrane fusion activity in pathogenic and oncolytic strains. Virus Res. 2013. [Google Scholar]
- Apte-Sengupta, S.; Negi, S.; Leonard, V.H.; Oezguen, N.; Navaratnarajah, C.K.; Braun, W.; Cattaneo, R. Base of the Measles virus fusion trimer head receives the signal that triggers membrane fusion. J. Biol. Chem. 2012, 287, 33026–33035. [Google Scholar] [CrossRef]
- Plattet, P.; Langedijk, J.P.; Zipperle, L.; Vandevelde, M.; Orvell, C.; Zurbriggen, A. Conserved leucine residue in the head region of Morbillivirus fusion protein regulates the large conformational change during fusion activity. Biochemistry 2009, 48, 9112–9121. [Google Scholar] [CrossRef]
- Devireddy, L.R.; Raghavan, R.; Ramachandran, S.; Shaila, M.S. The fusion protein of Peste des petits ruminants virus is a hemolysin. Arch. Virol. 1999, 144, 1241–1247. [Google Scholar] [CrossRef]
- Delpeut, S.; Rudd, P.A.; Labonte, P.; von Messling, V. Membrane fusion-mediated autophagy induction enhances morbillivirus cell-to-cell spread. J. Virol. 2012, 86, 8527–8535. [Google Scholar]
- Langedijk, J.P.; Daus, F.J.; van Oirschot, J.T. Sequence and structure alignment of Paramyxoviridae attachment proteins and discovery of enzymatic activity for a Morbillivirus hemagglutinin. J. Virol. 1997, 71, 6155–6167. [Google Scholar]
- Balamurugan, V.; Sen, A.; Saravanan, P.; Rasool, T.J.; Yadav, M.P.; Bandyopadhyay, S.K.; Singh, R.K. Development and characterization of a stable vero cell line constitutively expressing peste des petits ruminants virus (PPRV) hemagglutinin protein and its potential use as antigen in enzyme-linked immunosorbent assay for serosurveillance of PPRV. Clin. Vaccine Immunol. 2006, 13, 1367–1372. [Google Scholar]
- Galbraith, S.E.; Tiwari, A.; Baron, M.D.; Lund, B.T.; Barrett, T.; Cosby, S.L. Morbillivirus downregulation of CD46. J. Virol. 1998, 72, 10292–10297. [Google Scholar]
- Lecouturier, V.; Fayolle, J.; Caballero, M.; Carabana, J.; Celma, M.L.; Fernandez-Munoz, R.; Wild, T.F.; Buckland, R. Identification of two amino acids in the hemagglutinin glycoprotein of Measles virus (MV) that govern hemadsorption, Hela cell fusion, and CD46 downregulation: Phenotypic markers that differentiate vaccine and wild-type MV strains. J. Virol. 1996, 70, 4200–4204. [Google Scholar]
- Kumar, N.; Chaubey, K.K.; Chaudhary, K.; Singh, S.V.; Sharma, D.K.; Gupta, V.K.; Mishra, A.K.; Sharma, S. Isolation, identification and characterization of a peste des petits ruminants virus from an outbreak in Nanakpur, India. J. Virol. Methods 2013, 189, 388–392. [Google Scholar] [CrossRef]
- Malur, A.G.; Choudhary, S.K.; De, B.P.; Banerjee, A.K. Role of a highly conserved N-terminal domain of the human parainfluenza virus type 3 RNA polymerase. J. Virol. 2002, 76, 8101–8109. [Google Scholar] [CrossRef]
- Cartee, T.L.; Megaw, A.G.; Oomens, A.G.; Wertz, G.W. Identification of a single amino acid change in the human respiratory syncytial virus L protein that affects transcriptional termination. J. Virol. 2003, 77, 7352–7360. [Google Scholar] [CrossRef]
- Blumberg, B.M.; Crowley, J.C.; Silverman, J.I.; Menonna, J.; Cook, S.D.; Dowling, P.C. Measles virus L protein evidences elements of ancestral RNA polymerase. Virology 1988, 164, 487–497. [Google Scholar] [CrossRef]
- Bellini, W.J.; Englund, G.; Rozenblatt, S.; Arnheiter, H.; Richardson, C.D. Measles virus P gene codes for two proteins. J. Virol. 1985, 53, 908–919. [Google Scholar]
- Sweetman, D.A.; Miskin, J.; Baron, M.D. Rinderpest virus C and V proteins interact with the major (L) component of the viral polymerase. Virology 2001, 281, 193–204. [Google Scholar] [CrossRef]
- Escoffier, C.; Manie, S.; Vincent, S.; Muller, C.P.; Billeter, M.; Gerlier, D. Nonstructural C protein is required for efficient Measles virus replication in human peripheral blood cells. J. Virol. 1999, 73, 1695–1698. [Google Scholar]
- Baron, M.D.; Barrett, T. Rinderpest viruses lacking the C and V proteins show specific defects in growth and transcription of viral RNAs. J. Virol. 2000, 74, 2603–2611. [Google Scholar] [CrossRef]
- Patterson, J.B.; Thomas, D.; Lewicki, H.; Billeter, M.A.; Oldstone, M.B. V and C proteins of Measles virus function as virulence factors in vivo. Virology 2000, 267, 80–89. [Google Scholar] [CrossRef]
- Ito, M.; Iwasaki, M.; Takeda, M.; Nakamura, T.; Yanagi, Y.; Ohno, S. Measles virus non-structural C protein modulates viral RNA polymerase activity by interacting with host protein SHCBP1. J. Virol. 2013, 87, 9633–9642. [Google Scholar] [CrossRef]
- Boxer, E.L.; Nanda, S.K.; Baron, M.D. The rinderpest virus non-structural C protein blocks the induction of type 1 interferon. Virology 2009, 385, 134–142. [Google Scholar] [CrossRef]
- Chinnakannan, S.K.; Holzer, B.; Bernardo, B.S.; Nanda, S.K.; Baron, M.D. Different functions of the common P/V/W and V-specific domains of rinderpest virus V protein in blocking IFN signalling. J. Gen. Virol. 2014, 95, 44–51. [Google Scholar] [CrossRef]
- Tober, C.; Seufert, M.; Schneider, H.; Billeter, M.A.; Johnston, I.C.; Niewiesk, S.; ter Meulen, V.; Schneider-Schaulies, S. Expression of Measles virus V protein is associated with pathogenicity and control of viral RNA synthesis. J. Virol. 1998, 72, 8124–8132. [Google Scholar]
- Coetzer, J.A.W.; Tustin, R.C. Infectious Diseases of Livestock; Oxford University Press: New York, NY, USA, 2004; pp. 1–2352. [Google Scholar]
- Pawar, R.M.; Dhinakar Raj, G.; Balachandran, C. Relationship between the level of signaling lymphocyte activation molecule and replication of peste-des-petits-ruminants virus in peripheral blood mononuclear cells of host animals. Acta Virol. 2008, 52, 231–236. [Google Scholar]
- Adombi, C.M.; Lelenta, M.; Lamien, C.E.; Shamaki, D.; Koffi, Y.M.; Traore, A.; Silber, R.; Couacy-Hymann, E.; Bodjo, S.C.; Djaman, J.A. Monkey CV1 cell line expressing the sheep-goat SLAM protein: A highly sensitive cell line for the isolation of peste des petits ruminants virus from pathological specimens. J. Virol. Methods 2011, 173, 306–313. [Google Scholar] [CrossRef]
- Munir, M.; Zohari, S.; Berg, M. Genome organization. In Molecular Biology and Pathogenesis of Peste Des Petits Ruminants Virus; Springer: Heidelberg, Germany, 2013; pp. 1–18. [Google Scholar]
- Noyce, R.S.; Bondre, D.G.; Ha, M.N.; Lin, L.T.; Sisson, G.; Tsao, M.S.; Richardson, C.D. Tumor cell marker PVRl4 (Nectin-4) is an epithelial cell receptor for Measles virus. PLoS Pathog. 2011, 7, e1002240. [Google Scholar] [CrossRef]
