Identification of Cis-Acting Elements on Positive-Strand Subgenomic mRNA Required for the Synthesis of Negative-Strand Counterpart in Bovine Coronavirus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Plasmid Constructs
2.3. RT-PCR for Detecting (−)-Strand RNA Products
2.4. Quantitation of (−)-Strand sgmRNA Synthesis by RT-qPCR
2.5. Northern Blot Assay for DI RNA Replication
2.6. Statistical Analyses
3. Results
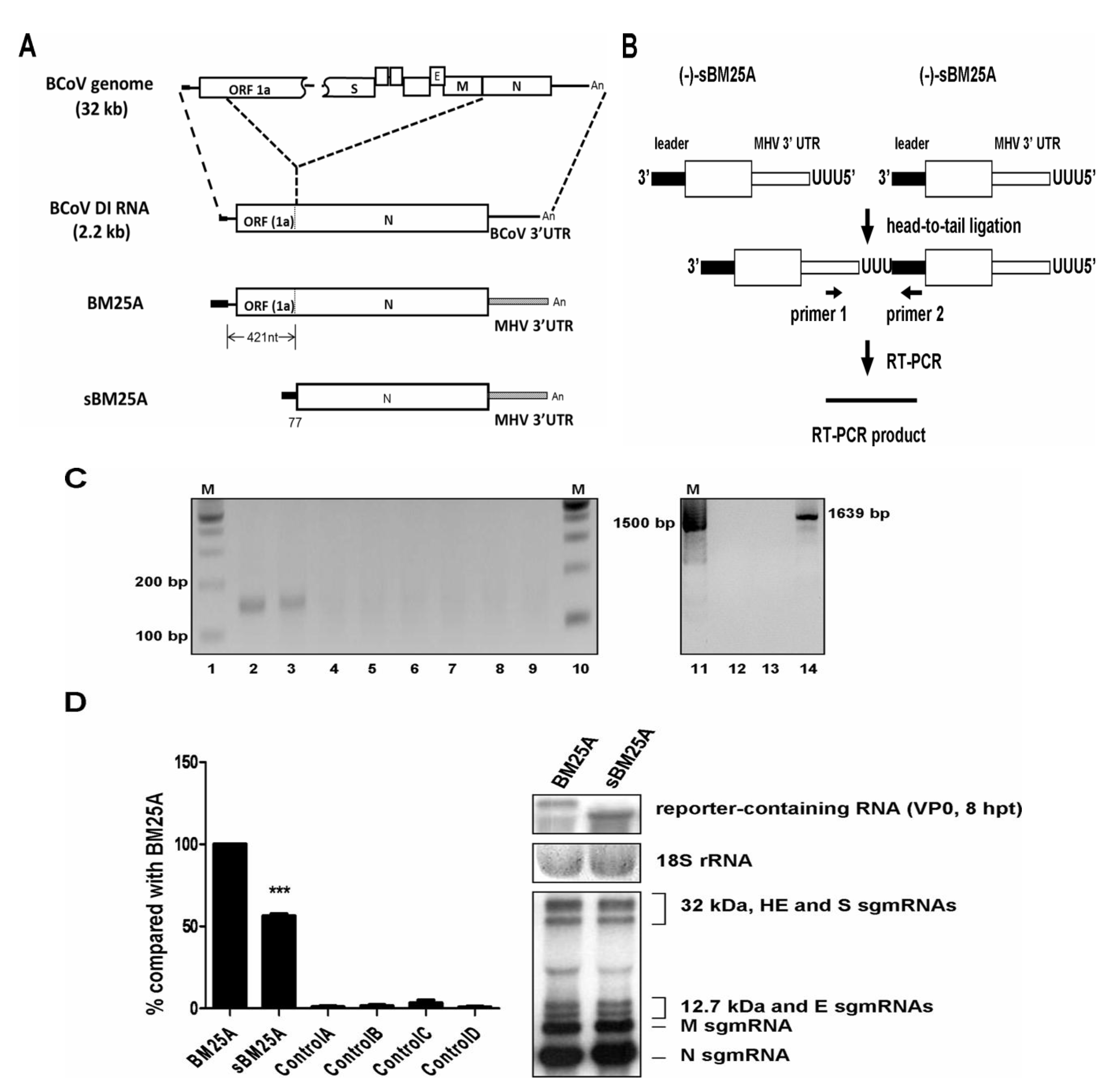
3.1. The (−)-Strand Synthesis from sgmRNA 7 Is ~2-Fold less than that from BCoV DI RNA, a Surrogate for BCoV Genome

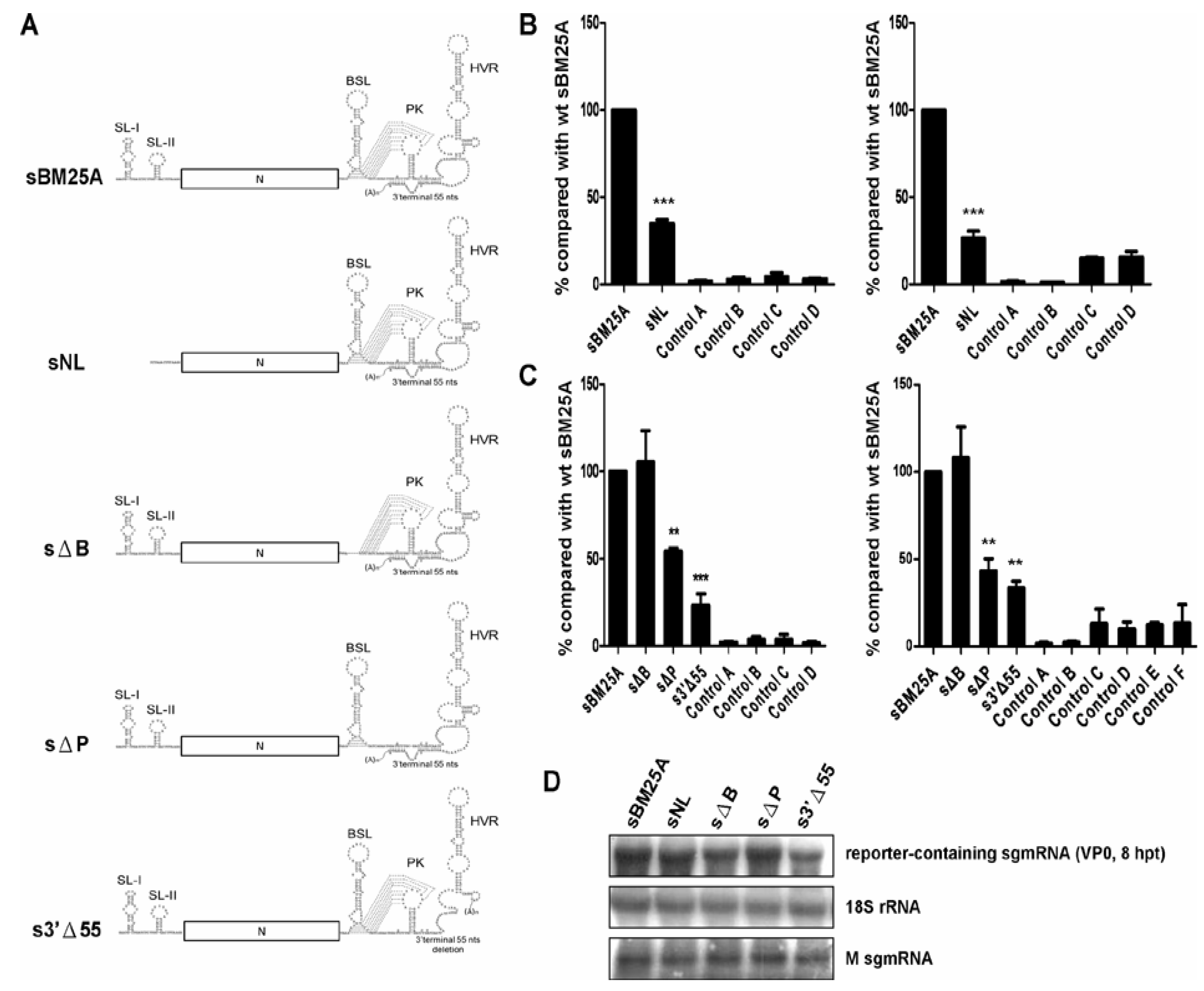
3.2. Screening for cis-Acting Elements in the 5' and 3' UTRs Required for (−)-Strand sgmRNA 7 Synthesis
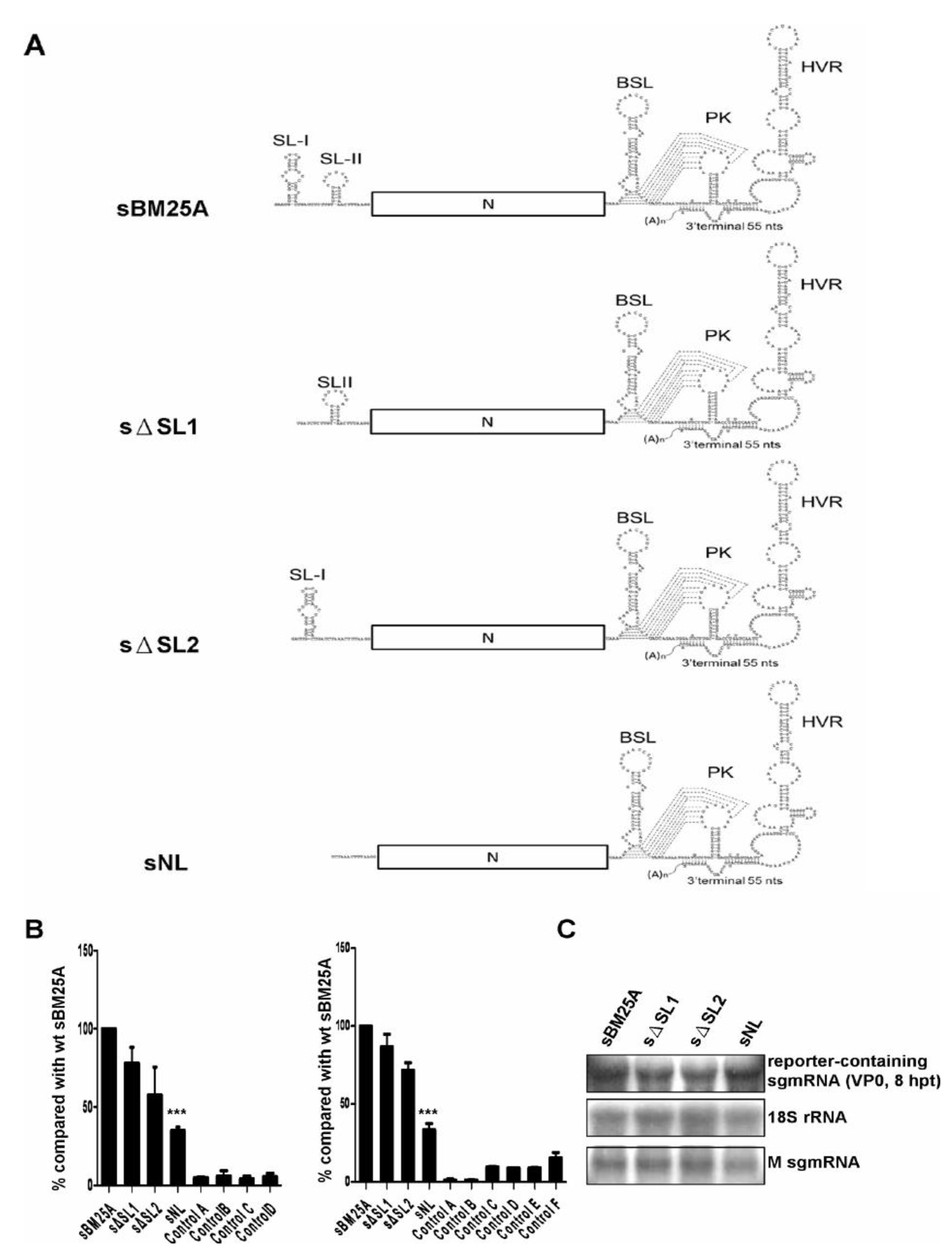
3.3. Deletion of Individual Stem-Loops within the Leader Sequence of sgmRNA 7 only Shows Minor Effects on (−)-Strand sgmRNA Synthesis


3.4. The 3'-Terminal nts Positioned from −15 to −34 is Required for Efficient (−)-Strand sgmRNA Synthesis
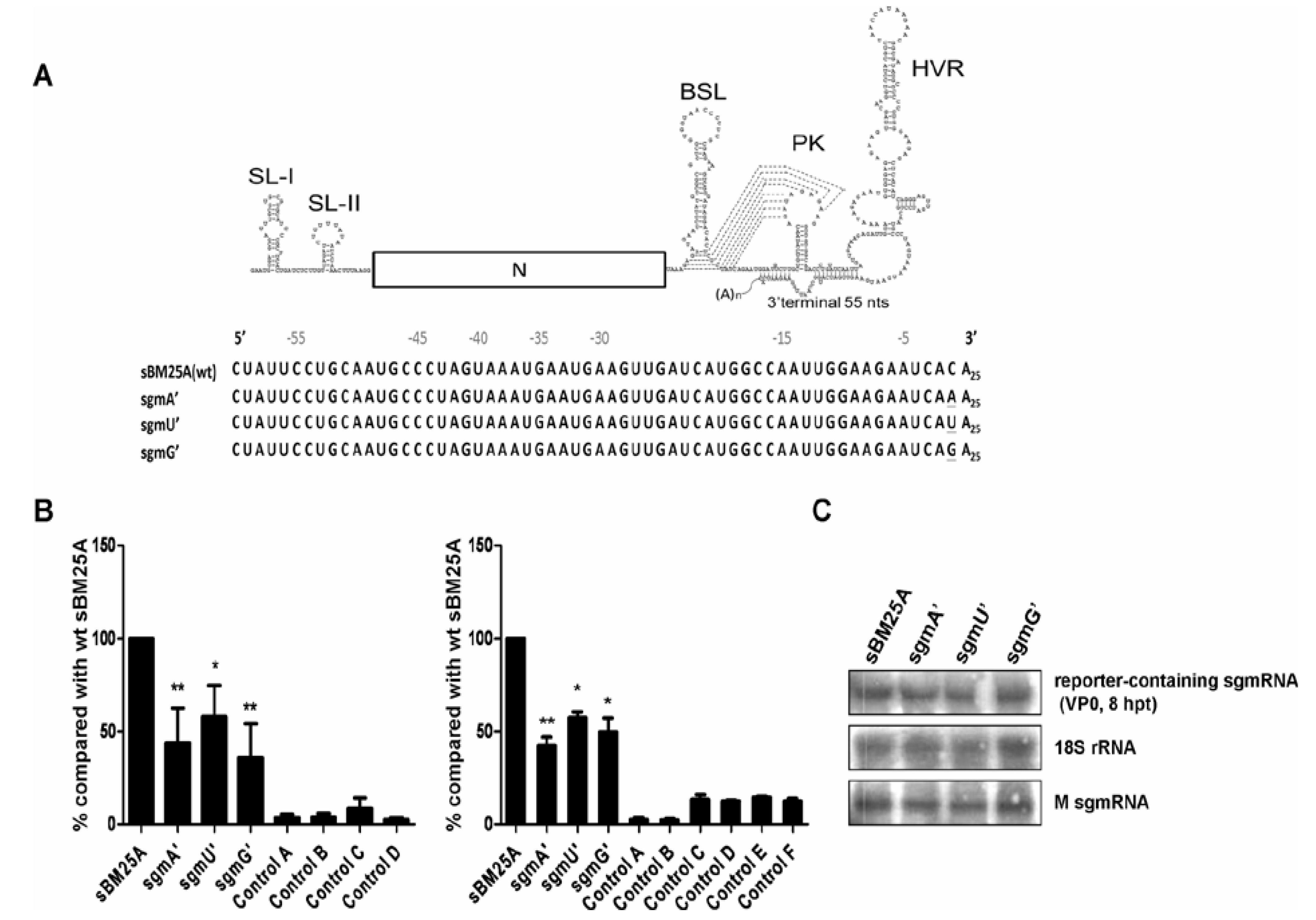
3.5. The 3'-Most Nucleotide Species of sgmRNA 7 Affects the Efficiency of (−)-Strand sgmRNA Synthesis


4. Discussion
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Brian, D.A.; Baric, R.S. Coronavirus genome structure and replication. Curr. Top. Microbiol. Immunol. 2005, 287, 1–30. [Google Scholar] [PubMed]
- Enjuanes, L.; Almazan, F.; Sola, I.; Zuniga, S. Biochemical aspects of coronavirus replication and virus-host interaction. Annu. Rev. Microbiol. 2006, 60, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, A.O.; Spaan, W.J.; Snijder, E.J. Nidovirus transcription: How to make sense...? J. Gen. Virol. 2006, 87, 1403–1421. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, A.L.; Smits, S.L.; Rottier, P.J.; de Groot, R.J. Discontinuous and non-discontinuous subgenomic rna transcription in a nidovirus. EMBO J. 2002, 21, 6571–6580. [Google Scholar] [CrossRef] [PubMed]
