cfa-miR-143 Promotes Apoptosis via the p53 Pathway in Canine Influenza Virus H3N2-Infected Cells


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Infection
2.2.1. Beagle Infections
2.2.2. Cell Infections
2.3. RNA Isolation
2.4. Real-Time Quantitative PCR (qPCR)
2.5. Luciferase Reporter Assay
2.6. Western Blotting
2.7. Flow Cytometry
2.8. Statistical Analysis
3. Results
3.1. Experimental Infection with CIV H3N2 in Dogs
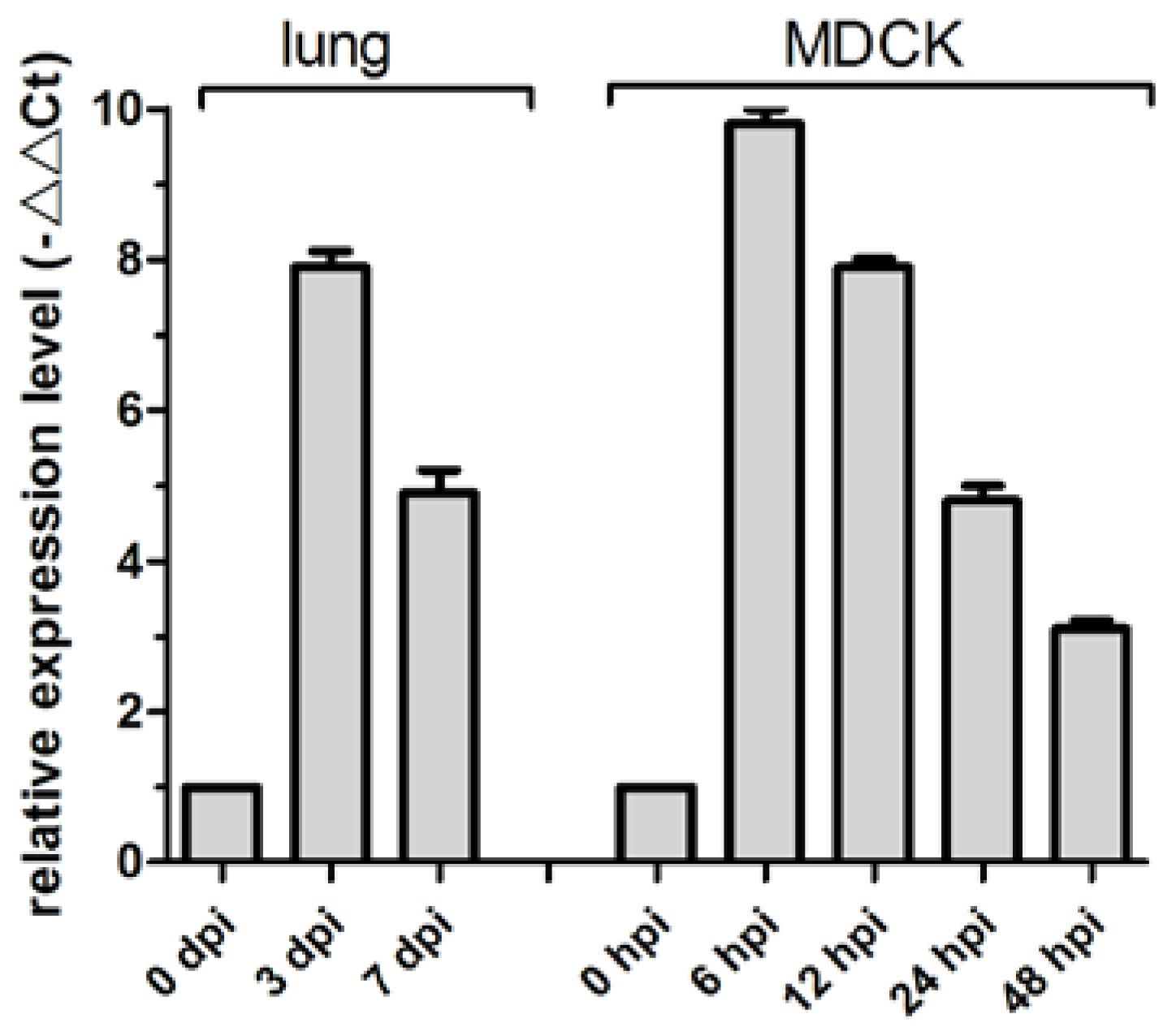
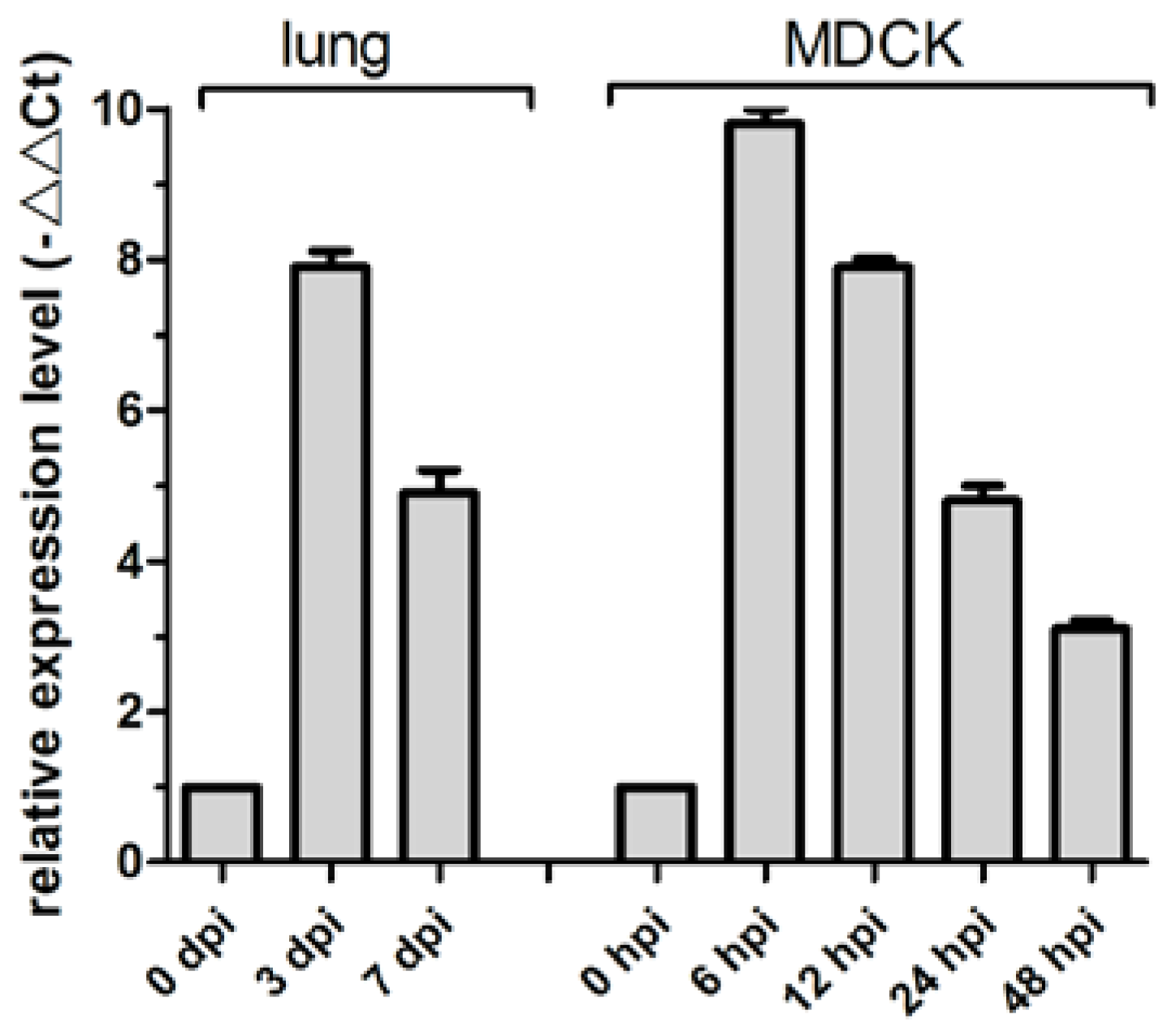
3.2. cfa-miR-143 Upregulation with CIV H3N2 Infection
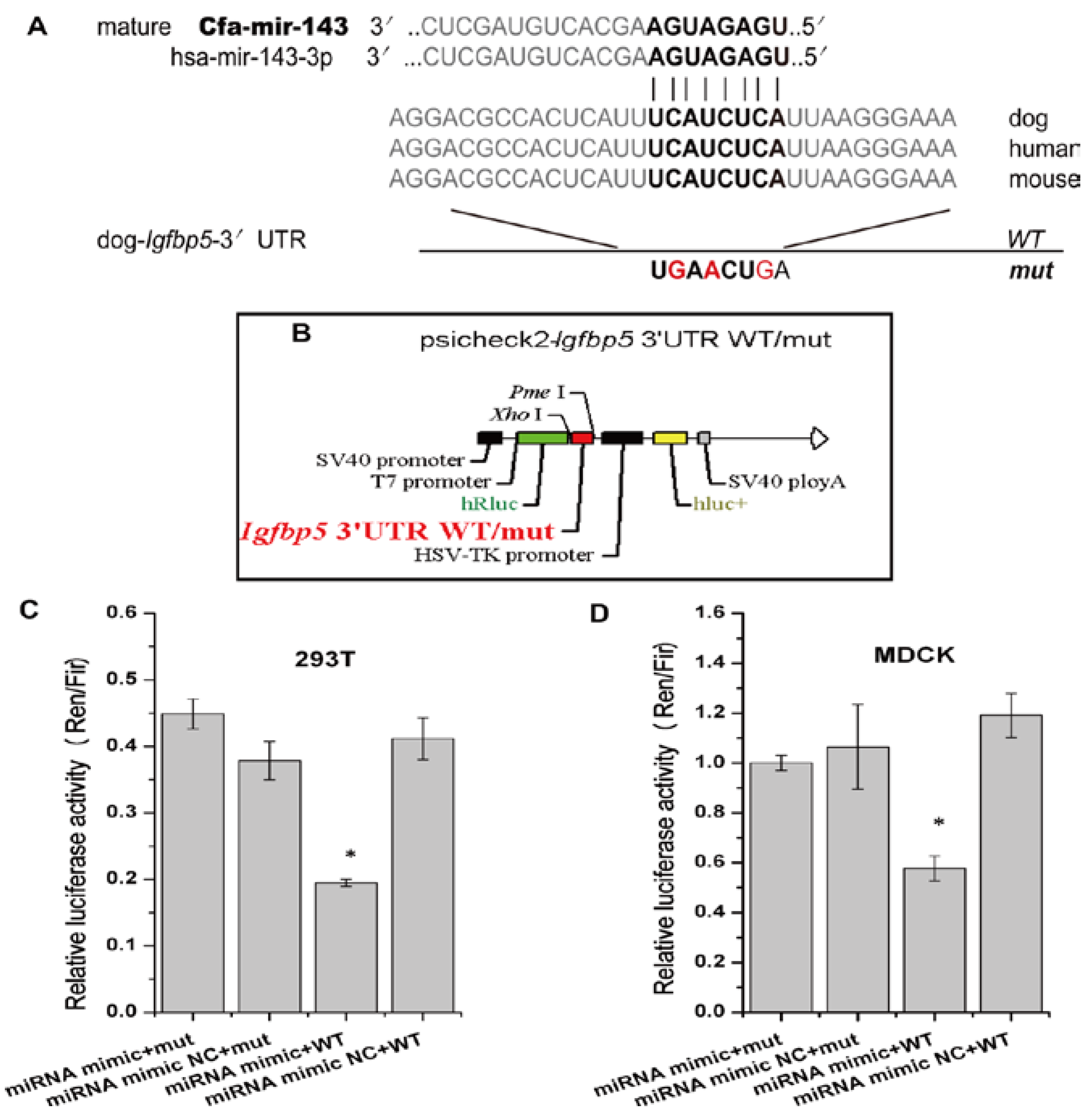
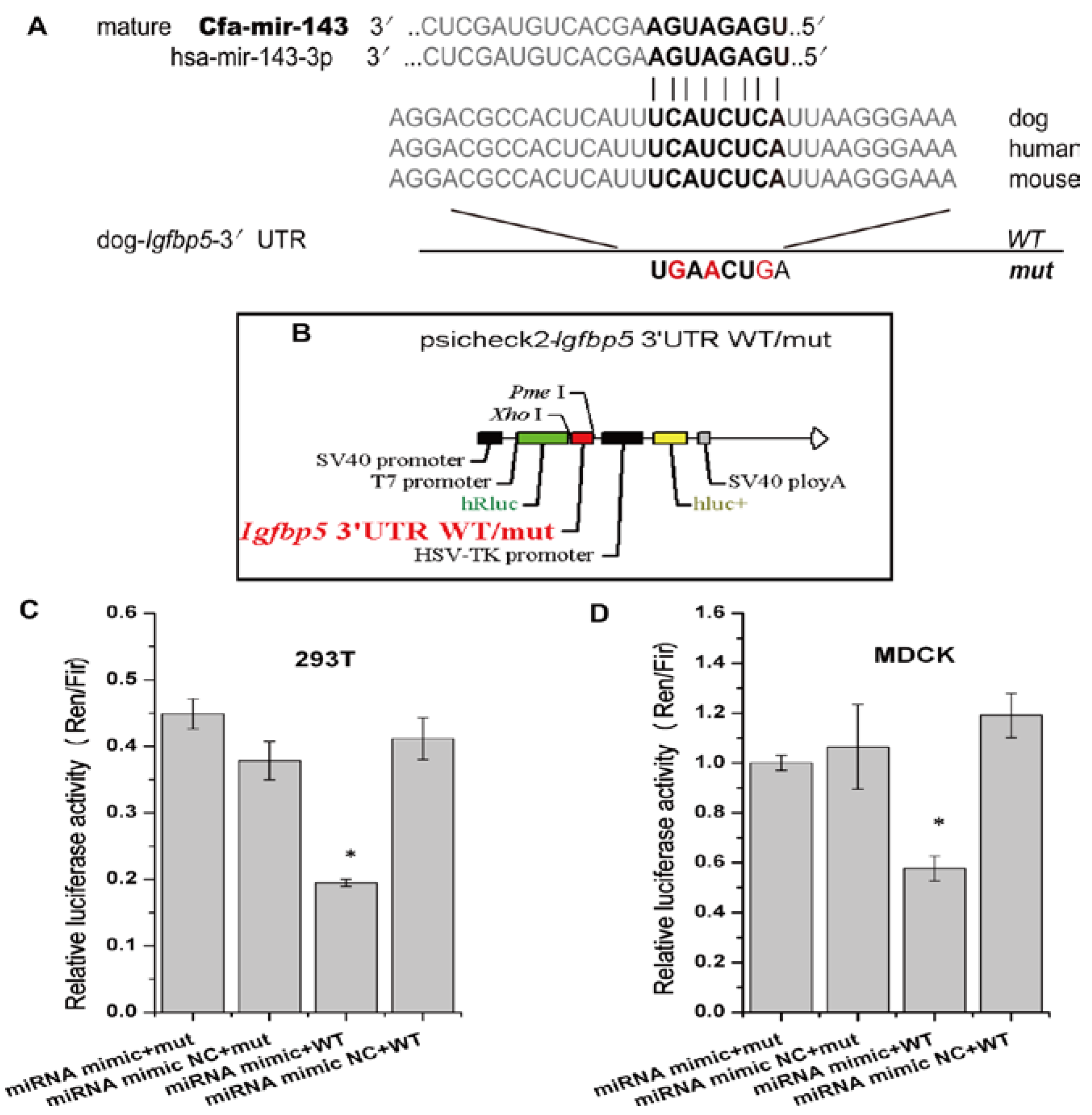
3.3. cfa-miR-143 Targets the Igfbp5 Gene
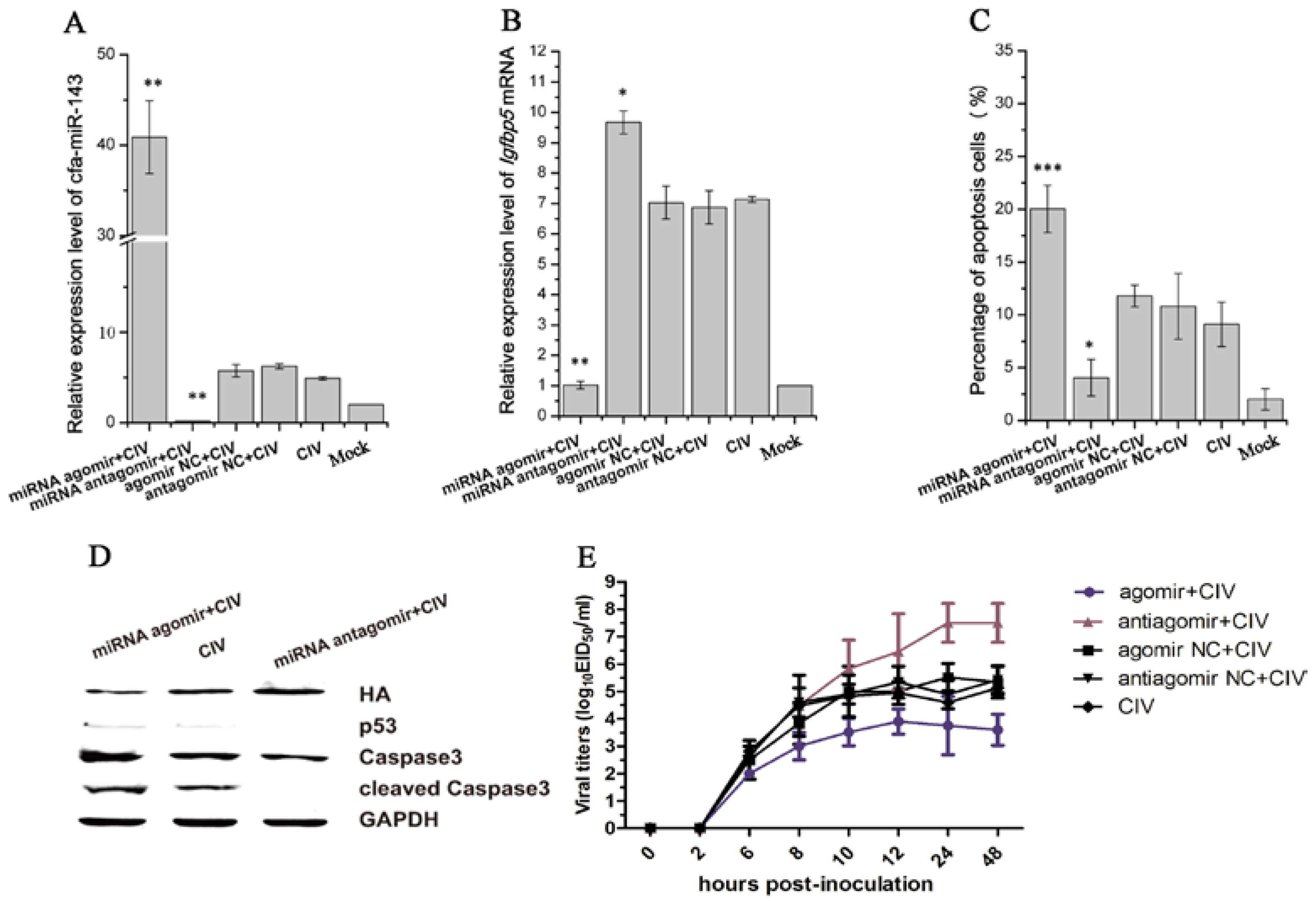
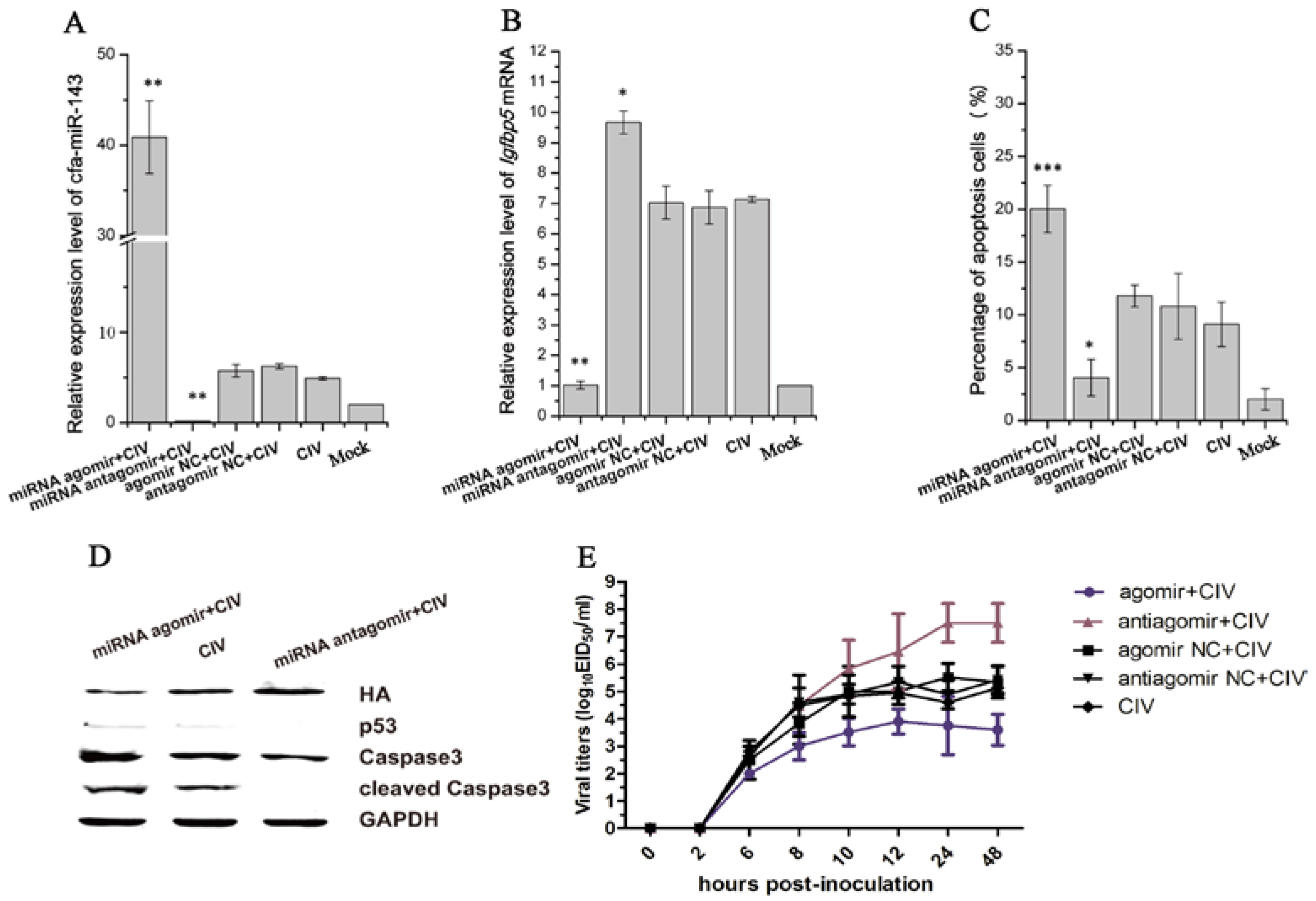
3.4. CIV H3N2 Triggers Apoptosis
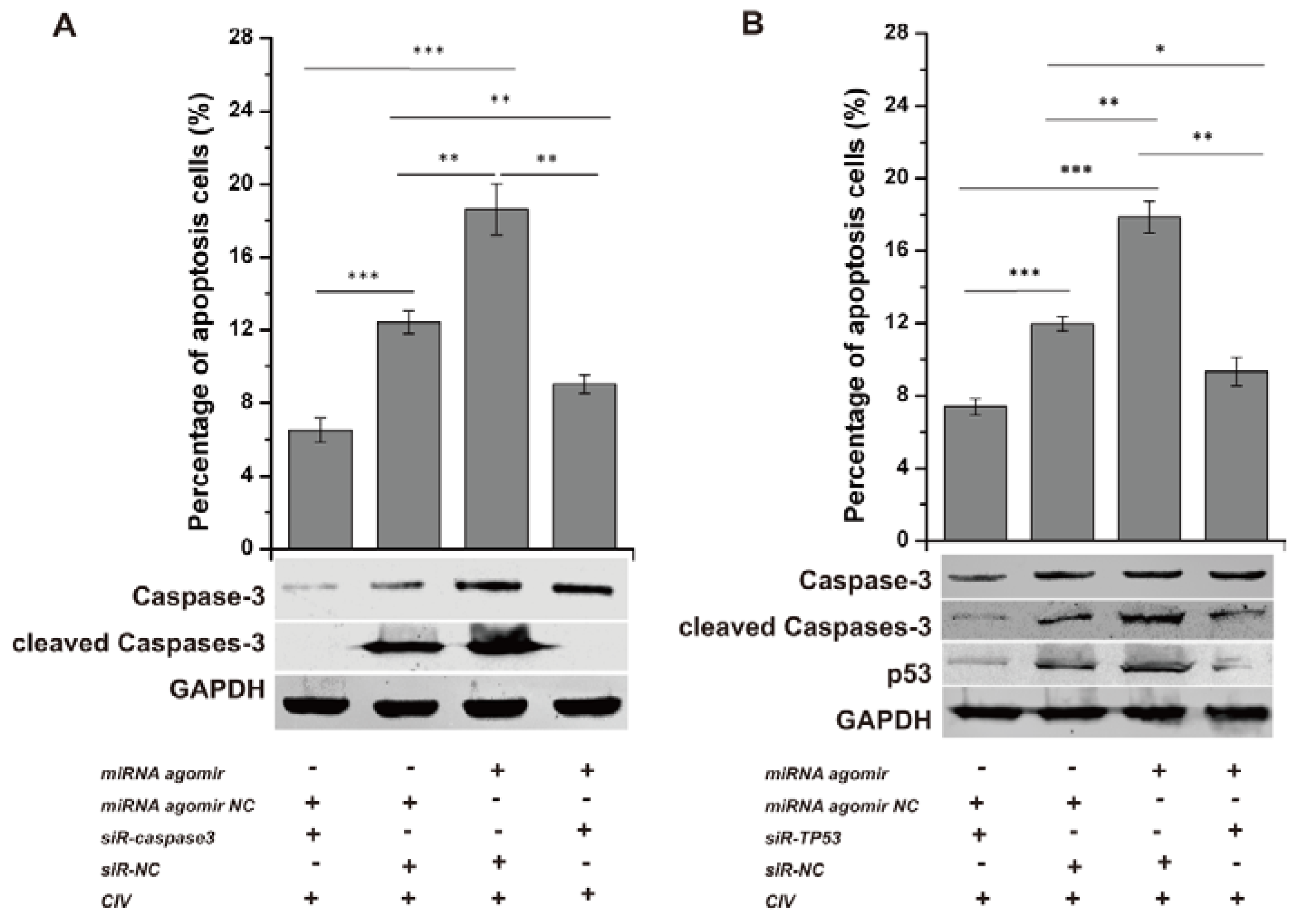
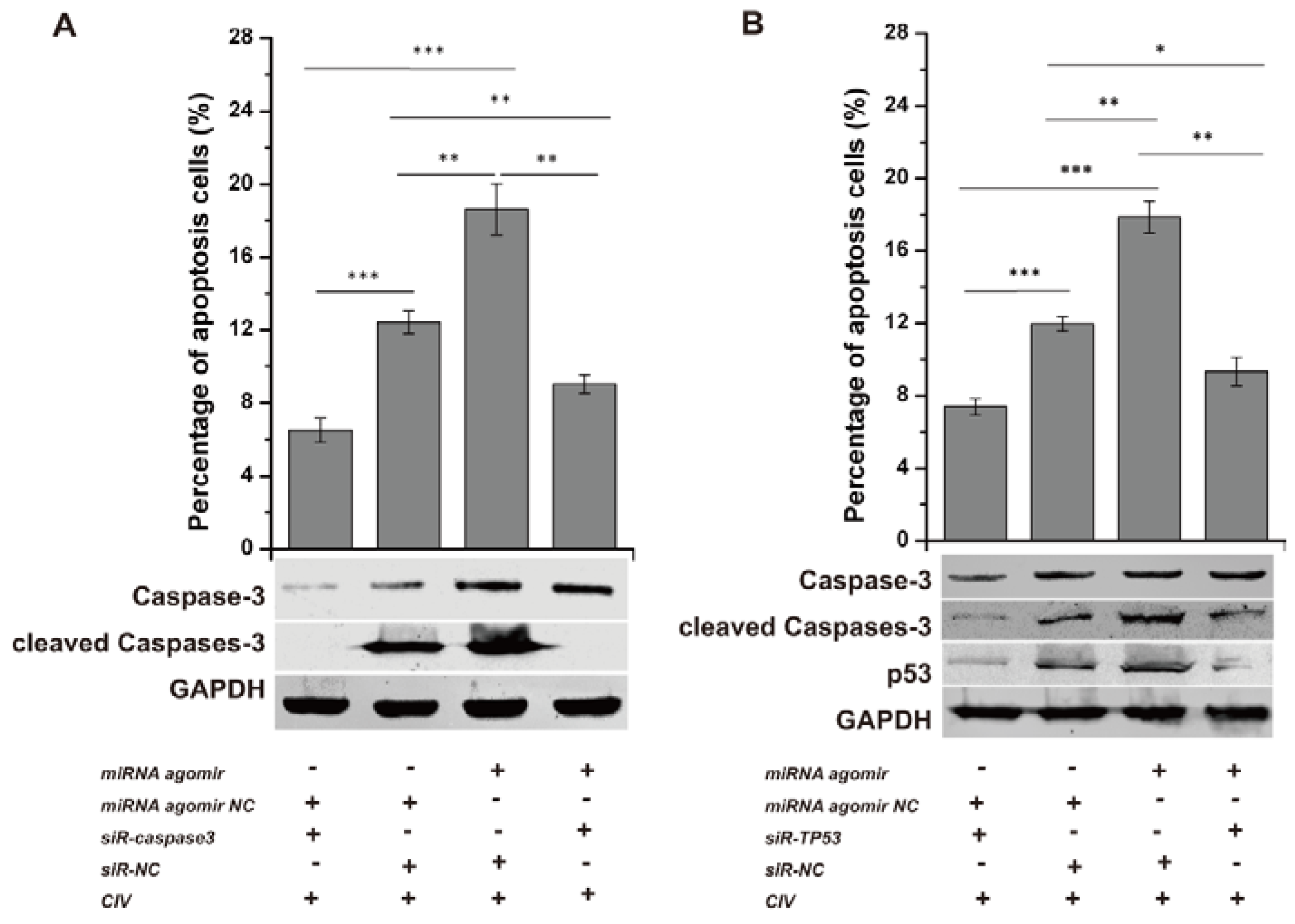
3.5. cfa-miR-143 Induces Apoptosis via the p53 Pathway in CIV-Infected MDCK Cells
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-κB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed]
- Hayashita, Y.; Osada, H.; Tatematsu, Y.; Yamada, H.; Yanagisawa, K.; Tomida, S.; Yatabe, Y.; Kawahara, K.; Sekido, Y.; Takahashi, T. A polycistronic microRNA cluster, miR-17-92, is overexpressed in human lung cancers and enhances cell proliferation. Cancer Res. 2005, 65, 9628–9632. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.Y.; Henson, R.; Wehbe-Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Schee, K.; Lorenz, S.; Worren, M.M.; Gunther, C.; Holden, M.; Hovig, E.; Fodstad, O.; Meza-Zepeda, L.A.; Flatmark, K. Deep sequencing the microrna transcriptome in colorectal cancer. PLoS ONE 2013, 8, e661656. [Google Scholar] [CrossRef] [PubMed]
- Vosa, U.; Vooder, T.; Kolde, R.; Vilo, J.; Metspalu, A.; Annilo, T. Meta-analysis of microRNA expression in lung cancer. Int. J. Cancer 2013, 132, 2884–2893. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Sevignani, C.; Dan Dumitru, C.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.L.; Yi, M.K.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of hepatitis C virus RNA abundance by a liver-specific microRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed]
