
Porcine Rotaviruses: Epidemiology, Immune Responses and Control Strategies



Abstract
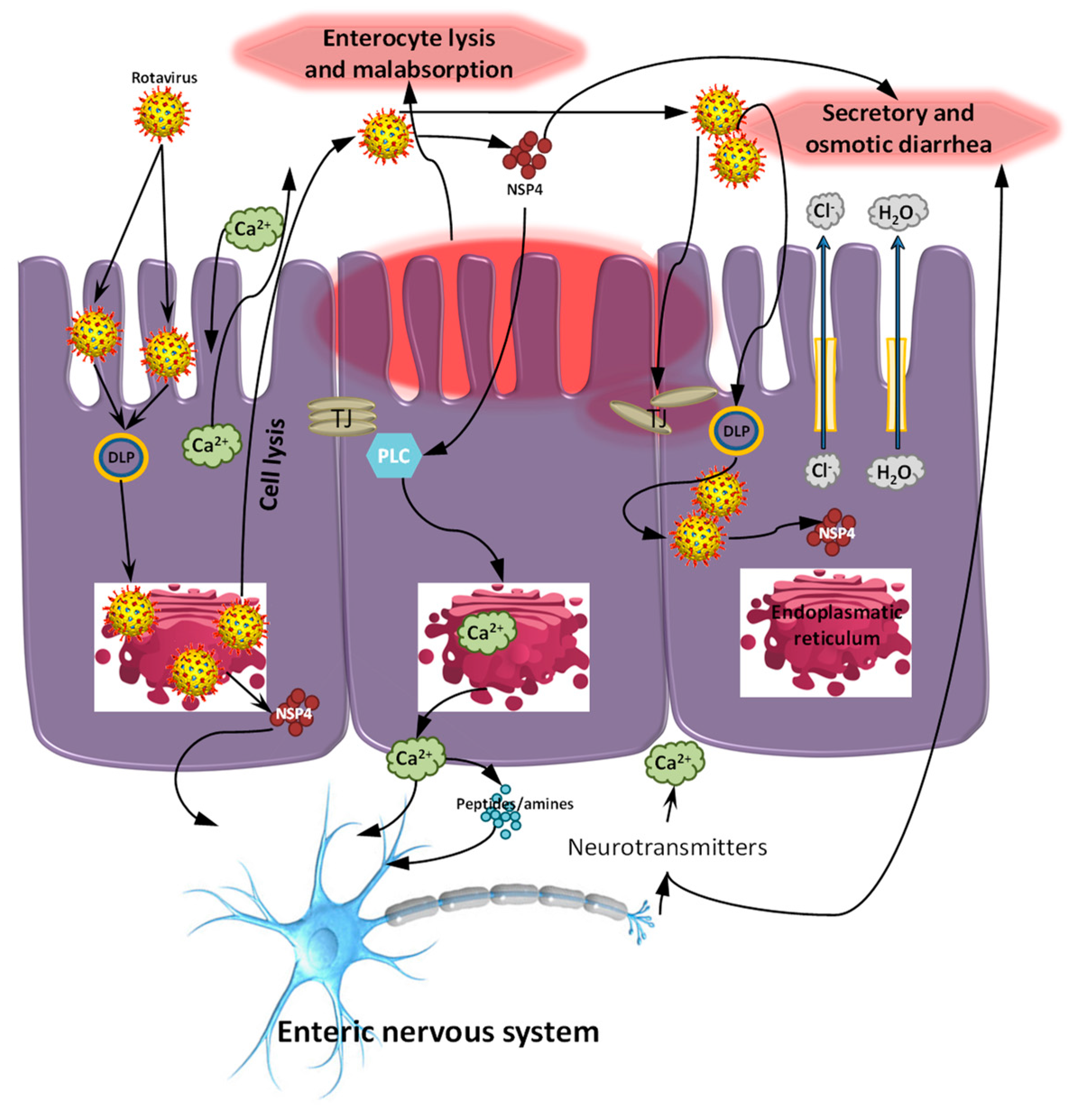
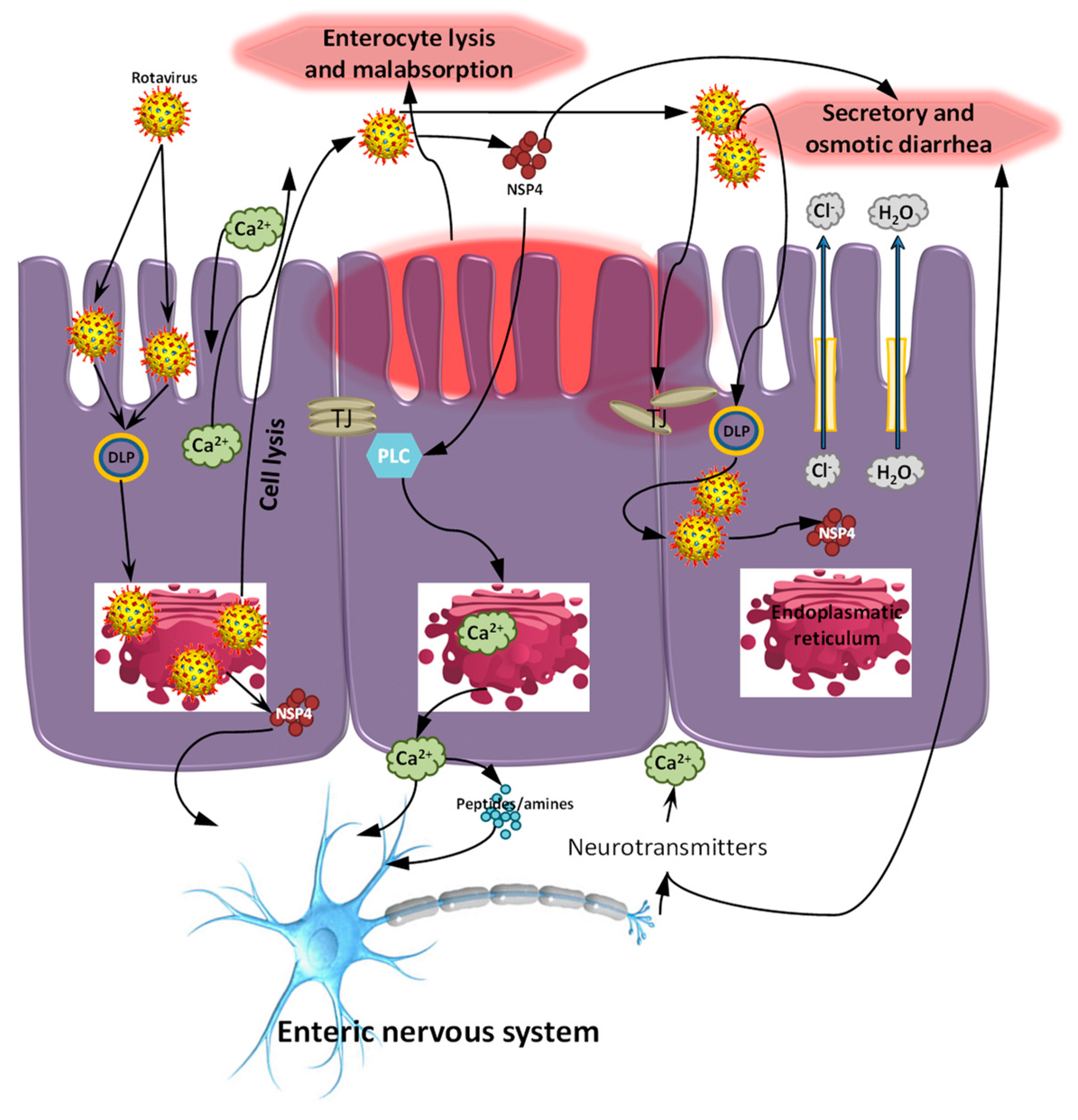
:1. Introduction
2. RV Genogroup/Genotype Classification and Prevalence in Swine
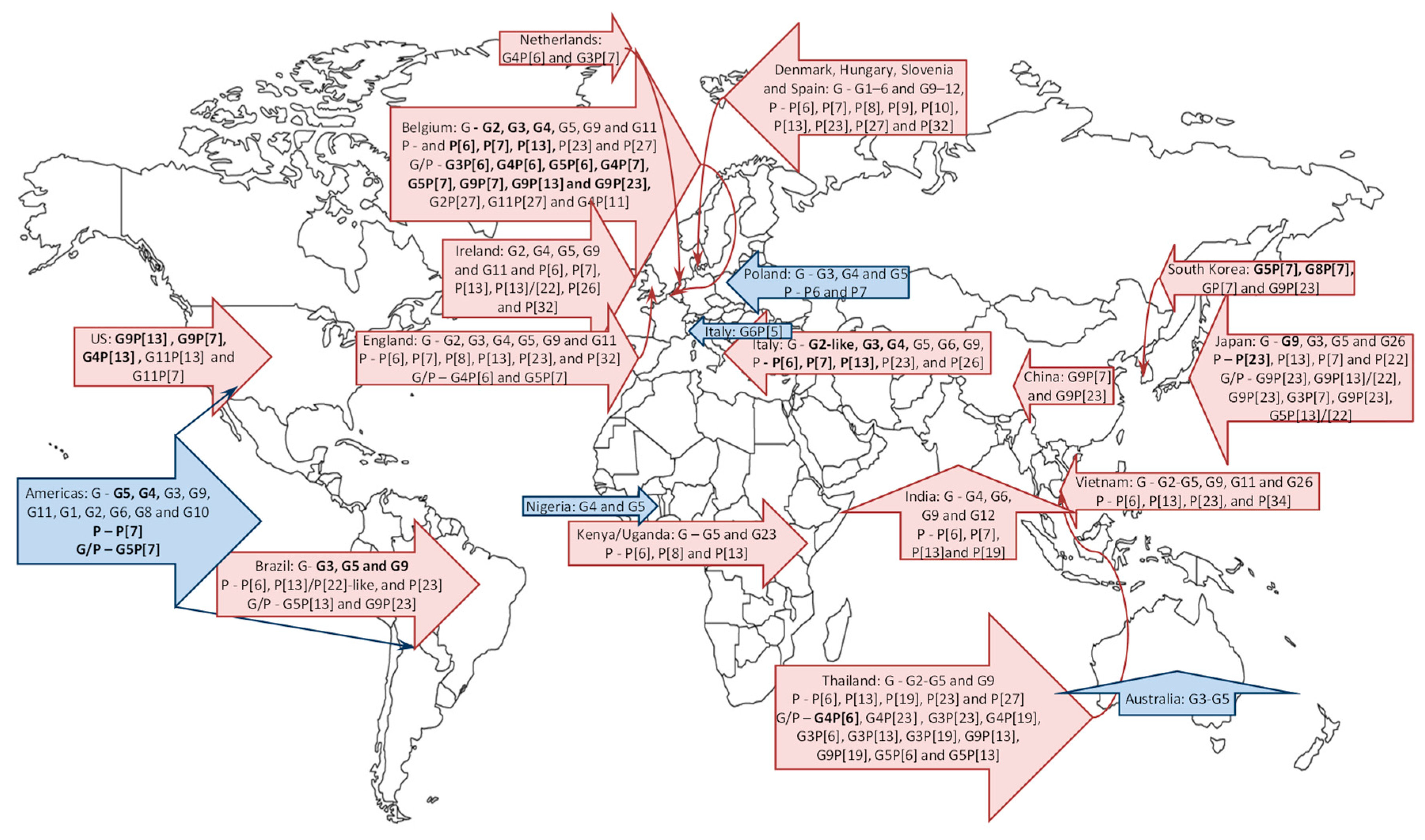
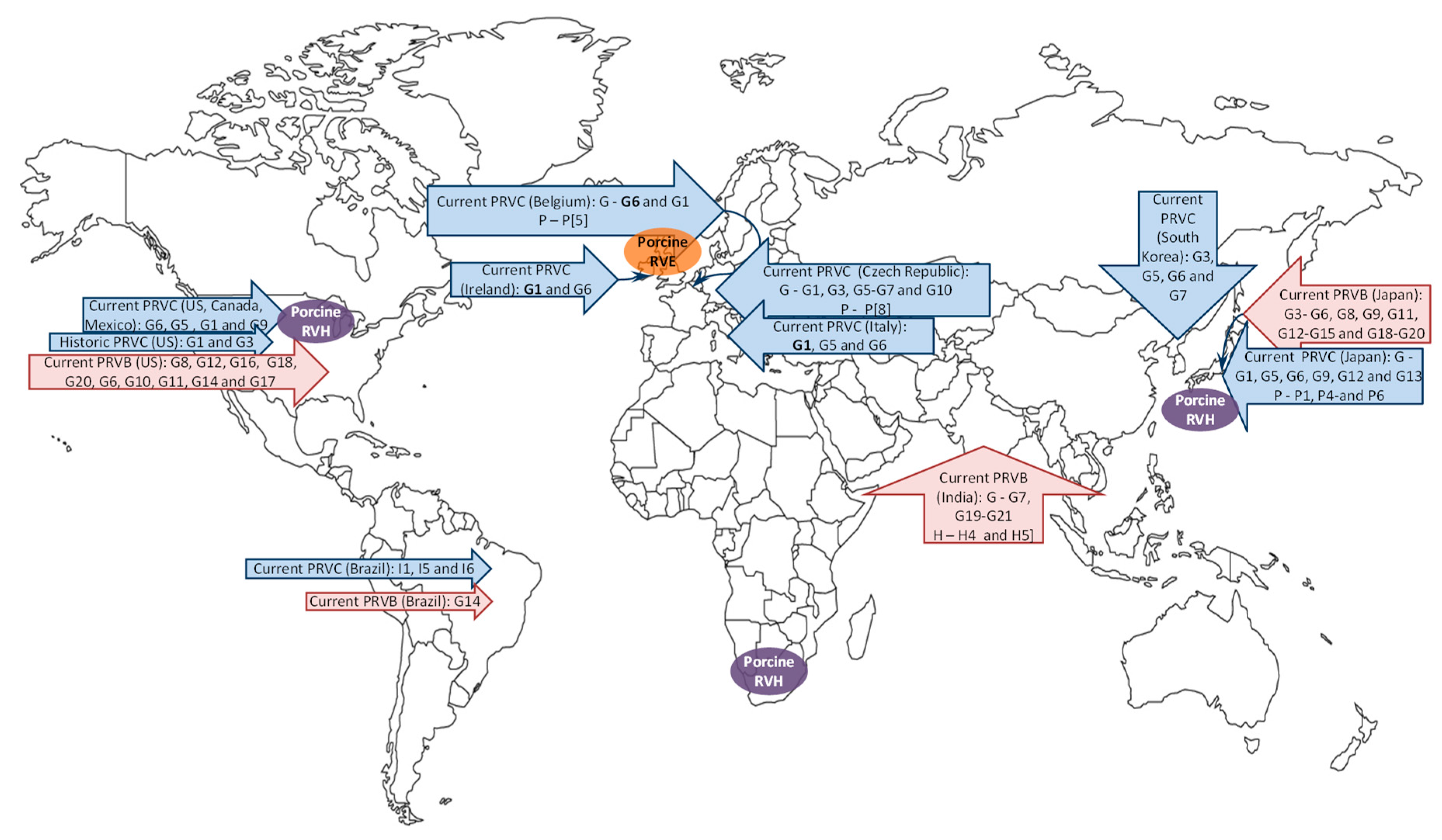
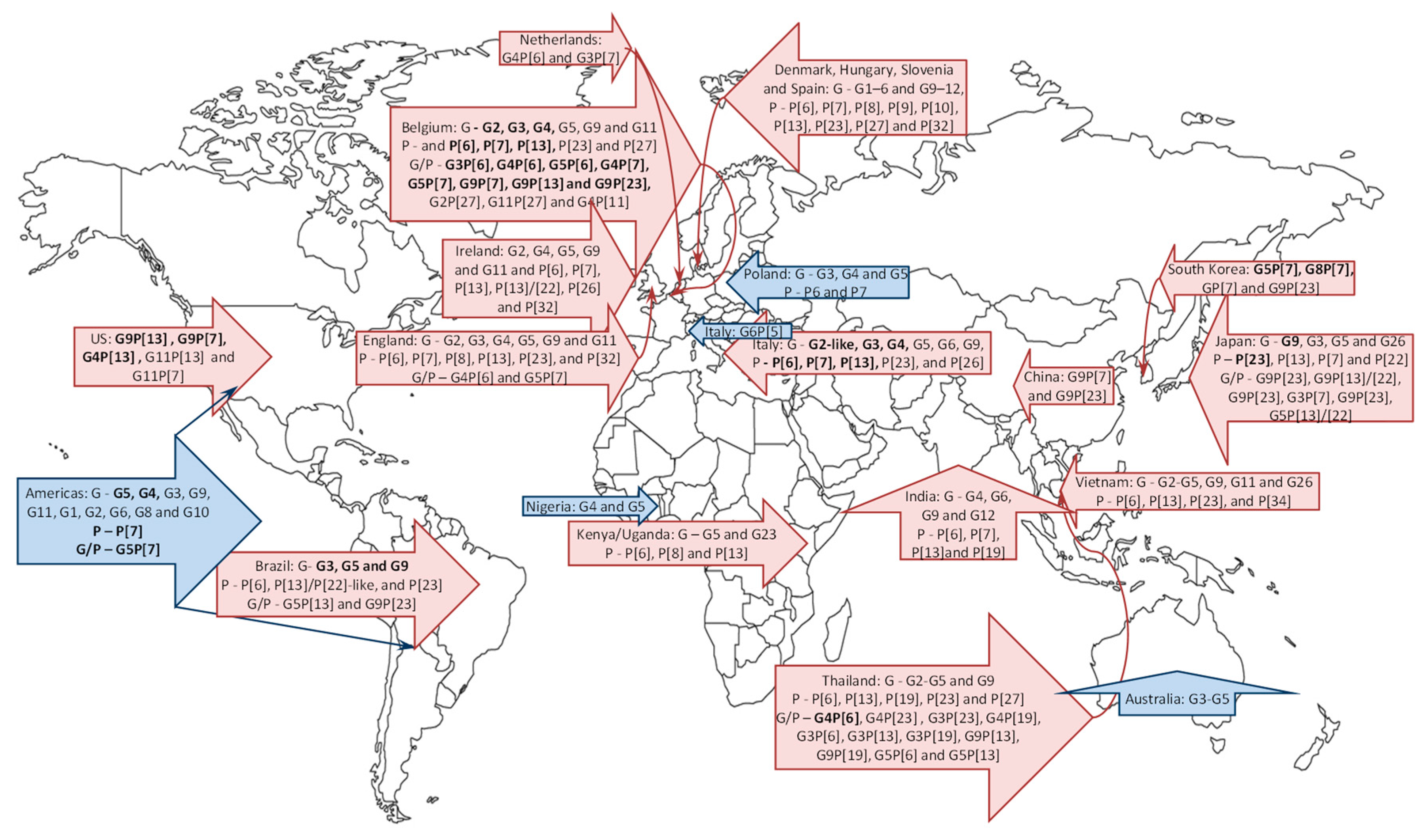
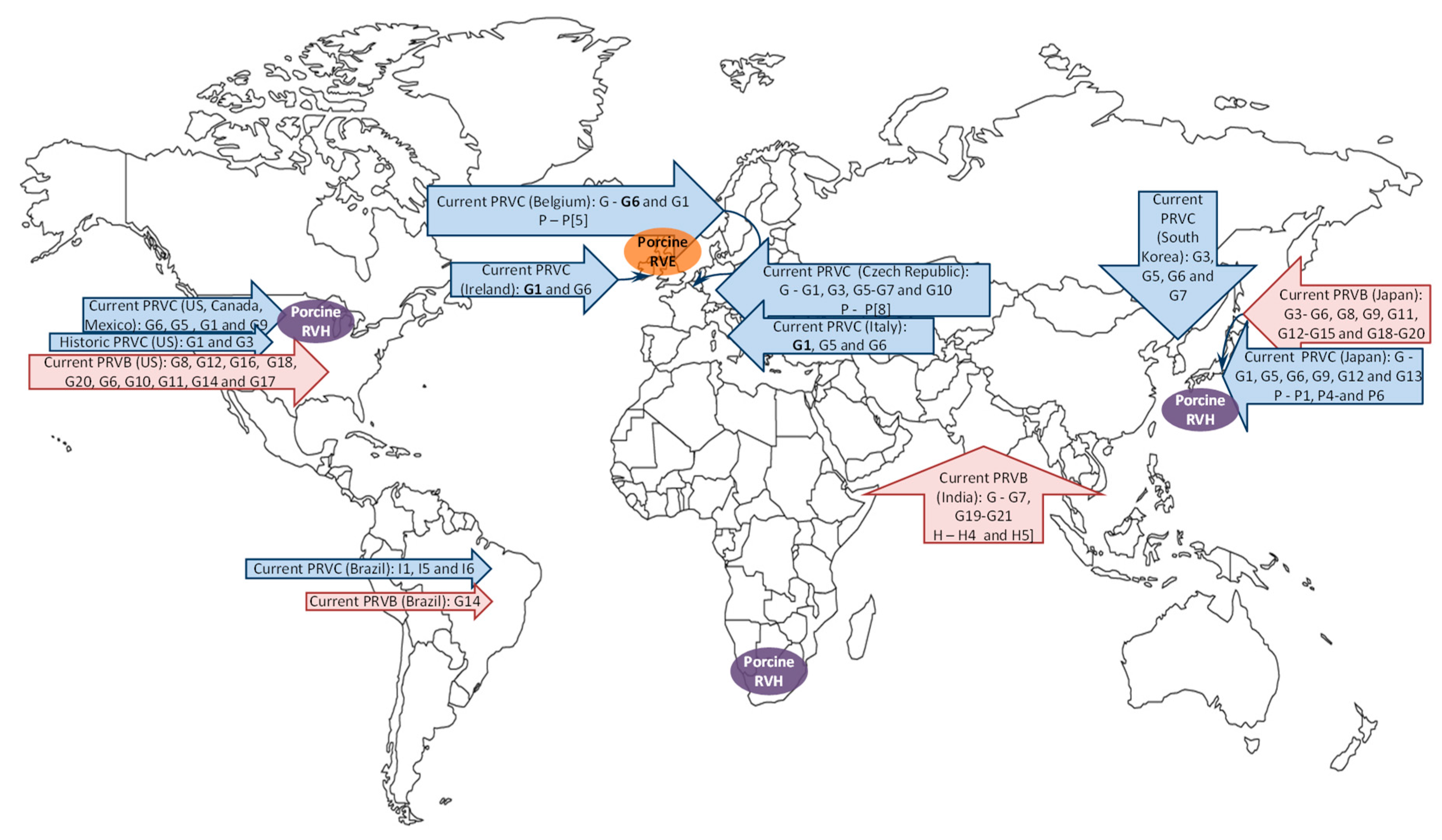
3. Porcine RV Distribution, Genotype Prevalence and Spatio-Temporal Variations in the Americas
3.1. North America
3.2. South America
4. Global Porcine RV Distribution and Genotype Prevalence: Africa, Europe, Asia and Australia
4.1. Africa
4.2. Europe
4.3. Asia
4.4. Australia
5. Zoonotic Potential of Porcine RV Strains
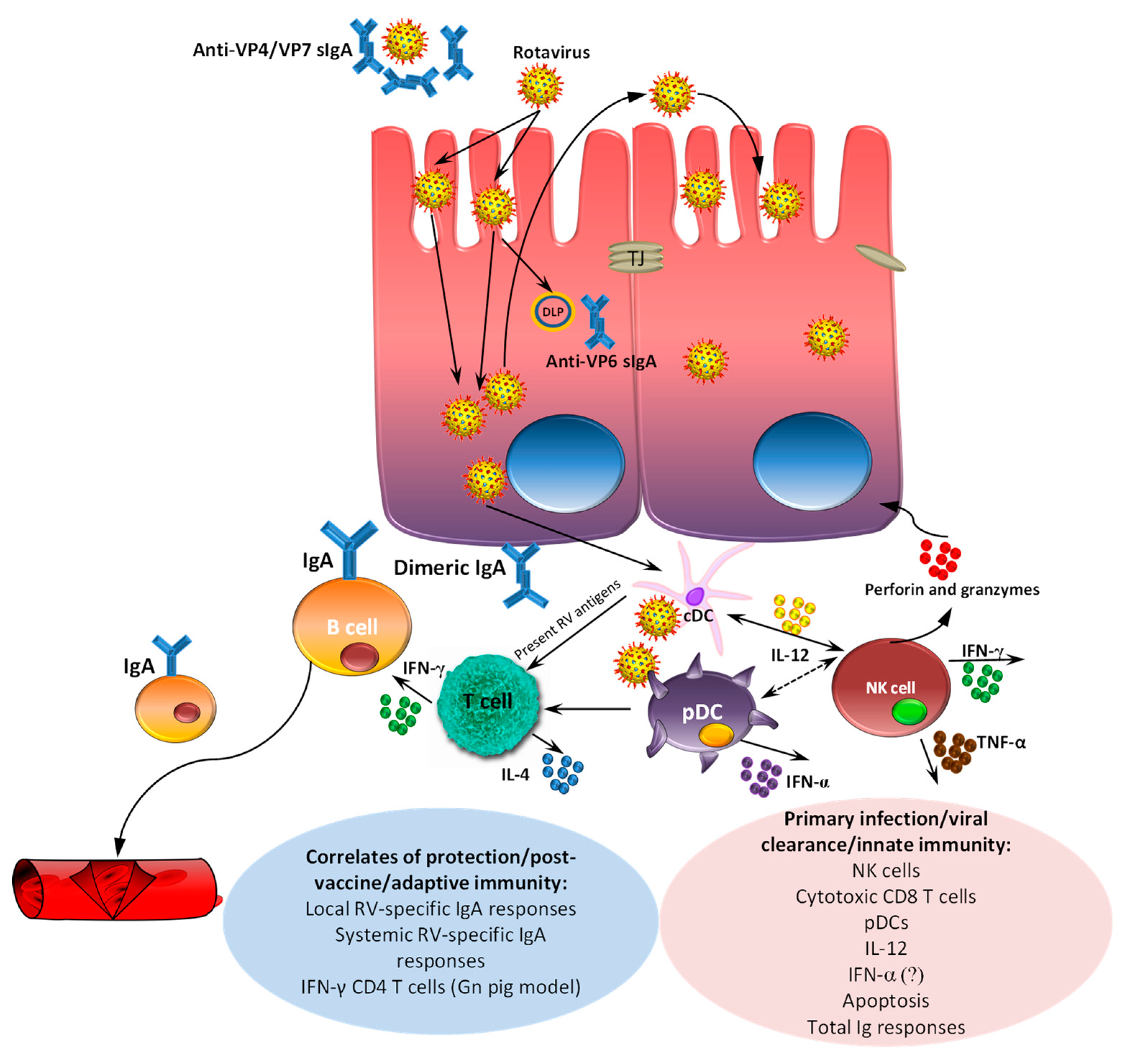
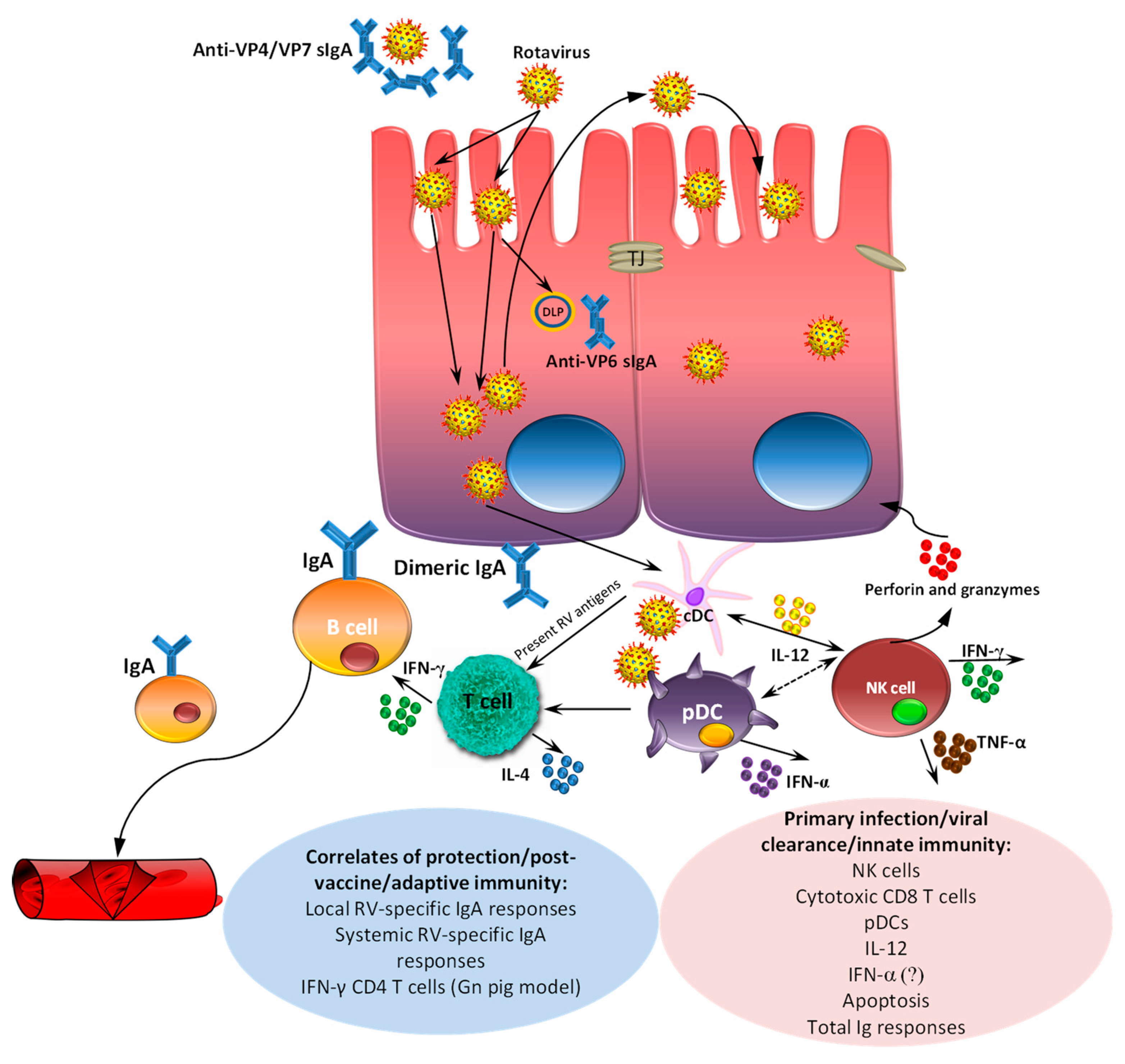
6. Passive and Active Immunity
7. Porcine RVA Vaccines and Control Strategies: Potential Impact of Vaccines on Porcine RVA Genetic Diversity
8. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chang, K.; Kim, Y.; Saif, L.J. Rotavirus and reovirus. In Diseases of Swine, 10 ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell: West Sussex, UK, 2012; pp. 621–634. [Google Scholar]
- Estes, M.K.; Kang, G.; Zeng, C.Q.; Crawford, S.E.; Ciarlet, M. Pathogenesis of rotavirus gastroenteritis. Novartis Found. Symp. 2001, 238, 82–96. [Google Scholar] [PubMed]
- Nava, P.; Lopez, S.; Arias, C.F.; Islas, S.; Gonzalez-Mariscal, L. The rotavirus surface protein VP8 modulates the gate and fence function of tight junctions in epithelial cells. J. Cell Sci. 2004, 117 Pt 23, 5509–5519. [Google Scholar] [CrossRef] [PubMed]
- Obert, G.; Peiffer, I.; Servin, A.L. Rotavirus-induced structural and functional alterations in tight junctions of polarized intestinal Caco-2 cell monolayers. J. Virol. 2000, 74, 4645–4651. [Google Scholar] [CrossRef] [PubMed]
- Dickman, K.G.; Hempson, S.J.; Anderson, J.; Lippe, S.; Zhao, L.; Burakoff, R.; Shaw, R.D. Rotavirus alters paracellular permeability and energy metabolism in Caco-2 cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G757–G766. [Google Scholar] [PubMed]
- Beau, I.; Cotte-Laffitte, J.; Amsellem, R.; Servin, A.L. A protein kinase A-dependent mechanism by which rotavirus affects the distribution and mRNA level of the functional tight junction-associated protein, occludin, in human differentiated intestinal Caco-2 cells. J. Virol. 2007, 81, 8579–8586. [Google Scholar] [CrossRef]
- Iturriza-Gomara, M.; Clarke, I.; Desselberger, U.; Brown, D.; Thomas, D.; Gray, J. Seroepidemiology of group C rotavirus infection in England and Wales. Eur. J. Epidemiol. 2004, 19, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Blutt, S.E.; Conner, M.E. Rotavirus: to the gut and beyond! Curr. Opin. Gastroenterol. 2007, 23, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.; Greenberg, H.B. Rotaviruses. In Fields Virology, 5 ed.; Knipe, D.M., Howley, P., Eds.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1347–1395. [Google Scholar]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef]
- Mihalov-Kovacs, E.; Gellert, A.; Marton, S.; Farkas, S.L.; Feher, E.; Oldal, M.; Jakab, F.; Martella, V.; Banyai, K. Candidate new rotavirus species in sheltered dogs, Hungary. Emerg. Infect. Dis. 2015, 21, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Banyai, K.; Kemenesi, G.; Budinski, I.; Foldes, F.; Zana, B.; Marton, S.; Varga-Kugler, R.; Oldal, M.; Kurucz, K.; Jakab, F. Candidate new rotavirus species in Schreiber’s bats, Serbia. Infect. Genet. Evol. 2017, 48, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Banyai, K.; Brister, J.R.; Buesa, J.; Esona, M.D.; Estes, M.K.; Gentsch, J.R.; et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- Trojnar, E.; Sachsenroder, J.; Twardziok, S.; Reetz, J.; Otto, P.H.; Johne, R. Identification of an avian group A rotavirus containing a novel VP4 gene with a close relationship to those of mammalian rotaviruses. J. Gen. Virol. 2013, 94 Pt 1, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gomara, M.; Maes, P.; Patton, J.T.; et al. Full genome-based classification of rotaviruses reveals a common origin between human Wa-Like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Banyai, K.; Estes, M.K.; Gentsch, J.R.; Iturriza-Gomara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the classification of group A rotaviruses using all 11 genomic RNA segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Banyai, K.; Matthijnssens, J.; Buonavoglia, C.; Ciarlet, M. Zoonotic aspects of rotaviruses. Vet. Microbiol. 2010, 140, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Suzuki, T.; Rossow, K.; Culhane, M.; Collins, J.; Goyal, S.; Tsunemitsu, H.; Ciarlet, M.; Matthijnssens, J. VP6 genetic diversity, reassortment, intragenic recombination and classification of rotavirus B in American and Japanese pigs. Vet. Microbiol. 2014, 172, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Taraporewala, Z.F.; Yang, H.; Rao, S.; Yuan, L.; Cao, D.; Hoshino, Y.; Mertens, P.P.; Carner, G.R.; McNeal, M.; et al. Simian rotaviruses possess divergent gene constellations that originated from interspecies transmission and reassortment. J. Virol. 2010, 84, 2013–2026. [Google Scholar] [CrossRef] [PubMed]
- McNulty, M.S. Rotaviruses. J. Gen. Virol. 1978, 40, 1–18. [Google Scholar] [CrossRef] [PubMed]
- McNulty, M.S.; Allan, G.M.; Connor, T.J.; McFerran, J.B.; McCracken, R.M. An entero-like virus associated with the runting syndrome in broiler chickens. Avian Pathol. 1984, 13, 429–439. [Google Scholar] [CrossRef] [PubMed]
- McNulty, M.S.; Allan, G.M.; McFerran, J.B. Prevalence of antibody to conventional and atypical rotaviruses in chickens. Vet. Rec. 1984, 114, 219. [Google Scholar] [CrossRef] [PubMed]
- McNulty, M.S.; Todd, D.; Allan, G.M.; McFerran, J.B.; Greene, J.A. Epidemiology of rotavirus infection in broiler chickens: recognition of four serogroups. Arch. Virol. 1984, 81, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Otto, P.; Liebler-Tenorio, E.M.; Elschner, M.; Reetz, J.; Lohren, U.; Diller, R. Detection of rotaviruses and intestinal lesions in broiler chicks from flocks with runting and stunting syndrome (RSS). Avian Dis. 2006, 50, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Chasey, D.; Bridger, J.C.; McCrae, M.A. A new type of atypical rotavirus in pigs. Arch. Virol. 1986, 89, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Janke, B.H.; Nelson, J.K.; Benfield, D.A.; Nelson, E.A. Relative prevalence of typical and atypical strains among rotaviruses from diarrheic pigs in conventional swine herds. J. Vet. Diagn. Investig. 1990, 2, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Wakuda, M.; Ide, T.; Sasaki, J.; Komoto, S.; Ishii, J.; Sanekata, T.; Taniguchi, K. Porcine rotavirus closely related to novel group of human rotaviruses. Emerg. Infect. Dis. 2011, 17, 1491–1493. [Google Scholar] [CrossRef]
- Bridger, J.C.; Woode, G.N. Neonatal calf diarrhoea: identification of a reovirus-like (rotavirus) agent in faeces by immunofluorescence and immune electron microscopy. Br. Vet. J. 1975, 131, 528–535. [Google Scholar] [PubMed]
- Bridger, J.C.; Brown, J.F. Prevalence of antibody to typical and atypical rotaviruses in pigs. Vet. Rec. 1985, 116, 50. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Rosen, B.; Parwani, A. Animal rotaviruses. In Virus Infections of the Gastrointestinal Tract; Kapikian, A.Z., Ed.; Marcel-Dekker: New York, NY, USA, 1994; pp. 279–367. [Google Scholar]
- Saif, L.J.; Bohl, E.H.; Theil, K.W.; Cross, R.F.; House, J.A. Rotavirus-like, calicivirus-like, and 23-nm virus-like particles associated with diarrhea in young pigs. J. Clin. Microbiol. 1980, 12, 105–111. [Google Scholar] [PubMed]
- Saif, L.J.; Jiang, B. Nongroup A rotaviruses of humans and animals. Curr. Top. Microbiol. Immunol. 1994, 185, 339–371. [Google Scholar]
