
Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance

1
Laboratory of Medical Parasitology, Central Institute of the Bundeswehr Medical Service, Koblenz, Andernacherstrasse 100, 56070 Koblenz, Germany
2
Institute of Integrated Sciences, Department of Biology, Parasitology and Infection Biology Group, University of Koblenz-Landau, Universitätsstrasse 1, 56070 Koblenz, Germany
*
Author to whom correspondence should be addressed.
Viruses 2017, 9(4), 65; https://doi.org/10.3390/v9040065
Submission received: 20 December 2016
/
Revised: 22 March 2017
/
Accepted: 24 March 2017
/
Published: 1 April 2017
(This article belongs to the Special Issue Viruses of Protozoa)
Abstract
:Free-living amoebae (FLA) are parasites within both humans and animals causing a wide range of symptoms and act as hosts of, and vehicles for phylogenetically diverse microorganisms, called endocytobionts. The interaction of the FLA with sympatric microorganisms leads to an exceptional diversity within FLA. Some of these bacteria, viruses, and even eukaryotes, can live and replicate intracellularly within the FLA. This relationship provides protection to the microorganisms from external interventions and a dispersal mechanism across various habitats. Among those intracellularly-replicating or -residing organisms there are obligate and facultative pathogenic microorganisms affecting the health of humans or animals and are therefore of interest to Public Health Authorities. Mimiviruses, Pandoraviruses, and Pithoviruses are examples for interesting viral endocytobionts within FLA. Future research is expected to reveal further endocytobionts within free-living amoebae and other protozoa through co-cultivation studies, genomic, transcriptomic, and proteomic analyses.
Keywords:
free-living amoebae; FLA; Acanthamoeba; vectors; hosts; endocytobionts; endosymbionts; Pandoravirus; Mimivirus; Pithovirus; public health1. Introduction
Free-living amoeba (FLA) are protozoa ubiquitously found in nature. They form a rather heterogeneous group of facultative parasitic amoebae within the free-living protozoa without common phylogenetic, systematic, or taxonomic origin [1]. Among the FLA, only four genera are pathogenic for humans and animals: Acanthamoeba, Balamuthia, Naegleria, and Sappinia. The taxonomy of FLA was revised several times in consequence of the availability of genomic sequence data. Regarding morphologic, biochemical and molecular data, a new taxonomic classification was proposed by the International Society of Protozoologists. According to this classification these pathogens has been placed in two ‘Super Groups’. Acanthamoeba and Balamuthia were classified under the super group Amoebozoa: Acanthamoebidae; Naegleria fowleri under the super group Excavata: Heterolobosia: Vahlkampfiidae; and Sappinia under the super group Amoebozoa: Flabellinea: Thecamoebidae [2].
The FLA are widespread in many habitats including soil, dust, air, seawater, drinking water, swimming pools, sewage, eyewash solutions, contact lenses, dialysis units, and dental treatment units [3]. Their prevalence in water networks is associated with biofilms, where they live sympatric within a biocoenosis together with other microorganisms. These biofilms serve as feeding grounds for the FLA and provide protection to a certain degree while the FLA adhere to their surfaces. They play a role in the reduction of bacterial biomass and the regeneration of nutrients. In such a biocoenosis there are multiple interactions between FLA and other microorganisms.
The trophozoites of FLA are the vegetative form and in this state, they can serve as prey for other microorganisms, such as amoebophagous fungi [4] or act as aerobic heterotrophic predators by feeding on algae, bacteria, cyanobacteria, and fungi, as well as on smaller protozoa. However, there are numerous pathogenic microorganisms of hygienic relevance which may survive and evade phagocytosis and digestion within the FLA trophozoites. One protective mechanism used by some species is the cyst. This life stage protects the organism from adverse environmental conditions and, therefore, makes these pathogenic organisms enormously resistant, persistent, and tenacious.
This review highlights the pathogenic potential of certain FLA and the consequences for public health caused by their interactions with other microorganisms.
2. Pathogenic Free-Living Amoebae
For a long time, FLA were considered to be harmless protozoa of soil and water. However, research since the 1960s has demonstrated that FLA can be pathogenic to humans and animals [5] with near 100% morbidity from some strains. When FLA are in intimate contact with humans they can provoke serious infections, for example by using contaminated contact lenses. The following section describes the diseases caused by the four well-characterized pathogenic genera of FLA, Acanthamoeba, Balamuthia, Naegleria, and Sappinia, as well as genera most recently recognized to be involved in infections, and even newly-described species.