- Samols, M.A.; Hu, J.H.; Skalsky, R.L.; Renne, R. Cloning and identification of a MicroRNA cluster within the latency-associated region of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2005, 79, 9301–9305. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Feng, G.; Chen, H.; Wang, L.; Wang, Y. Identification of host encoded microRNAs interacting with novel swine-origin influenza A (H1N1) virus and swine influenza virus. Bioinformation 2009, 4, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Aravalli, R.N.; Steer, C.J. Circulating microRNAs: Novel biomarkers for early detection of colorectal cancer. Transl. Res. 2015, 166, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Sakamoto, N.; Oue, N.; Yashiro, M.; Sentani, K.; Yanagihara, K.; Hirakawa, K.; Yasui, W. MicroRNA-143 regulates collagen type III expression in stromal fibroblasts of scirrhous type gastric cancer. Cancer Sci. 2014, 105, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Haasnoot, J.; Berkhout, B. RNA interference: Its use as antiviral therapy. Handb. Exp. Pharmacol. 2006, 173, 117–150. [Google Scholar]
- Pento, J.T. siRNA and microRNA for the treatment of cancer. Drug Future 2007, 32, 1061–1066. [Google Scholar] [CrossRef]
- Horvath, C.M.; Buggele, W.; Johnson, K. Influenza A virus infection of human respiratory cells induces primary MicroRNA expression. Cytokine 2011, 56, 5. [Google Scholar] [CrossRef]
- Kok, K.H.; Jin, D. Influenza A virus NS1 protein does not suppress RNA interference in mammalian cells. J. Gen. Virol. 2006, 87, 2639–2644. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chan, E.Y.; Li, J.; Ni, C.; Peng, X.; Rosenzweig, E.; Tumpey, T.M.; Katze, M.G. MicroRNA Expression and virulence in pandemic influenza virus-infected mice. J. Virol. 2010, 84, 3023–3032. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.T.; Pham, A.M.; Lorini, M.H.; Chua, M.A.; Steel, J.; TenOever, B.R. MicroRNA-mediated species-specific attenuation of influenza A virus. Nat. Biotechnol. 2009, 27, 117–572. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, Q.; Guo, Y.; Liu, S.; Song, R.; Gao, X.; Dai, L.; Li, B.; Zhang, D.; Cheng, J. Microarray analysis of microRNA expression in peripheral blood mononuclear cells of critically ill patients with influenza A (H1N1). BMC Infect. Dis. 2013, 13, 257. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.V.; Price, J.A.; Wendling, M.Q.; Long, J.P.; Bresler, H.S. Preliminary microRNA analysis in lung tissue to identify potential therapeutic targets against H5N1 infection. Viral Immunol. 2012, 25, 3–11. [Google Scholar] [PubMed]