- Mebus, C.A.; Underdahl, N.R.; Rhodes, M.B.; Twiehaus, M.J. Calf diarrhea (scours): reproduced with a virus from field outbreak. Neb. Agric. Exp. Stn. Res. Bull. 1969, 233, 1–16. [Google Scholar]
- Mebus, C.A.; Underdahl, N.R.; Rhodes, M.B.; Twiehaus, M.J. Further studies on neonatal calf diarrhea virus. Proc. Annu. Meet. U. S. Anim. Health Assoc. 1969, 73, 97–99. [Google Scholar] [PubMed]
- Bishop, R.F.; Davidson, G.P.; Holmes, I.H.; Ruck, B.J. Virus particles in epithelial cells of duodenal mucosa from children with acute non-bacterial gastroenteritis. Lancet 1973, 2, 1281–1283. [Google Scholar] [CrossRef]
- Theil, K.W. Group A rotaviruses. In Viral Diarrheas of Man and Animals; Saif, L.J., Theil, K.W., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 35–72. [Google Scholar]
- Holmes, I.H. Rotaviruses. In The Reoviridae; Joklik, W.T., Ed.; Plenum Press: New York, NY, USA, 1983; pp. 359–423. [Google Scholar]
- Rodger, S.M.; Bishop, R.F.; Holmes, I.H. Detection of a rotavirus-like agent associated with diarrhea in an infant. J. Clin. Microbiol. 1982, 16, 724–726. [Google Scholar] [PubMed]
- Bridger, J.C.; Pedley, S.; McCrae, M.A. Group C rotaviruses in humans. J. Clin. Microbiol. 1986, 23, 760–763. [Google Scholar] [PubMed]
- Torres-Medina, A. Isolation of an atypical rotavirus causing diarrhea in neonatal ferrets. Lab. Anim. Sci. 1987, 37, 167–171. [Google Scholar] [PubMed]
- Tsunemitsu, H.; Saif, L.J.; Jiang, B.M.; Shimizu, M.; Hiro, M.; Yamaguchi, H.; Ishiyama, T.; Hirai, T. Isolation, characterization, and serial propagation of a bovine group C rotavirus in a monkey kidney cell line (MA104). J. Clin. Microbiol. 1991, 29, 2609–2613. [Google Scholar] [PubMed]
- Theil, K.W.; Saif, L.J.; Moorhead, P.D.; Whitmoyer, R.E. Porcine rotavirus-like virus (group B rotavirus): characterization and pathogenicity for gnotobiotic pigs. J. Clin. Microbiol. 1985, 21, 340–345. [Google Scholar] [PubMed]
- Barman, P.; Ghosh, S.; Das, S.; Varghese, V.; Chaudhuri, S.; Sarkar, S.; Krishnan, T.; Bhattacharya, S.K.; Chakrabarti, A.; Kobayashi, N.; et al. Sequencing and sequence analysis of VP7 and NSP5 genes reveal emergence of a new genotype of bovine group B rotaviruses in India. J. Clin. Microbiol. 2004, 42, 2816–2818. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.O.; Parwani, A.V.; Smith, D.; Saif, L.J. Detection of group B rotaviruses in fecal samples from diarrheic calves and adult cows and characterization of their VP7 genes. J. Clin. Microbiol. 1997, 35, 2107–2110. [Google Scholar] [PubMed]
- Ghosh, S.; Varghese, V.; Sinha, M.; Kobayashi, N.; Naik, T.N. Evidence for interstate transmission and increase in prevalence of bovine group B rotavirus strains with a novel VP7 genotype among diarrhoeic calves in Eastern and Northern states of India. Epidemiol. Infect. 2007, 135, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Tsunemitsu, H.; Morita, D.; Takaku, H.; Nishimori, T.; Imai, K.; Saif, L.J. First detection of bovine group B rotavirus in Japan and sequence of its VP7 gene. Arch. Virol. 1999, 144, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; McKee, T.A.; Wang, Z.D.; Desselberger, U.; Liu, D.X. Sequence analysis and in vitro expression of genes 6 and 11 of an ovine group B rotavirus isolate, KB63: Evidence for a non-defective, C-terminally truncated NSP1 and a phosphorylated NSP5. J. Gen. Virol. 1999, 80 Pt 8, 2077–2085. [Google Scholar] [CrossRef]
- Eiden, J.J.; Nataro, J.; Vonderfecht, S.; Petric, M. Molecular cloning, sequence analysis, in vitro expression, and immunoprecipitation of the major inner capsid protein of the IDIR strain of group B rotavirus (GBR). Virology 1992, 188, 580–589. [Google Scholar] [CrossRef]
- Chen, G.M.; Hung, T.; Mackow, E.R. Identification of the gene encoding the group B rotavirus VP7 equivalent: primary characterization of the ADRV segment 9 RNA. Virology 1990, 178, 311–315. [Google Scholar] [CrossRef]
- Dai, G.Z.; Sun, M.S.; Liu, S.Q.; Ding, X.F.; Chen, Y.D.; Wang, L.C.; Du, D.P.; Zhao, G.; Su, Y.; Li, J.; et al. First report of an epidemic of diarrhoea in human neonates involving the new rotavirus and biological characteristics of the epidemic virus strain (KMB/R85). J. Med. Virol. 1987, 22, 365–373. [Google Scholar] [PubMed]
- Fang, Z.Y.; Ye, Q.; Ho, M.S.; Dong, H.; Qing, S.; Penaranda, M.E.; Hung, T.; Wen, L.; Glass, R.I. Investigation of an outbreak of adult diarrhea rotavirus in China. J. Infect. Dis. 1989, 160, 948–953. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.; Chen, G.M.; Wang, C.G.; Yao, H.L.; Fang, Z.Y.; Chao, T.X.; Chou, Z.Y.; Ye, W.; Chang, X.J.; Den, S.S.; et al. Waterborne outbreak of rotavirus diarrhoea in adults in China caused by a novel rotavirus. Lancet 1984, 1, 1139–1142. [Google Scholar]
- Kelkar, S.D.; Zade, J.K. Group B rotaviruses similar to strain CAL-1, have been circulating in Western India since 1993. Epidemiol. Infect. 2004, 132, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Lahon, A.; Chitambar, S.D. Molecular characterization of VP4, VP6, VP7 and NSP4 genes of group B rotavirus strains from outbreaks of gastroenteritis. Asian Pac. J. Trop. Med. 2011, 4, 846–849. [Google Scholar] [CrossRef]
- Ahmed, M.U.; Kobayashi, N.; Wakuda, M.; Sanekata, T.; Taniguchi, K.; Kader, A.; Naik, T.N.; Ishino, M.; Alam, M.M.; Kojima, K.; et al. Genetic analysis of group B human rotaviruses detected in Bangladesh in 2000 and 2001. J. Med. Virol. 2004, 72, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Saiada, F.; Rahman, H.N.; Moni, S.; Karim, M.M.; Pourkarim, M.R.; Azim, T.; Rahman, M. Clinical presentation and molecular characterization of group B rotaviruses in diarrhoea patients in Bangladesh. J. Med. Microbiol. 2011, 60 Pt 4, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Hassan, Z.M.; Zafrul, H.; Saiada, F.; Banik, S.; Faruque, A.S.; Delbeke, T.; Matthijnssens, J.; Van Ranst, M.; Azim, T. Sequence analysis and evolution of group B rotaviruses. Virus Res. 2007, 125, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Sanekata, T.; Ahmed, M.U.; Kader, A.; Taniguchi, K.; Kobayashi, N. Human group B rotavirus infections cause severe diarrhea in children and adults in Bangladesh. J. Clin. Microbiol. 2003, 41, 2187–2190. [Google Scholar] [CrossRef] [PubMed]
- Aung, T.S.; Kobayashi, N.; Nagashima, S.; Ghosh, S.; Aung, M.S.; Oo, K.Y.; Win, N. Detection of group B rotavirus in an adult with acute gastroenteritis in Yangon, Myanmar. J. Med. Virol. 2009, 81, 1968–1974. [Google Scholar] [CrossRef] [PubMed]
- Chasey, D.; Davies, P. Atypical rotaviruses in pigs and cattle. Vet. Rec. 1984, 114, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Nelson, M.; Ciarlet, M. Widespread rotavirus H in commercially raised pigs, United States. Emerg. Infect. Dis. 2014, 20, 1195–1198. [Google Scholar] [CrossRef] [PubMed]
- Molinari, B.L.; Lorenzetti, E.; Otonel, R.A.; Alfieri, A.F.; Alfieri, A.A. Species H rotavirus detected in piglets with diarrhea, Brazil, 2012. Emerg. Infect. Dis. 2014, 20, 1019–1022. [Google Scholar] [CrossRef] [PubMed]