- Tahara, M.; Takeda, M.; Shirogane, Y.; Hashiguchi, T.; Ohno, S.; Yanagi, Y. Measles virus infects both polarized epithelial and immune cells by using distinctive receptor-binding sites on its hemagglutinin. J. Virol. 2008, 82, 4630–4637. [Google Scholar] [CrossRef]
- Leonard, V.H.; Sinn, P.L.; Hodge, G.; Miest, T.; Devaux, P.; Oezguen, N.; Braun, W.; McCray, P.B., Jr.; McChesney, M.B.; Cattaneo, R. Measles virus blind to its epithelial cell receptor remains virulent in rhesus monkeys but cannot cross the airway epithelium and is not shed. J. Clin. Invest. 2008, 118, 2448–2458. [Google Scholar]
- Sato, H.; Yoneda, M.; Honda, T.; Kai, C. Morbillivirus receptors and tropism: Multiple pathways for infection. Front. Microbiol. 2012, 3, 75. [Google Scholar]
- Hashimoto, K.; Ono, N.; Tatsuo, H.; Minagawa, H.; Takeda, M.; Takeuchi, K.; Yanagi, Y. SLAM (CD150)-independent Measles virus entry as revealed by recombinant virus expressing green fluorescent protein. J. Virol. 2002, 76, 6743–6749. [Google Scholar] [CrossRef]
- Fujita, K.; Miura, R.; Yoneda, M.; Shimizu, F.; Sato, H.; Muto, Y.; Endo, Y.; Tsukiyama-Kohara, K.; Kai, C. Host range and receptor utilization of canine distemper virus analyzed by recombinant viruses: Involvement of heparin-like molecule in CDV infection. Virology 2007, 359, 324–335. [Google Scholar] [CrossRef]
- Kozak, M. Selection of initiation sites by eucaryotic ribosomes: Effect of inserting AUG triplets upstream from the coding sequence for preproinsulin. Nucleic Acids Res. 1984, 12, 3873–3893. [Google Scholar] [CrossRef]
- Schneider, H.; Kaelin, K.; Billeter, M.A. Recombinant Measles viruses defective for RNA editing and V protein synthesis are viable in cultured cells. Virology 1997, 227, 314–322. [Google Scholar] [CrossRef]
- Ward, S.V.; George, C.X.; Welch, M.J.; Liou, L.Y.; Hahm, B.; Lewicki, H.; de la Torre, J.C.; Samuel, C.E.; Oldstone, M.B. RNA editing enzyme adenosine deaminase is a restriction factor for controlling Measles virus replication that also is required for embryogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 331–336. [Google Scholar] [CrossRef]
- Otsuki, N.; Nakatsu, Y.; Kubota, T.; Sekizuka, T.; Seki, F.; Sakai, K.; Kuroda, M.; Yamaguchi, R.; Takeda, M. The V protein of canine distemper virus is required for virus replication in human epithelial cells. PLoS One 2013, 8, e82343. [Google Scholar] [CrossRef]
- Liermann, H.; Harder, T.C.; Lochelt, M.; von Messling, V.; Baumgartner, W.; Moennig, V.; Haas, L. Genetic analysis of the central untranslated genome region and the proximal coding part of the F gene of wild-type and vaccine canine distemper morbilliviruses. Virus Genes 1998, 17, 259–270. [Google Scholar] [CrossRef]
- Evans, S.A.; Belsham, G.J.; Barrett, T. The role of the 5' nontranslated regions of the fusion protein mRNAs of canine distemper virus and rinderpest virus. Virology 1990, 177, 317–323. [Google Scholar] [CrossRef]
- Anderson, D.E.; Castan, A.; Bisaillon, M.; von Messling, V. Elements in the canine distemper virus m 3' UTR contribute to control of replication efficiency and virulence. PLoS One 2012, 7, e31561. [Google Scholar]
- Esteban, D.J.; Upton, C.; Bartow-McKenney, C.; Buller, R.M.; Chen, N.G.; Schriewer, J.; Lefkowitz, E.J.; Wang, C. Expression of a non-coding RNA in Ectromelia virus is required for normal plaque formation. Virus Genes 2014, 48, 38–47. [Google Scholar] [CrossRef]
- Chang, R.Y.; Hsu, T.W.; Chen, Y.L.; Liu, S.F.; Tsai, Y.J.; Lin, Y.T.; Chen, Y.S.; Fan, Y.H. Japanese encephalitis virus non-coding RNA inhibits activation of interferon by blocking nuclear translocation of interferon regulatory factor 3. Vet. Microbiol. 2013, 166, 11–21. [Google Scholar] [CrossRef]
- Samanta, M.; Takada, K. Modulation of innate immunity system by Epstein-Barr virus-encoded non-coding RNA and oncogenesis. Canc. Sci. 2010, 101, 29–35. [Google Scholar] [CrossRef]
- Wu, Y.L.; Wu, C.P.; Liu, C.Y.; Hsu, P.W.; Wu, E.C.; Chao, Y.C. A non-coding RNA of insect HzNV-1 virus establishes latent viral infection through microRNA. Sci. Rep. 2011, 1, 60. [Google Scholar]
- Wertz, G.W.; Perepelitsa, V.P.; Ball, L.A. Gene rearrangement attenuates expression and lethality of a nonsegmented negative strand RNA virus. Proc. Natl. Acad. Sci. USA 1998, 95, 3501–3506. [Google Scholar] [CrossRef]
- Gupta, A.K.; Shaji, D.; Banerjee, A.K. Identification of a novel tripartite complex involved in replication of vesicular stomatitis virus genome RNA. J. Virol. 2003, 77, 732–738. [Google Scholar] [CrossRef]
- Kolakofsky, D.; Le Mercier, P.; Iseni, F.; Garcin, D. Viral DNA polymerase scanning and the gymnastics of sendai virus RNA synthesis. Virology 2004, 318, 463–473. [Google Scholar] [CrossRef]
- Harrison, M.S.; Sakaguchi, T.; Schmitt, A.P. Paramyxovirus assembly and budding: Building particles that transmit infections. Int. J. Biochem. Cell. Biol. 2010, 42, 1416–1429. [Google Scholar] [CrossRef]
- Kondo, T.; Yoshida, T.; Miura, N.; Nakanishi, M. Temperature-sensitive phenotype of a mutant Sendai virus strain is caused by its insufficient accumulation of the M protein. J. Biol. Chem. 1993, 268, 21924–21930. [Google Scholar]
- Yoshida, T.; Nagai, Y.; Maeno, K.; Iinuma, M.; Hamaguchi, M.; Matsumoto, T.; Nagayoshi, S.; Hoshino, M. Studies on the role of M protein in virus assembly using a ts mutant of HVJ (sendai virus). Virology 1979, 92, 139–154. [Google Scholar] [CrossRef]
- Iwasaki, M.; Takeda, M.; Shirogane, Y.; Nakatsu, Y.; Nakamura, T.; Yanagi, Y. The matrix protein of Measles virus regulates viral RNA synthesis and assembly by interacting with the nucleocapsid protein. J. Virol. 2009, 83, 10374–10383. [Google Scholar] [CrossRef]
- Vincent, S.; Gerlier, D.; Manie, S.N. Measles virus assembly within membrane rafts. J. Virol. 2000, 74, 9911–9915. [Google Scholar] [CrossRef]
- Shaikh, F.Y.; Crowe, J.E., Jr. Molecular mechanisms driving respiratory syncytial virus assembly. Future Microbiol. 2013, 8, 123–131. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Finberg, R.W.; Kurt-Jones, E.A. Viruses and Toll-like receptors. Microbes Infect. 2004, 6, 1356–1360. [Google Scholar] [CrossRef]
- Finberg, R.W.; Wang, J.P.; Kurt-Jones, E.A. Toll like receptors and viruses. Rev. Med. Virol. 2007, 17, 35–43. [Google Scholar] [CrossRef]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef]