- Vela, E.M.; Kasoji, M.D.; Wendling, M.Q.; Price, J.A.; Knostman, K.A.B.; Bresler, H.S.; Long, J.P. MicroRNA expression in mice infected with seasonal H1N1, swine H1N1 or highly pathogenic H5N1. J. Med. Microbiol. 2014, 63, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Moran, J.; Ramirez-Martinez, G.; Jimenez-Alvarez, L.; Cruz, A.; Perez-Patrigeon, S.; Hidalgo, A.; Orozco, L.; Martinez, A.; Padilla-Noriega, L.; Avila-Moreno, F.; et al. Circulating levels of miR-150 are associated with poorer outcomes of A/H1N1 infection. Exp. Mol. Pathol. 2015, 99, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Meng, D.; Wei, T.; Zhang, S.; Hu, Y.; Wang, M. Apoptosis and pro-inflammatory cytokine response of mast cells induced by influenza A viruses. PLoS ONE 2014, 9, e100109. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.M.; Kim, H.M.; Ku, K.B.; Park, E.H.; Yum, J.; Seo, S.H. H3N2 canine influenza virus causes severe morbidity in dogs with induction of genes related to inflammation and apoptosis. Vet. Res. 2013, 44, 92. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.M.; Byrom, M.W.; Shelton, J.; Ford, L.P. Antisense inhibition of human miRNAs and indications for an involvement of miRNA in cell growth and apoptosis. Nucleic Acids Res. 2005, 33, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhu, J.; Zhou, H.; Zhao, Z.; Zou, Z.; Liu, X.; Lin, X.; Zhang, X.; Deng, X.; Wang, R.; et al. Identification of cellular microRNA-136 as a dual regulator of RIG-I-mediated innate immunity that antagonizes H5N1 IAV replication in A549 cells. Sci. Rep. UK 2015, 5, 14991. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Su, S.; Zhou, D.; Zhou, P.; Xu, T.; Zhang, L.; Cao, N.; Qi, W.; Zhang, G.; Li, S. Comparative analysis of microRNAs from the lungs and trachea of dogs (Canis familiaris) infected with canine influenza virus. Infect. Genet. Evol. 2014, 21, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Tuo, Y.L.; Li, X.M.; Luo, J. Long noncoding RNA UCA1 modulates breast cancer cell growth and apoptosis through decreasing tumor suppressive miR-143. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3403–3411. [Google Scholar] [PubMed]