- Ozdarendeli, A.; Ku, S.; Rochat, S.; Williams, G.D.; Senanayake, S.D.; Brian, D.A. Downstream sequences influence the choice between a naturally occurring noncanonical and closely positioned upstream canonical heptameric fusion motif during bovine coronavirus subgenomic mrna synthesis. J. Virol. 2001, 75, 7362–7374. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, S.G.; Sawicki, D.L. Coronavirus transcription: Subgenomic mouse hepatitis virus replicative intermediates function in rna synthesis. J. Virol. 1990, 64, 1050–1056. [Google Scholar] [PubMed]
- Sawicki, S.G.; Sawicki, D.L. Coronavirus transcription: A perspective. Curr. Top. Microbiol. Immunol. 2005, 287, 31–55. [Google Scholar] [PubMed]
- Sethna, P.B.; Hung, S.L.; Brian, D.A. Coronavirus subgenomic minus-strand rnas and the potential for mrna replicons. Proc. Natl. Acad. Sci. USA 1989, 86, 5626–5630. [Google Scholar] [CrossRef] [PubMed]
- Sola, I.; Moreno, J.L.; Zuniga, S.; Alonso, S.; Enjuanes, L. Role of nucleotides immediately flanking the transcription-regulating sequence core in coronavirus subgenomic mrna synthesis. J. Virol. 2005, 79, 2506–2516. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Brian, D.A. 5'-proximal hot spot for an inducible positive-to-negative-strand template switch by coronavirus RNA-dependent RNA polymerase. J. Virol. 2007, 81, 3206–3215. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Ozdarendeli, A.; Brian, D.A. Bovine coronavirus 5'-proximal genomic acceptor hotspot for discontinuous transcription is 65 nucleotides wide. J. Virol. 2006, 80, 2183–2193. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, S.; Sola, I.; Alonso, S.; Enjuanes, L. Sequence motifs involved in the regulation of discontinuous coronavirus subgenomic RNA synthesis. J. Virol. 2004, 78, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.Y.; Hofmann, M.A.; Sethna, P.B.; Brian, D.A. A cis-acting function for the coronavirus leader in defective interfering RNA replication. J. Virol. 1994, 68, 8223–8231. [Google Scholar] [PubMed]