- Ichinohe, T.; Lee, H.K.; Ogura, Y.; Flavell, R.; Iwasaki, A. Inflammasome recognition of influenza virus is essential for adaptive immune responses. J. Exp. Med. 2009, 206, 79–87. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Platanias, L.C. Mechanisms of Type-I- and Type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef]
- Baron, J.; Bin-Tarif, A.; Herbert, R.; Frost, L.; Taylor, G.; Baron, M.D. Early changes in cytokine expression in peste des petits ruminants disease. Vet. Res. 2014, 45, 22. [Google Scholar] [CrossRef]
- Atmaca, H.T.; Kul, O. Examination of epithelial tissue cytokine response to natural peste des petits ruminants virus (PPRV) infection in sheep and goats by immunohistochemistry. Histol. Histopathol. 2012, 27, 69–78. [Google Scholar]
- Caignard, G.; Bourai, M.; Jacob, Y.; Tangy, F.; Vidalain, P.O. Inhibition of IFN-alpha/beta signaling by two discrete peptides within measles virus V protein that specifically bind STAT1 and STAT2. Virology 2009, 383, 112–120. [Google Scholar]
- Fontana, J.M.; Bankamp, B.; Bellini, W.J.; Rota, P.A. Regulation of interferon signaling by the C and V proteins from attenuated and wild-type strains of measles virus. Virology 2008, 374, 71–81. [Google Scholar] [CrossRef]
- Ramachandran, A.; Parisien, J.P.; Horvath, C.M. STAT2 is a primary target for measles virus V protein-mediated alpha/beta interferon signaling inhibition. J. Virol. 2008, 82, 8330–8338. [Google Scholar]
- Caignard, G.; Guerbois, M.; Labernardiere, J.L.; Jacob, Y.; Jones, L.M.; Infectious Mapping Project I-MAP; Wild, F.; Tangy, F.; Vidalain, P.O. Measles virus v protein blocks JAK1-mediated phosphorylation of STAT1 to escape IFN-alpha/beta signaling. Virology 2007, 368, 351–362. [Google Scholar] [CrossRef]
- Nanda, S.K.; Baron, M.D. Rinderpest virus blocks Type I and Type II interferon action: Role of structural and nonstructural proteins. J. Virol. 2006, 80, 7555–7568. [Google Scholar] [CrossRef]
- Ohno, S.; Ono, N.; Takeda, M.; Takeuchi, K.; Yanagi, Y. Dissection of measles virus V protein in relation to its ability to block alpha/beta interferon signal transduction. J. Gen. Virol. 2004, 85, 2991–2999. [Google Scholar] [CrossRef]
- Palosaari, H.; Parisien, J.P.; Rodriguez, J.J.; Ulane, C.M.; Horvath, C.M. STAT protein interference and suppression of cytokine signal transduction by measles virus V protein. J. Virol. 2003, 77, 7635–7644. [Google Scholar] [CrossRef]
- Takeuchi, K.; Kadota, S.I.; Takeda, M.; Miyajima, N.; Nagata, K. Measles virus V protein blocks interferon (IFN)-alpha/beta but not IFN-gamma signaling by inhibiting STAT1 and STAT2 phosphorylation. FEBS Lett. 2003, 545, 177–182. [Google Scholar] [CrossRef]
- Chinnakannan, S.K.; Nanda, S.K.; Baron, M.D. Morbillivirus v proteins exhibit multiple mechanisms to block Type 1 and Type 2 interferon signalling pathways. PLoS One 2013, 8, e57063. [Google Scholar]
- Naik, S.; Renukaradhya, G.J.; Rajasekhar, M.; Shaila, M.S. Immunogenic and protective properties of haemagglutinin protein (H) of rinderpest virus expressed by a recombinant baculovirus. Vaccine 1997, 15, 603–607. [Google Scholar] [CrossRef]
- Naik, S.; Shaila, M.S. Characterization of membrane-bound and membrane anchor-less forms of hemagglutinin glycoprotein of rinderpest virus expressed by baculovirus recombinants. Virus Genes 1997, 14, 95–104. [Google Scholar] [CrossRef]
- Sinnathamby, G.; Nayak, R.; Shaila, M.S. Mapping of T-helper epitopes of rinderpest virus hemagglutinin protein. Viral Immunol. 2001, 14, 83–92. [Google Scholar] [CrossRef]
- Griffin, D.E. Immune responses during Measles virus infection. Curr. Top. Microbiol. Immunol. 1995, 191, 117–134. [Google Scholar]
- Munir, M. Mononegaviruses of Veterinary Importance, Pathobiology and Molecular Diagnosis; CABI International: Oxfordshire, UK, 2013; Volume 1, pp. 65–98. [Google Scholar]
- Romero, C.H.; Barrett, T.; Kitching, R.P.; Carn, V.M.; Black, D.N. Protection of cattle against rinderpest and lumpy skin disease with a recombinant capripoxvirus expressing the fusion protein gene of rinderpest virus. Vet. Rec. 1994, 135, 152–154. [Google Scholar] [CrossRef]
- Sinnathamby, G.; Renukaradhya, G.J.; Rajasekhar, M.; Nayak, R.; Shaila, M.S. Immune responses in goats to recombinant hemagglutinin-neuraminidase glycoprotein of peste des petits ruminants virus: Identification of a T-cell determinant. Vaccine 2001, 19, 4816–4823. [Google Scholar] [CrossRef]
- Diallo, A.; Minet, C.; Le Goff, C.; Berhe, G.; Albina, E.; Libeau, G.; Barrett, T. The threat of peste des petits ruminants: Progress in vaccine development for disease control. Vaccine 2007, 25, 5591–5597. [Google Scholar] [CrossRef]
- Hickman, C.J.; Khan, A.S.; Rota, P.A.; Bellini, W.J. Use of synthetic peptides to identify Measles nucleoprotein T-cell epitopes in vaccinated and naturally infected humans. Virology 1997, 235, 386–397. [Google Scholar] [CrossRef]
- Ohishi, K.; Inui, K.; Yamanouchi, K.; Barrett, T. Cell-mediated immune responses in cattle vaccinated with a vaccinia virus recombinant expressing the nucleocapsid protein of rinderpest virus. J. Gen. Virol. 1999, 80, 1627–1634. [Google Scholar]
- Herbert, R.; Baron, J.; Batten, C.; Baron, M.; Taylor, G. Recombinant adenovirus expressing the haemagglutinin of Peste des petits ruminants virus (PPRV) protects goats against challenge with pathogenic virus; a DIVA vaccine for PPR. Vet. Res. 2014, 45, 24. [Google Scholar] [CrossRef]
- Lund, B.T.; Tiwari, A.; Galbraith, S.; Baron, M.D.; Morrison, W.I.; Barrett, T. Vaccination of cattle with attenuated rinderpest virus stimulates CD4(+) T cell responses with broad viral antigen specificity. J. Gen. Virol. 2000, 81, 2137–2146. [Google Scholar]
- Gans, H.A.; Maldonado, Y.A. Loss of passively acquired maternal antibodies in highly vaccinated populations: An emerging need to define the ontogeny of infant immune responses. J. Infect. Dis. 2013, 208, 1–3. [Google Scholar] [CrossRef]
- Ata, F.A.; al Sumry, H.S.; King, G.J.; Ismaili, S.I.; Ata, A.A. Duration of maternal immunity to peste des petits ruminants. Vet. Rec. 1989, 124, 590–591. [Google Scholar]
- Bassiri, M.; Ahmad, S.; Giavedoni, L.; Jones, L.; Saliki, J.T.; Mebus, C.; Yilma, T. Immunological responses of mice and cattle to baculovirus-expressed F and H proteins of rinderpest virus: Lack of protection in the presence of neutralizing antibody. J. Virol. 1993, 67, 1255–1261. [Google Scholar]
- Zohari, S.; Department of Virology, Immunobiology and Parasitology, National Veterinary Institute, Uppsala, Sweden. Personnel communication, 14 Febuary 2014.