- Du, F.; Feng, Y.; Fang, J.; Yang, M. MicroRNA-143 enhances chemosensitivity of Quercetin through autophagy inhibition via target GABARAPL1 in gastric cancer cells. Biomed. Pharmacother. 2015, 74, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Qiao, D.; Xu, J.; Le, C.; Huang, E.; Liu, C.; Qiu, P.; Lin, Z.; Xie, W.; Wang, H. Insulin-like growth factor binding protein 5 (IGFBP5) mediates methamphetamine-induced dopaminergic neuron apoptosis. Toxicol. Lett. 2014, 230, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, G.; Salih, D.A.M.; Drozd, A.C.; Cosgrove, R.A.; Cobb, L.J.; Pell, J.M. IGF-independent effects of insulin-like growth factor binding protein-5 (Igfbp5) in vivo. FASEB J. 2009, 23, 2616–2626. [Google Scholar] [CrossRef] [PubMed]
- Butt, A.J.; Dickson, K.A.; McDougall, F.; Baxter, R.C. Insulin-like growth factor-binding protein-5 inhibits the growth of human breast cancer cells in vitro and in vivo. J. Biol. Chem. 2003, 278, 29676–29685. [Google Scholar] [CrossRef] [PubMed]
- Eisfeld, A.J.; Neumann, G.; Kawaoka, Y. Influenza A virus isolation, culture and identification. Nat. Protoc. 2014, 9, 2663–2681. [Google Scholar] [CrossRef] [PubMed]
- Zhirnov, O.P.; Ovcharenko, A.V.; Bukrinskaya, A.G. A modified plaque assay method for accurate analysis of infectivity of influenza viruses with uncleaved hemagglutinin. Arch. Virol. 1982, 71, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Bandres, E.; Cubedo, E.; Agirre, X.; Malumbres, R.; Zarate, R.; Ramirez, N.; Abajo, A.; Navarro, A.; Moreno, I.; Monzo, M.; et al. Identification by real-time PCR of 13 mature microRNAs differentially expressed in colorectal cancer and non-tumoral tissues. Mol. Cancer 2006, 5, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-∆Ct) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Del Puerto, H.L.; Martins, A.S.; Moro, L.; Milsted, A.; Alves, F.; Braz, G.F.; Vasconcelos, A.C. Caspase-3/-8/-9, bax and bcl-2 expression in the cerebellum, lymph nodes and leukocytes of