- Wu, H.Y.; Brian, D.A. Subgenomic messenger RNA amplification in coronaviruses. Proc. Natl. Acad. Sci. USA 2010, 107, 12257–12262. [Google Scholar] [CrossRef] [PubMed]
- Baric, R.S.; Fu, K.; Schaad, M.C.; Stohlman, S.A. Establishing a genetic recombination map for murine coronavirus strain a59 complementation groups. Virology 1990, 177, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Baric, R.S. Map locations of mouse hepatitis virus temperature-sensitive mutants: Confirmation of variable rates of recombination. J. Virol. 1994, 68, 7458–7466. [Google Scholar] [PubMed]
- Koetzner, C.A.; Parker, M.M.; Ricard, C.S.; Sturman, L.S.; Masters, P.S. Repair and mutagenesis of the genome of a deletion mutant of the coronavirus mouse hepatitis virus by targeted RNA recombination. J. Virol. 1992, 66, 1841–1848. [Google Scholar] [PubMed]
- Masters, P.S.; Koetzner, C.A.; Kerr, C.A.; Heo, Y. Optimization of targeted RNA recombination and mapping of a novel nucleocapsid gene mutation in the coronavirus mouse hepatitis virus. J. Virol. 1994, 68, 328–337. [Google Scholar] [PubMed]
- Fu, K.; Baric, R.S. Evidence for variable rates of recombination in the mhv genome. Virology 1992, 189, 88–102. [Google Scholar] [CrossRef]
- Garcia-Arriaza, J.; Ojosnegros, S.; Davila, M.; Domingo, E.; Escarmis, C. Dynamics of mutation and recombination in a replicating population of complementing, defective viral genomes. J. Mol. Biol. 2006, 360, 558–572. [Google Scholar] [CrossRef] [PubMed]
- Eckerle, L.D.; Lu, X.; Sperry, S.M.; Choi, L.; Denison, M.R. High fidelity of murine hepatitis virus replication is decreased in nsp14 exoribonuclease mutants. J. Virol. 2007, 81, 12135–12144. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.G.; Nixon, K.S.; Senanayake, S.D.; Brian, D.A. An RNA stem-loop within the bovine coronavirus nsp1 coding region is a cis-acting element in defective interfering RNA replication. J. Virol. 