- Chinivasagam, H.N.; Thomas, R.J.; Casey, K.; McGahan, E.; Gardner, E.A.; Rafiee, M.; Blackall, P.J. Microbiological status of piggery effluent from 13 piggeries in the south east Queensland region of Australia. J. Appl. Microbiol. 2004, 97, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; El Zowalaty, M.E.; Githae, D.; Wamalwa, M.; Djikeng, A.; Nasrallah, G.K. Metagenomic analysis demonstrates the diversity of the fecal virome in asymptomatic pigs in East Africa. Arch. Virol. 2016, 161, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; Junga, J.O.; Ogara, W.O.; Vlasova, A.N.; Njahira, M.N.; Maina, S.; Okoth, E.A.; Bishop, R.P.; Saif, L.J.; Djikeng, A. Detection and genetic characterization of porcine group A rotaviruses in asymptomatic pigs in smallholder farms in East Africa: Predominance of P[8] genotype resembling human strains. Vet. Microbiol. 2015, 175, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; Otieno, T.F.; Okoth, E.; Onono, J.O.; Bett, B. Risk factors for rotavirus infection in pigs in Busia and Teso subcounties, Western Kenya. Trop. Anim. Health Prod. 2017, 49, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; Vlasova, A.N.; Saif, L.J. Detection and genetic diversity of porcine group A rotaviruses in historic (2004) and recent (2011 and 2012) swine fecal samples in Ohio: predominance of the G9P[13] genotype in nursing piglets. J. Clin. Microbiol. 2013, 51, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.F.; Tort, L.F.; Gomez, M.M.; Assis, R.M.; de Mendonca, M.C.; Volotao Ede, M.; Leite, J.P. Phylogenetic analysis of VP1, VP2, and VP3 gene segments of genotype G5 group A rotavirus strains circulating in Brazil between 1986 and 2005. Virus Res. 2011, 160, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Homwong, N.; Diaz, A.; Rossow, S.; Ciarlet, M.; Marthaler, D. Three-Level Mixed-Effects Logistic Regression Analysis Reveals Complex Epidemiology of Swine Rotaviruses in Diagnostic Samples from North America. PLoS ONE 2016, 11, e0154734. [Google Scholar]
- Martella, V.; Ciarlet, M.; Banyai, K.; Lorusso, E.; Arista, S.; Lavazza, A.; Pezzotti, G.; Decaro, N.; Cavalli, A.; Lucente, M.S.; et al. Identification of group A porcine rotavirus strains bearing a novel VP4 (P) Genotype in Italian swine herds. J. Clin. Microbiol. 2007, 45, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Homwong, N.; Rossow, K.; Culhane, M.; Goyal, S.; Collins, J.; Matthijnssens, J.; Ciarlet, M. Rapid detection and high occurrence of porcine rotavirus A, B, and C by RT-qPCR in diagnostic samples. J. Virol. Methods 2014, 209, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Papp, H.; Matthijnssens, J.; Martella, V.; Ciarlet, M.; Banyai, K. Global distribution of group A rotavirus strains in horses: a systematic review. Vaccine 2013, 31, 5627–5633. [Google Scholar] [CrossRef] [PubMed]
- Theuns, S.; Vyt, P.; Desmarets, L.M.; Roukaerts, I.D.; Heylen, E.; Zeller, M.; Matthijnssens, J.; Nauwynck, H.J. Presence and characterization of pig group A and C rotaviruses in feces of Belgian diarrheic suckling piglets. Virus Res. 2016, 213, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.A.; Carrique-Mas, J.J.; Nguyen, V.C.; Ngo, T.H.; Nguyet, L.A.; Do, T.D.; Vo, B.H.; Phan, V.T.; Rabaa, M.A.; Farrar, J.; et al. The prevalence and genetic diversity of group A rotaviruses on pig farms in the Mekong Delta region of Vietnam. Vet. Microbiol. 2014, 170, 258–265. [Google Scholar] [PubMed]
- Halaihel, N.; Masia, R.M.; Fernandez-Jimenez, M.; Ribes, J.M.; Montava, R.; De Blas, I.; Girones, O.; Alonso, J.L.; Buesa, J. Enteric calicivirus and rotavirus infections in domestic pigs. Epidemiol. Infect. 2010, 138, 542–548. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Park, S.I.; Ha, T.P.; Jeong, Y.J.; Kim, H.H.; Kwon, H.J.; Kang, M.I.; Cho, K.O.; Park, S.J. Detection and genotyping of Korean porcine rotaviruses. Vet. Microbiol. 2010, 144, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, K.; Kohmoto, M.; Kawashima, K.; Tsunemitsu, H. Frequency of enteropathogen detection in suckling and weaned pigs with diarrhea in Japan. J. Vet. Diagn. Investig. 2006, 18, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Saikruang, W.; Khamrin, P.; Chaimongkol, N.; Suantai, B.; Kongkaew, A.; Kongkaew, S.; Ushijima, H.; Maneekarn, N. Genetic diversity and novel combinations of G4P[19] and G9P[19] porcine rotavirus strains in Thailand. Vet. Microbiol. 2013, 161, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Lamhoujeb, S.; Cook, A.; Pollari, F.; Bidawid, S.; Farber, J.; Mattison, K. Rotaviruses from Canadian farm samples. Arch. Virol. 2010, 155, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Racz, M.L.; Kroeff, S.S.; Munford, V.; Caruzo, T.A.; Durigon, E.L.; Hayashi, Y.; Gouvea, V.; Palombo, E.A. Molecular characterization of porcine rotaviruses from the southern region of Brazil: characterization of an atypical genotype G[9] strain. J. Clin. Microbiol. 2000, 38, 2443–2446. [Google Scholar] [PubMed]
- Kusumakar, A.L.; Savita; Malik, Y.S.; Minakshi; Prasad, G. Genomic diversity among group A rotaviruses from diarrheic children, piglets, buffalo and cow calves of Madhya Pradesh. Indian J. Microbiol. 2010, 50, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Parra, G.I.; Vidales, G.; Gomez, J.A.; Fernandez, F.M.; Parreno, V.; Bok, K. Phylogenetic analysis of porcine rotavirus in Argentina: increasing diversity of G4 strains and evidence of interspecies transmission. Vet. Microbiol. 2008, 126, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Wieler, L.H.; Ilieff, A.; Herbst, W.; Bauer, C.; Vieler, E.; Bauerfeind, R.; Failing, K.; Klos, H.; Wengert, D.; Baljer, G.; et al. Prevalence of enteropathogens in suckling and weaned piglets with diarrhoea in southern Germany. J. Vet. Med. B Infect. Dis. Vet. Public Health 2001, 48, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.J.; Martella, V.; Sleator, R.D.; Fanning, S.; O’Shea, H. Detection and characterisation of group A rotavirus in asymptomatic piglets in southern Ireland. Arch. Virol. 2010, 155, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Midgley, S.E.; Banyai, K.; Buesa, J.; Halaihel, N.; Hjulsager, C.K.; Jakab, F.; Kaplon, J.; Larsen, L.E.; Monini, M.; Poljsak-Prijatelj, M.; et al. Diversity and zoonotic potential of rotaviruses in swine and cattle across Europe. Vet. Microbiol. 2012, 156, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Morin, M.; Turgeon, D.; Jolette, J.; Robinson, Y.; Phaneuf, J.B.; Sauvageau, R.; Beauregard, M.; Teuscher, E.; Higgins, R.; Lariviere, S. Neonatal diarrhea of pigs in Quebec: Infectious causes of significant outbreaks. Can. J. Comp. Med. 1983, 47, 11–17. [Google Scholar] [PubMed]
- Khamrin, P.; Peerakome, S.; Tonusin, S.; Malasao, R.; Okitsu, S.; Mizuguchi, M.; Ushijima, H.; Maneekarn, N. Changing pattern of rotavirus G genotype distribution in Chiang Mai, Thailand from 2002 to 2004: Decline of G9 and reemergence of G1 and G2. J. Med. Virol. 2007, 79, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Okitsu, S.; Khamrin, P.; Thongprachum, A.; Kongkaew, A.; Maneekarn, N.; Mizuguchi, M.; Hayakawa, S.; Ushijima, H. Whole-genomic analysis of G3P[23], G9P[23] and G3P[13] rotavirus strains isolated from piglets with diarrhea in Thailand, 2006–2008. Infect. Genet. Evol. 2013, 18, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Okitsu, S.; Khamrin, P.; Thongprachum, A.; Maneekarn, N.; Mizuguchi, M.; Ushijima, H. Predominance of porcine P[23] genotype rotaviruses in piglets with diarrhea in northern Thailand. J. Clin. Microbiol. 2011, 49, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, A.; Kuga, K.; Suzuki, T.; Kohmoto, M.; Katsuda, K.; Tsunemitsu, H. Genetic diversity of group A rotaviruses associated with repeated outbreaks of diarrhea in a farrow-to-finish farm: identification of a porcine rotavirus strain bearing a novel VP7 genotype, G26. Vet. Res. 2011, 42, 112. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.J.; Martella, V.; Buonavoglia, C.; O’Shea, H. Identification of a G2-like porcine rotavirus bearing a novel VP4 type, P[32]. Vet. Res. 2010, 41, 73. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Kobayashi, N.; Nagashima, S.; Zhou, X.; Ghosh, S.; Peng, J.S.; Hu, Q.; Zhou, D.J.; Yang, Z.Q. Full genomic analysis of a porcine-bovine reassortant G4P[6] rotavirus strain R479 isolated from an infant in China. J. Med. Virol. 2010, 82, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Chang, K.O.; Straw, B.; Saif, L.J. Characterization of group C rotaviruses associated with diarrhea outbreaks in feeder pigs. J. Clin. Microbiol. 1999, 37, 1484–1488. [Google Scholar] [PubMed]