dogs naturally infected with canine distemper virus. Genet. Mol. Res. 2014, 9, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Vitoriano-Souza, J.; das Dores Moreira, N.; Menezes-Souza, D.; Roatt, B.M.; de Oliveira Aguiar-Soares, R.D.; Siqueira-Mathias, F.A.; de Oliveira Cardoso, J.M.; Giunchetti, R.C.; de Sá, R.G.; Corrêa-Oliveira, R.; et al. Dogs immunized with lbsap vaccine displayed high levels of IL-12 and IL-10 cytokines and ccl4, ccl5 and cxcl8 chemokines in the dermis. Mol. Immunol. 2013, 56, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Cong, S.; Zhang, X.; Bao, X.; Wang, W.; Li, H.; Wang, Z.; Wang, G.; Xu, J.; Du, B.; et al. MicroRNA-192 targeting retinoblastoma 1 inhibits cell proliferation and induces cell apoptosis in lung cancer cells. Nucleic Acids Res. 2011, 39, 6669–6678. [Google Scholar] [CrossRef] [PubMed]
- Hale, B.G.; Randall, R.E. PI3K signalling during influenza A virus infections. Biochem. Soc. Trans. 2007, 35, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Zhirnov, O.P.; Klenk, H.D. Control of apoptosis in influenza virus-infected cells by up-regulation of Akt and p53 signaling. Apoptosis 2007, 12, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, G.; Zhang, H.; Shen, Y.; Dai, J.; Wu, L.; Zhou, J.; Jiang, Z.; Li, K. Inability of NS1 protein from an H5N1 influenza virus to activate PI3K/Akt signaling pathway correlates to the enhanced virus replication upon PI3K inhibition. Vet. Res. 2012, 43, 36. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.J.; Yang, J.; Fan, X.L.; Zhao, H.B.; Hu, W.; Li, Z.P.; Yu, G.C.; Ding, X.R.; Wang, J.Z.; Bo, X.C.; et al. Cellular microRNA let-7c inhibits M1 protein expression of the H1N1 influenza A virus in infected human lung epithelial cells. J. Cell. Mol. Med. 2012, 16, 2539–2546. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Ma, C.; Li, W.; Yang, S.; Liu, Z. miR-143 suppresses epithelial-mesenchymal transition and inhibits tumor growth of breast cancer through down-regulation of ERK5. Mol. Carcinog. 2015, 55, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mao, Y.; Zhang, D.; Hao, S.; Zhang, Z.; Li, Z.; Li, B. MiR-143 inhibits tumor cell proliferation and invasion by targeting STAT3 in esophageal squamous cell carcinoma. Cancer Lett. 2016, 373, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xie, Y.Q.; Tang, S.Q.; Wu, X.B.; Zhu, H.Y. miR-143 regulates proliferation and apoptosis of colorectal cancer cells and exhibits altered expression in colorectal cancer tissue. Int. J. Clin. Exp. Med. 2015, 8, 15308–15312. [Google Scholar] [PubMed]