2007, 81, 7716–7724. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.Y.; Krishnan, R.; Brian, D.A. The ucuaaac promoter motif is not required for high-frequency leader recombination in bovine coronavirus defective interfering RNA. J. Virol. 1996, 70, 2720–2729. [Google Scholar] [PubMed]
- Kang, H.; Feng, M.; Schroeder, M.E.; Giedroc, D.P.; Leibowitz, J.L. Putative cis-acting stem-loops in the 5' untranslated region of the severe acute respiratory syndrome coronavirus can substitute for their mouse hepatitis virus counterparts. J. Virol. 2006, 80, 10600–10614. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Lai, M.M. Deletion mapping of a mouse hepatitis virus defective interfering RNA reveals the requirement of an internal and discontiguous sequence for replication. J. Virol. 1993, 67, 6110–6118. [Google Scholar] [PubMed]
- Lin, Y.J.; Liao, C.L.; Lai, M.M. Identification of the cis-acting signal for minus-strand RNA synthesis of a murine coronavirus: Implications for the role of minus-strand RNA in RNA replication and transcription. J. Virol. 1994, 68, 8131–8140. [Google Scholar] [PubMed]
- Raman, S.; Bouma, P.; Williams, G.D.; Brian, D.A. Stem-loop iii in the 5' untranslated region is a cis-acting element in bovine coronavirus defective interfering RNA replication. J. Virol. 2003, 77, 6720–6730. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.; Brian, D.A. Stem-loop iv in the 5' untranslated region is a cis-acting element in bovine coronavirus defective interfering RNA replication. J. Virol. 2005, 79, 12434–12446. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, J.F.; Hogue, B.G. Host protein interactions with the 3' end of bovine coronavirus RNA and the requirement of the poly(a) tail for coronavirus defective genome replication. J. Virol. 2000, 74, 5053–5065. [Google Scholar] [CrossRef] [PubMed]