- Balamurugan, V.; Sen, A.; Venkatesan, G.; Rajak, K.K.; Bhanuprakash, V.; Singh, R.K. Study on passive immunity: Time of vaccination in kids born to goats vaccinated against peste des petits ruminants. Virol. Sin. 2012, 27, 228–233. [Google Scholar] [CrossRef]
- Cosby, S.L.; Chieko, K.; Yamaguchi, K. Rinderpest and Peste Des Petits Ruminants Virus: Plague of Large and Small Ruminants; Biology of Animal Infections; Academic Press: London, UK, 2006; pp. 197–221. [Google Scholar]
- Borrow, P.; Oldstone, M.B. Measles virus-mononuclear cell interactions. Curr. Trop. Microbiol. Immunol. 1995, 191, 85–100. [Google Scholar]
- Yanagi, Y.; Takeda, M.; Ohno, S. Measles virus: Cellular receptors, tropism and pathogenesis. J. Gen. Virol. 2006, 87, 2767–2779. [Google Scholar] [CrossRef]
- Pope, R.A.; Parida, S.; Bailey, D.; Brownlie, J.; Barrett, T.; Banyard, A.C. Early events following experimental infection with peste-des-petits ruminants virus suggest immune cell targeting. PLoS One 2013, 8, e55830. [Google Scholar]
- Esolen, L.M.; Ward, B.J.; Moench, T.R.; Griffin, D.E. Infection of monocytes during Measles. J. Infect. Dis. 1993, 168, 47–52. [Google Scholar] [CrossRef]
- Osunkoya, B.O.; Ukaejiofo, E.O.; Ajayi, O.; Akinyemi, A.A. Evidence that circulating lymphocytes act as vehicles or viraemia in Measles. West Afr. J. Med. 1990, 9, 35–39. [Google Scholar]
- Kumar, P.; Tripathi, B.N.; Sharma, A.K.; Kumar, R.; Sreenivasa, B.P.; Singh, R.P.; Dhar, P.; Bandyopadhyay, S.K. Pathological and immunohistochemical study of experimental Peste des petits ruminants virus infection in goats. J. Vet. Med. B Infect. Dis. Vet. Public Health 2004, 51, 153–159. [Google Scholar] [CrossRef]
- Kul, O.; Kabakci, N.; Atmaca, H.T.; Ozkul, A. Natural peste des petits ruminants virus infection: Novel pathologic findings resembling other Morbillivirus infections. Vet. Pathol. 2007, 44, 479–486. [Google Scholar] [CrossRef]
- Mondal, B.; Sreenivasa, B.P.; Dhar, P.; Singh, R.P.; Bandyopadhyay, S.K. Apoptosis induced by peste des petits ruminants virus in goat peripheral blood mononuclear cells. Virus Res. 2001, 73, 113–119. [Google Scholar] [CrossRef]
- Olaleye, O.D.; Oyejide, A.; Ikede, B.O. Correlation of humoral immune response with clinical presentation, pulmonary lesions and mortality patterns of goats experimentally infected with peste des petits ruminants virus. Cytobios 1989, 57, 141–147. [Google Scholar]
- Couacy-Hymann, E.; Bodjo, C.; Danho, T.; Libeau, G.; Diallo, A. Evaluation of the virulence of some strains of Peste-des-petits-ruminants virus (PPRV) in experimentally infected West African dwarf goats. Vet. J. 2007, 173, 178–183. [Google Scholar] [CrossRef]
- Ugochukwu, E.I.; Agwu, C.O. Aerobic bacteria from nasal discharge of goats suffering from clinical PPR: Isolation and identification. Microbios. 1991, 65, 81–85. [Google Scholar]
- Galbraith, S.E.; McQuaid, S.; Hamill, L.; Pullen, L.; Barrett, T.; Cosby, S.L. Rinderpest and peste des petits ruminants viruses exhibit neurovirulence in mice. J. Neurovirol. 2002, 8, 45–52. [Google Scholar]
- Toplu, N.; Oguzoglu, T.C.; Albayrak, H. Dual infection of fetal and neonatal small ruminants with border disease virus and peste des petits ruminants virus (PPRV): Neuronal tropism of PPRV as a novel finding. J. Comp. Pathol. 2012, 146, 289–297. [Google Scholar] [CrossRef]
- El Harrak, M.; Touil, N.; Loutfi, C.; Hammouchi, M.; Parida, S.; Sebbar, G.; Chaffai, N.; Harif, B.; Messoudi, N.; Batten, C. A reliable and reproducible experimental challenge model for peste des petits ruminants virus. J. Clin. Microbiol. 2012, 50, 3738–3740. [Google Scholar] [CrossRef]
- Hammouchi, M.; Loutfi, C.; Sebbar, G.; Touil, N.; Chaffai, N.; Batten, C.; Harif, B.; Oura, C.; El Harrak, M. Experimental infection of Alpine goats with a Moroccan strain of peste des petits ruminants virus (PPRV). Vet. Microbiol. 2012, 160, 240–244. [Google Scholar] [CrossRef]
- Bundza, A.; Afshar, A.; Dukes, T.W.; Myers, D.J.; Dulac, G.C.; Becker, S.A. Experimental peste des petits ruminants (goat plague) in goats and sheep. Can. J. Vet. Res. 1988, 52, 46–52. [Google Scholar]
- Truong, T.; Boshra, H.; Embury-Hyatt, C.; Nfon, C.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Chetty, T.; Mather, A. Peste des petits ruminants virus tissue tropism and pathogenesis in sheep and goats following experimental infection. PLoS One 2014, 9, e87145. [Google Scholar] [CrossRef]
- Couacy-Hymann, E.; Bodjo, S.C.; Danho, T.; Koffi, M.Y.; Libeau, G.; Diallo, A. Early detection of viral excretion from experimentally infected goats with peste-des-petits ruminants virus. Prev. Vet. Med. 2007, 78, 85–88. [Google Scholar] [CrossRef]
- Ezeibe, M.C.; Okoroafor, O.N.; Ngene, A.A.; Eze, J.I.; Eze, I.C.; Ugonabo, J.A. Persistent detection of peste de petits ruminants antigen in the faeces of recovered goats. Trop. Anim. Health Prod. 2008, 40, 517–519. [Google Scholar] [CrossRef]
- Gulyaz, V.; Ozkul, A. Pathogenicity of a local peste des petits ruminants virus isolate in sheep in Turkey. Trop. Anim. Health Prod. 2005, 37, 541–547. [Google Scholar] [CrossRef]
- Minet, C.; Yami, M.; Egzabhier, B.; Gil, P.; Tangy, F.; Bremont, M.; Libeau, G.; Diallo, A.; Albina, E. Sequence analysis of the large (L) polymerase gene and trailer of the peste des petits ruminants virus vaccine strain Nigeria 75/1: Expression and use of the L protein in reverse genetics. Virus Res. 2009, 145, 9–17. [Google Scholar] [CrossRef]