- Marthaler, D.; Rossow, K.; Culhane, M.; Collins, J.; Goyal, S.; Ciarlet, M.; Matthijnssens, J. Identification, phylogenetic analysis and classification of porcine group C rotavirus VP7 sequences from the United States and Canada. Virology 2013, 446, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Tsunemitsu, H.; Jiang, B.; Saif, L.J. Sequence comparison of the VP7 gene encoding the outer capsid glycoprotein among animal and human group C rotaviruses. Arch. Virol. 1996, 141, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Banik, S.; Faruque, A.S.; Taniguchi, K.; Sack, D.A.; van Ranst, M.; Azim, T. Detection and characterization of human group C rotaviruses in Bangladesh. J. Clin. Microbiol. 2005, 43, 4460–4465. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Banyai, K.; Lorusso, E.; Decaro, N.; Bellacicco, A.; Desario, C.; Corrente, M.; Greco, G.; Moschidou, P.; Tempesta, M.; et al. Genetic heterogeneity in the VP7 of group C rotaviruses. Virology 2007, 367, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Stipp, D.T.; Alfieri, A.F.; Lorenzetti, E.; da Silva Medeiros, T.N.; Possatti, F.; Alfieri, A.A. VP6 gene diversity in 11 Brazilian strains of porcine group C rotavirus. Virus Genes 2015, 50, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hasebe, A.; Miyazaki, A.; Tsunemitsu, H. Phylogenetic characterization of VP6 gene (inner capsid) of porcine rotavirus C collected in Japan. Infect. Genet. Evol. 2014, 26, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hasebe, A.; Miyazaki, A.; Tsunemitsu, H. Analysis of genetic divergence among strains of porcine rotavirus C, with focus on VP4 and VP7 genotypes in Japan. Virus Res. 2015, 197, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Soma, J.; Tsunemitsu, H.; Miyamoto, T.; Suzuki, G.; Sasaki, T.; Suzuki, T. Whole-genome analysis of two bovine rotavirus C strains: Shintoku and Toyama. J. Gen. Virol. 2013, 94 Pt 1, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, D.; Ghosh, S.; Kuzuya, M.; Wang, Y.H.; Zhou, X.; Chawla-Sarkar, M.; Paul, S.K.; Ishino, M.; Kobayashi, N. Whole-genome characterization of human group C rotaviruses: identification of two lineages in the VP3 gene. J. Gen. Virol. 2011, 92 Pt 2, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moutelikova, R.; Prodelalova, J.; Dufkova, L. Diversity of VP7, VP4, VP6, NSP2, NSP4, and NSP5 genes of porcine rotavirus C: phylogenetic analysis and description of potential new VP7, VP4, VP6, and NSP4 genotypes. Arch. Virol. 2015, 160, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Collins, P.J.; Martella, V.; O’Shea, H. Detection and characterization of group C rotaviruses in asymptomatic piglets in Ireland. J. Clin. Microbiol. 2008, 46, 2973–2979. [Google Scholar] [CrossRef] [PubMed]
- Niira, K.; Ito, M.; Masuda, T.; Saitou, T.; Abe, T.; Komoto, S.; Sato, M.; Yamasato, H.; Kishimoto, M.; Naoi, Y.; et al. Whole genome sequences of Japanese porcine species C rotaviruses reveal a high diversity of genotypes of individual genes and will contribute to a comprehensive, generally accepted classification system. Infect. Genet. Evol. 2016, 44, 106–113. [Google Scholar] [CrossRef]
- Kobayashi, N.; Naik, T.N.; Kusuhara, Y.; Krishnan, T.; Sen, A.; Bhattacharya, S.K.; Taniguchi, K.; Alam, M.M.; Urasawa, T.; Urasawa, S. Sequence analysis of genes encoding structural and nonstructural proteins of a human group B rotavirus detected in Calcutta, India. J. Med. Virol. 2001, 64, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, D.; Ghosh, S.; Ganesh, B.; Krishnan, T.; Chawla-Sarkar, M.; Alam, M.M.; Aung, T.S.; Kobayashi, N. Analysis of genetic diversity and molecular evolution of human group B rotaviruses based on whole genome segments. J. Gen. Virol. 2010, 91 Pt 7, 1772–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.H.; Kobayashi, N.; Wang, Y.H.; Zhou, X.; Li, Y.; Zhou, D.J.; Hu, Z.H.; Ishino, M.; Alam, M.M.; Naik, T.N.; et al. Phylogenetic analysis of a human group B rotavirus WH-1 detected in China in 2002. J. Med. Virol. 2004, 74, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Petric, M.; Mayur, K.; Vonderfecht, S.; Eiden, J.J. Comparison of group B rotavirus genes 9 and 11. J. Gen. Virol. 1991, 72 Pt 11, 2801–2804. [Google Scholar] [CrossRef] [PubMed]
- Kuga, K.; Miyazaki, A.; Suzuki, T.; Takagi, M.; Hattori, N.; Katsuda, K.; Mase, M.; Sugiyama, M.; Tsunemitsu, H. Genetic diversity and classification of the outer capsid glycoprotein VP7 of porcine group B rotaviruses. Arch. Virol. 2009, 154, 1785–1795. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Rossow, K.; Gramer, M.; Collins, J.; Goyal, S.; Tsunemitsu, H.; Kuga, K.; Suzuki, T.; Ciarlet, M.; Matthijnssens, J. Detection of substantial porcine group B rotavirus genetic diversity in the United States, resulting in a modified classification proposal for G genotypes. Virology 2012, 433, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Molinari, B.L.; Possatti, F.; Lorenzetti, E.; Alfieri, A.F.; Alfieri, A.A. Unusual outbreak of post-weaning porcine diarrhea caused by single and mixed infections of rotavirus groups A, B, C, and H. Vet. Microbiol. 2016, 193, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Makeyev, E.V.; Kang, Z.; Ji, S.; Bamford, D.H.; van Dijk, A.A. Cloning and sequence analysis of dsRNA segments 5, 6 and 7 of a novel non-group A, B, C adult rotavirus that caused an outbreak of gastroenteritis in China. Virus Res. 2004, 106, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Kobayashi, N.; Ishino, M.; Ahmed, M.S.; Ahmed, M.U.; Paul, S.K.; Muzumdar, B.K.; Hussain, Z.; Wang, Y.H.; Naik, T.N. Genetic analysis of an ADRV-N-like novel rotavirus strain B219 detected in a sporadic case of adult diarrhea in Bangladesh. Arch. Virol. 2007, 152, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Ji, S.; Tang, Q.; Cui, X.; Yang, H.; Kan, B.; Gao, S. Molecular characterization of a novel adult diarrhoea rotavirus strain J19 isolated in China and its significance for the evolution and origin of group B rotaviruses. J. Gen. Virol. 2008, 89 Pt 10, 2622–2629. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Kobayashi, N.; Ishino, M.; Alam, M.M.; Ahmed, M.U.; Paul, S.K.; Ganesh, B.; Chawla-Sarkar, M.; Krishnan, T.; Naik, T.N.; et al. Whole genomic characterization of a human rotavirus strain B219 belonging to a novel group of the genus Rotavirus. J. Med. Virol. 2008, 80, 2023–2033. [Google Scholar] [CrossRef] [PubMed]
- Nyaga, M.M.; Peenze, I.; Potgieter, C.A.; Seheri, L.M.; Page, N.A.; Yinda, C.K.; Steele, A.D.; Matthijnssens, J.; Mphahlele, M.J. Complete genome analyses of the first porcine rotavirus group H identified from a South African pig does not provide evidence for recent interspecies transmission events. Infect. Genet. Evol. 2016, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Winiarczyk, S.; Paul, P.S.; Mummidi, S.; Panek, R.; Gradzki, Z. Survey of porcine rotavirus G and P genotype in Poland and the United States using RT-PCR. J. Vet. Med. B Infect. Dis. Vet. Public Health 2002, 49, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; Vlasova, A.N.; Saif, L.J. Prevalence and genetic heterogeneity of porcine group C rotaviruses in nursing and weaned piglets in Ohio, USA and identification of a potential new VP4 genotype. Vet. Microbiol. 2013, 164, 27–38. [Google Scholar] [PubMed]
- Tonietti, P.O.; Hora, A.S.; Silva, F.D.; Ruiz, V.L.; Gregori, F. Phylogenetic analyses of the VP4 and VP7 genes of porcine group A rotaviruses in Sao Paulo State, Brazil: First identification of G5P[23] in piglets. J. Clin. Microbiol. 2013, 51, 2750–2753. [Google Scholar] [CrossRef] [PubMed]
- Nyaga, M.M.; Jere, K.C.; Esona, M.D.; Seheri, M.L.; Stucker, K.M.; Halpin, R.A.; Akopov, A.; Stockwell, T.B.; Peenze, I.; Diop, A.; et al. Whole genome detection of rotavirus mixed infections in human, porcine and bovine samples co-infected with various rotavirus strains collected from sub-Saharan Africa. Infect. Genet. Evol. 2015, 31, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Atii, D.J.; Ojeh, C.K. Subgroup determination of group A rotaviruses recovered from piglets in Nigeria. Viral Immunol. 1995, 8, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Geyer, A.; Sebata, T.; Peenze, I.; Steele, A.D. Group B and C porcine rotaviruses identified for the first time in South Africa. J. S. Afr. Vet. Assoc. 1996, 67, 115–116. [Google Scholar] [PubMed]
