
The Envelope Gene of Hepatitis B Virus Is Implicated in Both Differential Virion Secretion and Genome Replication Capacities between Genotype B and Genotype C Isolates

Abstract
:1. Introduction
2. Materials and Methods
2.1. The 1.1mer over-Length HBV Construct and SphI Dimer for Genome Replication
2.2. DNA Fragment Exchange and Site-Directed Mutagenesis
2.3. Subgenomic Expression Constructs for ENVELOPE Proteins
2.4. Transient Transfection and Measurement of Secreted HBsAg
2.5. Detection of HBV DNA Replication and Virion Secretion
3. Results
3.1. Rationale for Using 1.1mer Construct to Compare Virion Secretion between Genotypes B and C
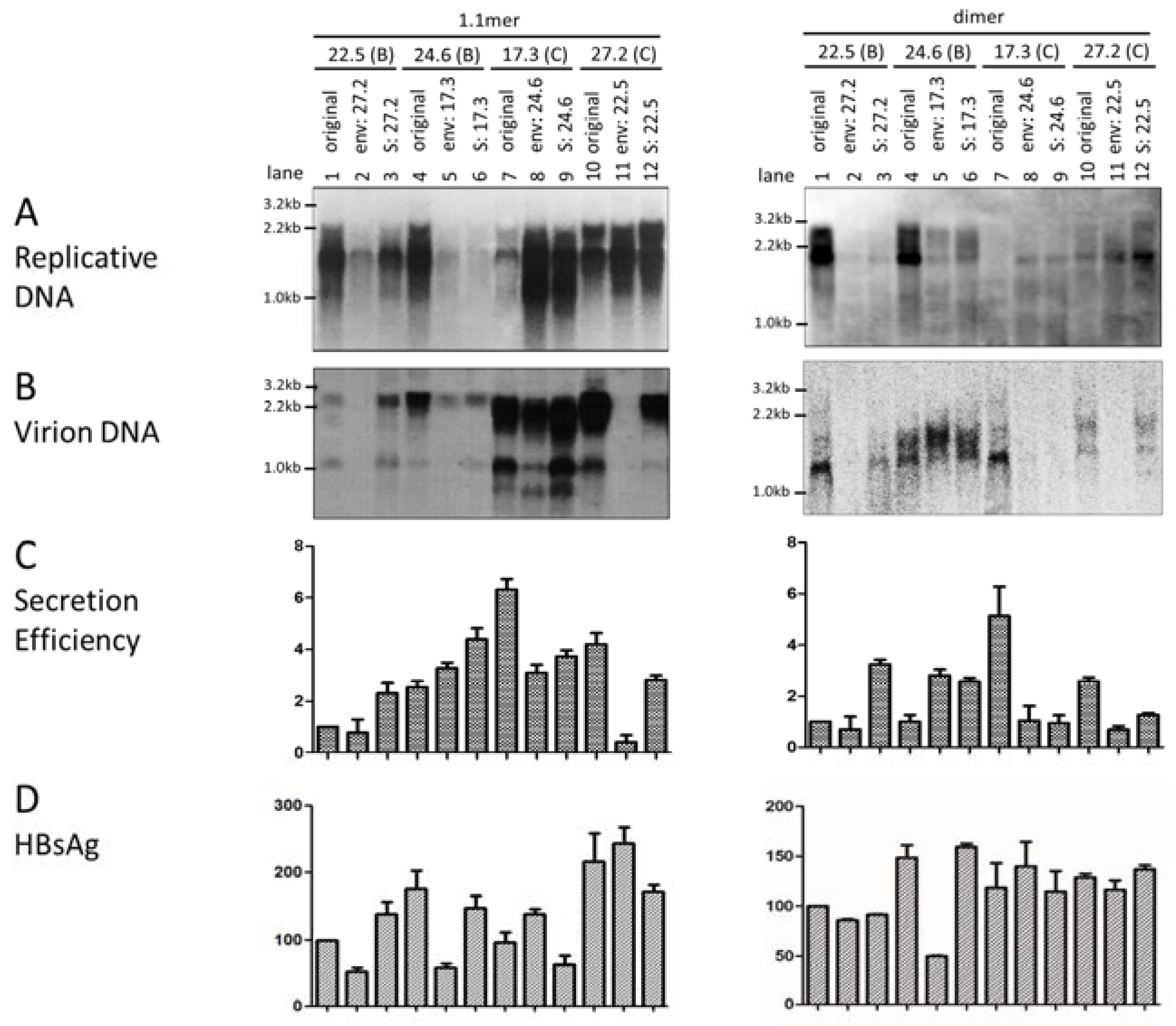
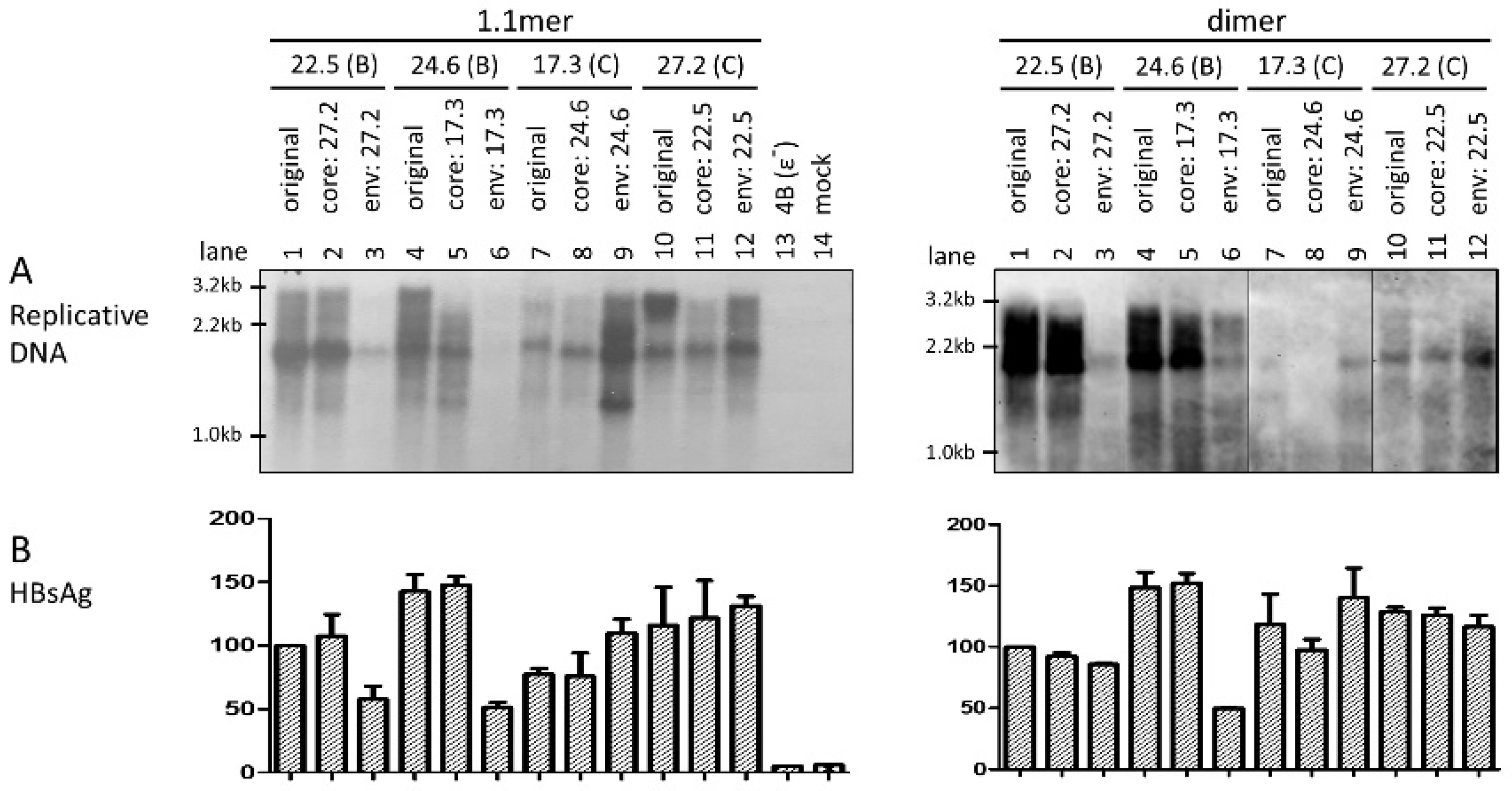
3.2. Swapping the Envelope Gene Could Reverse the Replication Phenotype between Clones of the Two Genotypes
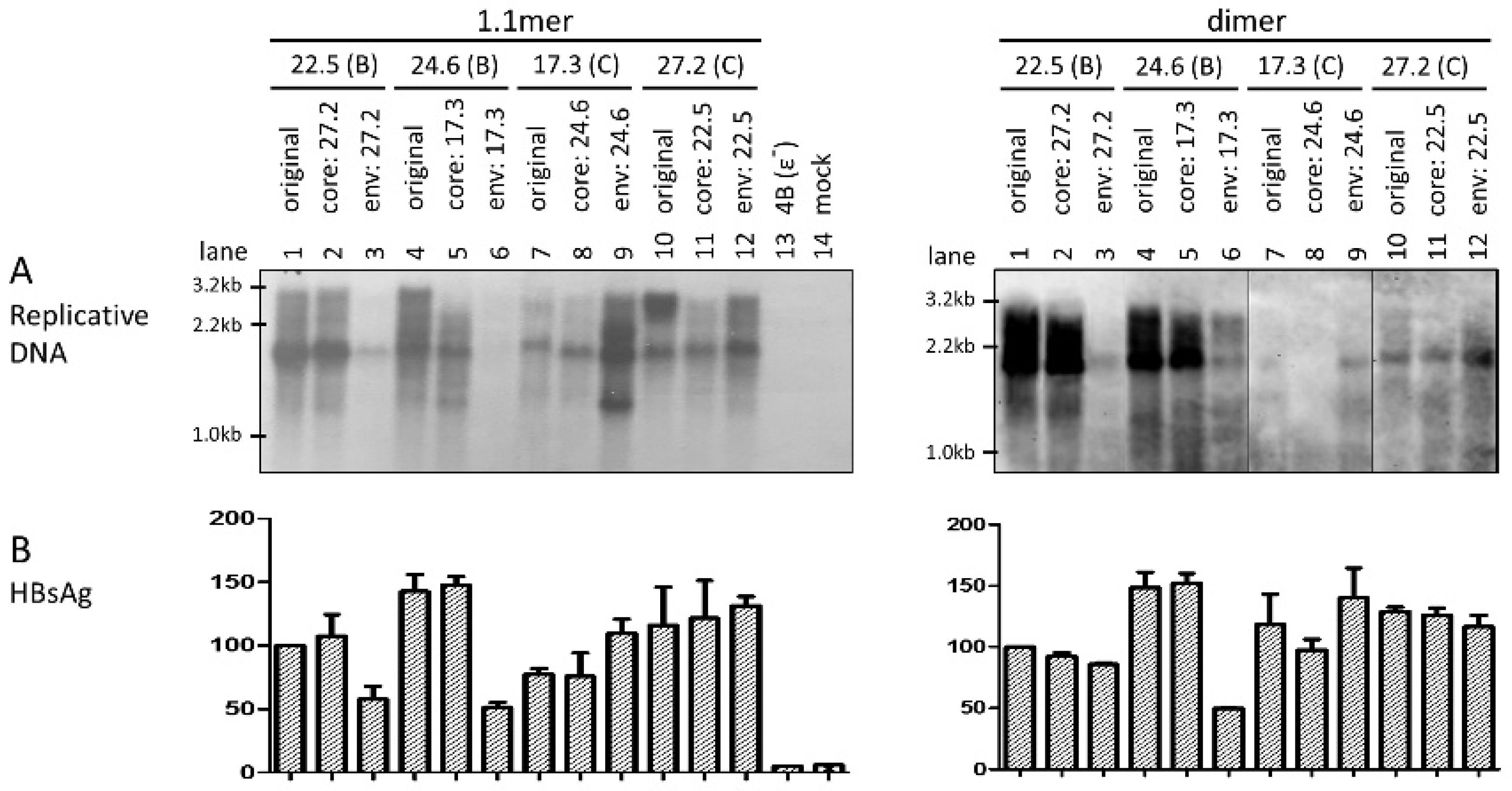
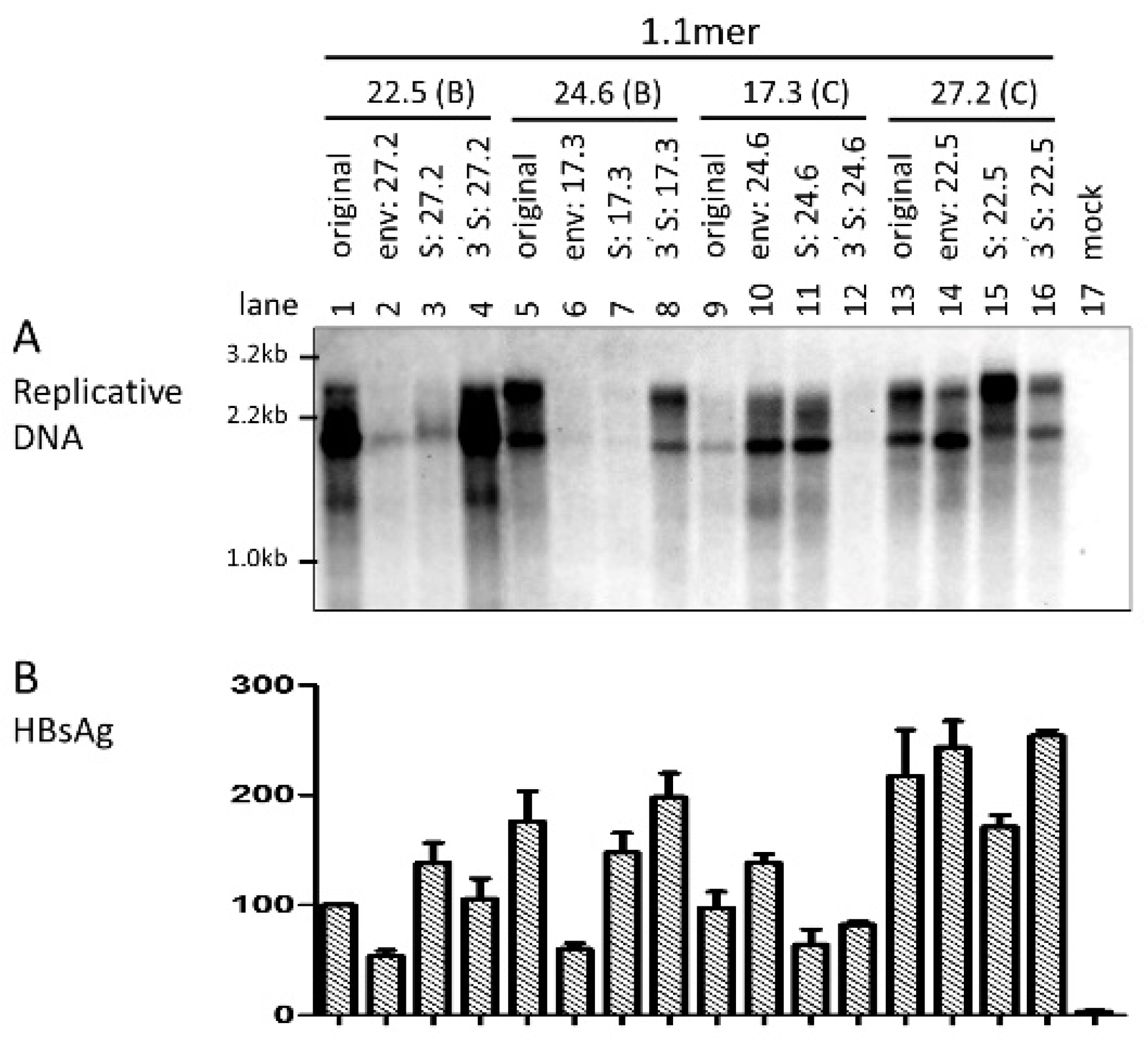
3.3. Level of Replicative DNA Was Unaffected by Exchanging the Core Gene or 3′ S Region between the Two HBV Genotypes
3.4. Exchanging the Entire Envelope Gene Also Affected HBsAg Level in Culture Supernatant, Especially for 1.1mer Construct
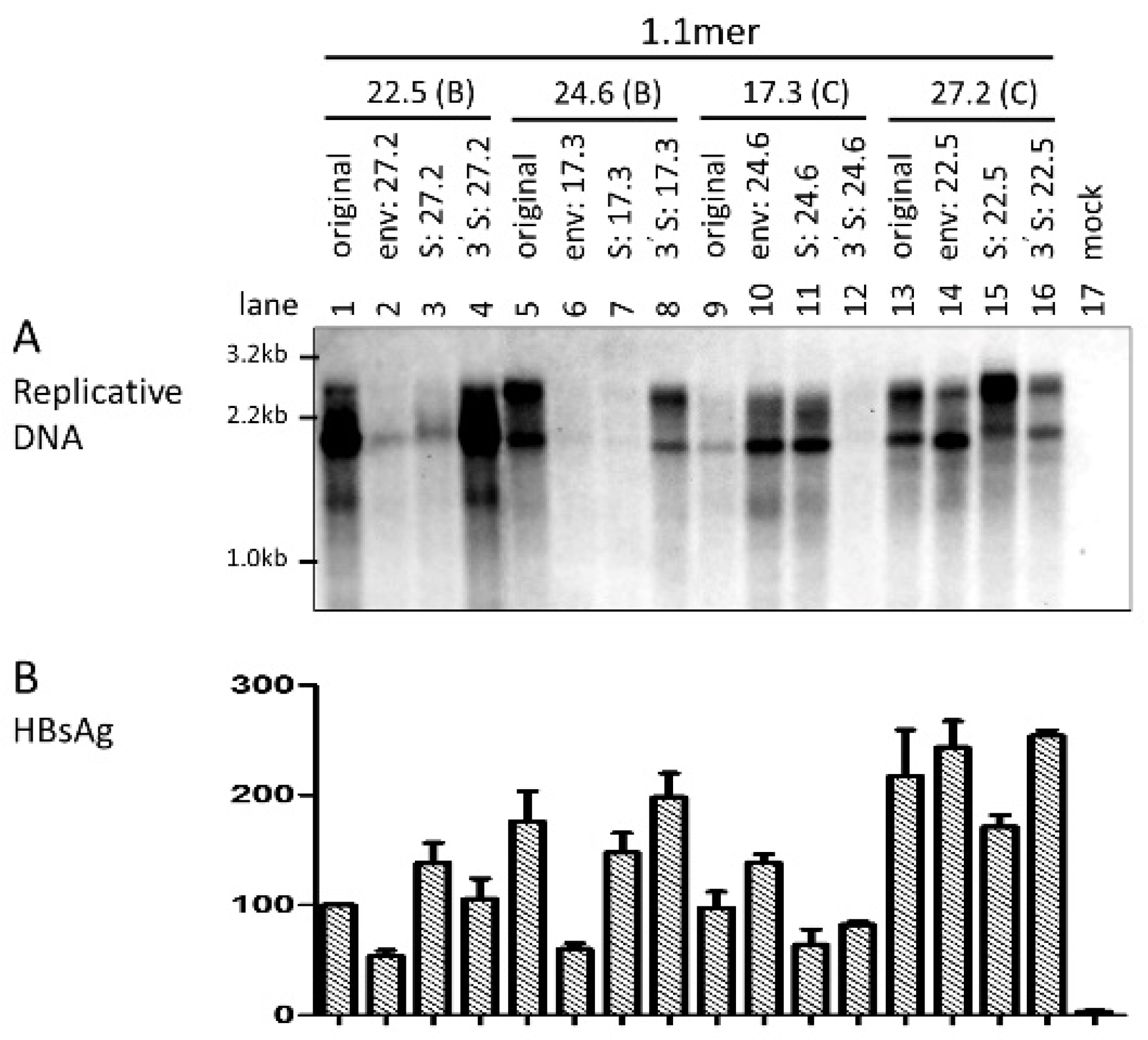
3.5. The Envelope Gene of Genotype C Was Associated with Higher Efficiency of Virion Secretion
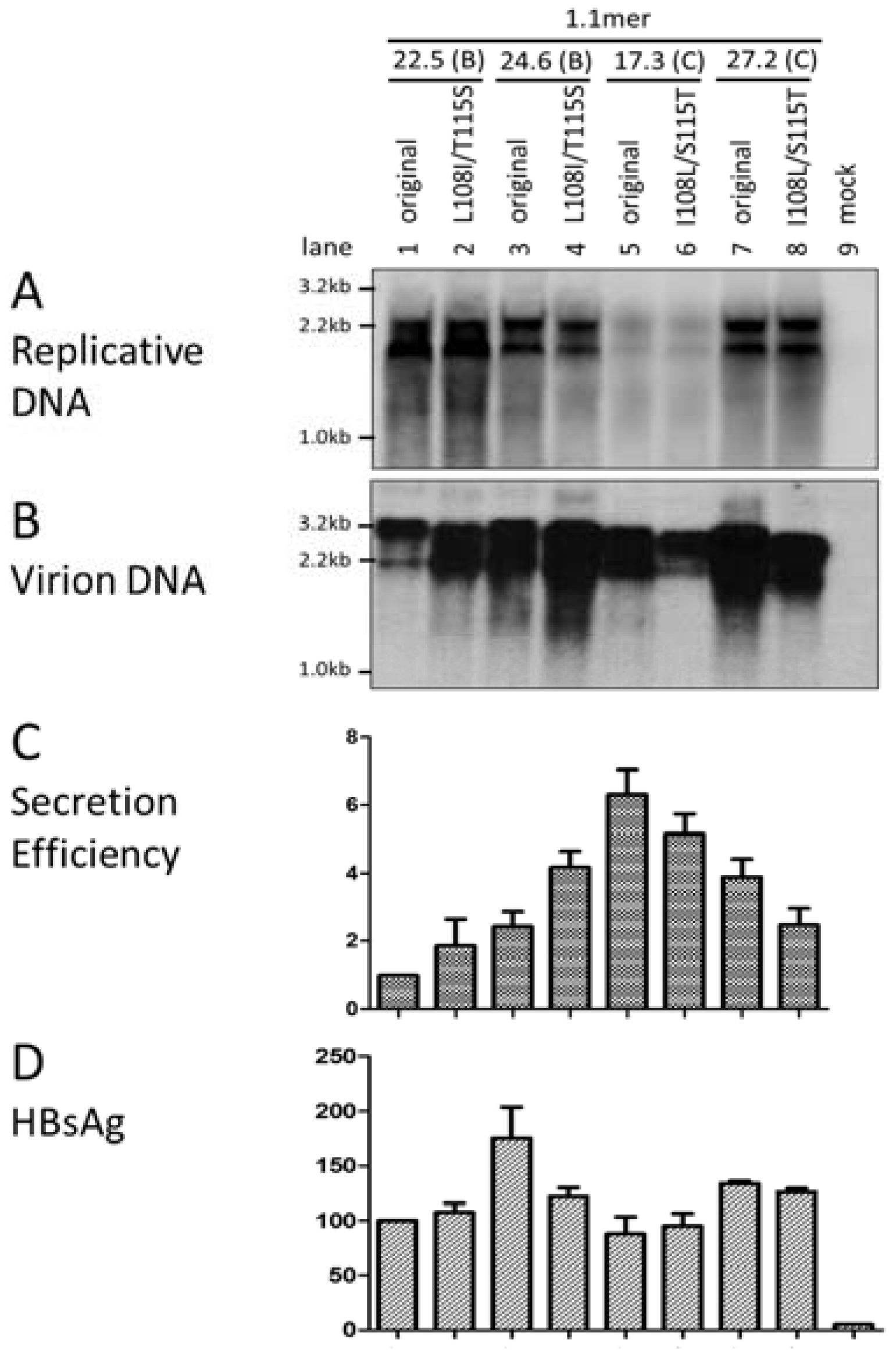
