
Attacked from All Sides: RNA Decay in Antiviral Defense


Department of Microbiology, University of Pennsylvania Perelman School of Medicine, Philadelphia, PA 19104, USA
*
Author to whom correspondence should be addressed.
Viruses 2017, 9(1), 2; https://doi.org/10.3390/v9010002
Submission received: 9 November 2016
/
Revised: 22 December 2016
/
Accepted: 26 December 2016
/
Published: 4 January 2017
(This article belongs to the Special Issue Viral Interactions with Host RNA Decay Pathways)
{kind=link}
Abstract
:The innate immune system has evolved a number of sensors that recognize viral RNA (vRNA) to restrict infection, yet the full spectrum of host-encoded RNA binding proteins that target these foreign RNAs is still unknown. The RNA decay machinery, which uses exonucleases to degrade aberrant RNAs largely from the 5′ or 3′ end, is increasingly recognized as playing an important role in antiviral defense. The 5′ degradation pathway can directly target viral messenger RNA (mRNA) for degradation, as well as indirectly attenuate replication by limiting specific pools of endogenous RNAs. The 3′ degradation machinery (RNA exosome) is emerging as a downstream effector of a diverse array of vRNA sensors. This review discusses our current understanding of the roles of the RNA decay machinery in controlling viral infection.
Keywords:
RNA decay; RNA-protein interactions; decapping; Xrn1; exosome; TRAMP; exonuclease; RNAse; intrinsic immunity; antiviral1. Viral RNAs (vRNAs) as Foreign RNAs
RNA viruses produce RNAs which differ substantially from normal cellular RNAs, leading to their recognition by host-encoded RNA-binding proteins. Unlike cellular RNAs, the genomes of RNA viruses are replicated by RNA-dependent RNA polymerases (RdRp) through antigenome intermediates, creating both transient double-stranded RNA (dsRNA) structures and 5′ triphosphate ends not normally present in cellular messenger RNAs (mRNAs) [1]. For 5′ end protection and recruitment of the translational machinery, endogenous mRNAs are capped in the nucleus. However, cytoplasmic RNA viruses have no access to the normal cellular capping machinery and, thus, many of these viruses go to great lengths to acquire a cap, including enzymatically synthesizing a cap or cap mimic, or acquiring one from cellular mRNA through a process known as cap-snatching [2]. For translation and 3′ end protection, most mRNAs are polyadenylated; some cytoplasmic viral mRNAs achieve this through RdRp-mediated polyadenylation [3]. Furthermore, some viruses encode 3′ structures which impede exonucleases [4,5]. These are but a few examples of the complexity of viral RNA (vRNA) metabolism, which can render viruses susceptible to both immune sensors and the cellular RNA decay machinery.
2. Innate Immune Recognition of vRNA
2.1. RIG-I Like Receptors (RLRs) and DEAD-Box Helicases
The RNAs produced during viral replication serve as an important sign of infection, and a series of sensors have evolved to detect these RNAs. In mammals, many cytosolic vRNAs are recognized by the RIG-I-like receptors (RLRs), RIG-I and MDA-5, which are homologous DEAD-box RNA helicases [6]. Each recognizes different RNA structures. RIG-I recognizes short dsRNAs and RNAs with 5′ triphosphates, and plays a role in restricting viruses, including paramyxoviruses, orthomyxoviruses, and flaviviruses [7,8]. In contrast, MDA-5 recognizes longer dsRNAs and higher-order RNA structures, and is integral for recognition of picornaviruses [9,10]. Both can respond to the synthetic dsRNA polyinosinic-polycytidylic acid (poly(I:C)) dependent on length; long poly(I:C) is a ligand of MDA-5, while short poly(I:C) can activate RIG-I [11]. The RLRs primarily act by signaling the interferon (IFN) system through their adaptor, MAVS; IFN then signals an extensive transcriptional antiviral program [12,13,14,15]. A third RLR helicase, LGP2, lacks the ability to signal through MAVS, and in different contexts has been found to either inhibit or potentiate the signaling of the other two RLRs [16,17,18,19,20,21,22]. In response, many RNA viruses have evolved mechanisms to evade the IFN system and, thus, avoid the consequences of RLR detection; for example, both Rift Valley fever virus (RVFV) and Sindbis virus (SINV) encode accessory proteins (NSs and nsP2) which inhibit the transcription of IFNs [23,24,25,26,27].
Just as mammalian cells utilize the RLR DEAD-box RNA helicases to recognize cytosolic vRNA, the closest Drosophila homolog, Dicer-2, recognizes viral dsRNA intermediates generated during infection [28,29,30]. However, Dicer-2 functions as both sensor and effector; in addition to its helicase domain, it has a ribonuclease (RNase) III domain which cleaves dsRNAs into siRNAs which, in turn, are loaded into the Argonaute 2-containing RNA-induced silencing complex, preventing RNA translation and cleaving vRNA [31]. The antiviral RNA silencing pathway in Drosophila is essential for immune defense; flies with mutations in this pathway rapidly succumb to viral infection. To evade this immune defense, natural insect pathogens such as Drosophila C virus encode suppressors of RNAi [29,32]. Dicer-2 also has silencing-independent antiviral functions which closely parallel the signaling functions of the RLRs; Dicer-2 is required to induce the transcription of the antiviral factor Vago, suggesting that it is also a regulator of antiviral transcription during viral infection in insects [33].
New roles are continually emerging for the larger family of DEAD-box helicases in recognizing vRNA. Many of these genes have roles both in normal cellular metabolism as well as in the control of viral infection. For example, DDX17 normally binds stem-loop structures of primary microRNAs (pri-miRNAs) in the nucleus and recruits the Drosha-anchored microprocessor complex to produce pre-miRNAs [34,35,36,37]. However, upon viral infection, DDX17 is repurposed and exported to the cytoplasm, where it binds a stem-loop miRNA-like structure in RVFV RNA in order to restrict viral replication [37]. Additional helicases are involved in innate recognition, such as the DEAD-box helicase DDX60, which interacts with RIG-I, and the complex of DDX1, DDX21, and DHX36, which bind the innate immune adaptor TRIF, in each case facilitating their activity [38,39].
2.2. Toll-Like Receptors (TLRs)
Endosomal RNAs are sensed by toll-like receptors (TLRs) including TLR3 and TLR7, which detect dsRNA and ssRNA, respectively. These sensors signal through the adaptors TRIF and MyD88 to activate antiviral transcription programs [6]. Endosomal TLRs are highly expressed in dedicated immune cells such as dendritic cells, but are missing from many other cell types, which can only effectively sense cytosolic RNA [40,41,42].
2.3. Protein Kinase R (PKR)
Mammalian cells possess additional cytosolic sensors of vRNA including the dsRNA-activated protein kinase R (PKR) [43,44]. Activation of PKR by dsRNAs from viruses or poly(I:C) induces autophosphorylation of PKR and subsequent phosphorylation of eIF2α, shutting down global protein translation thereby preventing viral protein synthesis [45,46,47,48]. Many viruses prevent PKR-mediated translational shutdown by binding dsRNA or PKR to prevent its activation [44,49]. Other viruses, such as RVFV and poliovirus, induce PKR degradation [50,51,52]. In contrast, several other viruses, such as hepatitis C virus (HCV) and SINV, encode RNA structures which bypass the PKR-dependent global translational arrest and continue to be efficiently translated under these stress conditions [27,53,54,55,56]. Moreover, studies have found that RLR-dependent transcriptional activation is dependent on PKR and vice-versa, suggesting crosstalk between these pathways [57,58].
2.4. Ribonucleases (RNases)
Another cytoplasmic sensor of viral dsRNA, 2′-5′ oligoadenylate synthetase (OAS), is an IFN-inducible enzyme and, thus, is up-regulated in response to viral detection by sensors, such as the RLRs and TLRs [59,60]. Upon sensing vRNA, OAS synthesizes 2,5-adenylate, which, in turn, activates the latent cytoplasmic endoribonuclease RNASEL. RNASEL cleaves vRNA and cellular RNA, thereby inhibiting viral replication [61,62,63]. Furthermore, these cleaved RNAs can, in turn, act as substrates for RLR detection, amplifying the antiviral program [64]. In addition, RNASEL promotes apoptosis in response to viral infection, preventing further viral spread [65]. Recent studies have begun to characterize RNAs as more or less susceptible to RNASEL and to postulate further functions for RNASEL-mediated regulation of specific RNAs [66].
Drosha, a nuclear RNase III enzyme, has a canonical role in processing pri-miRNAs to pre-miRNAs before they are exported to the cytoplasm for Dicer processing. Recent studies have shown that Drosha has antiviral activity [67]. Drosha is exported to the cytoplasm in response to diverse RNA viruses, and restricts RNA virus infection by unknown mechanisms, although it is likely that Drosha is recognizing stem-loop structures in vRNA.