- Van der Most, R.G.; Luytjes, W.; Rutjes, S.; Spaan, W.J. Translation but not the encoded sequence is essential for the efficient propagation of the defective interfering rnas of the coronavirus mouse hepatitis virus. J. Virol. 1995, 69, 3744–3751. [Google Scholar] [PubMed]
- Williams, G.D.; Chang, R.Y.; Brian, D.A. A phylogenetically conserved hairpin-type 3' untranslated region pseudoknot functions in coronavirus RNA replication. J. Virol. 1999, 73, 8349–8355. [Google Scholar] [PubMed]
- Liao, W.Y.; Ke, T.Y.; Wu, H.Y. The 3'-terminal 55 nucleotides of bovine coronavirus defective interfering RNA harbor cis-acting elements required for both negative- and positive-strand RNA synthesis. PLoS One 2014, 9, e98422. [Google Scholar] [CrossRef] [PubMed]
- Gustin, K.M.; Guan, B.J.; Dziduszko, A.; Brian, D.A. Bovine coronavirus nonstructural protein 1 (p28) is an RNA binding protein that binds terminal genomic cis-replication elements. J. Virol. 2009, 83, 6087–6097. [Google Scholar] [CrossRef] [PubMed]
- Hsue, B.; Hartshorne, T.; Masters, P.S. Characterization of an essential RNA secondary structure in the 3' untranslated region of the murine coronavirus genome. J. Virol. 2000, 74, 6911–6921. [Google Scholar] [CrossRef] [PubMed]
- Hsue, B.; Masters, P.S. A bulged stem-loop structure in the 3' untranslated region of the genome of the coronavirus mouse hepatitis virus is essential for replication. J. Virol. 1997, 71, 7567–7578. [Google Scholar] [PubMed]
- Goebel, S.J.; Miller, T.B.; Bennett, C.J.; Bernard, K.A.; Masters, P.S. A hypervariable region within the 3' cis-acting element of the murine coronavirus genome is nonessential for RNA synthesis but affects pathogenesis. J. Virol. 2007, 81, 1274–1287. [Google Scholar] [CrossRef] [PubMed]
- Lapps, W.; Hogue, B.G.; Brian, D.A. Sequence analysis of the bovine coronavirus nucleocapsid and matrix protein genes. Virology 1987, 157, 47–57. [Google Scholar] [CrossRef] [PubMed]