The Acanthamoebae are the most well-known facultative parasitic FLA. Their pathogenicity varies among their species or strains which are currently divided into 18 sequence types (genotypes; T1–18) based on the nuclear small-subunit ribosomal RNA (rRNA) gene, rather than their morphology. The predominant genotype in human pathogenic Acanthamoebae is T4; other genotypes have also been reported as pathogens [6,7,8,9]. As etiological agents of the so-called acanthamoebiasis, they can trigger several specific disease symptoms in humans. The amoebiasis of the central nervous system is called granulomatous amebic encephalitis (GAE). The infection route of Acanthamoebae mostly include the lower respiratory tract and skin lesions followed by hematogenous spread, finally reaching the central nervous system through the blood-brain barrier [3]. GAE differs clinically and morphologically from the primary amoebic meningoencephalitis (PAM), which is caused by Naegleria fowleri, showing subacute to chronic progress [10,11]. To date, approximately 150 cases have been reported worldwide and the infection is almost always fatal because of the difficulty and delay in diagnosis and lack of optimal therapy [3].
Cutaneous acanthamoebiasis is a rare opportunistic infection and occurs almost exclusively in immunocompromised persons, similar to GAE. Characteristic lesions of the skin, such as ulcers and erythematous sores, contain Acanthamoeba trophozoites and cysts and are a route of entry into the blood stream, with subsequent hematogenous spread to different tissues. Alternatively, the Acanthamoeba keratitis is not necessarily associated with an immune suppression, but rather with a trauma, exposure to contaminated water or, particularly, the improper handling of contact lenses, which promotes infection [12,13].
The Naegleria infection is a rare disease, which is caused by Naegleria fowleri. Up to now, only Naegleria fowleri is known in humans as an etiological agent of PAM, which has a worldwide distribution of more than 300 reported cases, mostly from the United States [14]. It is almost always fatal; only three people in the United States out of 138 have survived infection [15]. This infection generally occurs in previously healthy children and young adults with a history of swimming and other recreational activities in warm freshwater lakes and ponds [16,17]. When contaminated water enters the nose N. fowleri reaches the brain by migration alongside the olfactory nerves [18]. The naegleriosis shows up as a diffuse meningoencephalitis with swollen and edematous cerebral hemispheres. After a critical period with ataxia, convulsions, and coma, the patient may die within a very short period of time [16,17].
The clinical picture of GAE by Balamuthia mandrillaris is characterized by headache and neck stiffness. The Balamuthia infection is also known as granulomatous Balamuthia encephalitis (BAE) [19]. So far, approximately 100 cases have been reported, and only three survivors are known. The infection is chronic and the time between infection and appearance of neurological symptoms may range from one month to about two years [3]. The portal of entry are mostly skin lesions contaminated by soil and most cases of BAE are reported from warm regions [20]. However, quite recently, DNA of B. mandrillaris was found unexpectedly in a routine screen for Acanthamoeba in a contact lens sample. This demonstrates how easily pathogens might be missed if their occurrence is not expected [21].
The two-nucleated Sappiniae have been identified only once as an etiological agent of an amoeba-associated encephalitis with a favorable clinical outcome [22]. This amoebic encephalitis (SAE) was first associated with Sappinia diploidea, while an affiliation to Sappinia pedata was later proved using molecular biological studies [23].
Hartmannella sp. has also been discovered in keratitis cases, but determining whether they were pathogens or merely contaminants, still remains controversial [24,25]. The same applies to Vahlkampfiae, which were found together with Hartmannellae while diagnosing a keratitis of a contact lenses wearer [26]. Paravahlkampfia francinae, a recently described species within the genus Paravahlkampfia, was isolated from the cerebrospinal fluid of a patient with PAM-like symptoms, usually caused by Naegleria fowleri [27]. Allovahlkampfia spelae, a newly-discovered amoeba, was identified as a cause of chronic human keratitis in an Egyptian patient in 2016 [28]. The only involvement of Dictyostelium polycephalum, a social amoeba, was described in 2010 in a human keratitis in India [29]. The infection occurred in an immunocompetent person without history of ocular injury or surgery.
3. FLA as Vectors of Diverse Microorganisms (Biodiversity within FLA)
Biofilms serve the FLA as basic food and protection sources, where the FLA adhere as trophozoites on the surfaces [30], and they interact with many different microorganisms in their biocoenoses, especially in these biofilms. While the FLA usually use bacteria and other microorganisms as food sources, some of the prey organisms manage to survive within their grazing predators or even proliferate within their cytoplasm or their nucleus.
The generic term ’endocytobionts’ was introduced because the interaction of FLA with phylogenetically diverse microorganisms lead to a plethora of different outcomes [5,12,31]. This term avoids any predetermination of possible features of these ’relationships’, such as parasitism, symbiosis (‘endosymbionts’), phoresy, zoochory, or mutualism, especially since the behavior of the endocytobionts may vary depending on the temperature (see Legionella) or other conditions [5].