- Hu, Q.; Chen, W.; Huang, K.; Baron, M.D.; Bu, Z. Rescue of recombinant peste des petits ruminants virus: Creation of a GFP-expressing virus and application in rapid virus neutralization test. Vet. Res. 2012, 43, 48. [Google Scholar] [CrossRef]
- Schneider-Schaulies, S.; Niewiesk, S.; Schneider-Schaulies, J.; ter Meulen, V. Measles virus induced immunosuppression: Targets and effector mechanisms. Curr. Mol. Med. 2001, 1, 163–181. [Google Scholar] [CrossRef]
- Jagtap, S.P.; Rajak, K.K.; Garg, U.K.; Sen, A.; Bhanuprakash, V.; Sudhakar, S.B.; Balamurugan, V.; Patel, A.; Ahuja, A.; Singh, R.K. Effect of immunosuppression on pathogenesis of peste des petits ruminants (PPRV) virus infection in goats. Microb. Pathog. 2012, 52, 217–226. [Google Scholar] [CrossRef]
- Yoneda, M.; Bandyopadhyay, S.K.; Shiotani, M.; Fujita, K.; Nuntaprasert, A.; Miura, R.; Baron, M.D.; Barrett, T.; Kai, C. Rinderpest virus H protein: Role in determining host range in rabbits. J. Gen. Virol. 2002, 83, 1457–1463. [Google Scholar]
- Yoneda, M.; Miura, R.; Barrett, T.; Tsukiyama-Kohara, K.; Kai, C. Rinderpest virus phosphoprotein gene is a major determinant of species-specific pathogenicity. J. Virol. 2004, 78, 6676–6681. [Google Scholar]
- Schlender, J.; Schnorr, J.J.; Spielhoffer, P.; Cathomen, T.; Cattaneo, R.; Billeter, M.A.; ter Meulen, V.; Schneider-Schaulies, S. Interaction of Measles virus glycoproteins with the surface of uninfected peripheral blood lymphocytes induces immunosuppression in vitro. Proc. Natl. Acad. Sci. USA 1996, 93, 13194–13199. [Google Scholar] [CrossRef]
- Heaney, J.; Barrett, T.; Cosby, S.L. Inhibition of in vitro leukocyte proliferation by morbilliviruses. J. Virol. 2002, 76, 3579–3584. [Google Scholar] [CrossRef]
- Yokota, S.; Saito, H.; Kubota, T.; Yokosawa, N.; Amano, K.; Fujii, N. Measles virus suppresses interferon-alpha signaling pathway: Suppression of JAK1 phosphorylation and association of viral accessory proteins, C and V, with interferon-alpha receptor complex. Virology 2003, 306, 135–146. [Google Scholar] [CrossRef]
- Kerdiles, Y.M.; Cherif, B.; Marie, J.C.; Tremillon, N.; Blanquier, B.; Libeau, G.; Diallo, A.; Wild, T.F.; Villiers, M.B.; Horvat, B. Immunomodulatory properties of Morbillivirus nucleoproteins. Viral Immunol. 2006, 19, 324–334. [Google Scholar] [CrossRef]
- Marie, J.C.; Kehren, J.; Trescol-Biemont, M.C.; Evlashev, A.; Valentin, H.; Walzer, T.; Tedone, R.; Loveland, B.; Nicolas, J.F.; Rabourdin-Combe, C. Mechanism of Measles virus-induced suppression of inflammatory immune responses. Immunity 2001, 14, 69–79. [Google Scholar] [CrossRef]
- Bao, J.; Wang, Z.; Li, L.; Wu, X.; Sang, P.; Wu, G.; Ding, G.; Suo, L.; Liu, C.; Wang, J. Detection and genetic characterization of peste des petits ruminants virus in free-living bharals (pseudois nayaur) in Tibet, China. Res. Vet. Sci. 2011, 90, 238–240. [Google Scholar] [CrossRef]
- Lefevre, P.C.; Diallo, A. Peste des petits ruminants. Rev. Sci. Tech. 1990, 9, 935–981. [Google Scholar]
- Furley, C.W.; Taylor, W.P.; Obi, T.U. An outbreak of peste des petits ruminants in a zoological collection. Vet. Rec. 1987, 121, 443–447. [Google Scholar]
- Balamurugan, V.; Sen, A.; Venkatesan, G.; Bhanot, V.; Yadav, V.; Bhanuprakash, V.; Singh, R.K. Peste des petits ruminants virus detected in tissues from an Asiatic lion (Panthera leo persica) belongs to Asian lineage IV. J. Vet. Sci. 2012, 13, 203–206. [Google Scholar] [CrossRef]
- Bidjeh, K.; Bornarel, P.; Imadine, M.; Lancelot, R. First- time isolation of the peste des petits ruminants (PPR) virus in Chad and experimental induction of the disease. Rev. Elev. Med. Vet. Pays. Trop. 1995, 48, 295–300. [Google Scholar]
- Couacy-Hymann, E.; Bodjo, C.; Danho, T.; Libeau, G.; Diallo, A. Surveillance of wildlife as a tool for monitoring rinderpest and peste des petits ruminants in West Africa. Rev. Sci. Tech. 2005, 24, 869–877. [Google Scholar]
- Yesilbag, K.; Yilmaz, Z.; Golcu, E.; Ozkul, A. Peste des petits ruminants outbreak in Western Turkey. Vet. Rec. 2005, 157, 260–261. [Google Scholar]
- Ozkul, A.; Akca, Y.; Alkan, F.; Barrett, T.; Karaoglu, T.; Dagalp, S.B.; Anderson, J.; Yesilbag, K.; Cokcaliskan, C.; Gencay, A. Prevalence, distribution, and host range of peste des petits ruminants virus, Turkey. Emerg. Infect. Dis. 2002, 8, 708–712. [Google Scholar] [CrossRef]
- Abraham, G.; Sintayehu, A.; Libeau, G.; Albina, E.; Roger, F.; Laekemariam, Y.; Abayneh, D.; Awoke, K.M. Antibody seroprevalences against peste des petits ruminants (PPR) virus in camels, cattle, goats and sheep in Ethiopia. Prev. Vet. Med. 2005, 70, 51–57. [Google Scholar] [CrossRef]
- Ayari-Fakhfakh, E.; Ghram, A.; Bouattour, A.; Larbi, I.; Gribaa-Dridi, L.; Kwiatek, O.; Bouloy, M.; Libeau, G.; Albina, E.; Cetre-Sossah, C. First serological investigation of peste-des-petits-ruminants and rift valley fever in Tunisia. Vet. J. 2011, 187, 402–404. [Google Scholar] [CrossRef]
- Delil, F.; Asfaw, Y.; Gebreegziabher, B. Prevalence of antibodies to peste des petits ruminants virus before and during outbreaks of the disease in Awash fentale district, Afar, Ethiopia. Trop. Anim. Health Prod. 2012, 44, 1329–1330. [Google Scholar] [CrossRef]
- Al-Majali, A.M.; Hussain, N.O.; Amarin, N.M.; Majok, A.A. Seroprevalence of, and risk factors for, Peste des petits ruminants in sheep and goats in Northern Jordan. Prev. Vet. Med. 2008, 85, 1–8. [Google Scholar] [CrossRef]