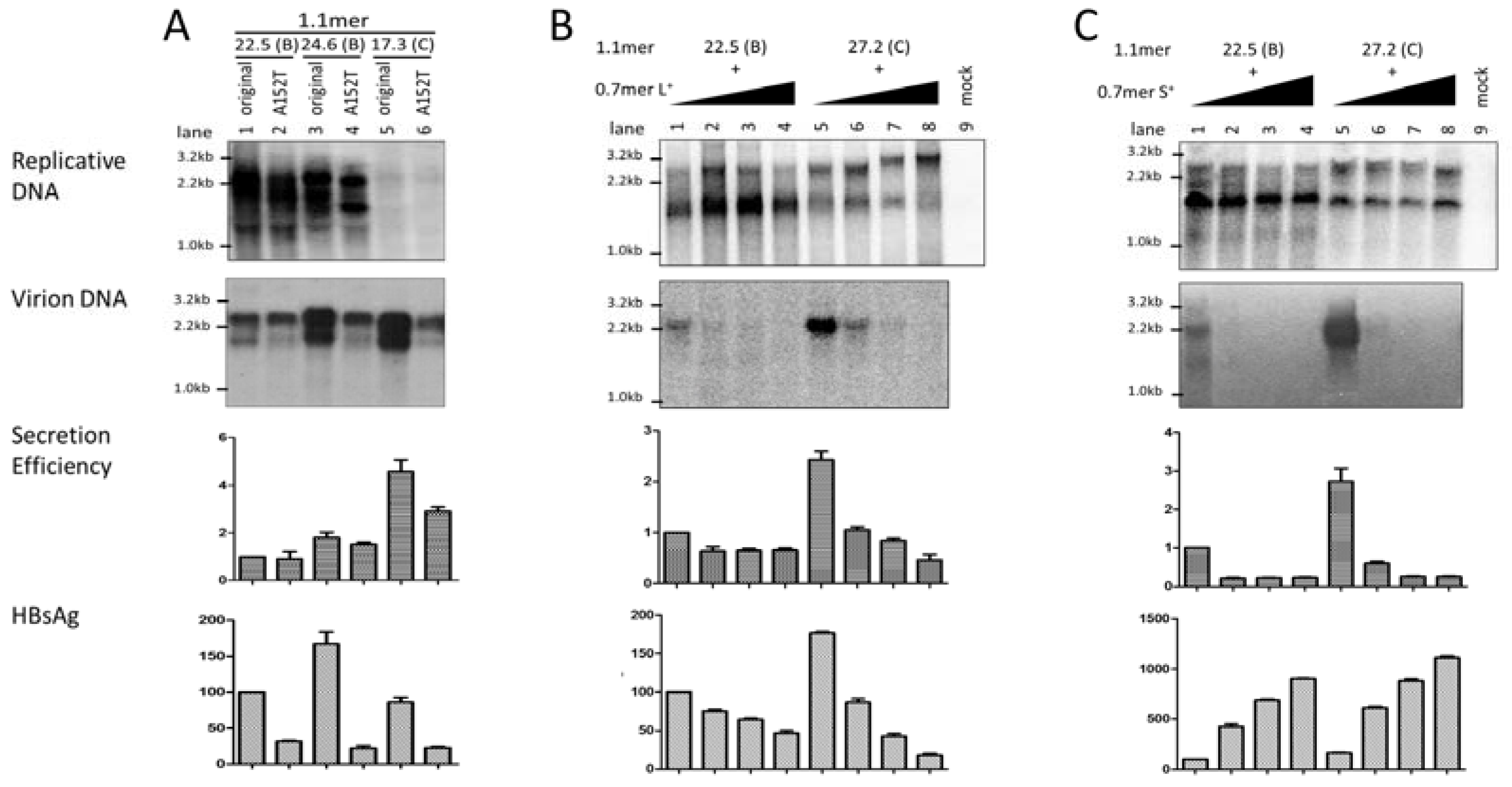
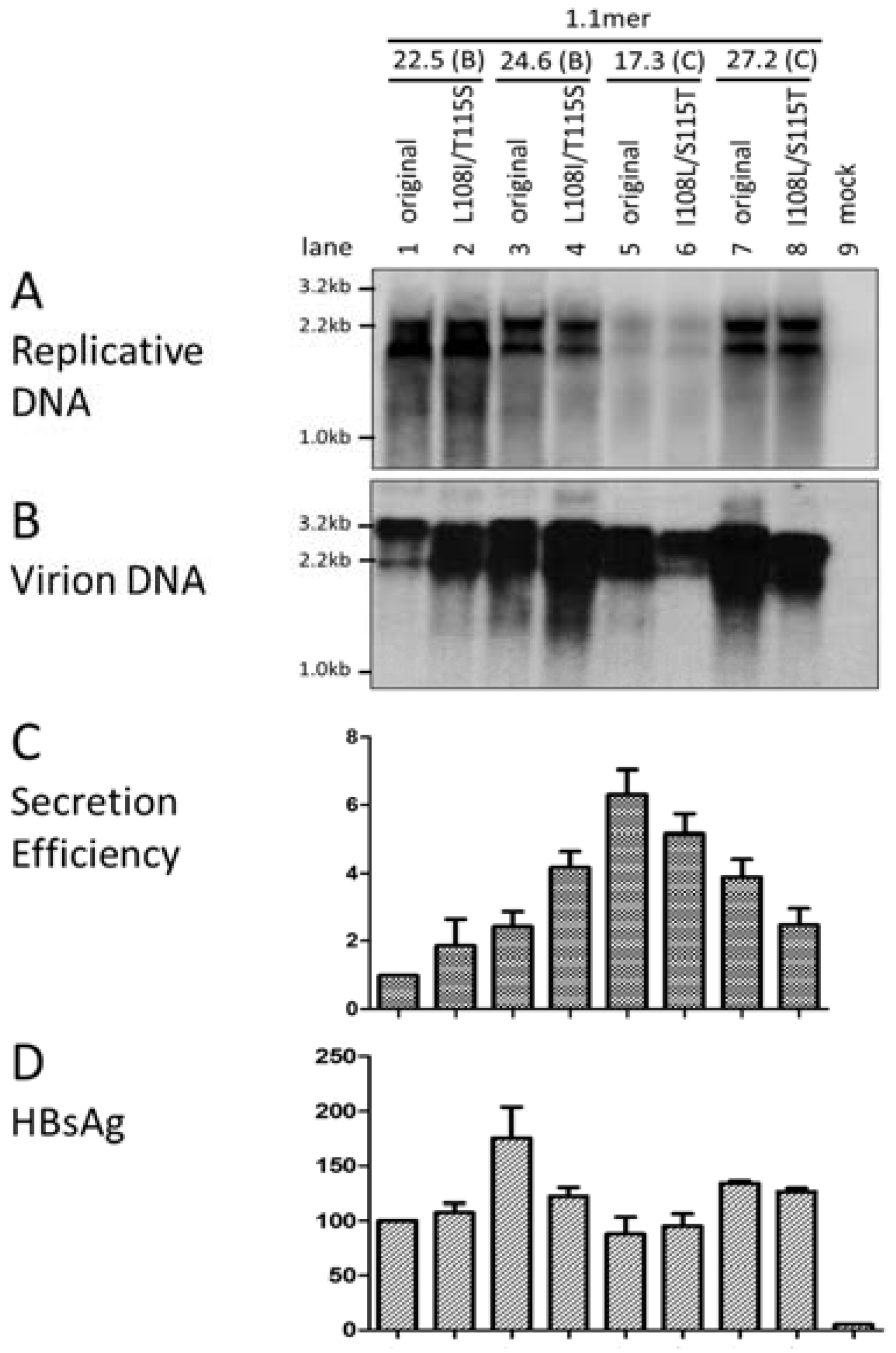
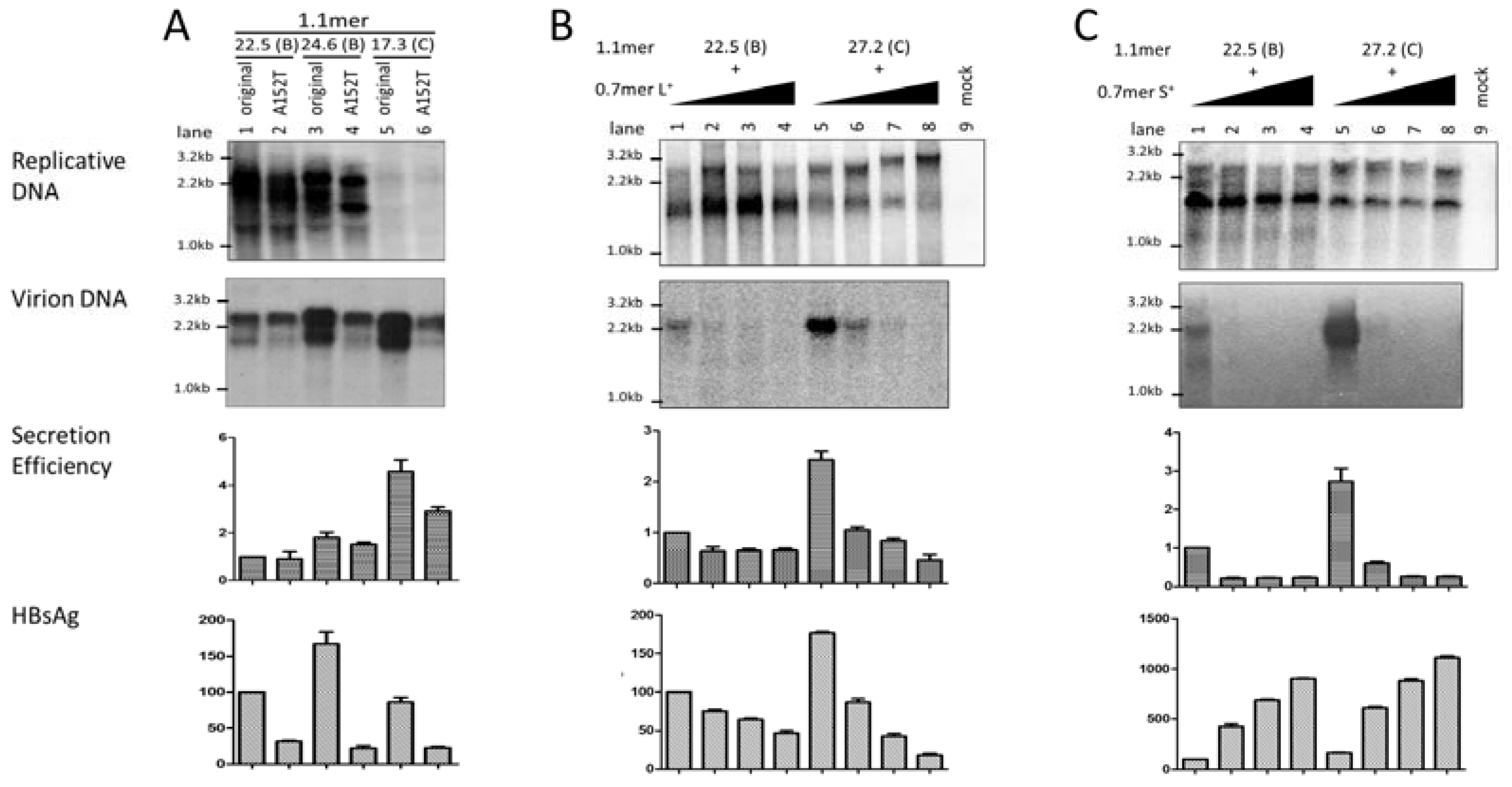
3.6. Two Residues in the preS1 Domain Contributed to Genotypic Difference in Virion Secretion
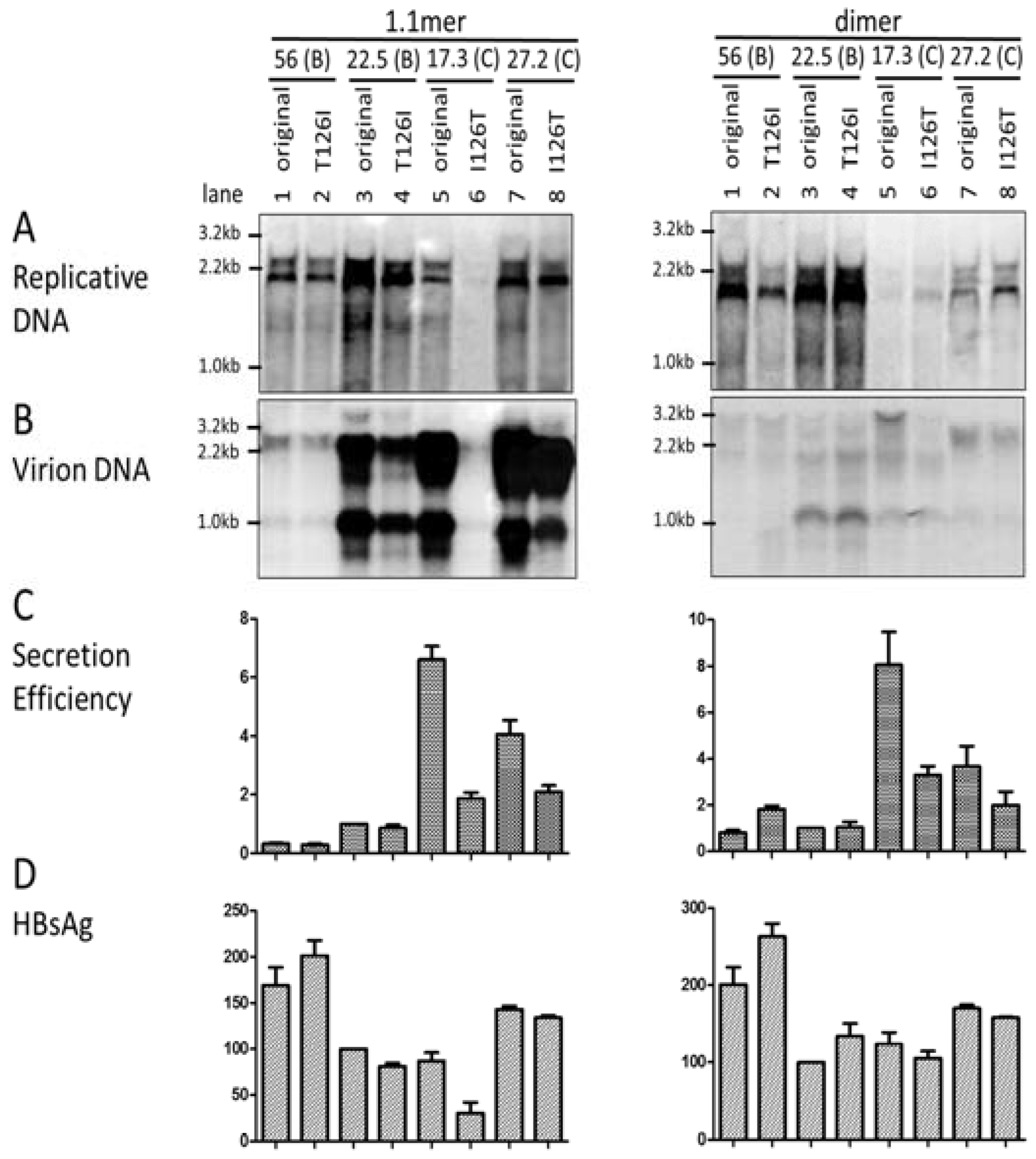
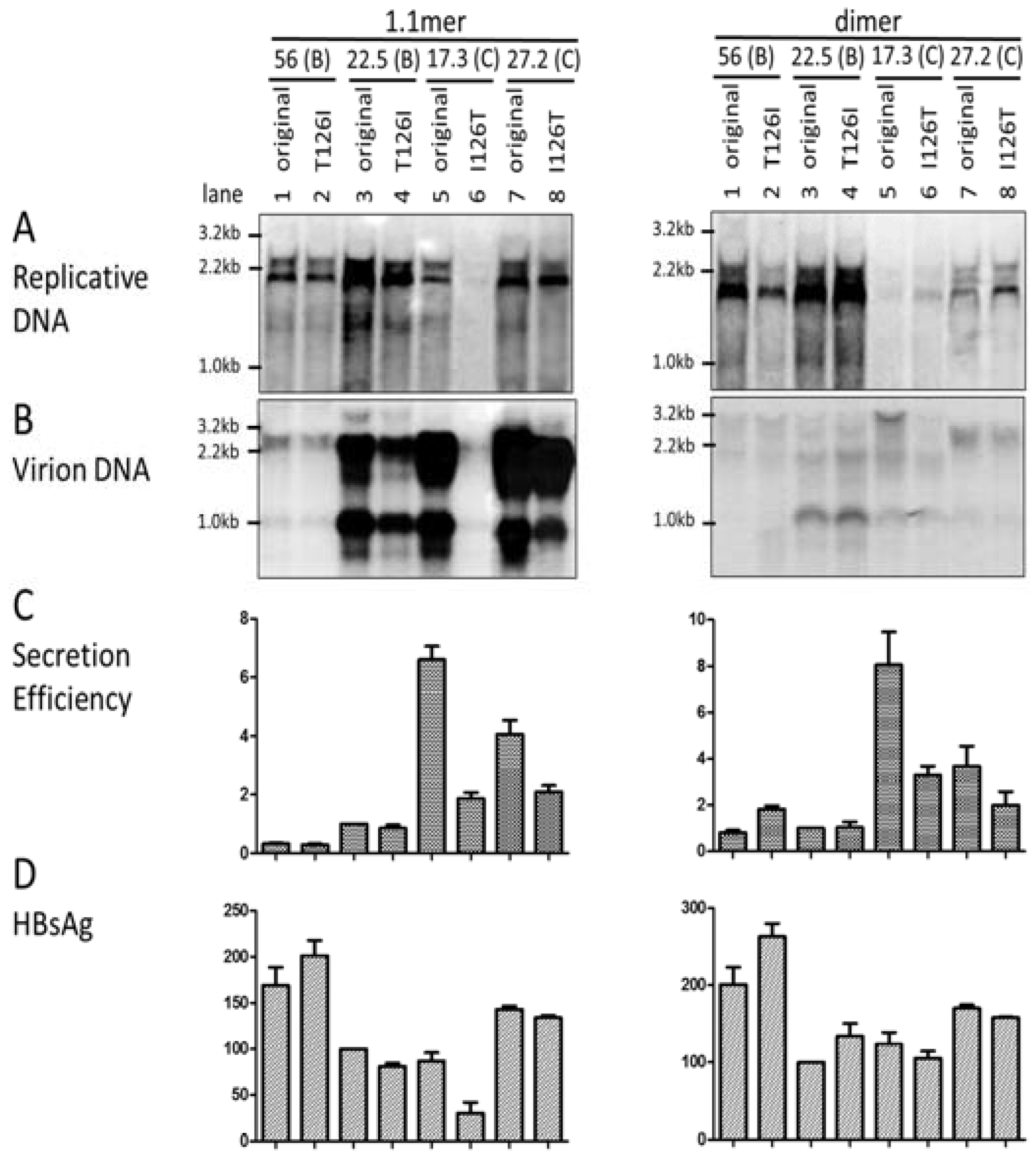
3.7. I126 in the S Domain Was Partly Responsible for Efficient Virion Secretion by Genotype C
3.8. The L/S Protein Ratio Produced by the 1.1mer Construct Appeared Optimal for Virion Secretion
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ganem, D.; Prince, A.M. Hepatitis B virus infection--natural history and clinical consequences. N. Engl. J. Med. 2004, 350, 1118–1129. [Google Scholar] [CrossRef] [PubMed]
- Trépo, C.; Chan, H.L.Y.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- Norder, H.; Courouce, A.M.; Coursaget, P.; Echevarria, J.M.; Lee, S.D.; Mushahwar, I.K.; Robertson, B.H.; Locarnini, S.; Magnius, L.O. Genetic diversity of hepatitis B virus strains derived worldwide: Genotypes, subgenotypes, and HBsAg subtypes. Intervirology 2004, 47, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, S. Hepatitis B virus: Significance of genotypes. J. Viral Hepat. 