FLA play an important role in the transmission and dispersal of pathogenic mircroorganisms by acting as a ‘trojan horse’ serving as vehicles for the colonization of new habitats or hosts. The relationships between FLA and the diverse intracellular microorganisms shows similarities to the relationships between some prokaryonts and macrophages: the virulence of pathogenic microorganisms to their amoebal host indicates a virulence and an adaptation towards human or animal cells, e.g., macrophages [32,33,34,35,36]. FLA serves as a ‘training ground’, a prerequisite for a predation-induced adaptation to life in a eukaryotic host [5,37,38,39,40]. It is known that the virulence in bacteria is often correlated to resistance to antibiotics or disinfection measures [41,42]. Conclusively, FLA-endocytobiont relationships are of relevance for the origin of pathogens and they make an impact on evolutionary processes [5]. The impact of the host-endocytobiont relationship may, in turn, also lead to an increase of virulence of the FLA [5,41,43].
4. Host-Microorganism Interactions
4.1. Amoebal Co-Culture vs. In Situ Interactions
The analysis of interactions of FLA and pathogenic bacteria species, or other microorganisms, are of great value for public health. Since mechanisms allowing bacteria to escape from phagocytosis and digestion by FLA are believed to be similar to the mechanisms allowing the same microorganisms to evade a similar fate by macrophages, co-culture with FLA has been proposed as a new tool to recover and identify potentially new pathogens from environmental samples [44]. By incubating samples on axenically-grown FLA, and subsequent staining and visualization of microorganisms, the putative development of bacteria inside FLA can be analyzed. Molecular methods, like sequencing of 16S ribosomal DNA (rDNA), are subsequently used to identify bacterial isolates [45]. The careful selection of specific FLA for these experiments is necessary since the host range of pathogens might be narrow and, therefore, by choosing the wrong (those not susceptible or insensitive) FLA-strain, the isolation of a specific pathogen might be missed. Expanding the panel of used FLA or other protozoa is, therefore, promising [46].
A restriction of this widely-used co-culture experiments is the in vitro framework of the laboratory assays. Although the chosen FLA and the microorganism might be inhabitants of the same natural habitat, the interaction is exclusively shown for the precise experimental conditions. Hence, chosen protocols for the recovery of intra-amoebic microorganisms might miss the particular condition required for a specific interaction (e.g., bacteria/amoeba ratio or temperature). There is a significant qualitative difference, as to whether the microorganisms could be isolated from a real specimen (e.g., an environmental sample or patient specimen) or solitary under laboratory conditions. It is another qualitative differentiation mark, whether the endocytobionts are only transported by FLA as vector/carriers, or whether they are able to proliferate intracellularly with the FLA as the host [5]. Table 1 gives an overview of a selection of microorganisms of public health significance interacting with FLA in which in situ and in vitro interactions are listed separately. A comprehensive list of pathogens, including the nature of interaction between bacterial pathogen and amoebal host, was published by Thomas et al. [44].
4.2. Interactions of FLA and Bacteria
Among the numerous bacterial species interacting successfully with FLA, or even other protozoa, many are known to be pathogenic for humans [118]. More than 100 proven pathogens for humans are described as surviving and/or proliferating when in contact with various amoebal species [44]. These impressive numbers seem to be underestimated since in most studies analyzing host-pathogen interactions often just one, typically A. polyphaga, or just a few protozoa species/strains were used and a range of bacterial pathogens have not yet been tested to interact with FLA. Important pathogens like mycobacteria, Legionella pneumophila, Legionella micdadei, Vibrio cholera, and Francisella tularensis are able to survive not only in trophozoites but even in cysts [100,101,119,120,121,122,123]. Since bacteria incorporated in trophozoites show increased resistance to biocides, the resistance of bacteria inside cysts is even more increased [74,121,124]. Hence, pathogenic bacteria internalized in amoebae represent a high risk for public health because they are not reached by conventional biocide treatment. Some of these bacteria-FLA interactions are described below in more detail.
4.2.1. Legionella pneumophilia
Legionella pneumophilia was first described more than 35 years ago, as the etiological agent of the Legionaries’ disease. It is widespread in nature and can proliferate inside FLA like Acanthamoeba spp., Naegleria sp., Hartmannella sp. and other protozoa [5]. Legionella is able to avoid degradation by the phagolysosomes in the amoeba and hence, FLA might also contribute to the virulence characteristics of the bacteria interacting with human macrophages [125].