New research continues to uncover RNase activity among previously identified antiviral proteins. SAMHD1 was identified as a restriction factor for human immunodeficiency virus (HIV) that is antagonized by the viral protein Vpx [68]. An initial search for the antiviral mechanism revealed that SAMHD1 is a deoxynucleotide triphosphohydrolase that degrades DNA nucleotides, restricting the nucleotide pool available to HIV [69,70]. However, recent work has also identified 3′-5′ DNase and RNase activity for SAMHD1, suggesting additional antiviral functions that may be active against HIV and other viruses [71].
3. The Canonical RNA Decay Machinery and vRNA Targeting
Emerging data suggest that the canonical RNA decay machinery, which is largely dependent on exonucleases, also plays an important role in antiviral immunity. In general, RNA decay proceeds from either the 5′ or 3′ end of an RNA transcript, and has roles in RNA biogenesis, RNA quality control, and normal RNA turnover. Ribosomal RNAs (rRNAs), small nuclear RNAs (snRNAs), and small nucleolar RNAs (snoRNAs) all require RNA processing in the nucleus to reach their mature forms [72]. Quality control also begins in the nucleus, where RNAs which fail to be properly matured, such as hypoadenylated mRNAs and hypomodified tRNAs, are degraded before they can leave the nucleus [73,74]. In the cytoplasm, additional quality-control checkpoints, such as nonsense-mediated decay or no-go decay, detect stalled ribosomes or premature stop codons and degrade these aberrant mRNAs to release and recycle the translational machinery [75]. Recent studies have shown that RNA decay machinery also serves a key role in post-transcriptional regulation of groups of RNAs, called regulons, which are rapidly co-regulated through specific recognition of sequences in their 5′ and 3′ untranslated regions (UTRs) [76]. These include sequences such as terminal oligopyrimidines (TOPs) at the 5′ and AU-rich elements (AREs) at the 3′ ends of RNAs [77,78,79]. As viruses possess many features of aberrant RNAs, they are increasingly recognized as targets of the RNA decay machinery.
4. Antiviral Roles for Nonsense-Mediated Decay (NMD)
Nonsense-mediated decay (NMD) is the process by which mRNAs with stop codons far from the 3′ end of an mRNA, either due to a premature stop codon or a long 3′ UTR, are detected and degraded at either the 5′ or 3′ ends [75,80,81]. Several RNA viruses have been shown to be sensitive to this pathway. In particular, the NMD components Upf1, Smg5, and Smg7 restrict the replication of Semliki Forest virus (SFV) in mammalian cells [82]. The mechanism of this restriction is unclear, and may act through degradation of vRNA or indirect effects; it is independent of viral 3′ UTR length, suggesting that the long 3′ UTR of SFV mRNA is not necessary for NMD sensitivity. The antiviral effect of NMD is ancient; several plant NMD orthologs, including Upf1, were found to restrict potato virus X by recognizing vRNAs with long 3′ UTRs [83]. NMD is also antagonized by viruses; both HCV and human T-cell leukemia virus type 1 produce proteins which inhibit NMD, suggesting evolutionary pressure to evade this antiviral mechanism [84,85].
5. 5′ Decapping and Decay
Endogenous mRNAs targeted for 5′ decay are typically first deadenylated by the CCR4-NOT complex, often assisted by other deadenylating enzymes, before they can be targeted for decay [80,86]. Although deadenylation is the first regulated step towards mRNA degradation, it is sometimes reversible, and can act to “pause” mRNA translation rather than degrading these targets [87]. Removal of the 5′ cap of RNA by decappers (e.g., Dcp2) is irreversible, and permits degradation by the 5′–3′ exonucleases Xrn1 and Xrn2, in the cytoplasm and nucleus, respectively [88,89,90]. This process is largely conserved from yeast to mammals, though mammals have evolved multiple, partially-redundant decapping enzymes with specificity for different targets; Dcp2 is preferentially utilized in NMD and Nudt16 is preferentially involved in degradation of mRNAs containing AREs or 5′ TOPs [77,91].
Deadenylation, decapping, and 5′ degradation activities coalesce in ribonucleoprotein (RNP) structures known as processing bodies (P bodies). These RNPs consist of mRNAs targeted for decay as well as components of the decapping and 5′ degradation machinery (including Dcp2, its activators, and Xrn1) [86,92]. RNAs that accumulate in P bodies are removed from translation, and normally degraded [93]. Although P-bodies are present in normal cells at baseline, their number and size increase in response to a variety of stressors [94,95]. P bodies can interact with and exchange RNAs with other RNP granules, such as stress granules, which are composed of translationally-stalled RNAs and chaperone proteins [95,96]. There is evidence that visible P bodies are a consequence of high concentrations of mRNAs undergoing decay rather than being necessary for decay, as P body structure can be disrupted without preventing RNA degradation [97,98]. Furthermore, up-regulation of 5′ decapping and decay leads to the loss of visible P bodies due to the depletion of RNA targets [77]. These data suggest that P bodies are dynamic structures which form and dissolve in response to RNA target levels.
6. Antiviral Roles for 5′ to 3′ RNA Decay
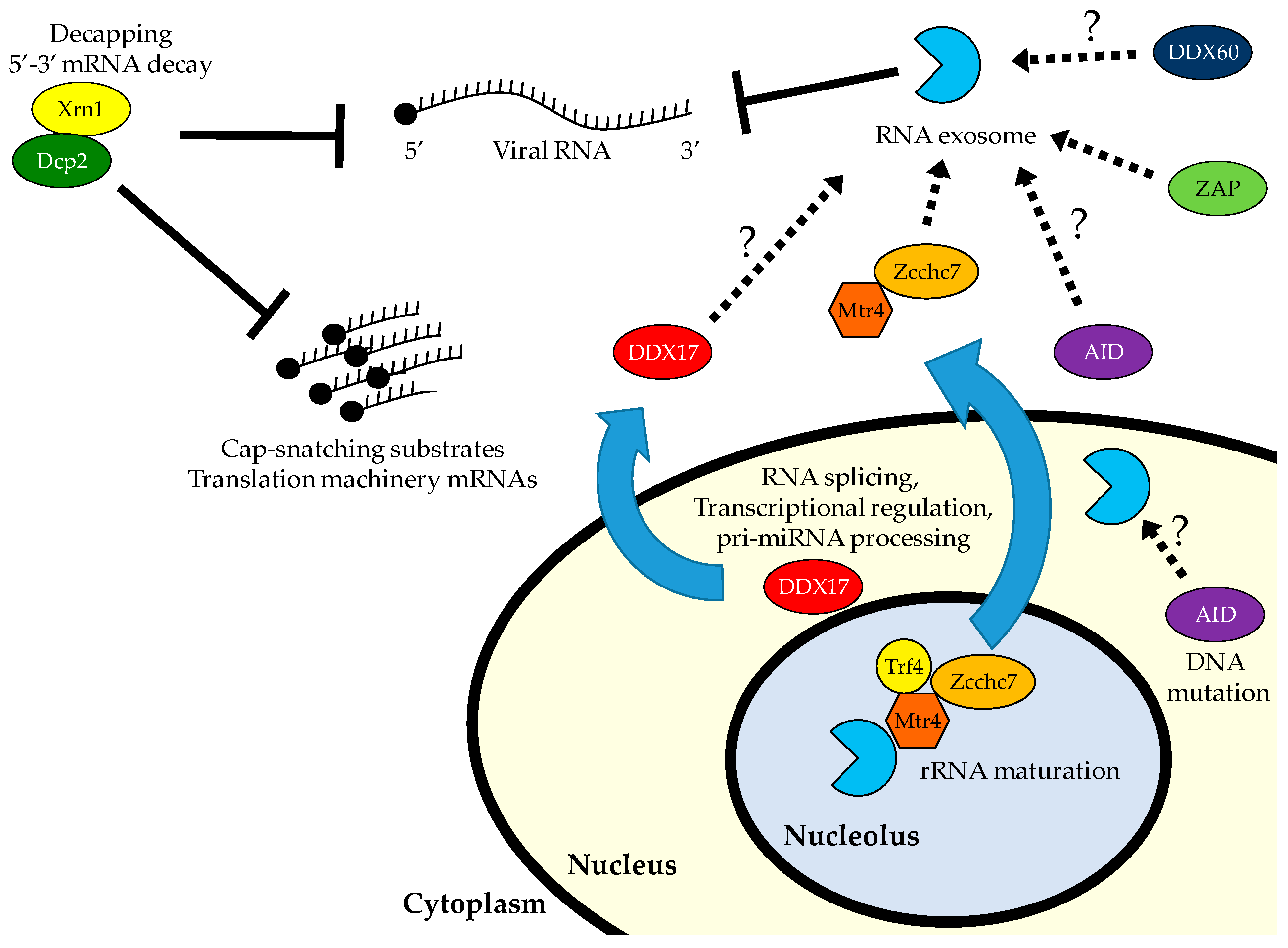
The 5′ to 3′ RNA decay machinery can inhibit viral replication in a number of different ways (Figure 1). Studies have shown that the cytoplasmic 5′ RNA exonuclease, Xrn1, can target flavivirus RNAs and in response these viruses antagonize Xrn1 by encoding structured RNAs that result in Xrn1 stalling [99,100,101]. Furthermore, poliovirus induces the degradation of host 5′ decay factors, such as Xrn1 and Dcp2, through a combination of viral and host proteases, suggesting evolutionary pressure to evade host 5′ RNA decay machinery [102]. In addition to directly targeting vRNAs, the 5′ decay machinery can also impact viral replication indirectly. Recent studies have shown that decappers limit the pool of host mRNAs available for RVFV to cap snatch from, attenuating replication in both insects and mammals [77,103]. Additionally, in mammals, RVFV infection induces NUDT16-mediated decapping and decay of 5′ TOP-containing mRNAs encoding the translational machinery, limiting both global and virus-specific translation [77].