- Skovgaard, K.; Cirera, S.; Vasby, D.; Podolska, A.; Breum, S.O.; Durrwald, R.; Schlegel, M.; Heegaard, P.M. Expression of innate immune genes, proteins and microRNAs in lung tissue of pigs infected experimentally with influenza virus (H1N2). Innate Immun. 2013, 19, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Globinska, A.; Pawelczyk, M.; Kowalski, M.L. MicroRNAs and the immune response to respiratory virus infections. Expert Rev. Clin. Immunol. 2014, 10, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.; Allan, G.J.; Lochrie, J.D.; Flint, D.J. Insulin-like growth factor-binding protein-5 (IGFBP-5): A critical member of the IGF axis. Biochem. J. 2006, 395, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.Y.; Kwok, H.H.; Chan, R.W.Y.; Peiris, M.J.S.; Mak, N.K.; Wong, R.N.S.; Chan, M.C.W.; Yue, P.Y.K. Dual functions of ginsenosides in protecting human endothelial cells against influenza H9N2-induced inflammation and apoptosis. J. Ethnopharmacol. 2011, 137, 1542–1546. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zhang, Y.; Fu, H.; Wu, D.; Zhou, H. Overexpression of microRNA-143 inhibits growth and induces apoptosis in human leukemia cells. Oncol. Rep. 2014, 31, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Seu, Y.B.; Baek, S.H.; Kim, M.J.; Kim, K.J.; Kim, J.H.; Kim, J.R. Induction of cellular senescence by insulin-like growth factor binding protein-5 through a p53-dependent mechanism. Mol. Biol. Cell. 2007, 18, 4543–4552. [Google Scholar] [CrossRef] [PubMed]
- Iwai, A.; Shiozaki, T.; Miyazaki, T. Relevance of signaling molecules for apoptosis induction on influenza A virus replication. Biochem. Biophys. Res. Commun. 2013, 441, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Wlodkowic, D.; Telford, W.; Skommer, J.; Darzynkiewicz, Z. Apoptosis and beyond: Cytometry in studies of programmed cell death. Methods Cell. Biol. 2011, 103, 55–98. [Google Scholar] [PubMed]