Co-cultivation with FLA leads to regeneration of bacteria from the so called ’viable-but-non culturable’ state due to starvation or biocide application as was shown for Legionella pneumophilia [126]. Beside this resuscitation effect of amoebae on Legionella, the interaction also enables the bacterium to tolerate elevated concentrations of biocides. Only a 10-fold kill was achieved with an isothiazolone derivate against intra-amoebal-grown L. pneumophilia whereas, for in vitro-grown bacteria, a 1000-fold kill was observed [127]. Thus, insufficient biocide application might generate serious problems since internalization makes pathogens motile with the FLA as vectors or transport vehicles for the pathogenic bacteria.
4.2.2. FLA-Bacteria Interactions Other than Legionella
The facultative intracellular pathogen Listeria monocytogenes is a food-borne opportunistic pathogen which can switch from an environmental saprophyte to a potentially fatal human pathogen. L. monocytogenes is not digested when incorporated in Acanthamoeba and bacteria are released from vegetative trophozoites, whereas bacteria are killed in cysts [56].
Campylobacter jejuni is able to resist digestion in Acanthamoeba for a long time and cells survived for longer periods when co-cultured with Acanthamoeba sp. than when grown in culture alone [75,128]. In addition, moderately acidic conditions were shown to trigger Campylobacter jejuni adhesion and internalization into Acanthamoeba polyphaga [129]. Therefore, improper biocide application might increase internalization of pathogens. Co-cultivation of C. jejuni with A. castellanii increases its survival of chemical disinfection with an iodine-containing product [75]. Additionally, transmission of endocytobionts has been reported (e.g., Campylobacter jejuni in Acanthamoeba castellanii to broilers) [130].
Within the Mycobacterium genus, the species Mycobacterium tuberculosis and Mycobacterium leprae are considered to be the most important human pathogens. Mycobacterium avium and other species of the Mycobacterium genus are opportunistic pathogens having a high impact for HIV/AIDS patients [131]. Mycobacterium avium enters and replicates in A. castellanii and amoeba-grown mycobacteria have an enhanced virulence in the mouse model [41].
Arcobacter butzleri can establish a long-lasting relation with Acanthamoeba castellanii for up to 10 days in laboratory co-culture experiments [132]. The pathogen is also able to replicate inside the amoeba suggesting a possible endocytobiotic interaction and a possible role of the FLA as a transport vehicle for the bacteria [76,77]. In co-culture experiments with Acanthamoeba castellanii, Yersinia enterocolitica can evade digestion and survives at least 14 days in nutrient-poor and nutrient-rich medium. The bacteria seem to evade from digestive vacuoles and replicate within the cytoplasm of the amoeba [133]. Ingested Yersinia were protected from chlorination since the pathogen has increased resistance to free chlorine residues [74]. Chlorine is a widely used chemical agent for disinfection of surfaces and, therefore, a failure of disinfection due to internalization of Yersinia by FLA has to be considered.
Salmonella typhimurium replicates inside contractile vacuoles and even food vacuoles of Acanthamoeba polyphaga to a high degree (100–200 cells per vacuole) [72]. Due to the high density of cell growth, the protozoa is now considered to be an environmental reservoir for this bacteria [78]. The interaction of S. typhimurium and Salmonella dublin with Acanthamoeba polyphaga and Acanthamoeba rhysodes upregulates pathogenicity factors of the bacteria indicating an enhancement of virulence through the amoebal passage [73,134]. In addition, exposure of Salmonella typhimurium to different rumen protozoa also enhances virulence of the bacterium indicating that virulence-enhancement is a common mechanism in this relationship [135,136,137].
5. Viruses in Close Association with FLA
Viruses are highly abundant and ubiquitous components of the biosphere. Their genomes are extremely variable concerning their composition, organization and size [138]. Thus, viruses represent specialized, simple organisms that interact in their intracellular environment with potential hosts and sympatric occurring microorganisms [139]. The story of viruses in close association with FLA began in 1974 when Schuster and Dunnebacke detected virus-like particles in Naegleria sp. [140].
5.1. Coxsackievirus and Adenovirus
The human pathogenic coxsackievirus B3 (CVB-3) is incorporated in Acanthamoeba trophozoites without changing the infective activity of the virion [112]. In co-culture experiments CVB-3 remains infectious even after a complete 6-month cycle of encystment and excystment. Acanthamoeba are therefore suggested to play an active role as a vector in the survival of coxsackieviruses. Acanthamoebae are also considered to be a potential reservoir and a vector/carrier of adenoviruses to humans [111]. Viral DNA of different human adenoviruses (HAdv) serotypes has been found in Acanthamoeba isolates from water sources in the Canary Islands [108]. The serotype HAdV-2 was found to be the most frequently detected.