2005, 12, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A. Genotypes and genetic variability of hepatitis B virus. Intervirology 2013, 57, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Revill, P. Overview of hepatitis B viral replication and genetic variability. J. Hepatol. 2016, 64, S4–S16. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.F.; Tanaka, Y.; Shinkai, N.; Poon, R.T.; But, Y.K.; Fong, Y.T.; Fung, J.; Wong, K.H.; Yuen, C.H.; Mizokami, M. Risk for hepatocellular carcinoma with respect to hepatitis B virus genotypes B/C, specific mutations of enhancer II/core promoter/precore regions and HBV DNA levels. Gut 2008, 57, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Yuen, M.F.; Sablon, E.; Yuan, H.J.; Wong, K.H.; Hui, C.K.; Wong, C.Y.; Chan, O.O.; Lai, C.L. Significance of hepatitis B genotype in acute exacerbation, HBeAg seroconversion, cirrhosis-related complications, and hepatocellular carcinoma. Hepatology 2003, 37, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Orito, E.; Ichida, T.; Sakugawa, H.; Sata, M.; Horiike, N.; Hino, K.; Okita, K.; Okanoue, T.; Iino, S.; Tanaka, E. Geographic distribution of hepatitis B virus (HBV) genotype in patients with chronic HBV infection in Japan. Hepatology 2001, 34, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.J.; Hussain, M.; Lok, A.S. Hepatitis B virus genotype B is associated with earlier HBeAg seroconversion compared with hepatitis B virus genotype