P body structure is also altered during many viral infections. The up-regulation of 5′ decay during RVFV infection prevents the formation of P bodies due to depletion of the RNA targets around which they nucleate [77]. Poliovirus induces the degradation of 5′ decay proteins such as Xrn1 and Dcp1a, preventing P body formation [102]. In addition to the destruction of P bodies during some viral infections, P body components can be repurposed by viruses to facilitate infection. Flaviviruses relocalize P body components to viral replication centers, where they bind viral 3′ UTRs, promoting efficient viral replication [104,105,106].
7. The RNA Exosome
3′ to 5′ degradation is largely mediated by the RNA exosome. This complex consists of a hexameric barrel (six proteins with RNase PH homology) and a cap structure (three proteins with S1 RNA-binding domains) [107,108,109]. These structural components of the exosome form an internal channel wide enough to permit entry of single-stranded but not double-stranded RNA [110]. In contrast to the 5′ decay machinery which localizes to cytoplasmic P bodies, exosome components are both nuclear and cytoplasmic, and can accumulate in poorly-understood RNP granules [92,111,112,113]. While exosome proteins share structural and sequence homology to RNases, the structural components of the exosome are not believed to contribute directly to RNA degradation in vivo. Rather, in yeast, where it has been extensively characterized, 3′ to 5′ exonuclease activity is performed by two exosome-associated RNA exonucleases: Rrp6, which is exclusive to the nucleus, and Dis3, which is present in both the nucleus and the cytoplasm [114,115,116].
The exosome is both structurally and functionally conserved from yeast to humans [117]. Indeed, mutations in the yeast exosomal genes Rrp4, Csl4, or Dis3 can be complemented with the human orthologs [116]. Interestingly, the localization of the exosome exonucleases has diverged over evolutionary history. Rrp6 is present in both the nuclear and cytoplasmic fractions of human cells, and Dis3 has two additional paralogs in humans, Dis3L1 and Dis3L2, which function exclusively in the cytoplasm [118]. Dis3L2, which lacks the exosome-associating PIN domain, operates independently of the larger exosome complex in a separate 3′ to 5′ degradation system which favors terminally uridylated RNAs [119,120,121].
Though the exosome degrades RNAs indiscriminately in vitro, it degrades RNAs in vivo in a regulated fashion by relying on RNA-binding cofactor complexes that bind specific targets and recruit the exosome for degradation [122]. All known exosome cofactor complexes are anchored by helicases which are thought to unwind higher-order RNA structures to permit single-stranded RNA to be inserted into the exosome barrel for decay [123]. Different RNAs are targeted by the exosome in the nucleus, nucleolus, and cytoplasm; therefore, the exosome relies on different cofactors in each subcellular compartment to target these diverse RNAs for decay. Two major complexes, the cytoplasmic Ski and nuclear TRAMP complexes, have been extensively characterized in yeast.
The Superkiller (Ski) complex is the major cytoplasmic exosome cofactor complex in yeast, named for the “superkilling” phenotype of dsRNA viruses, which are lethal to yeast deficient in these genes [124]. The Ski genes were identified before the discovery of the exosome, and some components of the exosome barrel were also assigned Ski names. Though mutants in cofactor Ski genes lead to increased vRNA, this has not yet been definitively linked to exosomal RNA degradation [125]. The Ski complex consists of a DExH/D-box helicase, Ski2, a tetratricopeptide repeat-containing protein, Ski3, and a WD repeat-containing protein, Ski8 [126]. An adaptor G-protein, Ski7, physically links the Ski complex and the exosome and is required for Ski complex-mediated decay [127]. The Ski complex is involved in recruiting the exosome to RNAs targeted for NMD, as well as nonstop decay [128,129,130]. Orthologs for all three Ski genes are present in higher organisms, though their specific targets have not been clearly defined [131,132]. Interestingly, a recent paper found that the human Ski complex-associated helicase SKI2L prevents hyper-activation of RIG-I in uninfected cells, protecting cells from autoimmune activation and patients harboring mutations in this gene presented with anomalously high IFN signatures [133]. Though the exosome was not shown to be required for this activity, it does suggest that the Ski-associated helicase, potentially with the exosome, may serve to protect the intracellular milieu from overactive RIG-I signaling, paralleling the role of the DNA exonuclease TREX, which degrades cytoplasmic DNA to prevent hyper-activation of the DNA sensor cGAS [134,135,136].
The yeast TRAMP (Trf4/5-Air1/2-Mtr4-Polyadenylation) complex, located in the nucleus, has known roles in degrading hypomodified tRNAs, hypoadenylated mRNAs, cryptic unstable transcripts (CUTs), and in the biogenesis of rRNA, snRNA, and snoRNA [72,73,74,122,137,138]. The complex is anchored by a DExD/H box helicase, Mtr4, which binds the other TRAMP components through its arch domain [123,139]. The Zn-knuckle RNA-binding proteins Air1 and Air2 bind specific RNAs and target them for degradation [140]. These two proteins are partially functionally redundant; mutants in each protein accumulate overlapping but non-identical populations of snRNAs, snoRNAs, and mRNAs, and double-mutant strains fail to grow. Trf4 and Trf5 are non-canonical poly(A) polymerases which add 5–6 adenines to RNAs bound to the TRAMP complex [137]. The addition of short poly(A) tails creates an unstructured 3′ end which is thought to facilitate insertion of the RNA into the exosome barrel [141]. This adenylation parallels the role of polyadenylation in Escherichia coli, which, unlike eukaryotic polyadenylation, targets RNAs for decay [142,143]. In addition to the canonical TRAMP complex, Mtr4 can form other modular cofactor complexes by associating with the adaptors Nop53 or Utp18, which assist in some rRNA maturation steps [144].
The TRAMP complex is conserved in humans, but nuclear RNA degradation machinery has additional complexity. As in yeast, human TRAMP is composed of a helicase, hMTR4, a zinc-finger Air-like protein, hZCCHC7, and a poly(A) polymerase, hTRF4-1 or hTRF4-2 [145,146]. However, unlike yeast TRAMP, the human TRAMP complex is restricted to the nucleolus, and is only known to process rRNA [145,147]. Most of the yeast TRAMP targets, such as mRNAs, snRNAs, snoRNAs, and promoter upstream transcripts (PROMPTs, which are analogous to yeast CUTs) appear to be regulated in human cells by the nuclear exosome targeting (NEXT) complex, which shares hMTR4 with the TRAMP complex, but also contains the zinc-finger protein hZCCHC8 and the RNA-binding motif protein hRBM7 [145,148,149]. Other targets are likely to exist for mammalian TRAMP-like complexes; murine cells depleted of Mtr4 accumulate adenylated 5′ miRNA fragments, suggesting that adenylation and Mtr4-mediated degradation may be important for these RNAs [150]. The full spectrum of Mtr4-anchored complexes in mammals and the regulation of other classical yeast TRAMP targets (such as misprocessed tRNAs) remain unclear.
8. Antiviral Roles for the RNA Exosome and 3′ Decay
Studies have implicated the exosome in antiviral defense. A number of antiviral RNA-binding proteins co-immunoprecipitate with the exosome, suggesting that their mechanism of action may involve exosomal degradation. DDX17 restricts RVFV by binding a miRNA-like stem loop structure encoded in the vRNA [37]. Though its mechanism of restriction is unknown, DDX17 binds to the exosome, suggesting that it directly recruits the exosome to degrade these bound vRNAs [145,151]. DDX60, which is antiviral against vesicular stomatitis virus (VSV), also binds the exosome [39]. However, DDX60 does not depend on the exosome for its antiviral function, but rather bridges vRNA and RLRs to potentiate signaling. The cytidine deaminase AID, which binds the exosome and hepatitis B virus (HBV) RNA in a complex, is antiviral when overexpressed only if the exosome is present, suggesting the possibility that it recruits the exosome to degrade HBV RNA [152]. The zinc-finger antiviral protein (ZAP) binds SINV and retrovirus RNA, as well as components of the exosome [153]. In overexpression systems, ZAP restricts MLV viral replication in an exosome-dependent fashion, as well as affecting the expression and stability of viral luciferase reporters for both MLV and HIV [154,155]. It remains unclear if the exosome is required for the activity of endogenous ZAP or degrades ZAP-bound vRNAs. The cell biology of these factors is largely unexplored, but overexpressed DDX60 and ZAP localize to the cytoplasm, while overexpressed AID binds the exosome in both the nucleus and cytoplasm [39,152,156]. In response to viral infection, DDX17 translocates from the nucleus to the cytoplasm where the vRNAs are located [37].