- Turpin, E.; Luke, K.; Jones, J.; Tumpey, T.; Konan, K.; Schultz-Cherry, S. Influenza virus infection increases p53 activity: Role of p53 in cell death and viral replication. J. Virol. 2005, 79, 8802–8811. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.B.; Holland, R.E.; Donofrio, J.C.; McCoy, M.H.; Tudor, L.R.; Chambers, T.M. Caspase activation in equine influenza virus induced apoptotic cell death. Vet. Microbiol. 2002, 84, 357–365. [Google Scholar] [CrossRef]
- Manikandan, M.; Deva, M.R.A.; Arunkumar, G.; Rajkumar, K.S.; Rajaraman, R.; Munirajan, A.K. Down Regulation of miR-34a and miR-143 may indirectly inhibit p53 in oral squamous cell carcinoma: A pilot study. Asian Pac. J. Cancer Prev. 2015, 16, 7619–7625. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Lee, J.S.; Oh, S.Y.; Jin, X.; Choi, Y.J.; Lee, T.H.; Lee, E.; Choi, Y.K.; You, S.; Chung, Y.G.; et al. Direct transcriptional activation of promyelocytic leukemia protein by IFN regulatory factor 3 induces the p53-dependent growth inhibition of cancer cells. Cancer Res. 2007, 67, 11133–11140. [Google Scholar] [CrossRef] [PubMed]
- Forys, J.T.; Kuzmicki, C.E.; Saporita, A.J.; Winkeler, C.L.; Maggi, L.J.; Weber, J.D. ARF and p53 coordinate tumor suppression of an oncogenic IFN-β-STAT1-ISG15 signaling axis. Cell Rep. 2014, 7, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Mancl, M.E.; Barnes, B.J. Signaling through IFN regulatory factor-5 sensitizes p53-deficient tumors to DNA damage-induced apoptosis and cell death. Cancer Res. 2005, 65, 7403–7412. [Google Scholar] [CrossRef] [PubMed]
- Wurzer, W.J.; Planz, O.; Ehrhardt, C.; Giner, M.; Silberzahn, T.; Pleschka, S.; Ludwig, S. Caspase 3 activation is essential for efficient influenza virus propagation. EMBO J. 2003, 22, 2717–2728. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.X.; Li, Y.; Sun, C.; Jiang, D.; Lin, Y.J.; Jin, F.X.; Lee, S.K.; Jin, Y.H. p53-dependent Fas expression is critical for Ginsenoside Rh2 triggered caspase-8 activation in HeLa cells. Protein Cell 2014, 5, 224–234. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence | References |
|---|---|---|
| Igfbp5 (WT)-Forward | 5′-CTCGAGGTCCTCCCCTCGCCCCATCCCATCC-3′ | NCBI Reference Sequence: XM_847792.4 |