5.2. Mimiviridae
Additionally, the interaction of these proven human pathogenic viruses with amoebae the discovery of the so-called giant viruses has increased interest in the association of viruses and FLA. The 0.8 µm large Acanthamoeba polyphaga Mimivirus (APMV) was described in 2003 and placed in the family of the Mimiviridae [109]. It was isolated in 1992 (from a cooling tower in the UK at Bradford) and this intracellular endocytobiont developing within Acanthamoebae was first considered as a bacterium which was finally named ‘Bradford coccus’ [109]. This misinterpretation still reflects in the naming of Mimiviruses (‘mimicking Microbe virus’). Virion factories were described as functional dynamic structures, modifying areas of the infected trophozoites to assure viral morphogenesis [141]. To date, about 100 amoebal Mimiviruses from various environmental water and soil samples, but also human samples have been isolated [142]. Phylogenomics separate these viruses in three lineages named A, B, C with their main representatives Mimivirus, Acanthamoeba polyphaga moumouvirus, and Megavirus chilensis, respectively. Megavirus chilensis was isolated from a marine sediment sample in the aftermath, which had even a larger genome than the Mimivirus. Already at this time, it was evident that the extent of the complexity of these viruses that multiply in protozoa, is not yet able to be predicted.
5.3. Marseilleviridae
About five years after the discovery of the Acanthamoeba polyphaga Mimivirus, the Acanthamoeba polyphaga Marseille virus was isolated from a cooling tower in Paris and placed taxonomically within the Iridoviridae family [143]. This giant virus belonged to