Recent work has implicated the exosome and components of a canonical cofactor complex in the direct recognition and degradation of specific vRNAs. RNAi screening revealed an antiviral role for exosome core components as well as the TRAMP components Mtr4 and Zcchc7 against three RNA viruses from distinct families, VSV, SINV, and RVFV, in both Drosophila and human cells [157]. Though TRAMP components are normally nucleolar, infection with these cytoplasmic viruses leads to the export of hMTR4 and hZCCHC7 to the cytoplasm, where they complex with the exosome and specifically bind viral mRNAs. Further study found that RVFV mRNA is destabilized by the exosome and hZCCHC7, and that the 3′ UTR of RVFV mRNA is sufficient to render a reporter RNA susceptible to exosomal degradation during viral infection. Cell biological studies showed that hZCCHC7 localizes to cytoplasmic punctae during viral infection, suggesting that it may be recruited to RNP granules for its antiviral function.
Taken together, these studies suggest that the exosome is a broad antiviral effector downstream of diverse sensors which bind distinct vRNAs to recruit the exosome for degradation (Figure 1). Though the exosome has been shown to degrade vRNA sensed by some of the proposed exosomal cofactors, such as Mtr4. Zcchc7, and ZAP, much work remains to describe the mechanism and exosomal involvement in antiviral restriction downstream of the other proteins.
9. Concluding Remarks
Increasing evidence suggests that the RNA decay machinery plays important roles in antiviral defense. This can involve either direct effects on vRNA stability or indirect regulation of the intracellular milieu. Furthermore, an emerging theme suggests that many RNA binding proteins can be repurposed from their endogenous roles in the nucleus to antiviral roles in the cytoplasm. Future studies are necessary to further elucidate how these RNA binding proteins recognize foreign RNAs and how they interface with the RNA decay machinery to restrict vRNA replication.
Acknowledgments
We thank E. Klinman, B. Hackett, and C. Sansone for critical reading of the manuscript. This work was supported by National Institute of Health grants R01AI074951, U54AI057168, and R01AI095500 to SC, and T32AI007324 to JMM. SC is a recipient of the Burroughs Wellcome Investigators in the Pathogenesis of Infectious Disease Award.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moon, S.L.; Wilusz, J. Cytoplasmic viruses: Rage against the (cellular RNA decay) machine. PLoS Pathog. 2013, 9, e1003762. [Google Scholar] [CrossRef] [PubMed]
- Decroly, E.; Ferron, F.; Lescar, J.; Canard, B. Conventional and unconventional mechanisms for capping viral mRNA. Nat. Rev. Microbiol. 2012, 10, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.L.; Barnhart, M.D.; Wilusz, J. Inhibition and avoidance of mRNA degradation by RNA viruses. Curr. Opin. Microbiol. 2012, 15, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Ford, L.P.; Wilusz, J. 3′-Terminal RNA structures and poly(U) tracts inhibit initiation by a 3′-5′ exonuclease in vitro. Nucleic Acids Res. 1999, 27, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Iwakawa, H.-O.; Tajima, Y.; Taniguchi, T.; Kaido, M.; Mise, K.; Tomari, Y.; Taniguchi, H.; Okuno, T. Poly(A)-binding protein facilitates translation of an uncapped/nonpolyadenylated viral RNA by binding to the 3′ untranslated region. J. Virol. 2012, 86, 7836–7849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbalat, R.; Ewald, S.E.; Mouchess, M.L.; Barton, G.M. Nucleic acid recognition by the innate immune system. Annu. Rev. Immunol. 2011, 29, 185–214. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Hornung, V.; Ellegast, J.; Kim, S.; Brzózka, K.; Jung, A.; Kato, H.; Poeck, H.; Akira, S.; Conzelmann, K.-K.; Schlee, M.; Endres, S.; Hartmann, G. 5′-Triphosphate RNA is the ligand for RIG-I. Science 2006, 314, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Schulz, O.; Tan, C.-P.; Rehwinkel, J.; Kato, H.; Takeuchi, O.; Akira, S.; Way, M.; Schiavo, G.; Reis e Sousa, C. Activation of MDA5 requires higher-order RNA structures generated during virus infection. J. Virol. 2009, 83, 10761–10769. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, L.; Barchet, W.; Gilfillan, S.; Cella, M.; Beutler, B.; Flavell, R.A.; Diamond, M.S.; Colonna, M. Essential role of mda-5 in type I IFN responses to polyriboinosinic:polyribocytidylic acid and encephalomyocarditis picornavirus. Proc. Natl. Acad. Sci. USA 2006, 103, 8459–8464. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takeuchi, O.; Mikamo-Satoh, E.; Hirai, R.; Kawai, T.; Matsushita, K.; Hiiragi, A.; Dermody, T.S.; Fujita, T.; Akira, S. Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5. J. Exp. Med. 2008, 205, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Meylan, E.; Curran, J.; Hofmann, K.; Moradpour, D.; Binder, M.; Bartenschlager, R.; Tschopp, J. Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 2005, 437, 1167–1172. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.B.; Sun, L.; Ea, C.-K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF 3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.-G.; Wang, Y.-Y.; Han, K.-J.; Li, L.-Y.; Zhai, Z.; Shu, H.-B. VISA is an adapter protein required for virus-triggered IFN-beta signaling. Mol. Cell 2005, 19, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Bruns, A.M.; Leser, G.P.; Lamb, R.A.; Horvath, C.M. The innate immune sensor LGP2 activates antiviral signaling by regulating MDA5-RNA interaction and filament assembly. Mol. Cell 2014, 55, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Bruns, A.M.; Pollpeter, D.; Hadizadeh, N.; Myong, S.; Marko, J.F.; Horvath, C.M. ATP hydrolysis enhances RNA recognition and antiviral signal transduction by the innate immune sensor, laboratory of genetics and physiology 2 (LGP2). J. Biol. Chem. 2013, 288, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Komuro, A.; Horvath, C.M. RNA- and virus-independent inhibition of antiviral signaling by RNA helicase LGP2. J. Virol. 2006, 80, 12332–12342. [Google Scholar] [CrossRef] [PubMed]
- Rothenfusser, S.; Goutagny, N.; DiPerna, G.; Gong, M.; Monks, B.G.; Schoenemeyer, A.; Yamamoto, M.; Akira, S.; Fitzgerald, K.A. The RNA helicase Lgp2 inhibits TLR-independent sensing of viral replication by Retinoic Acid-Inducible Gene-I. J. Immunol. 2005, 175, 5260–5268. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S.; Takeuchi, O. LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses. Proc. Natl. Acad. Sci. USA 2010, 107, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, T.; Valdes, M.; Elsby, R.; Kakuta, S.; Caceres, G.; Saijo, S.; Iwakura, Y.; Barber, G.N. Loss of DExD/H Box RNA helicase LGP2 manifests disparate antiviral responses. J. Immunol. 2007, 178, 6444–6455. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Matsumoto, K.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Foy, E.; Loo, Y.-M.; Gale, M.; Akira, S.; et al. Shared and Unique Functions of the DExD/H-Box Helicases RIG-I, MDA5, and LGP2 in Antiviral Innate Immunity. J. Immunol. 2005, 175, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Billecocq, A.; Spiegel, M.; Vialat, P.; Kohl, A.; Weber, F.; Bouloy, M.; Haller, O. NSs protein of Rift Valley fever virus blocks interferon production by inhibiting host gene transcription. J. Virol. 2004, 78, 9798–9806. [Google Scholar] [CrossRef] [PubMed]
- Bouloy, M.; Janzen, C.; Vialat, P.; Khun, H.; Pavlovic, J.; Huerre, M.; Haller, O. Genetic evidence for an interferon-antagonistic function of rift valley fever virus nonstructural protein NSs. J. Virol. 2001, 75, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Frolova, E.I.; Fayzulin, R.Z.; Cook, S.H.; Griffin, D.E.; Rice, C.M.; Frolov, I. Roles of nonstructural protein nsP2 and α/β interferons in determining the outcome of sindbis virus infection. J. Virol. 2002, 76, 11254–11264. [Google Scholar] [CrossRef] [PubMed]
- Garmashova, N.; Gorchakov, R.; Frolova, E.; Frolov, I. Sindbis virus nonstructural protein nsP2 is cytotoxic and inhibits cellular transcription. J. Virol. 2006, 80, 5686–5696. [Google Scholar] [CrossRef] [PubMed]