c. Gastroenterology 2002, 122, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.L.; Hui, A.Y.; Wong, M.L.; Tse, A.M.; Hung, L.C.; Wong, V.W.; Sung, J.J. Genotype c hepatitis B virus infection is associated with an increased risk of hepatocellular carcinoma. Gut 2005, 53, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.M.; Liaw, Y.F. Genotype C hepatitis B virus infection is associated with a higher risk of reactivation of hepatitis B and progression to cirrhosis than genotype b: A longitudinal study of hepatitis B e antigen-positive patients with normal aminotransferase levels at base. J. Hepatol. 2005, 43, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Yin, J.H.; Li, Y.T.; Li, C.Z.; Ren, H.; Gu, C.Y.; Wu, H.Y.; Liang, X.S.; Zhang, P.; Zhao, J.F. Risk factors for acute hepatitis B and its progression to chronic hepatitis in Shanghai, China. Gut 2008, 57, 1713–1720. [Google Scholar] [CrossRef] [PubMed]
- Imamura, T. Distribution of hepatitis B viral genotypes and mutations in the core promoter and precore regions in acute forms of liver disease in patients from Chiba, Japan. Gut 2003, 52, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Ozasa, A.; Tanaka, Y.; Orito, E.; Sugiyama, M.; Kang, J.H.; Hige, S.; Kuramitsu, T.; Suzuki, K.; Tanaka, E.; Okada, S. Influence of genotypes and precore