- Mornet, P.; Orue, J.; Gilbert, Y.; Thiery, G.; Sow, M. La peste des petits ruminants en Afrique Occidentale Française. Ses rapports avec la peste bovine. Rev. Elev. Méd. Vét. Pays. Trop. 1956, 9, 313–342. [Google Scholar]
- Khalafalla, A.I.; Saeed, I.K.; Ali, Y.H.; Abdurrahman, M.B.; Kwiatek, O.; Libeau, G.; Obeida, A.A.; Abbas, Z. An outbreak of peste des petits ruminants (PPR) in camels in the Sudan. Acta Trop. 2010, 116, 161–165. [Google Scholar] [CrossRef]
- Megersa, B.; Biffa, D.; Abunna, F.; Regassa, A.; Bohlin, J.; Skjerve, E. Epidemic characterization and modeling within herd transmission dynamics of an "emerging trans-boundary" camel disease epidemic in Ethiopia. Trop. Anim. Health Prod. 2012, 44, 1643–1651. [Google Scholar] [CrossRef]
- Albina, E.; Kwiatek, O.; Minet, C.; Lancelot, R.; Servan de Almeida, R.; Libeau, G. Peste des petits ruminants, the next eradicated animal disease? Vet. Microbiol. 2013, 165, 38–44. [Google Scholar] [CrossRef]
- Capobianchi, M.R.; Giombini, E.; Rozera, G. Next-generation sequencing technology in clinical virology. Clin. Microbiol. Infect. 2013, 19, 15–22. [Google Scholar] [CrossRef]
- Kwiatek, O.; Ali, Y.H.; Saeed, I.K.; Khalafalla, A.I.; Mohamed, O.I.; Obeida, A.A.; Abdelrahman, M.B.; Osman, H.M.; Taha, K.M.; Abbas, Z. Asian lineage of peste des petits ruminants virus, Africa. Emerg. Infect. Dis. 2011, 17, 1223–1231. [Google Scholar]
- Shaila, M.S.; Shamaki, D.; Forsyth, M.A.; Diallo, A.; Goatley, L.; Kitching, R.P.; Barrett, T. Geographic distribution and epidemiology of peste des petits ruminants virus. Virus Res. 1996, 43, 149–153. [Google Scholar] [CrossRef]
- Kwiatek, O.; Minet, C.; Grillet, C.; Hurard, C.; Carlsson, E.; Karimov, B.; Albina, E.; Diallo, A.; Libeau, G. Peste des petits ruminants (PPR) outbreak in Tajikistan. J. Comp. Pathol. 2007, 136, 111–119. [Google Scholar] [CrossRef]
- Baron, M.D.; Banyard, A.C.; Parida, S.; Barrett, T. The plowright vaccine strain of rinderpest virus has attenuating mutations in most genes. J. Gen. Virol. 2005, 86, 1093–1101. [Google Scholar] [CrossRef]
- Rossiter, P.B.; Wardley, R.C. The differential growth of virulent and avirulent strains of rinderpest virus in bovine lymphocytes and macrophages. J. Gen. Virol. 1985, 66, 969–975. [Google Scholar] [CrossRef]
- Couacy-Hymann, E.; Bidjeh, K.; Angba, A.; Domenech, J.; Diallo, A. Protection of goats against rinderpest by vaccination with attenuated peste des petits ruminants virus. Res. Vet. Sci. 1995, 59, 106–109. [Google Scholar] [CrossRef]
- Sharon, D.; Chen, R.; Snyder, M. Systems biology approaches to disease marker discovery. Dis. Markers. 2010, 28, 209–224. [Google Scholar]
- Chakrabarti, A.K.; Vipat, V.C.; Mukherjee, S.; Singh, R.; Pawar, S.D.; Mishra, A.C. Host gene expression profiling in influenza a virus-infected lung epithelial (A549) cells: A comparative analysis between highly pathogenic and modified H5N1 viruses. Virol. J. 2010, 7, 219. [Google Scholar] [CrossRef]
- Rowland, A.C.; Bourdin, P. The histological relationship between "Peste des petits ruminants" and Kata in West Africa. Rev. Elev. Med. Vet. Pays Trop. 1970, 23, 301–307. [Google Scholar]
- Mariner, J.C.; House, J.A.; Mebus, C.A.; van den Ende, M.C. The use of thermostable Vero cell-adapted rinderpest vaccine as a heterologous vaccine against peste des petits ruminants. Res. Vet. Sci. 1993, 54, 212–216. [Google Scholar] [CrossRef]
- Sarkar, J.; Sreenivasa, B.P.; Singh, R.P.; Dhar, P.; Bandyopadhyay, S.K. Comparative efficacy of various chemical stabilizers on the thermostability of a live-attenuated peste des petits ruminants (PPR) vaccine. Vaccine 2003, 21, 4728–4735. [Google Scholar] [CrossRef]
- Singh, R.P.; De, U.K.; Pandey, K.D. Virological and antigenic characterization of two peste des petits ruminants (PPR) vaccine viruses of Indian origin. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 343–353. [Google Scholar] [CrossRef]
- Singh, R.P. Control strategies for peste des petits ruminants in small ruminants of India. Rev. Sci. Tech. Epiz. 2011, 30, 879–887. [Google Scholar]
- Saravanan, P.; Sen, A.; Balamurugan, V.; Bandyopadhyay, S.K.; Singh, R.K. Rapid quality control of a live attenuated peste des petits ruminants (PPR) vaccine by monoclonal antibody based sandwich ELISA. Biologicals 2008, 36, 1–6. [Google Scholar] [CrossRef]
- Rajak, K.K.; Sreenivasa, B.P.; Hosamani, M.; Singh, R.P.; Singh, S.K.; Singh, R.K.; Bandyopadhyay, S.K. Experimental studies on immunosuppressive effects of peste des petits ruminants (PPR) virus in goats. Comp. Immunol. Microbiol. Infect. Dis. 2005, 28, 287–296. [Google Scholar] [CrossRef]
- Worrall, E.E.; Litamoi, J.K.; Seck, B.M.; Ayelet, G. Xerovac: An ultra rapid method for the dehydration and preservation of live attenuated rinderpest and Peste des petits ruminants vaccines. Vaccine 2000, 19, 834–839. [Google Scholar] [CrossRef]
- Sen, A.; Saravanan, P.; Balamurugan, V.; Rajak, K.K.; Sudhakar, S.B.; Bhanuprakash, V.; Parida, S.; Singh, R.K. Vaccines against peste des petits ruminants virus. Expert Rev. Vaccines 2010, 9, 785–796. [Google Scholar] [CrossRef]
- Silva, A.C.; Carrondo, M.J.; Alves, P.M. Strategies for improved stability of peste des petits ruminants vaccine. Vaccine 2011, 29, 4983–4991. [Google Scholar] [CrossRef]