- Gorchakov, R.; Frolova, E.; Williams, B.R.G.; Rice, C.M.; Frolov, I. PKR-dependent and -independent mechanisms are involved in translational shutoff during Sindbis virus infection. J. Virol. 2004, 78, 8455–8467. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. RIG-I-like antiviral protein in flies. Nat. Immunol. 2008, 9, 1327–1328. [Google Scholar] [CrossRef] [PubMed]
- Galiana-Arnoux, D.; Dostert, C.; Schneemann, A.; Hoffmann, J.A.; Imler, J.-L. Essential function in vivo for Dicer-2 in host defense against RNA viruses in drosophila. Nat. Immunol. 2006, 7, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Nakahara, K.; Pham, J.W.; Kim, K.; He, Z.; Sontheimer, E.J.; Carthew, R.W.; Abrahante, J.E.; Daul, A.L.; Li, M.; et al. Distinct Roles for Drosophila Dicer-1 and Dicer-2 in the siRNA/miRNA Silencing Pathways. Cell 2004, 117, 69–81. [Google Scholar] [CrossRef]
- Wang, X.-H.; Aliyari, R.; Li, W.-X.; Li, H.-W.; Kim, K.; Carthew, R.; Atkinson, P.; Ding, S.-W. RNA interference directs innate immunity against viruses in adult Drosophila. Science 2006, 312, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, R.P.; Saleh, M.-C.; Berry, B.; Foo, C.; Houk, A.; Antoniewski, C.; Andino, R. The RNA silencing endonuclease Argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes Dev. 2006, 20, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffmann, J.A.; Imler, J.-L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.I.; Yan, K.-P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Triboulet, R.; Mohseni, M.; Schlegelmilch, K.; Shrestha, K.; Camargo, F.D.; Gregory, R.I. Hippo signaling regulates microprocessor and links cell-density-dependent miRNA biogenesis to cancer. Cell 2014, 156, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.I.; Yamagata, K.; Sugimoto, K.; Iwamoto, T.; Kato, S.; Miyazono, K. Modulation of microRNA processing by p53. Nature 2009, 460, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Moy, R.H.; Cole, B.S.; Yasunaga, A.; Gold, B.; Shankarling, G.; Varble, A.; Molleston, J.M.; tenOever, B.R.; Lynch, K.W.; Cherry, S. Stem-loop recognition by DDX17 facilitates miRNA processing and antiviral defense. Cell 2014, 158, 764–777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kim, T.; Bao, M.; Facchinetti, V.; Jung, S.Y.; Ghaffari, A.A.; Qin, J.; Cheng, G.; Liu, Y.-J. DDX1, DDX21, and DHX36 helicases form a complex with the adaptor molecule TRIF to sense dsRNA in dendritic cells. Immunity 2011, 34, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, M.; Oshiumi, H.; Matsumoto, M.; Seya, T. DDX60, a DEXD/H box helicase, is a novel antiviral factor promoting RIG-I-Like Receptor-mediated signaling. Mol. Cell. Biol. 2011, 31, 3802–3819. [Google Scholar] [CrossRef] [PubMed]
- Laredj, L.N.; Beard, P. Adeno-associated virus activates an innate immune response in normal human cells but not in osteosarcoma cells. J. Virol. 2011, 85, 13133–13143. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Müller, C.; Podszuweit, A.; Montino, C.; Vollmer, J.; Forsbach, A. Toll-like receptor (TLR) 3 immune modulation by unformulated small interfering RNA or DNA and the role of CD14 (in TLR-mediated effects). Immunology 2012, 136, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Clemens, M.J.; Hershey, J.W.; Hovanessian, A.C.; Jacobs, B.C.; Katze, M.G.; Kaufman, R.J.; Lengyel, P.; Samuel, C.E.; Sen, G.C.; Williams, B.R. PKR: Proposed nomenclature for the RNA-dependent protein kinase induced by interferon. J. Interf. Res. 1993, 13, 241. [Google Scholar] [CrossRef] [PubMed]
- Gale, M.; Katze, M.G. Molecular mechanisms of interferon resistance mediated by viral-directed inhibition of PKR, the interferon-induced protein kinase. Pharmacol. Ther. 1998, 78, 29–46. [Google Scholar] [CrossRef]
- Hinnebusch, A.G. The eIF-2α kinases: Regulators of protein synthesis in starvation and stress. Semin. Cell. Biol. 1994, 5, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Meurs, E.; Chong, K.; Galabru, J.; Thomas, N.S.B.; Kerr, I.M.; Williams, B.R.G.; Hovanessian, A.G. Molecular cloning and characterization of the human double-stranded RNA-activated protein kinase induced by interferon. Cell 1990, 62, 379–390. [Google Scholar] [CrossRef]
- Chacko, M.S.; Adamo, M.L. Double-stranded RNA decreases IGF-I gene expression in a protein kinase R-Dependent, but type I interferon-independent, mechanism in C6 Rat Glioma Cells. Endocrinol. Endocr. Soc. 2011. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, S.; Roberts, P.C.; Brown, L.E.; Truong, H.; Pattnaik, A.K.; Archer, D.R.; Barber, G.N. Essential role for the dsRNA-dependent protein kinase PKR in innate immunity to viral infection. Immunity 2000, 13, 129–141. [Google Scholar] [CrossRef]
- Short, J.A.L. Viral evasion of interferon stimulated genes. Biosci. Horiz. 2009, 2, 212–224. [Google Scholar] [CrossRef]
- Black, T.L.; Safer, B.; Hovanessian, A.; Katze, M.G. The cellular 68,000-Mr protein kinase is highly autophosphorylated and activated yet significantly degraded during poliovirus infection: Implications for translational regulation. J. Virol. 1989, 63, 2244–2251. [Google Scholar] [PubMed]
- Ikegami, T.; Narayanan, K.; Won, S.; Kamitani, W.; Peters, C.J.; Makino, S. Rift Valley fever virus NSs protein promotes post-transcriptional downregulation of protein kinase PKR and inhibits eIF2alpha phosphorylation. PLoS Pathog. 2009, 5, e1000287. [Google Scholar] [CrossRef] [PubMed]
- Kainulainen, M.; Lau, S.; Samuel, C.E.; Hornung, V.; Weber, F. NSs virulence factor of rift valley fever virus engages the F-Box proteins FBXW11 and β-TRCP1 to degrade the antiviral protein Kinase PKR. J. Virol. 2016, 90, 6140–6147. [Google Scholar] [CrossRef] [PubMed]
- Garaigorta, U.; Chisari, F.V. Hepatitis C virus blocks interferon effector function by inducing protein kinase R phosphorylation. Cell. Host Microbe 2009, 6, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, S.M.; Park, J.H.; Keum, S.J.; Jang, S.K. eIF2A mediates translation of hepatitis C viral mRNA under stress conditions. EMBO J. 2011, 30, 2454–2464. [Google Scholar] [CrossRef] [PubMed]
- Robert, F.; Kapp, L.D.; Khan, S.N.; Acker, M.G.; Kolitz, S.; Kazemi, S.; Kaufman, R.J.; Merrick, W.C.; Koromilas, A.E.; Lorsch, J.R.; et al. Initiation of protein synthesis by hepatitis C virus is refractory to reduced eIF2.GTP.Met-tRNA(i)(Met) ternary complex availability. Mol. Biol. Cell 2006, 17, 4632–4644. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.J.; Mohr, I. Translation initiation and viral tricks. Trends Biochem.Sci. 2003, 28, 130–136. [Google Scholar] [CrossRef]
- Sen, A.; Pruijssers, A.J.; Dermody, T.S.; García-Sastre, A.; Greenberg, H.B. The early interferon response to rotavirus is regulated by PKR and depends on MAVS/IPS-1, RIG-I, MDA-5, and IRF3. J. Virol. 2011, 85, 3717–3732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Langland, J.O.; Jacobs, B.L.; Samuel, C.E. Protein kinase PKR-dependent activation of mitogen-activated protein kinases occurs through mitochondrial adapter IPS-1 and is antagonized by vaccinia virus E3L. J. Virol. 2009, 83, 5718–5725. [Google Scholar] [CrossRef] [PubMed]
- Hovanessian, A.G.; Laurent, A.G.; Chebath, J.; Galabru, J.; Robert, N.; Svab, J. Identification of 69-kd and 100-kd forms of 2–5A synthetase in interferon-treated human cells by specific monoclonal antibodies. EMBO J. 1987, 6, 1273–1280. [Google Scholar] [PubMed]
- Jensen, S.; Thomsen, A.R. Sensing of RNA viruses: A review of innate immune receptors involved in recognizing RNA virus invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef] [PubMed]
- Floyd-Smith, G.; Slattery, E.; Lengyel, P. Interferon action: RNA cleavage pattern of a (2′-5′)oligoadenylate--dependent endonuclease. Science 1981, 212, 1030–1032. [Google Scholar] [CrossRef] [PubMed]