a further new family within the nucleocytoplasmatic large DNA viruses (NCLDV) and showed at that time the fifth-largest virus genome. A separate order (Megavirales) was proposed to combine the giant viruses such as the Marseillevirus and related viruses [142,144]. Since the discovery of the giant virus in amoebae it has been searched extensively (especially in biofilms and sediments) for other viruses as endocytobionts in FLA and many new representatives have been found [142]. It could never be excluded that the Megavirales could actually represent a separate branch of micro-organisms different from previously described domains of life and, thus, initiate a revolution in the classification of micro-organisms.
5.4. Pandoraviruses and Pithoviruses
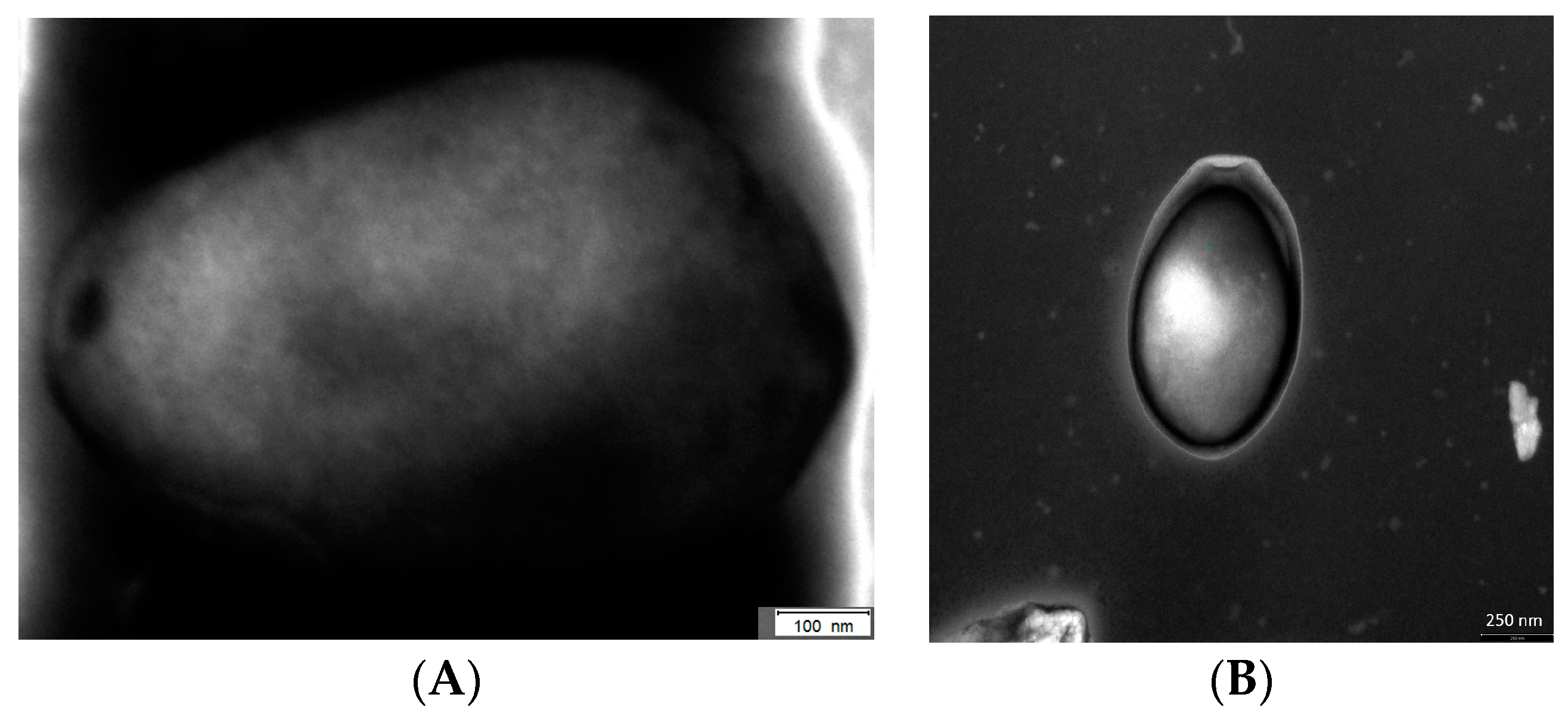
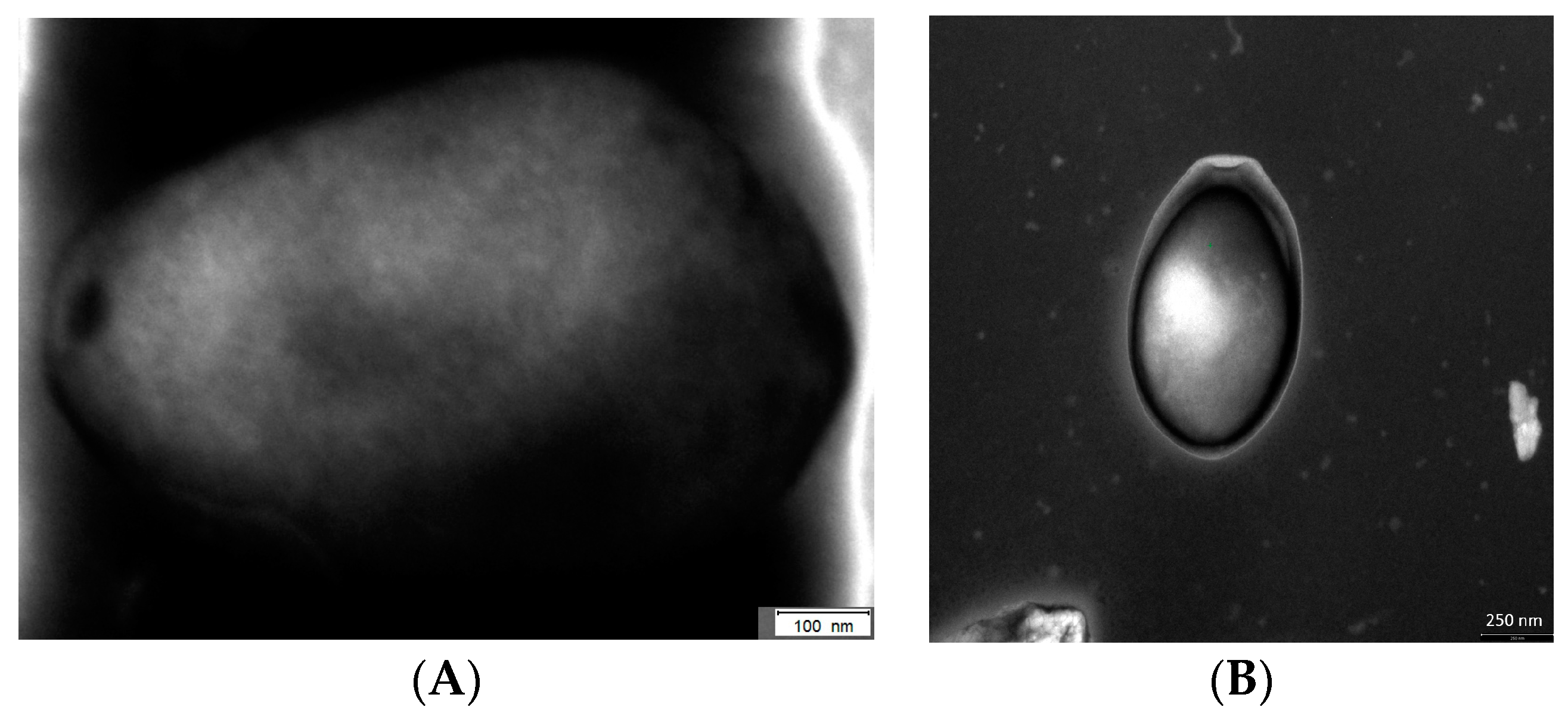
About 10 years after the discovery of the Mimivirus in Acanthamoebae, more giant viruses in FLA were investigated and named as Pandoraviruses. It turned out that in 2008 the first Pandoravirus has been described in its natural host while the taxonomic affiliation was not solved at that time [12,115,145]. These viruses do not have any similarity with the presently-known virus families on the genomic level. Even morphologically the virion was not comparable to any other viral structure (Figure 1) [146]. So far, three species of these extraordinary viruses are analyzed in detail, Pandoravirus salinus, Pandoravirus dulcis, and Pandoravirus inopinatum [115,147,148]. Virion factories have never been clearly observed in Pandoraviruses like in Mimivirus. However, similar structures have been proposed for the replication of Pandoraviruses [146,149].