- King, B.; Brian, D.A. Bovine coronavirus structural proteins. J. Virol. 1982, 42, 700–707. [Google Scholar] [PubMed]
- Wu, H.Y.; Ke, T.Y.; Liao, W.Y.; Chang, N.Y. Regulation of coronaviral poly(a) tail length during infection. PLoS One 2013, 8, e70548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GraphPad Prism, version 2.01; GraphPad Software, Inc.: San Diego, CA, USA, 1996.
- Hofmann, M.A.; Sethna, P.B.; Brian, D.A. Bovine coronavirus mrna replication continues throughout persistent infection in cell culture. J. Virol. 1990, 64, 4108–4114. [Google Scholar] [PubMed]
- Schenborn, E.T.; Mierendorf, R.C., Jr. A novel transcription property of sp6 and t7 RNA polymerases: Dependence on template structure. Nucleic Acids Res. 1985, 13, 6223–6236. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M. Genetic recombination in RNA viruses. Curr. Top. Microbiol. Immunol. 1992, 176, 21–32. [Google Scholar] [PubMed]
- Ke, T.Y.; Liao, W.Y.; Wu, H.Y. A leaderless genome identified during persistent bovine coronavirus infection is associated with attenuation of gene expression. PLoS One 2013, 8, e82176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zust, R.; Miller, T.B.; Goebel, S.J.; Thiel, V.; Masters, P.S. Genetic interactions between an essential 3' cis-acting RNA pseudoknot, replicase gene products, and the extreme 3' end of the mouse coronavirus genome. J. Virol. 2008, 82, 1214–1228. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kang, H.; Liu, P.; Makkinje, N.; Williamson, S.T.; Leibowitz, J.L.; Giedroc, D.P. Structural lability in stem-loop 1 drives a 5' utr-3' utr interaction in coronavirus replication. J. Mol. Biol. 2008, 377, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, L.; Keane, S.C.; Yang, D.; Leibowitz, J.L.; Giedroc, D.P. Mouse hepatitis virus stem-loop 2 adopts a uynmg(u)a-like tetraloop structure that is highly functionally tolerant of base substitutions. J. Virol. 