- Wreschner, D.H.; McCauley, J.W.; Skehel, J.J.; Kerr, I.M. Interferon action—Sequence specificity of the ppp(A2′p)nA-dependent ribonuclease. Nature 1981, 289, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Silverman, R.H. Viral encounters with 2′,5′-oligoadenylate synthetase and RNase L during the interferon antiviral response. J. Virol. 2007, 81, 12720–12729. [Google Scholar] [CrossRef] [PubMed]
- Malathi, K.; Dong, B.; Gale, M.; Silverman, R.H. Small self-RNA generated by RNase L amplifies antiviral innate immunity. Nature 2007, 448, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Castelli, J.C.; Hassel, B.A.; Wood, K.A.; Li, X.L.; Amemiya, K.; Dalakas, M.C.; Torrence, P.F.; Youle, R.J. A study of the interferon antiviral mechanism: Apoptosis activation by the 2–5A system. J. Exp. Med. 1997, 186, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Brennan-Laun, S.E.; Ezelle, H.J.; Li, X.-L.; Hassel, B.A. RNase-L control of cellular mRNAs: Roles in biologic functions and mechanisms of substrate targeting. J. Interf. Cytokine Res. 2014, 34, 275–288. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.S.; Schmid, S.; Aguado, L.C.; Sabin, L.R.; Yasunaga, A.; Shim, J.V.; Sachs, D.; Cherry, S.; tenOever, B.R. Drosha as an interferon-independent antiviral factor. Proc. Natl. Acad. Sci. USA 2014, 111, 7108–7113. [Google Scholar] [CrossRef] [PubMed]
- Laguette, N.; Sobhian, B.; Casartelli, N.; Ringeard, M.; Chable-Bessia, C.; Ségéral, E.; Yatim, A.; Emiliani, S.; Schwartz, O.; Benkirane, M. SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 2011, 474, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, D.C.; Ennis-Adeniran, V.; Hedden, J.J.; Groom, H.C.T.; Rice, G.I.; Christodoulou, E.; Walker, P.A.; Kelly, G.; Haire, L.F.; Yap, M.W.; et al. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature 2011, 480, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Powell, R.D.; Holland, P.J.; Hollis, T.; Perrino, F.W. Aicardi-Goutieres syndrome gene and HIV-1 restriction factor SAMHD1 is a dGTP-regulated deoxynucleotide triphosphohydrolase. J. Biol. Chem. 2011, 286, 43596–43600. [Google Scholar] [CrossRef] [PubMed]
- Beloglazova, N.; Flick, R.; Tchigvintsev, A.; Brown, G.; Popovic, A.; Nocek, B.; Yakunin, A.F. Nuclease activity of the human SAMHD1 protein implicated in the Aicardi-Goutieres syndrome and HIV-1 restriction. J. Biol. Chem. 2013, 288, 8101–8110. [Google Scholar] [CrossRef] [PubMed]
- Allmang, C.; Kufel, J.; Chanfreau, G.; Mitchell, P.; Petfalski, E.; Tollervey, D. Functions of the exosome in rRNA, snoRNA and snRNA synthesis. EMBO J. 1999, 18, 5399–5410. [Google Scholar] [CrossRef] [PubMed]
- Milligan, L.; Torchet, C.; Allmang, C.; Shipman, T.; Tollervey, D. A nuclear surveillance pathway for mRNAs with defective polyadenylation. Mol. Cell. Biol. 2005, 25, 9996–10004. [Google Scholar] [CrossRef] [PubMed]
- Kadaba, S. Nuclear surveillance and degradation of hypomodified initiator tRNAMet in S. cerevisiae. Genes Dev. 2004, 18, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Isken, O.; Maquat, L.E. Quality control of eukaryotic mRNA: Safeguarding cells from abnormal mRNA function. Genes Dev. 2007, 21, 1833–1856. [Google Scholar] [CrossRef] [PubMed]
- Keene, J.D. RNA regulons: Coordination of post-transcriptional events. Nat. Rev. Genet. 2007, 8, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.C.; Tartell, M.A.; Herrmann, C.; Hackett, B.A.; Taschuk, F.; Panda, D.; Menghani, S.V.; Sabin, L.R.; Cherry, S. Virus-induced translational arrest through 4EBP1/2-dependent decay of 5′-TOP mRNAs restricts viral infection. Proc. Natl. Acad. Sci. USA 2015, 112, 201418805. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, D.; Gao, M.; O′Connor, J.P.; Raijmakers, R.; Pruijn, G.; Lutz, C.S.; Wilusz, J. The mammalian exosome mediates the efficient degradation of mRNAs that contain AU-rich elements. EMBO J. 2002, 21, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Gherzi, R.; Ong, S.E.; Chan, E.L.; Raijmakers, R.; Pruijn, G.J.; Stoecklin, G.; Moroni, C.; Mann, M.; Karin, M. AU binding proteins recruit the exosome to degrade ARE-containing mRNAs. Cell 2001, 107, 451–464. [Google Scholar] [CrossRef]
- Lejeune, F.; Li, X.; Maquat, L.E. Nonsense-mediated mRNA decay in mammalian cells involves decapping, deadenylating, and exonucleolytic activities. Mol. Cell 2003, 12, 675–687. [Google Scholar] [CrossRef]
- Baker, K.E.; Parker, R. Nonsense-mediated mRNA decay: Terminating erroneous gene expression. Curr. Opin. Cell Biol. 2004, 16, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Balistreri, G.; Horvath, P.; Schweingruber, C.; Zünd, D.; McInerney, G.; Merits, A.; Mühlemann, O.; Azzalin, C.; Helenius, A. The host nonsense-mediated mRNA decay pathway restricts Mammalian RNA virus replication. Cell. Host Microbe 2014, 16, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Garcia, S.; Voinnet, O. Nonsense-mediated decay serves as a general viral restriction mechanism in plants. Cell. Host Microbe 2014, 16, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Nakano, K.; Ando, T.; Yamagishi, M.; Yokoyama, K.; Ishida, T.; Ohsugi, T.; Tanaka, Y.; Brighty, D.W.; Watanabe, T. Viral interference with host mRNA surveillance, the nonsense-mediated mRNA decay (NMD) pathway, through a new function of HTLV-1 Rex: Implications for retroviral replication. Microbes Infect. 2013, 15, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Ramage, H.R.; Kumar, G.R.; Verschueren, E.; Johnson, J.R.; Von Dollen, J.; Johnson, T.; Newton, B.; Shah, P.; Horner, J.; Krogan, N.J.; et al. A combined proteomics/genomics approach links hepatitis C virus infection with nonsense-mediated mRNA decay. Mol. Cell 2015, 57, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.; Sheth, U. P Bodies and the Control of mRNA Translation and Degradation. Mol. Cell 2007, 25, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Huarte, J.; Stutz, A.; O’Connell, M.L.; Gubler, P.; Belin, D.; Darrow, A.L.; Strickland, S.; Vassalli, J.-D. Transient translational silencing by reversible mRNA deadenylation. Cell 1992, 69, 1021–1030. [Google Scholar] [CrossRef]
- Muhlrad, D.; Decker, C.J.; Parker, R. Deadenylation of the unstable mRNA encoded by the yeast MFA2 gene leads to decapping followed by 5′-->3′ digestion of the transcript. Genes Dev. 1994, 8, 855–866. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Stevens, A. Yeast cells lacking 5′-->3′ exoribonuclease 1 contain mRNA species that are poly(A) deficient and partially lack the 5′ cap structure. Mol. Cell. Biol. 1993, 13, 4826–4835. [Google Scholar] [CrossRef] [PubMed]
- Brannan, K.; Kim, H.; Erickson, B.; Glover-Cutter, K.; Kim, S.; Fong, N.; Kiemele, L.; Hansen, K.; Davis, R.; Lykke-Andersen, J.; et al. mRNA decapping factors and the exonuclease Xrn2 function in widespread premature termination of RNA polymerase II transcription. Mol. Cell 2012, 46, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, M.; Kiledjian, M. Differential utilization of decapping enzymes in mammalian mRNA decay pathways. RNA 2011, 17, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Sheth, U.; Parker, R. Decapping and decay of messenger RNA occur in cytoplasmic processing bodies. Science 2003, 300, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Brengues, M.; Teixeira, D.; Parker, R. Movement of Eukaryotic mRNAs between polysomes and cytoplasmic processing bodies. Science 2005, 310, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Ayache, J.; Bénard, M.; Ernoult-Lange, M.; Minshall, N.; Standart, N.; Kress, M.; Weil, D. P-body assembly requires DDX6 repression complexes rather than decay or Ataxin2/2L complexes. Mol. Biol. Cell 2015, 26, 2579–2595. [Google Scholar] [CrossRef] [PubMed]