- Malik, Y.S.; Kumar, N.; Sharma, K.; Sircar, S.; Dhama, K.; Bora, D.P.; Dutta, T.; Prasad, M.; Tiwari, A.K. Rotavirus diarrhea in piglets: A review on epidemiology, genetic diversity and zoonotic risks. Indian J. Anim. Sci. 2014, 84, 1035–1042. [Google Scholar]
- Gachanja, E.; Buza, J.; Petrucka, P. Prevalence of group A rotavirus in piglets in a periurban setting of Arusha, Tanzania. J. Biosci. Med. 2016, 4, 37–44. [Google Scholar]
- Otto, P.H.; Rosenhain, S.; Elschner, M.C.; Hotzel, H.; Machnowska, P.; Trojnar, E.; Hoffmann, K.; Johne, R. Detection of rotavirus species A, B and C in domestic mammalian animals with diarrhoea and genotyping of bovine species A rotavirus strains. Vet. Microbiol. 2015, 179, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Pratelli, A.; Greco, G.; Tempesta, M.; Ferrari, M.; Losio, M.N.; Buonavoglia, C. Genomic characterization of porcine rotaviruses in Italy. Clin. Diagn. Lab. Immunol. 2001, 8, 129–132. [Google Scholar] [CrossRef]
- Theuns, S.; Desmarets, L.M.; Heylen, E.; Zeller, M.; Dedeurwaerder, A.; Roukaerts, I.D.; Van Ranst, M.; Matthijnssens, J.; Nauwynck, H.J. Porcine group A rotaviruses with heterogeneous VP7 and VP4 genotype combinations can be found together with enteric bacteria on Belgian swine farms. Vet. Microbiol. 2014, 172, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Ciarlet, M.; Banyai, K.; Lorusso, E.; Cavalli, A.; Corrente, M.; Elia, G.; Arista, S.; Camero, M.; Desario, C.; et al. Identification of a novel VP4 genotype carried by a serotype G5 porcine rotavirus strain. Virology 2006, 346, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Ciarlet, M.; Baselga, R.; Arista, S.; Elia, G.; Lorusso, E.; Banyai, K.; Terio, V.; Madio, A.; Ruggeri, F.M.; et al. Sequence analysis of the VP7 and VP4 genes identifies a novel VP7 gene allele of porcine rotaviruses, sharing a common evolutionary origin with human G2 rotaviruses. Virology 2005, 337, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Van der Heide, R.; Koopmans, M.P.; Shekary, N.; Houwers, D.J.; van Duynhoven, Y.T.; van der Poel, W.H. Molecular characterizations of human and animal group a rotaviruses in the Netherlands. J. Clin. Microbiol. 2005, 43, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Chandler-Bostock, R.; Hancox, L.R.; Nawaz, S.; Watts, O.; Iturriza-Gomara, M.; Mellits, K.H. Genetic diversity of porcine group A rotavirus strains in the UK. Vet. Microbiol. 2014, 173, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Smitalova, R.; Rodak, L.; Smid, B.; Psikal, I. Detection of nongroup A rotaviruses in faecal samples of pigs in the Czech Republic. Vet. Med. 2009, 54, 1–18. [Google Scholar]
- Chitambar, S.D.; Arora, R.; Chhabra, P. Molecular characterization of a rare G1P[19] rotavirus strain from India: evidence of reassortment between human and porcine rotavirus strains. J. Med. Microbiol. 2009, 58 Pt 12, 1611–1615. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.A.; Khamrin, P.; Trinh, Q.D.; Phan, T.G.; Pham le, D.; Hoang le, P.; Hoang, K.T.; Yagyu, F.; Okitsu, S.; Ushijima, H. Sequence analysis of Vietnamese P[6] rotavirus strains suggests evidence of interspecies transmission. J. Med. Virol. 2007, 79, 1959–1965. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.J.; Li, D.D.; Zhang, Q.; Liu, N.; Huang, C.P.; Jiang, X.; Jiang, B.; Glass, R.; Steele, D.; Tang, J.Y.; et al. Novel human rotavirus of genotype G5P[6] identified in a stool specimen from a Chinese girl with diarrhea. J. Clin. Microbiol. 2007, 45, 1614–1617. [Google Scholar] [PubMed]
- Matsushima, Y.; Nakajima, E.; Nguyen, T.A.; Shimizu, H.; Kano, A.; Ishimaru, Y.; Phan, T.G.; Ushijima, H. Genome sequence of an unusual human G10P[8] rotavirus detected in Vietnam. J. Virol. 2012, 86, 10236–10237. [Google Scholar] [CrossRef] [PubMed]
- Park, S.I.; Matthijnssens, J.; Saif, L.J.; Kim, H.J.; Park, J.G.; Alfajaro, M.M.; Kim, D.S.; Son, K.Y.; Yang, D.K.; Hyun, B.H.; et al. Reassortment among bovine, porcine and human rotavirus strains results in G8P[7] and G6P[7] strains isolated from cattle in South Korea. Vet. Microbiol. 2011, 152, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Teodoroff, T.A.; Tsunemitsu, H.; Okamoto, K.; Katsuda, K.; Kohmoto, M.; Kawashima, K.; Nakagomi, T.; Nakagomi, O. Predominance of porcine rotavirus G9 in Japanese piglets with diarrhea: close relationship of their VP7 genes with those of recent human G9 strains. J. Clin. Microbiol. 2005, 43, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Khamrin, P.; Maneekarn, N.; Peerakome, S.; Chan-it, W.; Yagyu, F.; Okitsu, S.; Ushijima, H. Novel porcine rotavirus of genotype P[27] shares new phylogenetic lineage with G2 porcine rotavirus strain. Virology 2007, 361, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, J.; Li, H.; Sun, D.; Wang, C.; Feng, L. Molecular characterization of a rare G9P[23] porcine rotavirus isolate from China. Arch. Virol. 2012, 157, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Z.; Wang, Y.; Wang, X.; Xia, M.; Wu, H. Isolation, molecular characterization and evaluation of the pathogenicity of a porcine rotavirus isolated from Jiangsu Province, China. Arch. Virol. 2015, 160, 1333–1338. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Li, D.D.; Cai, K.; Qin, J.J.; Wang, Y.X.; Lin, Q.; Guo, Y.Q.; Zhao, C.Y.; Duan, Z.J. The epidemiological characteristics of group C rotavirus in Lulong area and the analysis of diversity of VP6 gene. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 2013, 27, 164–166. [Google Scholar] [PubMed]
- Suzuki, T.; Soma, J.; Miyazaki, A.; Tsunemitsu, H. Phylogenetic analysis of nonstructural protein 5 (NSP5) gene sequences in porcine rotavirus B strains. Infect. Genet. Evol. 2012, 12, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Lahon, A.; Ingle, V.C.; Birade, H.S.; Raut, C.G.; Chitambar, S.D. Molecular characterization of group B rotavirus circulating in pigs from India: identification of a strain bearing a novel VP7 genotype, G21. Vet. Microbiol. 2014, 174, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Nagesha, H.S.; Dyall-Smith, M.L.; Holmes, I.H. Comparative sequence analysis of VP7 genes from five Australian porcine rotaviruses. Arch. Virol. 1989, 109, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.A.; Nagesha, H.S.; Holmes, I.H. Comparative sequence analysis of VP4s from five Australian porcine rotaviruses: implication of an apparent new P type. Virology 1993, 196, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Nagesha, H.S.; Huang, J.; Holmes, I.H. A variant serotype G3 rotavirus isolated from an unusually severe outbreak of diarrhoea in piglets. J. Med. Virol. 1992, 38, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Khamrin, P.; Maneekarn, N.; Peerakome, S.; Yagyu, F.; Okitsu, S.; Ushijima, H. Molecular characterization of a rare G3P[3] human rotavirus reassortant strain reveals evidence for multiple human-animal interspecies transmissions. J. Med. Virol. 2006, 78, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Marton, S.; Doro, R.; Feher, E.; Forro, B.; Ihasz, K.; Varga-Kugler, R.; Farkas, S.L.; Banyai, K. Whole genome sequencing of a rare rotavirus from archived stool sample demonstrates independent zoonotic origin of human G8P[14] strains in Hungary. Virus Res. 2017, 227, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Lin, X.D.; Huang, K.Y.; Zhang, B.; Shi, M.; Guo, W.P.; Wang, M.R.; Wang, W.; Xing, J.G.; Li, M.H.; et al. Identification of novel and diverse rotaviruses in rodents and insectivores, and evidence of cross-species transmission into humans. Virology 2016, 494, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Varghese, V.; Samajdar, S.; Bhattacharya, S.K.; Kobayashi, N.; Naik, T.N. Molecular characterization of a porcine Group A rotavirus strain with G12 genotype specificity. Arch. Virol. 