mutations on fulminant or chronic outcome of acute hepatitis B virus infection. Hepatology 2006, 44, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Xu, Z.; Liu, Y.; Li, X.; Bai, S.; Ding, N.; Zhong, Y.; Wang, L.; Mao, P.; Zoulim, F. Hepatitis B virus genotype and basal core promoter/precore mutations are associated with hepatitis B-related acute-on-chronic liver failure without pre-existing liver cirrhosis. J. Viral Hepat. 2010, 17, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Tang, X.; Garcia, T.; Hussain, M.; Zhang, J.; Lok, A.; Wands, J.; Li, J.; Tong, S. Hepatitis B virus genotype c isolates with wild-type core promoter sequence replicate less efficiently than genotype B isolates but possess higher virion secretion capacity. J. Virol. 2011, 85, 10167–10177. [Google Scholar] [CrossRef] [PubMed]
- Seeger, C.; Mason, W.S. Molecular biology of hepatitis b virus infection. Virology 2015, 479–480, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Boyer, M. Hepatitis B virus reverse transcriptase and epsilon RNA sequences required for specific interaction in vitro. J. Virol. 2006, 80, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Tavis, J.E.; Ganem, D. Evidence for activation of the hepatitis B virus polymerase by binding of its RNA template. J. Virol. 1996, 70, 5741–5750. [Google Scholar] [PubMed]