2009, 83, 12084–12093. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, L.; Millership, J.J.; Kang, H.; Leibowitz, J.L.; Giedroc, D.P. A u-turn motif-containing stem-loop in the coronavirus 5' untranslated region plays a functional role in replication. RNA 2007, 13, 763–780. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Johnson, R.F.; Leibowitz, J.L. Secondary structural elements within the 3' untranslated region of mouse hepatitis virus strain jhm genomic RNA. J. Virol. 2001, 75, 12105–12113. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.J.; Su, Y.P.; Wu, H.Y.; Brian, D.A. Genetic evidence of a long-range RNA-RNA interaction between the genomic 5' untranslated region and the nonstructural protein 1 coding region in murine and bovine coronaviruses. J. Virol. 2012, 86, 4631–4643. [Google Scholar] [CrossRef] [PubMed]
- Guan, B.J.; Wu, H.Y.; Brian, D.A. An optimal cis-replication stem-loop iv in the 5' untranslated region of the mouse coronavirus genome extends 1. J. Virol. 2011, 85, 5593–5605. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.Y.; Brian, D.A. Cis requirement for n-specific protein sequence in bovine coronavirus defective interfering RNA replication. J. Virol. 1996, 70, 2201–2207. [Google Scholar] [PubMed]
- Liao, C.L.; Lai, M.M. A cis-acting viral protein is not required for the replication of a coronavirus defective-interfering RNA. Virology 1995, 209, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yang, D.; Carter, K.; Masud, F.; Leibowitz, J.L. Functional analysis of the stem loop s3 and s4 structures in the coronavirus 3'utr. Virology 2013, 443, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.E.; Filomatori, C.V.; Gamarnik, A.V. Functional analysis of dengue virus cyclization sequences located at the 5' and 3'utrs. Virology 2008, 375, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, D.E.; Lodeiro, M.F.; Luduena, S.J.; Pietrasanta, L.I.; Gamarnik, A.V. Long-range RNA-RNA interactions circularize the dengue virus genome. J. Virol. 2005, 79, 6631–6643. [Google Scholar] [CrossRef] [PubMed]