- Kedersha, N.; Stoecklin, G.; Ayodele, M.; Yacono, P.; Lykke-Andersen, J.; Fritzler, M.J.; Scheuner, D.; Kaufman, R.J.; Golan, D.E.; Anderson, P. Stress granules and processing bodies are dynamically linked sites of mRNP remodeling. J. Cell Biol. 2005, 169, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Wilczynska, A.; Aigueperse, C.; Kress, M.; Dautry, F.; Weil, D. The translational regulator CPEB1 provides a link between dcp1 bodies and stress granules. J. Cell. Sci. 2005, 118, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Behm-Ansmant, I.; Schweizer, D.; Izaurralde, E. P-body formation is a consequence, not the cause, of RNA-mediated gene silencing. Mol. Cell. Biol. 2007, 27, 3970–3981. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, D.; Sheth, U.; Valencia-Sanchez, M.A.; Brengues, M.; Parker, R. Processing bodies require RNA for assembly and contain nontranslating mRNAs. RNA 2005, 11, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.M.; Domingues, P.; Targett-Adams, P.; McLauchlan, J. Comparison of U2OS and Huh-7 cells for identifying host factors that affect hepatitis C virus RNA replication. J. Gen. Virol. 2010, 91, 2238–2248. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.L.; Anderson, J.R.; Kumagai, Y.; Wilusz, C.J.; Akira, S.; Khromykh, A.A.; Wilusz, J. A noncoding RNA produced by arthropod-borne flaviviruses inhibits the cellular exoribonuclease XRN1 and alters host mRNA stability. RNA 2012, 18, 2029–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, P.A.G.C.; Pereira, C.F.; Dalebout, T.J.; Spaan, W.J.M.; Bredenbeek, P.J. An RNA pseudoknot is required for production of yellow fever virus subgenomic RNA by the host nuclease XRN1. J. Virol. 2010, 84, 11395–11406. [Google Scholar] [CrossRef] [PubMed]
- Dougherty, J.D.; White, J.P.; Lloyd, R.E. Poliovirus-mediated disruption of cytoplasmic processing bodies. J. Virol. 2011, 85, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.C.; McLane, L.M.; Maqbool, T.; Panda, D.; Gordesky-Gold, B.; Cherry, S. A genome-wide RNAi screen reveals that mRNA decapping restricts bunyaviral replication by limiting the pools of Dcp2-accessible targets for cap-snatching. Genes Dev. 2013, 27, 1511–1525. [Google Scholar] [CrossRef] [PubMed]
- Emara, M.M.; Brinton, M.A. Interaction of TIA-1/TIAR with West Nile and dengue virus products in infected cells interferes with stress granule formation and processing body assembly. Proc. Natl. Acad. Sci. USA 2007, 104, 9041–9046. [Google Scholar] [CrossRef] [PubMed]
- Chahar, H.S.; Chen, S.; Manjunath, N. P-body components LSM1, GW182, DDX3, DDX6 and XRN1 are recruited to WNV replication sites and positively regulate viral replication. Virology 2013, 436, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.M.; Bidet, K.; Yinglin, A.; Ler, S.G.; Hogue, K.; Blackstock, W.; Gunaratne, J.; Garcia-Blanco, M.A. Quantitative mass spectrometry of DENV-2 RNA-interacting proteins reveals that the DEAD-box RNA helicase DDX6 binds the DB1 and DB2 3′ UTR structures. RNA Biol. 2011, 8, 1173–1186. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Tollervey, D. Threading the barrel of the RNA exosome. Trends Biochem. Sci. 2013, 38, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Greimann, J.C.; Lima, C.D. Reconstitution, activities, and structure of the eukaryotic RNA exosome. Cell 2006, 127, 1223–1237. [Google Scholar] [CrossRef] [PubMed]
- Raijmakers, R.; Egberts, W.V.; van Venrooij, W.J.; Pruijn, G.J.M. Protein-Protein Interactions between Human Exosome Components Support the Assembly of RNase PH-type Subunits into a Six-membered PNPase-like Ring. J. Mol. Biol. 2002, 323, 653–663. [Google Scholar] [CrossRef]
- Makino, D.L.; Baumgärtner, M.; Conti, E. Crystal structure of an RNA-bound 11-subunit eukaryotic exosome complex. Nature 2013, 495, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-J.; Duffy, A.; Chen, C.-Y. Localization of AU-rich element-containing mRNA in cytoplasmic granules containing exosome subunits. J. Biol. Chem. 2007, 282, 19958–19968. [Google Scholar] [CrossRef] [PubMed]
- Graham, A.C.; Kiss, D.L.; Andrulis, E.D. Differential distribution of exosome subunits at the nuclear lamina and in cytoplasmic foci. Mol. Biol. Cell 2006, 17, 1399–1409. [Google Scholar] [CrossRef] [PubMed]
- Zurla, C.; Lifland, A.W.; Santangelo, P.J. Characterizing mRNA interactions with RNA granules during translation initiation inhibition. PLoS ONE 2011, 6, e19727. [Google Scholar] [CrossRef] [PubMed]
- Dziembowski, A.; Lorentzen, E.; Conti, E.; Séraphin, B. A single subunit, Dis3, is essentially responsible for yeast exosome core activity. Nat. Struct. Mol. Biol. 2007, 14, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.; Petfalski, E.; Shevchenko, A.; Mann, M.; Tollervey, D. The exosome: A conserved eukaryotic RNA processing complex containing multiple 3′-->5′ exoribonucleases. Cell 1997, 91, 457–466. [Google Scholar] [CrossRef]
- Allmang, C.; Petfalski, E.; Podtelejnikov, A.; Mann, M.; Tollervey, D.; Mitchell, P. The yeast exosome and human PM-Scl are related complexes of 3′-->5′ exonucleases. Genes Dev. 1999, 13, 2148–2158. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; Tollervey, D. The many pathways of RNA degradation. Cell 2009, 136, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Tomecki, R.; Kristiansen, M.S.; Lykke-Andersen, S.; Chlebowski, A.; Larsen, K.M.; Szczesny, R.J.; Drazkowska, K.; Pastula, A.; Andersen, J.S.; Stepien, P.P.; et al. The human core exosome interacts with differentially localized processive RNases: hDIS3 and hDIS3L. EMBO J. 2010, 29, 2342–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.-M.; Triboulet, R.; Thornton, J.E.; Gregory, R.I. A role for the Perlman syndrome exonuclease Dis3l2 in the Lin28-let-7 pathway. Nature 2013, 497, 244–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.P.; Liu, X.; Whangbo, J.; McCrossan, G.; Sanborn, K.B.; Basar, E.; Walch, M.; Lieberman, J. Apoptosis triggers specific, rapid, and global mRNA Decay with 3′ uridylated intermediates degraded by DIS3L2. Cell. Rep. 2015, 11, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Ustianenko, D.; Hrossova, D.; Potesil, D.; Chalupnikova, K.; Hrazdilova, K.; Pachernik, J.; Cetkovska, K.; Uldrijan, S.; Zdrahal, Z.; Vanacova, S. Mammalian DIS3L2 exoribonuclease targets the uridylated precursors of let-7 miRNAs. RNA 2013, 19, 1632–1638. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; LaCava, J.; Tollervey, D. RNA-quality control by the exosome. Nat. Rev. Mol. Cell Biol. 2006, 7, 529–539. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, J.; Kressler, D.; Tollervey, D.; Linder, P. Dob1p (Mtr4p) is a putative ATP-dependent RNA helicase required for the 3′ end formation of 5.8S rRNA in Saccharomyces cerevisiae. EMBO J. 1998, 17, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Toh-E, A.; Guerry, P.; Wickner, R.B. Chromosomal superkiller mutants of Saccharomyces cerevisiae. J. Bacteriol. 1978, 136, 1002–1007. [Google Scholar] [PubMed]
- Masison, D.C.; Blanc, A.; Ribas, J.C.; Carroll, K.; Sonenberg, N.; Wickner, R.B. Decoying the cap- mRNA degradation system by a double-stranded RNA virus and poly(A)- mRNA surveillance by a yeast antiviral system. Mol. Cell. Biol. 1995, 15, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.T.; Bai, X.; Johnson, A.W. The yeast antiviral proteins Ski2p, Ski3p, and Ski8p exist as a complex in vivo. RNA 2000, 6, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Takahashi, S.; Kobayashi, T.; Kajiho, H.; Hoshino, S.; Katada, T. Ski7p G protein interacts with the exosome and the Ski complex for 3′-to-5′ mRNA decay in yeast. EMBO J. 2001, 20, 4684–4693. [Google Scholar] [CrossRef] [PubMed]
- Jacobs Anderson, J.S.; Parker, R. The 3′ to 5′ degradation of yeast mRNAs is a general mechanism for mRNA turnover that requires the SKI2 DEVH box protein and 3′ to 5′ exonucleases of the exosome complex. EMBO J. 1998, 17, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Van Hoof, A.; Frischmeyer, P.A.; Dietz, H.C.; Parker, R. Exosome-mediated recognition and degradation of mRNAs lacking a termination codon. Science 2002, 295, 2262–2264. [Google Scholar] [CrossRef] [PubMed]
- Horikawa, W.; Endo, K.; Wada, M.; Ito, K. Mutations in the G-domain of Ski7 cause specific dysfunction in non-stop decay. Sci. Rep. 2016, 6, 29295. [Google Scholar] [CrossRef] [PubMed]