| Igfbp5 (WT)-Reverse | 5′-GTTTAAACTAAATGAGATGAAATGAGTGGCGTC-3′ | |
| Igfbp5 (mut)-Forward | 5′-CATTTGAACTGATTTAGT-3′ (5′ phosphorylation) | |
| Igfbp5 (mut)-Forward | 5′-CAGTTCAAATGAGTGGCGTCCT-3′ (5′ phosphorylation) | |
| Igfbp5-Forward (qPCR) | 5′-GACTCCAGCCAGCACCT-3′ | |
| Igfbp5-Reverse (qPCR) | 5′-AGACCTTGCTAGCGATTCCGA-3′ | |
| TP53-Forward | 5′-ACAGTAGTGACGGTCTTGCC-3′ | GenBank: AB020761.1 |
| TP53-Reverse | 5′-CAACCTCGGGTGGCTCATAA-3′ | |
| bcl-2-Forward | 5′-CATGCCAAGAGGGAAACACCAGAA-3′ | [37] |
| bcl-2-Reverse | 5′-GTGCTTTGCATTCTTGGATGAGGG-3′ | |
| bax-Forward | 5′-TTCCGAGTGGCAGCTGAGATGTTT-3′ | |
| bax-Reverse | 5′-TGCTGGCAAAGTAGAAGAGGGCAA-3′ | |
| caspase3-Forward | 5′-TTCATTATTCAGGCCTGCCGAGG-3′ | |
| caspase3-Reverse | 5′-TTCTGACAGGCCATGTCATCCTCA-3′ | |
| caspase8-Forward | 5′-ACAAGGGCATCATCTATGGCTCTGA-3′ | |
| caspase8-Reverse | 5′-CCAGTGAAGTAAGAGGTCAGCTCAT-3′ | |
| caspase9-Forward | 5′-TCAGTGACGTCTGTGTTCAGGAGA-3′ | |
| caspase9-Reverse | 5′-TTGTTGATGATGAGGCAGTAGCCG-3′ | |
| GAPDH-Forward | 5′-TTCCACGGCACAGTCAAG-3′ | [38] |
| GAPDH-Reverse | 5′-ACTCAGCACCAGCATCAC-3′ |
| siRNA Name | Strand | Sequence |
|---|---|---|
| siR-caspase3 | positive-sense strand | 5′ GCAGCAAACCUCAGGGAAA dTdT 3′ |
| siR-caspase3 | antisense strand | 3′ dTdT CGUCGUUUGGAGUCCCUUU 5′ |
| siR-TP53 | positive-sense strand | 5′ CCAUCCACUACAACUACAU dTdT 3′ |
| siR-TP53 | antisense strand | 3′ dTdT GGUAGGUGAUGUUGAUGUA 5′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, P.; Tu, L.; Lin, X.; Hao, X.; Zheng, Q.; Zeng, W.; Zhang, X.; Zheng, Y.; Wang, L.; Li, S. cfa-miR-143 Promotes Apoptosis via the p53 Pathway in Canine Influenza Virus H3N2-Infected Cells. Viruses 2017, 9, 360. https://doi.org/10.3390/v9120360
Zhou P, Tu L, Lin X, Hao X, Zheng Q, Zeng W, Zhang X, Zheng Y, Wang L, Li S. cfa-miR-143 Promotes Apoptosis via the p53 Pathway in Canine Influenza Virus H3N2-Infected Cells. Viruses. 2017; 9(12):360. https://doi.org/10.3390/v9120360
Chicago/Turabian StyleZhou, Pei, Liqing Tu, Xi Lin, Xiangqi Hao, Qingxu Zheng, Weijie Zeng, Xin Zhang, Yun Zheng, Lifang Wang, and Shoujun Li. 2017. "cfa-miR-143 Promotes Apoptosis via the p53 Pathway in Canine Influenza Virus H3N2-Infected Cells" Viruses 9, no. 12: 360. https://doi.org/10.3390/v9120360