- Enjuanes, L.; Almazan, F.; Sola, I.; Zuniga, S.; Alvarez, E.; Reguera, J.; Capiscol, C. Biochemical aspects of coronavirus replication. Adv. Exp. Med. Biol. 2006, 581, 13–24. [Google Scholar] [PubMed]
- Filomatori, C.V.; Lodeiro, M.F.; Alvarez, D.E.; Samsa, M.M.; Pietrasanta, L.; Gamarnik, A.V. A 5' RNA element promotes dengue virus RNA synthesis on a circular genome. Genes Dev. 2006, 20, 2238–2249. [Google Scholar] [CrossRef] [PubMed]
- Herold, J.; Andino, R. Poliovirus RNA replication requires genome circularization through a protein-protein bridge. Mol. Cell 2001, 7, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Ooms, M.; Abbink, T.E.; Pham, C.; Berkhout, B. Circularization of the HIV-1 RNA genome. Nucleic Acids Res. 2007, 35, 5253–5261. [Google Scholar] [CrossRef] [PubMed]
- Villordo, S.M.; Gamarnik, A.V. Genome cyclization as strategy for flavivirus RNA replication. Virus Res. 2009, 139, 230–239. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yeh, P.-Y.; Wu, H.-Y. Identification of Cis-Acting Elements on Positive-Strand Subgenomic mRNA Required for the Synthesis of Negative-Strand Counterpart in Bovine Coronavirus. Viruses 2014, 6, 2938-2959. https://doi.org/10.3390/v6082938
Yeh P-Y, Wu H-Y. Identification of Cis-Acting Elements on Positive-Strand Subgenomic mRNA Required for the Synthesis of Negative-Strand Counterpart in Bovine Coronavirus. Viruses. 2014; 6(8):2938-2959. https://doi.org/10.3390/v6082938
Chicago/Turabian StyleYeh, Po-Yuan, and Hung-Yi Wu. 2014. "Identification of Cis-Acting Elements on Positive-Strand Subgenomic mRNA Required for the Synthesis of Negative-Strand Counterpart in Bovine Coronavirus" Viruses 6, no. 8: 2938-2959. https://doi.org/10.3390/v6082938