2006, 151, 1329–1344. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, Y.; Honma, S.; Jones, R.W.; Ross, J.; Santos, N.; Gentsch, J.R.; Kapikian, A.Z.; Hesse, R.A. A porcine G9 rotavirus strain shares neutralization and VP7 phylogenetic sequence lineage 3 characteristics with contemporary human G9 rotavirus strains. Virology 2005, 332, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Matthijnssens, J.; Yang, X.; Delbeke, T.; Arijs, I.; Taniguchi, K.; Iturriza-Gomara, M.; Iftekharuddin, N.; Azim, T.; Van Ranst, M. Evolutionary history and global spread of the emerging g12 human rotaviruses. J. Virol. 2007, 81, 2382–2390. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Mullick, S.; Deb, A.K.; Panda, S.; Chawla-Sarkar, M. First report of human rotavirus G8P[4] gastroenteritis in India: evidence of ruminants-to-human zoonotic transmission. J. Med. Virol. 2013, 85, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Doan, Y.H.; Nakagomi, T.; Aboudy, Y.; Silberstein, I.; Behar-Novat, E.; Nakagomi, O.; Shulman, L.M. Identification by full-genome analysis of a bovine rotavirus transmitted directly to and causing diarrhea in a human child. J. Clin. Microbiol. 2013, 51, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Luchs, A.; Cilli, A.; Morillo, S.G.; Carmona Rde, C.; Timenetsky Mdo, C. Rare G3P[3] rotavirus strain detected in Brazil: possible human-canine interspecies transmission. J. Clin. Virol. 2012, 54, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Ben Hadj Fredj, M.; Heylen, E.; Zeller, M.; Fodha, I.; Benhamida-Rebai, M.; Van Ranst, M.; Matthijnssens, J.; Trabelsi, A. Feline origin of rotavirus strain, Tunisia, 2008. Emerg. Infect. Dis. 2013, 19, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, P.; Tan, M.; Liu, Y.; Biesiada, J.; Meller, J.; Castello, A.A.; Jiang, B.; Jiang, X. Rotavirus VP8*: phylogeny, host range, and interaction with histo-blood group antigens. J. Virol. 2012, 86, 9899–9910. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ramelot, T.A.; Huang, P.; Liu, Y.; Li, Z.; Feizi, T.; Zhong, W.; Wu, F.T.; Tan, M.; Kennedy, M.A.; et al. Glycan Specificity of P[19] Rotavirus and Comparison with Those of Related P Genotypes. J. Virol. 2016, 90, 9983–9996. [Google Scholar] [CrossRef]
- Huang, P.; Xia, M.; Tan, M.; Zhong, W.; Wei, C.; Wang, L.; Morrow, A.; Jiang, X. Spike protein VP8* of human rotavirus recognizes histo-blood group antigens in a type-specific manner. J. Virol. 2012, 86, 4833–4843. [Google Scholar] [CrossRef] [PubMed]
- Van Trang, N.; Vu, H.T.; Le, N.T.; Huang, P.; Jiang, X.; Anh, D.D. Association between norovirus and rotavirus infection and histo-blood group antigen types in Vietnamese children. J. Clin. Microbiol. 2014, 52, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, F.; Yamamoto, M. Molecular genetic basis of porcine histo-blood group AO system. Blood 2001, 97, 3308–3310. [Google Scholar] [CrossRef] [PubMed]
- Cooling, L. Blood Groups in Infection and Host Susceptibility. Clin. Microbiol. Rev. 2015, 28, 801–870. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Banyai, K.; Ciarlet, M.; Iturriza-Gomara, M.; Lorusso, E.; De Grazia, S.; Arista, S.; Decaro, N.; Elia, G.; Cavalli, A.; et al. Relationships among porcine and human P[6] rotaviruses: evidence that the different human P[6] lineages have originated from multiple interspecies transmission events. Virology 2006, 344, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, J.D.; Leite, J.P.; Lima, J.C.; Heinemann, M.B.; Oliveira, D.S.; Araujo, I.T.; Soares, L.S.; Gusmao, R.H.; Gabbay, Y.B.; Linhares, A.C. Detection of a neonatal human rotavirus strain with VP4 and NSP4 genes of porcine origin. J. Med. Microbiol. 2007, 56 Pt 4, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Shintani, T.; Ghosh, S.; Wang, Y.H.; Zhou, X.; Zhou, D.J.; Kobayashi, N. Whole genomic analysis of human G1P[8] rotavirus strains from different age groups in China. Viruses 2012, 4, 1289–1304. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, R.G.; James, W.D.; Bohl, E.H.; Theil, K.W.; Saif, L.J.; Kalica, A.R.; Greenberg, H.B.; Kapikian, A.Z.; Chanock, R.M. Human rotavirus type 2: Cultivation in vitro. Science 1980, 207, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Do, L.P.; Nakagomi, T.; Otaki, H.; Agbemabiese, C.A.; Nakagomi, O.; Tsunemitsu, H. Phylogenetic inference of the porcine Rotavirus A origin of the human G1 VP7 gene. Infect. Genet. Evol. 2016, 40, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Do, L.P.; Nakagomi, T.; Nakagomi, O. A rare G1P[6] super-short human rotavirus strain carrying an H2 genotype on the genetic background of a porcine rotavirus. Infect. Genet. Evol. 2014, 21, 334–350. [Google Scholar] [CrossRef] [PubMed]
- Heylen, E.; Batoko Likele, B.; Zeller, M.; Stevens, S.; De Coster, S.; Conceicao-Neto, N.; Van Geet, C.; Jacobs, J.; Ngbonda, D.; Van Ranst, M.; et al. Rotavirus surveillance in Kisangani, the Democratic Republic of the Congo, reveals a high number of unusual genotypes and gene segments of animal origin in non-vaccinated symptomatic children. PLoS ONE 2014, 9, e100953. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Y.H.; Ghosh, S.; Tang, W.F.; Pang, B.B.; Liu, M.Q.; Peng, J.S.; Zhou, D.J.; Kobayashi, N. Genomic characterization of G3P[6], G4P[6] and G4P[8] human rotaviruses from Wuhan, China: Evidence for interspecies transmission and reassortment events. Infect. Genet. Evol. 2015, 33, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Martella, V.; Colombrita, D.; Lorusso, E.; Draghin, E.; Fiorentini, S.; De Grazia, S.; Banyai, K.; Ciarlet, M.; Caruso, A.; Buonavoglia, C. Detection of a porcine-like rotavirus in a child with enteritis in Italy. J. Clin. Microbiol. 2008, 46, 3501–3507. [Google Scholar] [CrossRef] [PubMed]
- Steyer, A.; Poljsak-Prijatelj, M.; Barlic-Maganja, D.; Marin, J. Human, porcine and bovine rotaviruses in Slovenia: evidence of interspecies transmission and genome reassortment. J. Gen. Virol. 2008, 89 Pt 7, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.T.; Banyai, K.; Huang, J.C.; Wu, H.S.; Chang, F.Y.; Hsiung, C.A.; Huang, Y.C.; Lin, J.S.; Hwang, K.P.; Jiang, B.; et al. Human infection with novel G3P[25] rotavirus strain in Taiwan. Clin. Microbiol. Infec. 2011, 17, 1570–1573. [Google Scholar] [CrossRef] [PubMed]
- Hwang, K.P.; Wu, F.T.; Banyai, K.; Wu, H.S.; Yang, D.C.; Huang, Y.C.; Lin, J.S.; Hsiung, C.A.; Huang, J.C.; Jiang, B.; et al. Identification of porcine rotavirus-like genotype P[6] strains in Taiwanese children. J. Med. Microbiol. 2012, 61 Pt 7, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Papp, H.; Borzak, R.; Farkas, S.; Kisfali, P.; Lengyel, G.; Molnar, P.; Melegh, B.; Matthijnssens, J.; Jakab, F.; Martella, V.; et al. Zoonotic transmission of reassortant porcine G4P[6] rotaviruses in Hungarian pediatric patients identified sporadically over a 15 year period. Infect. Genet. Evol. 2013, 19, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.J.; Qian, Y.; Huang, T.; Zhu, R.N.; Zhao, L.Q.; Zhang, Y.; Li, R.C.; Li, Y.P. Identification of circulating porcine-human reassortant G4P[6] rotavirus from children with acute diarrhea in China by whole genome analyses. Infect. Genet. Evol. 2013, 20, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Degiuseppe, J.I.; Beltramino, J.C.; Millan, A.; Stupka, J.A.; Parra, G.I. Complete genome analyses of G4P[6] rotavirus detected in Argentinean children with diarrhoea provides evidence of interspecies transmission from swine. Clin. Microbiol. Infec. 2013, 19, E367–E371. [Google Scholar] [CrossRef]
- Stupka, J.A.; Carvalho, P.; Amarilla, A.A.; Massana, M.; Parra, G.I.; Argentinean National Surveillance Network for Diarrheas. National Rotavirus Surveillance in Argentina: High incidence of G9P[8] strains and detection of G4P[6] strains with porcine characteristics. Infect. Genet. Evol. 2009, 9, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Razafindratsimandresy, R.; Heraud, J.M.; Ramarokoto, C.E.; Rabemanantsoa, S.; Randremanana, R.; Andriamamonjy, N.S.; Richard, V.; Reynes, J.M. Rotavirus genotypes in children in the community with diarrhea in Madagascar. J. Med. Virol. 2013, 85, 1652–1660. [Google Scholar] [CrossRef] [PubMed]