Pithoviruses were analyzed on the genomic level in 2014 when a sample from >30,000-year-old Siberian permafrost was inoculated on Acanthamoeba castellanii host amoeba [116]. Similar to Pandoravirus, a relative of Pithoviruses, the endocytobiont KC5/2 was first described by Hofmann et al. before the identity as a giant virus was evident [110]. The Pithovirus sibericum particle/virion is 1.5 µm in length but has a smaller genome compared with the Pandoraviruses [116,150].
5.5. Faustovirus and Mollivirus
Several isolates of Faustovirus, an Asfarvirus-related lineage of giant viruses, were isolated using Vermamoeba vermiformis, the first time an FLA other than Acanthamoeba sp. has been used [46]. This virus forms icosahedral virions of 200 nm and the genome encodes for 451 predicted proteins from which about two-thirds do not show significant similarity to known protein-encoding genes. Prominent virus factories were built within the host cells and complete viral particles were released at 18 to 20 h post-infection [46].
Mollivirus sibericum was isolated from the same permafrost sample as Pithovirus sibericum [117]. The spherical virion of 0.6 µm diameter differs significantly from P. sibericum and encloses a genome which encodes 523 proteins, 16% of them have their closest homologies to proteins from Pandoravirus. Astonishing 10% of the encoded proteins may have been acquired by horizontal gene transfer from its host since their closest homologs are from Acanthamoeba castellani [117].
5.6. Giant Viruses as Putative Pathogens
Most of the viruses associated with FLA are isolated by using amoebal co-cultivation. Therefore, except for APMV and Pandoravirus inopinatum, which were isolated together with their respective Acanthamoeba host, and Pandoravirus inopinatum even from a patient specimen, the real host in the environment is not known [12]. Although their potential pathogenicity for humans is not clear, at least for Pandoravirus inopinatum, it was shown how easily theses giant viruses get in close contact to humans. In addition, APMV interacts with the interferon system of vertebrates and replicate in human and murine phagocytes [151]. Serology studies support the role of mimiviruses in hospital-acquired pneumonia [152]. Hence, the possible role of these giant viruses as pathogen needs to be clarified.
5.7. Giant Viruses and Evolution
The discovery of these large DNA viruses led to questioning the evolutionary significance of these unusual viruses, because viruses are generally in much closer relationship to their host cells as to other viruses [153]. Features of complex genetic interaction such as interviral gene transfer, the correlation to virophages (e.g., Sputnik), lateral gene transfer, and their role with respect to their evolution, remained enigmatic for a long time. It was known that the laterally acquired genes are associated with the sympatric lifestyle (in association with viruses, bacteria, and eukaryotes) and the simultaneous occurrence of virophages [154,155]. Lateral gene transfer obviously occurred to a great extent in Acanthamoeba castellanii and led to integration of viral genes from all known giant NCLDVs [156]. Some of these genes are actively transcribed in the amoeba suggesting their domestication during amoebal evolution. Giant viruses, such as the Marseillevirus use their amoebal hosts (e.g., Acanthamoebae) as ‘gene melting pots’, a place where horizontal gene transfer occurs. The FLA play a role in the genesis of complex, chimeric genomes [113], for example Mimivirus acquired tyrosyl-tRNA synthetase from the host amoeba Acanthamoeba polyphaga [155].
Most recently, a comprehensive analysis of the nuclear genome of the A. castellanii Neff strain suggests the existence of a yet-undiscovered family of amoeba-infecting NCLDV [156]. A bioinformatics tool which especially screens metagenomes for giant virus-related sequences might improve our knowledge about the presence and prevalence of giant viruses in the environment and the human body [157]. Therefore, the story of giant viruses in amoebae and the discovery of new viruses seem to be going on.