- Orban, T.I.; Izaurralde, E. Decay of mRNAs targeted by RISC requires XRN1, the Ski complex, and the exosome. RNA 2005, 11, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Mandal, S.S.; Pham, A.-D.; Zheng, Y.; Erdjument-Bromage, H.; Batra, S.K.; Tempst, P.; Reinberg, D. The human PAF complex coordinates transcription with events downstream of RNA synthesis. Genes Dev. 2005, 19, 1668–1673. [Google Scholar] [CrossRef] [PubMed]
- Eckard, S.C.; Rice, G.I.; Fabre, A.; Badens, C.; Gray, E.E.; Hartley, J.L.; Crow, Y.J.; Stetson, D.B. The SKIV2L RNA exosome limits activation of the RIG-I-like receptors. Nat. Immunol. 2014, 15, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Chiu, Y.-H.; Chen, Z. J. The cGAS-cGAMP-STING Pathway of Cytosolic DNA Sensing and Signaling. Mol. Cell 2014, 54, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Crow, Y.J.; Rehwinkel, J. Aicardi-Goutieres syndrome and related phenotypes: Linking nucleic acid metabolism with autoimmunity. Hum. Mol. Genet. 2009, 18, R130–R136. [Google Scholar] [CrossRef] [PubMed]
- Stetson, D.B.; Ko, J.S.; Heidmann, T.; Medzhitov, R. Trex1 prevents cell-intrinsic initiation of autoimmunity. Cell 2008, 134, 587–598. [Google Scholar] [CrossRef] [PubMed]
- LaCava, J.; Houseley, J.; Saveanu, C.; Petfalski, E.; Thompson, E.; Jacquier, A.; Tollervey, D. RNA Degradation by the Exosome Is Promoted by a Nuclear Polyadenylation Complex. Cell 2005, 121, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Wyers, F.; Rougemaille, M.; Badis, G.; Rousselle, J.-C.; Dufour, M.-E.; Boulay, J.; Régnault, B.; Devaux, F.; Namane, A.; Séraphin, B. Cryptic Pol II Transcripts Are Degraded by a Nuclear Quality Control Pathway Involving a New Poly(A) Polymerase. Cell 2005, 121, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.N.; Klauer, A.A.; Hintze, B.J.; Robinson, H.; van Hoof, A.; Johnson, S.J. The crystal structure of Mtr4 reveals a novel arch domain required for rRNA processing. EMBO J. 2010, 29, 2205–2216. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Xu, Z.; Mathews, D.H.; Butler, J.S. Air proteins control differential TRAMP substrate specificity for nuclear RNA surveillance. RNA 2012, 18, 1934–1945. [Google Scholar] [CrossRef] [PubMed]
- Paolo, S.S.; Vanacova, S.; Schenk, L.; Scherrer, T.; Blank, D.; Keller, W.; Gerber, A.P. Distinct Roles of Non-Canonical Poly(A) Polymerases in RNA Metabolism. PLoS Genet. 2009, 5, e1000555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Reimers, S.; Pandit, S.; Deutscher, M.P.; Bradford, M.; Carpousis, A.; Vanzo, N.; Raynal, L.; Coburn, G.; Mackie, G.; et al. RNA quality control: Degradation of defective transfer RNA. EMBO J. 2002, 21, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Deutscher, M.P. Degradation of RNA in bacteria: Comparison of mRNA and stable RNA. Nucleic Acids Res. 2006, 34, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Thoms, M.; Thomson, E.; Baßler, J.; Gnädig, M.; Griesel, S.; Hurt, E. The exosome is recruited to RNA substrates through specific adaptor proteins. Cell 2015, 162, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Lubas, M.; Christensen, M.S.; Kristiansen, M.S.; Domanski, M.; Falkenby, L.G.; Lykke-Andersen, S.; Andersen, J.S.; Dziembowski, A.; Jensen, T.H. Interaction profiling identifies the human nuclear exosome targeting complex. Mol. Cell 2011, 43, 624–637. [Google Scholar] [CrossRef] [PubMed]
- Fasken, M.B.; Leung, S.W.; Banerjee, A.; Kodani, M.O.; Chavez, R.; Bowman, E.A.; Purohit, M.K.; Rubinson, M.E.; Rubinson, E.H.; Corbett, A.H. Air1 zinc knuckles 4 and 5 and a conserved IWRXY motif are critical for the function and integrity of the Trf4/5-Air1/2-Mtr4 polyadenylation (TRAMP) RNA quality control complex. J. Biol. Chem. 2011, 286, 37429–37445. [Google Scholar] [CrossRef] [PubMed]
- Shcherbik, N.; Wang, M.; Lapik, Y.R.; Srivastava, L.; Pestov, D.G. Polyadenylation and degradation of incomplete RNA polymerase I transcripts in mammalian cells. EMBO Rep. 2010, 11, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Lubas, M.; Andersen, P.R.; Schein, A.; Dziembowski, A.; Kudla, G.; Jensen, T.H. The human nuclear exosome targeting complex is loaded onto newly synthesized RNA to direct early ribonucleolysis. Cell Rep. 2015, 10, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.R.; Domanski, M.; Kristiansen, M.S.; Storvall, H.; Ntini, E.; Verheggen, C.; Schein, A.; Bunkenborg, J.; Poser, I.; Hallais, M.; et al. The human cap-binding complex is functionally connected to the nuclear RNA exosome. Nat. Struct. Mol. Biol. 2013, 20, 1367–1376. [Google Scholar] [CrossRef] [PubMed]
- Dorweiler, J.E.; Ni, T.; Zhu, J.; Munroe, S.H.; Anderson, J.T. Certain adenylated non-coding RNAs, including 5′ leader sequences of primary microrna transcripts, accumulate in mouse cells following depletion of the RNA Helicase MTR4. PLoS ONE 2014, 9, e99430. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Guo, X.; Lv, F.; Xu, Y.; Gao, G. p72 DEAD box RNA helicase is required for optimal function of the zinc-finger antiviral protein. Proc. Natl. Acad. Sci. USA 2008, 105, 4352–4357. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Liu, G.; Kitamura, K.; Wang, Z.; Chowdhury, S.; Monjurul, A.M.; Wakae, K.; Koura, M.; Shimadu, M.; Kinoshita, K.; et al. TGF-β suppression of HBV RNA through AID-dependent recruitment of an RNA exosome complex. PLOS Pathog. 2015, 11, e1004780. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Carroll, J.-W.N.; Macdonald, M.R.; Goff, S.P.; Gao, G. The zinc finger antiviral protein directly binds to specific viral mRNAs through the CCCH zinc finger motifs. J. Virol. 2004, 78, 12781–12787. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Ma, J.; Sun, J.; Gao, G. The zinc-finger antiviral protein recruits the RNA processing exosome to degrade the target mRNA. Proc. Natl. Acad. Sci. USA 2007, 104, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Chen, G.; Lv, F.; Wang, X.; Ji, X.; Xu, Y.; Sun, J.; Wu, L.; Zheng, Y.-T.; Gao, G. Zinc-finger antiviral protein inhibits HIV-1 infection by selectively targeting multiply spliced viral mRNAs for degradation. Proc. Natl. Acad. Sci. USA 2011, 108, 15834–15839. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, G.; Ji, X.; Gao, G. ZAP is a CRM1-dependent nucleocytoplasmic shuttling protein. Biochem. Biophys. Res. Commun. 2004, 321, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Molleston, J.M.; Sabin, L.R.; Moy, R.H.; Menghani, S.V.; Rausch, K.; Gordesky-Gold, B.; Hopkins, K.C.; Zhou, R.; Jensen, T.H.; Wilusz, J.E.; et al. A conserved virus-induced cytoplasmic TRAMP-like complex recruits the exosome to target viral RNA for degradation. Genes Dev. 2016, 30, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The 5′ to 3′ decay machinery can inhibit viral infection directly through degradation of viral RNA (vRNA; flaviviruses) or indirectly through decapping and degradation of RNAs needed for viral transcription and translation (bunyaviruses). The 3′ to 5′ decay machinery, the RNA exosome, interacts with a variety of RNA-binding proteins, some of which are exported to the cytoplasm in response to viral infection. Recruitment of the exosome can result in degradation of vRNA.
Figure 1.
The 5′ to 3′ decay machinery can inhibit viral infection directly through degradation of viral RNA (vRNA; flaviviruses) or indirectly through decapping and degradation of RNAs needed for viral transcription and translation (bunyaviruses). The 3′ to 5′ decay machinery, the RNA exosome, interacts with a variety of RNA-binding proteins, some of which are exported to the cytoplasm in response to viral infection. Recruitment of the exosome can result in degradation of vRNA.
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Molleston, J.M.; Cherry, S. Attacked from All Sides: RNA Decay in Antiviral Defense. Viruses 2017, 9, 2. https://doi.org/10.3390/v9010002
AMA Style
Molleston JM, Cherry S. Attacked from All Sides: RNA Decay in Antiviral Defense. Viruses. 2017; 9(1):2. https://doi.org/10.3390/v9010002
Chicago/Turabian StyleMolleston, Jerome M., and Sara Cherry. 2017. "Attacked from All Sides: RNA Decay in Antiviral Defense" Viruses 9, no. 1: 2. https://doi.org/10.3390/v9010002
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.