- Sinnathamby, G.; Naik, S.; Renukaradhya, G.J.; Rajasekhar, M.; Nayak, R.; Shaila, M.S. Recombinant hemagglutinin protein of rinderpest virus expressed in insect cells induces humoral and cell mediated immune responses in cattle. Vaccine 2001, 19, 3870–3876. [Google Scholar] [CrossRef]
- Romero, C.H.; Barrett, T.; Chamberlain, R.W.; Kitching, R.P.; Fleming, M.; Black, D.N. Recombinant capripoxvirus expressing the hemagglutinin protein gene of rinderpest virus: Protection of cattle against rinderpest and lumpy skin disease viruses. Virology 1994, 204, 425–429. [Google Scholar] [CrossRef]
- Belsham, G.J.; Anderson, E.C.; Murray, P.K.; Anderson, J.; Barrett, T. Immune response and protection of cattle and pigs generated by a vaccinia virus recombinant expressing the F protein of rinderpest virus. Vet. Rec. 1989, 124, 655–658. [Google Scholar]
- Berhe, G.; Minet, C.; Le Goff, C.; Barrett, T.; Ngangnou, A.; Grillet, C.; Libeau, G.; Fleming, M.; Black, D.N.; Diallo, A. Development of a dual recombinant vaccine to protect small ruminants against peste-des-petits-ruminants virus and capripoxvirus infections. J. Virol. 2003, 77, 1571–1577. [Google Scholar] [CrossRef]
- Chen, W.; Hu, S.; Qu, L.; Hu, Q.; Zhang, Q.; Zhi, H.; Huang, K.; Bu, Z. A goat poxvirus-vectored peste-des-petits-ruminants vaccine induces long-lasting neutralization antibody to high levels in goats and sheep. Vaccine 2010, 28, 4742–4750. [Google Scholar] [CrossRef]
- Diallo, A.; Minet, C.; Berhe, G.; Le Goff, C.; Black, D.N.; Fleming, M.; Barrett, T.; Grillet, C.; Libeau, G. Goat immune response to capripox vaccine expressing the hemagglutinin protein of peste des petits ruminants. Ann. N. Y. Acad. Sci. 2002, 969, 88–91. [Google Scholar] [CrossRef]
- Das, S.C.; Baron, M.D.; Barrett, T. Recovery and characterization of a chimeric rinderpest virus with the glycoproteins of peste-des-petits-ruminants virus: Homologous F and H proteins are required for virus viability. J. Virol. 2000, 74, 9039–9047. [Google Scholar] [CrossRef]
- Parida, S.; Mahapatra, M.; Kumar, S.; Das, S.C.; Baron, M.D.; Anderson, J.; Barrett, T. Rescue of a chimeric rinderpest virus with the nucleocapsid protein derived from peste-des-petits-ruminants virus: Use as a marker vaccine. J. Gen. Virol. 2007, 88, 2019–2027. [Google Scholar] [CrossRef]
- Chandran, D.; Reddy, K.B.; Vijayan, S.P.; Sugumar, P.; Rani, G.S.; Kumar, P.S.; Rajendra, L.; Srinivasan, V.A. MVA recombinants expressing the fusion and hemagglutinin genes of PPRV protects goats against vilrulent challenge. Ind. J. Microbol. 2010, 50, 266–274. [Google Scholar] [CrossRef]
- Cubillos-Zapata, C.; Guzman, E.; Turner, A.; Gilbert, S.C.; Prentice, H.; Hope, J.C.; Charleston, B. Differential effects of viral vectors on migratory afferent lymph dendritic cells in vitro predict enhanced immunogenicity in vivo. J. Virol. 2011, 85, 9385–9394. [Google Scholar] [CrossRef]
- Rahman, M.M.; Shaila, M.S.; Gopinathan, K.P. Baculovirus display of fusion protein of peste des petits ruminants virus and hemagglutination protein of rinderpest virus and immunogenicity of the displayed proteins in mouse model. Virology 2003, 317, 36–49. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, G.; Shi, L.; Li, W.; Li, C.; Chen, Z.; Jin, H.; Xu, B.; Li, G. Immune responses in mice vaccinated with a suicidal DNA vaccine expressing the hemagglutinin glycoprotein from the peste des petits ruminants virus. J. Virol. Methods 2013, 193, 525–530. [Google Scholar] [CrossRef]
- Kaufman, D.R.; Goudsmit, J.; Holterman, L.; Ewald, B.A.; Denholtz, M.; Devoy, C.; Giri, A.; Grandpre, L.E.; Heraud, J.M.; Franchini, G.; et al. Differential antigen requirements for protection against systemic and intranasal vaccinia virus challenges in mice. J. Virol. 2008, 82, 6829–6837. [Google Scholar] [CrossRef]
- Geutskens, S.B.; van der Eb, M.M.; Plomp, A.C.; Jonges, L.E.; Cramer, S.J.; Ensink, N.G.; Kuppen, P.J.; Hoeben, R.C. Recombinant adenoviral vectors have adjuvant activity and stimulate T cell responses against tumor cells. Gene Ther. 2000, 7, 1410–1416. [Google Scholar] [CrossRef]
- Abaitua, F.; Rodriguez, J.R.; Garzon, A.; Rodriguez, D.; Esteban, M. Improving recombinant MVA immune responses: Potentiation of the immune responses to HIV-1 with MVA and DNA vectors expressing env and the cytokines IL-12 and IFN-gamma. Virus Res. 2006, 116, 11–20. [Google Scholar] [CrossRef]
- Thacker, E.E.; Nakayama, M.; Smith, B.F.; Bird, R.C.; Muminova, Z.; Strong, T.V.; Timares, L.; Korokhov, N.; O'Neill, A.M.; de Gruijl, T.D.; et al. A genetically engineered adenovirus vector targeted to CD40 mediates transduction of canine dendritic cells and promotes antigen-specific immune responses in vivo. Vaccine 2009, 27, 7116–7124. [Google Scholar] [CrossRef]
- Cruz, P.E.; Silva, A.C.; Roldao, A.; Carmo, M.; Carrondo, M.J.; Alves, P.M. Screening of novel excipients for improving the stability of retroviral and adenoviral vectors. Biotechnol. Prog. 2006, 22, 568–576. [Google Scholar] [CrossRef]
- Lameiro, M.H.; Malpique, R.; Silva, A.C.; Alves, P.M.; Melo, E. Encapsulation of adenoviral vectors into chitosan-bile salt microparticles for mucosal vaccination. J. Biotechnol. 2006, 126, 152–162. [Google Scholar] [CrossRef]
- Ferreira, T.B.; Ferreira, A.L.; Carrondo, M.J.; Alves, P.M. Effect of re-feed strategies and non-ammoniagenic medium on adenovirus production at high cell densities. J. Biotechnol. 2005, 119, 272–280. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, G.; Chen, Z.; Li, C.; Shi, L.; Li, W.; Huang, H.; Tao, C.; Cheng, C.; Xu, B.; et al. Recombinant adenovirus expressing F and H fusion proteins of Peste des petits ruminants virus induces both humoral and cell-mediated immune responses in goats. Vet. Immunol. Immunopathol. 2013, 154, 1–7. [Google Scholar] [CrossRef]