- Bruss, V.; Ganem, D. The role of envelope proteins in hepatitis B virus assembly. Proc. Natl. Acad. Sci. USA 1991, 88, 1059–1063. [Google Scholar] [CrossRef] [PubMed]
- Garcia, T.; Li, J.; Sureau, C.; Ito, K.; Qin, Y.; Wands, J.; Tong, S. Drastic reduction in the production of subviral particles does not impair hepatitis B virus virion secretion. J. Virol. 2009, 83, 11152–11165. [Google Scholar] [CrossRef] [PubMed]
- Junker, M.; Galle, P.; Schaller, H. Expression and replication of the hepatitis B virus genome under foreign promoter control. Nucleic Acids Res. 1988, 15, 10117–10132. [Google Scholar] [CrossRef]
- Zong, L.; Qin, Y.; Jia, H.; Zhou, H.; Chen, C.; Qiao, K.; Zhang, J.; Wang, Y.; Li, J.; Tong, S. Two-way molecular ligation for efficient conversion of monomeric hepatitis B virus DNA constructs into tandem dimers. J. Virol. Methods 2016, 233, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Bang, G.; Kim, K.H.; Guarnieri, M.; Zoulim, F.; Kawai, S.; Li, J.; Wands, J.; Tong, S. Effect of mutating the two cysteines required for HBe antigenicity on hepatitis B virus DNA replication and virion secretion. Virology 2005, 332, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Parekh, S.; Zoulim, F.; Ahn, S.H.; Tsai, A.; Li, J.; Kawai, S.; Khan, N.; Trepo, C.; Wands, J.; Tong, S. Genome replication, virion secretion, and e antigen expression of naturally occurring hepatitis B virus core promoter mutants. J. Virol. 2003, 77, 6601–6612. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zong, L.; Sureau, C.; Barker, L.; Wands, J.R.; Tong, S. Unusual features of sodium taurocholate cotransporting polypeptide as a hepatitis B virus receptor. J. Virol. 2016, 90, 8302–8313. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Jia, H.; Zhang, F.; Qin, Y.; Zong, L.; Yuan, Q.; Wang, Y.; Xia, N.; Li, J.; Wen, Y. Functional characterization of hepatitis B virus core promoter mutants revealed transcriptional interference among co-terminal viral mRNAs. J. Gen. Virol. 2016, 97, 1–9. [Google Scholar]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The imagej ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518. [Google Scholar] [CrossRef] [PubMed]
- Bruss, V. A short linear sequence in the pre-S domain of the large hepatitis B virus envelope protein required for virion formation. J. Virol. 1997, 71, 9350–9357. [Google Scholar] [PubMed]
- Bruss, V. Envelopment of the hepatitis B virus nucleocapsid. Virus Res. 2004, 106, 199–209. [Google Scholar] [CrossRef] [PubMed]
- The Hepatitis B Virus Database (HBVdb). Available online: https://hbvdb.ibcp.fr/HBVdb/HBVdbIndex (accessed on 24 March 2017).