- Komoto, S.; Maeno, Y.; Tomita, M.; Matsuoka, T.; Ohfu, M.; Yodoshi, T.; Akeda, H.; Taniguchi, K. Whole genomic analysis of a porcine-like human G5P[6] rotavirus strain isolated from a child with diarrhoea and encephalopathy in Japan. J. Gen. Virol. 2013, 94 Pt 7, 1568–1575. [Google Scholar] [PubMed]
- Mladenova, Z.; Papp, H.; Lengyel, G.; Kisfali, P.; Steyer, A.; Steyer, A.F.; Esona, M.D.; Iturriza-Gomara, M.; Banyai, K. Detection of rare reassortant G5P[6] rotavirus, Bulgaria. Infect. Genet. Evol. 2012, 12, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.F.; Tort, L.F.; Gomez, M.M.; Assis, R.M.; Volotao Ede, M.; de Mendonca, M.C.; Bello, G.; Leite, J.P. VP7 Gene of human rotavirus A genotype G5: Phylogenetic analysis reveals the existence of three different lineages worldwide. J. Med. Virol. 2011, 83, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic-Rustempasic, S.; Banyai, K.; Esona, M.D.; Foytich, K.; Bowen, M.D.; Gentsch, J.R. Genome sequence based molecular epidemiology of unusual US Rotavirus A G9 strains isolated from Omaha, USA between 1997 and 2000. Infect. Genet. Evol. 2011, 11, 522–527. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Laso, J.; Roman, A.; Head, J.; Cervera, I.; Rodriguez, M.; Rodriguez-Avial, I.; Picazo, J.J. Phylogeny of G9 rotavirus genotype: a possible explanation of its origin and evolution. J. Clin. Virol. 2009, 44, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Dutta, D.; Ghosh, S.; Bagchi, P.; Chattopadhyay, S.; Nagashima, S.; Kobayashi, N.; Dutta, P.; Krishnan, T.; Naik, T.N.; et al. Full genomic analysis of a human group A rotavirus G9P[6] strain from Eastern India provides evidence for porcine-to-human interspecies transmission. Arch. Virol. 2009, 154, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Yodmeeklin, A.; Khamrin, P.; Chuchaona, W.; Kumthip, K.; Kongkaew, A.; Vachirachewin, R.; Okitsu, S.; Ushijima, H.; Maneekarn, N. Analysis of complete genome sequences of G9P[19] rotavirus strains from human and piglet with diarrhea provides evidence for whole-genome interspecies transmission of nonreassorted porcine rotavirus. Infect. Genet. Evol. 2017, 47, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Urushibara, N.; Taniguchi, K.; Kobayashi, N. Whole genomic analysis reveals the porcine origin of human G9P[19] rotavirus strains Mc323 and Mc345. Infect. Genet. Evol. 2012, 12, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.T.; Banyai, K.; Jiang, B.; Liu, L.T.; Marton, S.; Huang, Y.C.; Huang, L.M.; Liao, M.H.; Hsiung, C.A. Novel G9 rotavirus strains co-circulate in children and pigs, Taiwan. Sci. Rep. 2017, 7, 40731. [Google Scholar] [CrossRef] [PubMed]
- Do, L.P.; Kaneko, M.; Nakagomi, T.; Gauchan, P.; Agbemabiese, C.A.; Dang, A.D.; Nakagomi, O. Molecular epidemiology of Rotavirus A, causing acute gastroenteritis hospitalizations among children in Nha Trang, Vietnam, 2007–2008: Identification of rare G9P[19] and G10P[14] strains. J. Med. Virol. 2017, 89, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Rahman, M.; Ciarlet, M.; Zeller, M.; Heylen, E.; Nakagomi, T.; Uchida, R.; Hassan, Z.; Azim, T.; Nakagomi, O.; et al. Reassortment of human rotavirus gene segments into G11 rotavirus strains. Emerg. Infect. Dis. 2010, 16, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.A.; Mathur, M.; Deshpande, J.M. Complete genome analysis of a rare group A rotavirus, G11P[25], isolated from a child in Mumbai, India, reveals interspecies transmission and reassortment with human rotavirus strains. J. Med. Microbiol. 2014, 63 Pt 9, 1220–1227. [Google Scholar] [CrossRef] [PubMed]
- Mullick, S.; Mukherjee, A.; Ghosh, S.; Pazhani, G.P.; Sur, D.; Manna, B.; Nataro, J.P.; Levine, M.M.; Ramamurthy, T.; Chawla-Sarkar, M. Genomic analysis of human rotavirus strains G6P[14] and G11P[25] isolated from Kolkata in 2009 reveals interspecies transmission and complex reassortment events. Infect. Genet. Evol. 2013, 14, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Komoto, S.; Wandera Apondi, E.; Shah, M.; Odoyo, E.; Nyangao, J.; Tomita, M.; Wakuda, M.; Maeno, Y.; Shirato, H.; Tsuji, T.; et al. Whole genomic analysis of human G12P[6] and G12P[8] rotavirus strains that have emerged in Kenya: identification of porcine-like NSP4 genes. Infect. Genet. Evol. 2014, 27, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Ide, T.; Komoto, S.; Higo-Moriguchi, K.; Htun, K.W.; Myint, Y.Y.; Myat, T.W.; Thant, K.Z.; Thu, H.M.; Win, M.M.; Oo, H.N.; et al. Whole Genomic Analysis of Human G12P[6] and G12P[8] Rotavirus Strains that Have Emerged in Myanmar. PLoS ONE 2015, 10, e0124965. [Google Scholar] [CrossRef] [PubMed]
- My, P.V.; Rabaa, M.A.; Donato, C.; Cowley, D.; Phat, V.V.; Dung, T.T.; Anh, P.H.; Vinh, H.; Bryant, J.E.; Kellam, P.; et al. Novel porcine-like human G26P[19] rotavirus identified in hospitalized paediatric diarrhoea patients in Ho Chi Minh City, Vietnam. J. Gen. Virol. 2014, 95 Pt 12, 2727–2733. [Google Scholar] [CrossRef] [PubMed]
- Medici, K.C.; Barry, A.F.; Alfieri, A.F.; Alfieri, A.A. Genetic analysis of the porcine group B rotavirus NSP2 gene from wild-type Brazilian strains. Braz. J. Med. Biol. Res. 2010, 43, 13–16. [Google Scholar] [CrossRef] [PubMed]
- Gabbay, Y.B.; Borges, A.A.; Oliveira, D.S.; Linhares, A.C.; Mascarenhas, J.D.; Barardi, C.R.; Simoes, C.M.; Wang, Y.; Glass, R.I.; Jiang, B. Evidence for zoonotic transmission of group C rotaviruses among children in Belem, Brazil. J. Med. Virol. 2008, 80, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Iturriza-Gomara, M.; Dallman, T.; Banyai, K.; Bottiger, B.; Buesa, J.; Diedrich, S.; Fiore, L.; Johansen, K.; Koopmans, M.; Korsun, N.; et al. Rotavirus genotypes co-circulating in Europe between 2006 and 2009 as determined by EuroRotaNet, a pan-European collaborative strain surveillance network. Epidemiol. Infect. 2011, 139, 895–909. [Google Scholar] [CrossRef] [PubMed]
- Iturriza-Gomara, M.; Dallman, T.; Banyai, K.; Bottiger, B.; Buesa, J.; Diedrich, S.; Fiore, L.; Johansen, K.; Korsun, N.; Kroneman, A.; et al. Rotavirus surveillance in europe, 2005–2008: web-enabled reporting and real-time analysis of genotyping and epidemiological data. J. Infect. Dis. 2009, 200 (Suppl. S1), S215–S221. [Google Scholar] [CrossRef] [PubMed]
- Iturriza-Gomara, M.; Green, J.; Brown, D.W.; Ramsay, M.; Desselberger, U.; Gray, J.J. Molecular epidemiology of human group A rotavirus infections in the United Kingdom between 1995 and 1998. J. Clin. Microbiol. 2000, 38, 4394–4401. [Google Scholar] [PubMed]
- Chang, K.O.; Nielsen, P.R.; Ward, L.A.; Saif, L.J. Dual infection of gnotobiotic calves with bovine strains of group A and porcine-like group C rotaviruses influences pathogenesis of the group C rotavirus. J. Virol. 1999, 73, 9284–9293. [Google Scholar] [PubMed]
- Banyai, K.; Jiang, B.; Bogdan, A.; Horvath, B.; Jakab, F.; Meleg, E.; Martella, V.; Magyari, L.; Melegh, B.; Szucs, G. Prevalence and molecular characterization of human group C rotaviruses in Hungary. J. Clin. Virol. 2006, 37, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Desselberger, U.; Huppertz, H.I. Immune responses to rotavirus infection and vaccination and associated correlates of protection. J. Infect. Dis. 2011, 203, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Ward, L.A.; Yuan, L.; Rosen, B.I.; To, T.L. The gnotobiotic piglet as a model for studies of disease pathogenesis and immunity to human rotaviruses. Arch. Virol. Suppl. 1996, 12, 153–161. [Google Scholar] [PubMed]
- Vlasova, A.N.; Shao, L.; Kandasamy, S.; Fischer, D.D.; Rauf, A.; Langel, S.N.; Chattha, K.S.; Kumar, A.; Huang, H.C.; Rajashekara, G.; et al. Escherichia coli Nissle 1917 protects gnotobiotic pigs against human rotavirus by modulating pDC and NK-cell responses. Eur. J. Immunol. 2016, 46, 2426–2437. [Google Scholar] [CrossRef] [PubMed]