- Qin, J.; Huang, H.; Ruan, Y.; Hou, X.; Yang, S.; Wang, C.; Huang, G.; Wang, T.; Feng, N.; Gao, Y.; et al. A novel recombinant peste des petits ruminants-canine adenovirus vaccine elicits long-lasting neutralizing antibody response against PPR in goats. PLoS One 2012, 7, e37170. [Google Scholar] [CrossRef]
- Saravanan, P.; Sen, A.; Balamurugan, V.; Rajak, K.K.; Bhanuprakash, V.; Palaniswami, K.S.; Nachimuthu, K.; Thangavelu, A.; Dhinakarraj, G.; Hegde, R.; et al. Comparative efficacy of Peste des petits ruminants (PPR) vaccines. Biologicals 2010, 38, 479–485. [Google Scholar] [CrossRef]
- Hosamani, M.; Singh, S.K.; Mondal, B.; Sen, A.; Bhanuprakash, V.; Bandyopadhyay, S.K.; Yadav, M.P.; Singh, R.K. A bivalent vaccine against goat pox and peste des petits ruminants induces protective immune response in goats. Vaccine 2006, 24, 6058–6064. [Google Scholar] [CrossRef]
- Chaudhary, S.S.; Pandey, K.D.; Singh, R.P.; Verma, P.C.; Gupta, P.K. A vero cell derived combined vaccine against sheep pox and peste des petits ruminants for sheep. Vaccine 2009, 27, 2548–2553. [Google Scholar] [CrossRef]
- Horzinek, M.C. Rinderpest: The second viral disease eradicated. Vet. Microbiol. 2011, 149, 295–297. [Google Scholar] [CrossRef]
- Couacy-Hymann, E.; Bodjo, S.C.; Danho, T. Interference in the vaccination of cattle against rinderpest virus by antibodies against peste des petits ruminants (PPR) virus. Vaccine 2006, 24, 5679–5683. [Google Scholar] [CrossRef]
- Rossister, P. Progressive control of Peste des petits ruminants. In Proceedings of the E-Conference to Launch Global Research and Expertise Network (GREN) on PPR, Online. 3–28 February 2014.
- Goris, N.; Vandenbussche, F.; De Clercq, K. Potential of antiviral therapy and prophylaxis for controlling RNA viral infections of livestock. Antivir. Res. 2008, 78, 170–178. [Google Scholar] [CrossRef]
- Charleston, B.; Bankowski, B.M.; Gubbins, S.; Chase-Topping, M.E.; Schley, D.; Howey, R.; Barnett, P.V.; Gibson, D.; Juleff, N.D.; Woolhouse, M.E. Relationship between clinical signs and transmission of an infectious disease and the implications for control. Science 2011, 332, 726–729. [Google Scholar] [CrossRef]
- Raheel, R.; Ashraf, M.; Ejaz, S.; Javeed, A.; Altaf, I. Assessment of the cytotoxic and anti-viral potential of aqueous extracts from different parts of Acacia nilotica against peste des petits ruminants virus. Environ. Toxicol. Pharmacol. 2013, 35, 72–81. [Google Scholar] [CrossRef]
- Nizamani, Z.A.; Keil, G.M.; Albina, E.; Holz, C.; Minet, C.; Kwiatek, O.; Libeau, G.; Servan de Almeida, R. Potential of adenovirus and baculovirus vectors for the delivery of shRNA against Morbilliviruses. Antivir. Res. 2011, 90, 98–101. [Google Scholar] [CrossRef]
- Kumar, N.; Liang, Y.; Parslow, T.G. Receptor tyrosine kinase inhibitors block multiple steps of influenza A virus replication. J. Virol. 2011, 85, 2818–2827. [Google Scholar] [CrossRef]
- Kumar, N.; Sharma, N.R.; Ly, H.; Parslow, T.G.; Liang, Y. Receptor tyrosine kinase inhibitors that block replication of influenza A and other viruses. Antimicro. Agents Chemother. 2011, 55, 5553–5559. [Google Scholar] [CrossRef]
- Kumar, N.; Xin, Z.T.; Liang, Y.; Ly, H. NF-kappaB signaling differentially regulates influenza virus RNA synthesis. J. Virol. 2008, 82, 9880–9889. [Google Scholar] [CrossRef]
- Pleschka, S.; Wolff, T.; Ehrhardt, C.; Hobom, G.; Planz, O.; Rapp, U.R.; Ludwig, S. Influenza virus propagation is impaired by inhibition of the RAF/MEK/ERK signalling cascade. Nat. Cell Biol. 2001, 3, 301–305. [Google Scholar] [CrossRef]
- Borgeling, Y.; Schmolke, M.; Viemann, D.; Nordhoff, C.; Roth, J.; Ludwig, S. Inhibition of p38 mitogen-activated protein kinase impairs influenza virus-induced primary and secondary host gene responses and protects mice from lethal H5N1 infection. J. Biol. Chem. 2014, 289, 13–27. [Google Scholar]
- Hrincius, E.R.; Dierkes, R.; Anhlan, D.; Wixler, V.; Ludwig, S.; Ehrhardt, C. Phosphatidylinositol-3-kinase (PI3K) is activated by influenza virus vRNA via the pathogen pattern receptor RIG-I to promote efficient type I interferon production. Cell. Microbiol. 2011, 13, 1907–1919. [Google Scholar] [CrossRef]
- Kumar, N.; Maherchandani, S. Targeting host cell factors for development of antiviral therapeutics. Adv. Anim. Vet. Sci. 2014, 1S, 37–41. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kumar, N.; Maherchandani, S.; Kashyap, S.K.; Singh, S.V.; Sharma, S.; Chaubey, K.K.; Ly, H. Peste Des Petits Ruminants Virus Infection of Small Ruminants: A Comprehensive Review. Viruses 2014, 6, 2287-2327. https://doi.org/10.3390/v6062287
Kumar N, Maherchandani S, Kashyap SK, Singh SV, Sharma S, Chaubey KK, Ly H. Peste Des Petits Ruminants Virus Infection of Small Ruminants: A Comprehensive Review. Viruses. 2014; 6(6):2287-2327. https://doi.org/10.3390/v6062287
Chicago/Turabian StyleKumar, Naveen, Sunil Maherchandani, Sudhir Kumar Kashyap, Shoor Vir Singh, Shalini Sharma, Kundan Kumar Chaubey, and Hinh Ly. 2014. "Peste Des Petits Ruminants Virus Infection of Small Ruminants: A Comprehensive Review" Viruses 6, no. 6: 2287-2327. https://doi.org/10.3390/v6062287