- Ou, J.H.; Rutter, W.J. Regulation of secretion of the hepatitis B virus major surface antigen by the preS-1 protein. J. Virol. 1987, 61, 782–786. [Google Scholar] [PubMed]
- Persing, D.H.; Varmus, H.E.; Ganem, D. Inhibition of secretion of hepatitis B surface antigen by a related presurface polypeptide. Science 1986, 234, 1388–1391. [Google Scholar] [CrossRef] [PubMed]
- Ladner, S.K.; Otto, M.J.; Barker, C.S.; Zaifert, K.; Wang, G.H.; Guo, J.T.; Seeger, C.; King, R.W. Inducible expression of human hepatitis B virus (HBV) in stably transfected hepatoblastoma cells: A novel system for screening potential inhibitors of HBV replication. Antimicrob. Agents Chemother. 1997, 41, 1715–1720. [Google Scholar] [PubMed]
- Guo, H.; Jiang, D.; Zhou, T.; Cuconati, A.; Block, T.M.; Guo, J.T. Characterization of the intracellular deproteinized relaxed circular DNA of hepatitis B virus: An intermediate of covalently closed circular DNA formation. J. Virol. 2007, 81, 12472–12484. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, T.; Iwanski, A.; Will, H.; Sterneck, M. Deficiency in virion secretion and decreased stability of the hepatitis B virus immune escape mutant G145R. Hepatology 2003, 38, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Waters, J.A.; Kennedy, M.; Voet, P.; Hauser, P.; Petre, J.; Carman, W.; Thomas, H.C. Loss of the common “A” determinant of hepatitis B surface antigen by a vaccine-induced escape mutant. J. Clin. Investig. 1992, 90, 2543–2547. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, G.; Caccamo, G.; Filomia, R.; Pollicino, T. Occult HBV infection. Semin. Immunopathol. 2013, 35, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Carman, W.F.; Karayiannis, P.; Waters, J.; Thomas, H.; Zanetti, A.; Manzillo, G.; Zuckerman, A.J. Vaccine-induced escape mutant of hepatitis B virus. Lancet 1990, 336, 325–329. [Google Scholar] [CrossRef]
- Xiang, K.H.; Michailidis, E.; Ding, H.; Peng, Y.Q.; Su, M.Z.; Li, Y.; Liu, X.E.; Dao Thi, V.L.; Wu, X.F.; Schneider, W.M.; et al. Effects of amino acid substitutions in hepatitis B virus surface protein on virion secretion, antigenicity, HBsAg and viral DNA. J. Hepatol. 2016, 66, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Tsubota, A.; Hirokawa, T.; Kumada, H.; Yang, Z.; Tanaka, H. A unique amino acid substitution, T126I, in human genotype C of hepatitis B virus S gene and its possible influence on antigenic structural change. Gene 2006, 383, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Yuan, Q.; Chen, P.J.; Zhang, Y.L.; Chen, C.R.; Zheng, Q.B.; Yeh, S.H.; Yu, H.; Xue, Y.; Chen, Y.X. Influence of mutations in hepatitis B virus surface protein on viral antigenicity and phenotype in occult HBV strains from blood donors. J. Hepatol. 2012, 57, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Kwei, K.; Tang, X.; Lok, A.S.; Sureau, C.; Garcia, T.; Li, J.; Wands, J.; Tong, S. Impaired virion secretion by hepatitis B virus immune escape mutants and its rescue by wild-type envelope proteins or a second-site mutation. J. Virol. 2013, 87, 2352–2357. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zhou, X.; Jia, H.; Chen, C.; Zhao, W.; Zhang, J.; Tong, S. Stronger enhancer ii/core promoter activities of hepatitis B virus isolates of B2 subgenotype than those of C2 subgenotype. Sci. Rep. 2016, 6, 30374. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| preS1 aa Position | ||||||||||||
| 10 | 35 | 39 | 45 | 48 | 51 | 54 | 57 | 60 | 84 | 108 | 115 | |
| Genotype B (2246 isolates) | K | K | E | L | H | N | D | K | V | L | L | T |
| Genotype C (2227 isolates) | Q | G | N | F | N | H | E | Q | A | I | I | S |
| preS2 aa Position | ||||||||||||
| 130 | 132 | 138 | 152 | 154 | 155 | 156 | 158 | 160 | 165 | 167 | 172 | |
| Genotype B (2419 isolates) | T | Q | A | S | A | Q | N | V | A | L | K | V |
| Genotype C (2377 isolates) | A | L | G | N | V | P | T | A | P | F | R | A |
| S aa Position | ||||||||||||
| 4 | 5 | 8 | 24 | 45 | 47 | 49 | 56 | 57 | 59 | |||
| Genotype B (4224 isolates) | I | A | L | K | T | V | L | Q | I | S | ||
| Genotype C (7183 isolates) | T | T | F | R | A | T | P | P | T | N | ||
| 64 | 85 | 110 | 113 | 126 | 143 | 160 | 161 | 200 | 213 | |||
| Genotype B (4224 isolates) | C | C | I | S | T | T | K | Y | F | M | ||
| Genotype C (7183 isolates) | S | F | L | T | I | S | R | F | Y | L | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, H.; Qin, Y.; Chen, C.; Zhang, F.; Li, C.; Zong, L.; Wang, Y.; Zhang, J.; Li, J.; Wen, Y.; et al. The Envelope Gene of Hepatitis B Virus Is Implicated in Both Differential Virion Secretion and Genome Replication Capacities between Genotype B and Genotype C Isolates. Viruses 2017, 9, 62. https://doi.org/10.3390/v9040062
Jia H, Qin Y, Chen C, Zhang F, Li C, Zong L, Wang Y, Zhang J, Li J, Wen Y, et al. The Envelope Gene of Hepatitis B Virus Is Implicated in Both Differential Virion Secretion and Genome Replication Capacities between Genotype B and Genotype C Isolates. Viruses. 2017; 9(4):62. https://doi.org/10.3390/v9040062
Chicago/Turabian StyleJia, Haodi, Yanli Qin, Chaoyang Chen, Fei Zhang, Cheng Li, Li Zong, Yongxiang Wang, Jiming Zhang, Jisu Li, Yumei Wen, and et al. 2017. "The Envelope Gene of Hepatitis B Virus Is Implicated in Both Differential Virion Secretion and Genome Replication Capacities between Genotype B and Genotype C Isolates" Viruses 9, no. 4: 62. https://doi.org/10.3390/v9040062