- Narvaez, C.F.; Angel, J.; Franco, M.A. Interaction of rotavirus with human myeloid dendritic cells. J. Virol. 2005, 79, 14526–14535. [Google Scholar] [CrossRef] [PubMed]
- Feng, N.; Kim, B.; Fenaux, M.; Nguyen, H.; Vo, P.; Omary, M.B.; Greenberg, H.B. Role of interferon in homologous and heterologous rotavirus infection in the intestines and extraintestinal organs of suckling mice. J. Virol. 2008, 82, 7578–7590. [Google Scholar] [CrossRef] [PubMed]
- Angel, J.; Franco, M.A.; Greenberg, H.B.; Bass, D. Lack of a role for type I and type II interferons in the resolution of rotavirus-induced diarrhea and infection in mice. J. Interferon Cytokine Res. 1999, 19, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Vancott, J.L.; McNeal, M.M.; Choi, A.H.; Ward, R.L. The role of interferons in rotavirus infections and protection. J. Interferon Cytokine Res. 2003, 23, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Chattha, K.S.; Kandasamy, S.; Siegismund, C.S.; Saif, L.J. Prenatally acquired vitamin A deficiency alters innate immune responses to human rotavirus in a gnotobiotic pig model. J. Immunol. 2013, 190, 4742–4753. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Chattha, K.S.; Kandasamy, S.; Liu, Z.; Esseili, M.; Shao, L.; Rajashekara, G.; Saif, L.J. Lactobacilli and bifidobacteria promote immune homeostasis by modulating innate immune responses to human rotavirus in neonatal gnotobiotic pigs. PLoS ONE 2013, 8, e76962. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, S.; Vlasova, A.N.; Fischer, D.; Kumar, A.; Chattha, K.S.; Rauf, A.; Shao, L.; Langel, S.N.; Rajashekara, G.; Saif, L.J. Differential Effects of Escherichia coli Nissle and Lactobacillus rhamnosus Strain GG on Human Rotavirus Binding, Infection, and B Cell Immunity. J. Immunol. 2016, 196, 1780–1789. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.S.; Yuan, L.; Iosef, C.; Chang, K.O.; Kim, Y.; Nguyen, T.V.; Saif, L.J. Magnitude of serum and intestinal antibody responses induced by sequential replicating and nonreplicating rotavirus vaccines in gnotobiotic pigs and correlation with protection. Clin. Diagn. Lab. Immunol. 2004, 11, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Kang, S.Y.; Ward, L.A.; To, T.L.; Saif, L.J. Antibody-secreting cell responses and protective immunity assessed in gnotobiotic pigs inoculated orally or intramuscularly with inactivated human rotavirus. J. Virol. 1998, 72, 330–338. [Google Scholar] [PubMed]
- Yuan, L.; Wen, K.; Azevedo, M.S.; Gonzalez, A.M.; Zhang, W.; Saif, L.J. Virus-specific intestinal IFN-gamma producing T cell responses induced by human rotavirus infection and vaccines are correlated with protection against rotavirus diarrhea in gnotobiotic pigs. Vaccine 2008, 26, 3322–3331. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Iosef, C.; Azevedo, M.S.; Kim, Y.; Qian, Y.; Geyer, A.; Nguyen, T.V.; Chang, K.O.; Saif, L.J. Protective immunity and antibody-secreting cell responses elicited by combined oral attenuated Wa human rotavirus and intranasal Wa 2/6-VLPs with mutant Escherichia coli heat-labile toxin in gnotobiotic pigs. J. Virol. 2001, 75, 9229–9238. [Google Scholar] [CrossRef] [PubMed]
- Iosef, C.; Chang, K.O.; Azevedo, M.S.; Saif, L.J. Systemic and intestinal antibody responses to NSP4 enterotoxin of Wa human rotavirus in a gnotobiotic pig model of human rotavirus disease. J. Med. Virol. 2002, 68, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Hodgins, D.C.; Kang, S.Y.; deArriba, L.; Parreno, V.; Ward, L.A.; Yuan, L.; To, T.; Saif, L.J. Effects of maternal antibodies on protection and development of antibody responses to human rotavirus in gnotobiotic pigs. J. Virol. 1999, 73, 186–197. [Google Scholar] [PubMed]
- Nguyen, T.V.; Yuan, L.; Azevedo, M.S.; Jeong, K.I.; Gonzalez, A.M.; Iosef, C.; Lovgren-Bengtsson, K.; Morein, B.; Lewis, P.; Saif, L.J. High titers of circulating maternal antibodies suppress effector and memory B-cell responses induced by an attenuated rotavirus priming and rotavirus-like particle-immunostimulating complex boosting vaccine regimen. Clin. Vaccine Immunol. CVI 2006, 13, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.E.; Parashar, U.D. Rotavirus vaccines in routine use. Clin. Infect. Dis. 2014, 59, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Fernandez, F.M. Group A rotavirus veterinary vaccines. J. Infect. Dis. 1996, 174 (Suppl. S1), S98–S106. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.P.; Vlasova, A.N.; Saif, L.J. Human rotavirus virus-like particle vaccines evaluated in a neonatal gnotobiotic pig model of human rotavirus disease. Expert Rev. Vaccines 2013, 12, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Mijatovic-Rustempasic, S.; Esona, M.D.; Tam, K.I.; Quaye, O.; Bowen, M.D. One-step multiplex real-time RT-PCR assay for detecting and genotyping wild-type group A rotavirus strains and vaccine strains (Rotarix(R) and RotaTeq(R)) in stool samples. PeerJ 2016, 4, e1560. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Porcine RV Species | G and/or P Genotype | Geographic Region | Year Samples Collected | Epidemiological Status and Medical Relevance | Reference |
|---|---|---|---|---|---|
| A | G1-G4, G9, G12 and P8 | Worldwide | 2000s | Commonly seen in humans * | [152,153,165] |
| G3–G5, G9 and G11, as well as P[6] | Denmark, France, Hungary, Italy, Slovenia | 2003–2007 | G3-G5—common in humans, G5—regional in humans, P6—rare in humans | [85] | |
| G1 and G4 | Brazil | 2007 | Common | [166] | |
| G1P[8] | China; MD, USA | 2004–2009 | Common | [15,167,168] | |
| G1, G1P[6] | Japan | 2001 | Common | [169] | |
| G1P[6] | Japan | 1997 | Rare | [170] | |
| G1P[6], G4P[6] and G12P[6] | Democratic Republic of the Congo | 2007–2010 | Common | [171] | |
| G1P[19] | India | 1992 | Rare | [134] | |
| G2 | Europe | 1992 | Uncommon | [130] | |
| G3P[6], G4P[6] and G4P[8] | China, Italy, Slovenia | 2003–2013 | Common | [172,173,174] | |
| G3P[25] | Taiwan | 2009 | Rare | [175] | |
| G4P[6] strains, one G5P[6] | Taiwan | 2006–2012 | Common | [176] | |
| G4P[6] | Hungary, China, Argentina, Madagascar | 2006–2007 2008–2009 | Sporadic identification in humans worldwide | [177,178,179,180,181] | |
| G5P[6] | Japan, Bulgaria | 2011 2006 | Rare | [182,183] | |
| G5P[8] | Brazil, Argentina, Paraguay, Cameroon, China, Thailand, and Vietnam | 1986–2005 | Common in Asian, African and South American countries | [184] | |
| G9 | NE, USA; India | 1980s, 1190s, 1997–2000 | Uncommon, emerging worldwide | [153,185,186] | |
| G9P[6] | India | 2007 | Unusual | [187] | |
| G9P[19] | Thailand, India | 2012–2013 1989–1990 | Rare | [188,189] | |
| G9P[19] and G9P[13] | Taiwan | 2014–2015 | Rare | [190] | |
| G9P[19] and G10P[14] | Vietnam | 2007–2008 | Rare | [191] | |
| G11P[4], G11P[6], G11P[8] G11P[25] | Nepal, Bangladesh | 2001–2004 | Rare | [192] | |
| G11P[25] | India | 2005–2009 | Uncommon | [193,194] | |
| G12P[6] and G12P[8] | Kenya, Myanmar | 2010, 2011 | Common | [195,196] | |
| G26P[19] | Vietnam | 2009–2010 | Atypical in humans | [197] | |
| B | N/A | Brazil | 2000s | Regional significance | [198] |
| C | N/A | Japan, Brazil | 1982–1986, 2000–2007 | Regional significance | [100,199] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vlasova, A.N.; Amimo, J.O.; Saif, L.J. Porcine Rotaviruses: Epidemiology, Immune Responses and Control Strategies. Viruses 2017, 9, 48. https://doi.org/10.3390/v9030048
Vlasova AN, Amimo JO, Saif LJ. Porcine Rotaviruses: Epidemiology, Immune Responses and Control Strategies. Viruses. 2017; 9(3):48. https://doi.org/10.3390/v9030048
Chicago/Turabian StyleVlasova, Anastasia N., Joshua O. Amimo, and Linda J. Saif. 2017. "Porcine Rotaviruses: Epidemiology, Immune Responses and Control Strategies" Viruses 9, no. 3: 48. https://doi.org/10.3390/v9030048