6. Interactions of FLA with Protozoa or Fungi
Only a few examples of interaction between FLA and protozoa (and also fungi) have been analyzed so far, including examples of endozoochory or parasitism [61,105]. Cryptosporidium parvum causes cryptosporidiosis, a parasitic disease of the mammalian intestinal tract [106]. Especially immunocompromised individuals, including HIV/AIDS patients, often suffer from intractable diarrhea, which can be fatal. In co-cultivation experiments the capacity of Acanthamoeba to predate Cryptosporidium oocysts was demonstrated [105,107]. Winiecka-Krusnell et al. examined the interaction of FLA and Toxoplasma gondii [158]. Oocysts were actively internalized and can establish infection in a murine model.
Cryptococcus neoformans is a microscopic fungus that lives in the environment throughout the world. People can become infected after breathing in C. neoformans, but usually people who have weakened immune systems are at most risk, particular those who have advanced HIV/AIDS [159]. Cryptococcosis usually affects the lungs or the central nervous system, however, sometimes life-threatening brain infections (cryptococcal meningitis) occur. In vitro co-culture experiments showed an ingestion and even replication of the yeast cells inside A. castellanii (enclosed in membrane-bound vesicles, the phagocytic vacuole) leading to the death of the amoeba [42]. Both passage in Acanthamoeba castellanii and Dictyostelium discoideum enhance virulence of Cryptococcus neoformans [42,160].
In addition to human pathogenic endocytobionts with public health importance, fungal parasites of FLA are also observed. Cochlonema euryblastum belongs to the order Zoopagales and specifically infects Thecamoeba quadrilineata [104]. After the uptake by this FLA conidia germinate to produce coiled thalli and within two days conidiogenous hyphae are produced which break through the protoplasmic pellicle of the amoeba. Some experts stated that fungal virulence may have evolved, and been maintained, as a countermeasure to predation by amoeba since pathogenicity in mammals is based on multiple virulence factors also required for fungal survival during interactions with non-vertebrate hosts [161].
7. Conclusions
There are numerous examples of such microorganisms which survive, grow, or proliferate inside amoebae [5]. The intracellular life in protozoa protects the endocytobionts from adverse environmental conditions whether of natural or artificial origin. In particular, cyst-forming FLA species and their endocytobionts are protected from biocides (as well as disinfectants, contact lens cleaners, other antiseptics, herbicides, pesticides, PCBs, and heavy metals) [121,162,163]. The fact that FLA can serve as hosts of and vehicles (vectors) for microorganisms, has an impact on the sector of public health. Not only is the resistance to biocides insides cysts increased but also virulence of some pathogens is boosted by exposure to FLA [73,134,135,136,137].
The host-endocytobiont relationships in FLA promote evolutionary processes and development of pathogenicity. It could mean that the amoebal passage is a prerequisite for the development of virulence factors—and, thus, for the development of human-pathogenic microorganisms. FLA then play an important role for their endocytobionts in the selection and evolution of virulence characteristics and the adaptation to (human) macrophages [5,36].
Since many of the pathogenic microorganisms which interact with FLA are water-borne there is a clear risk for quality in water networks. The resistance of FLA to drinking water treatments favors distribution of incorporated pathogens. Extracellular and intracellular biofilms possess an importance for evolutionary processes and the emergence of pathogens in biofilms. Therefore, in this regard, FLA can be viewed as evolutionary triggers or incubators.
Acknowledgments
The authors thank Dimitrios Frangoulidis and Claudia Kahlhofer (Bundeswehr Institute of Microbiology, Munich) for the outstanding EM-photos of Pandoravirus inopinatum. We also wish to thank Richard F.D. Shepherd for review and English-language editing of the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Walochnik, J.; Aspöck, H. Amöben: Paradebeispiele für Probleme der Phylogenetik, Klassifikation und Nomenklatur. Denisia 2007, 20, 323–350. [Google Scholar]
- Adl, S.M.; Simpson, A.G.B.; Farmer, M.A.; Andresen, R.A.; Anderson, O.R.; Barta, J.R.; Bowser, S.S.; Brugerolle, G.; Fensome, R.A.; Fredericq, S.; et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J. Eukaryot Microbiol 2005, 52, 399–451. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, H.; Dendana, F.; Sellami, A.; Sellami, H.; Cheikhrouhou, F.; Neji, S.; Makni, F.; Ayadi, A. Pathogenic free-living amoebae: Epidemiology and clinical review. Pathol. Biol. 2012, 60, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kurek, R.; Scheid, P.; Michel, R. Darstellung von pilzartigen Endoparasiten bei freilebenden Amöben nach spezifischer Fluoreszenzanfärbung. Mikrokosmos 2010, 99, 327–330. [Google Scholar]
- Scheid, P. Relevance of free-living amoebae as hosts for phylogenetically diverse microorganisms. Parasitol. Res. 2014, 113, 2407–2414. [Google Scholar] [CrossRef] [PubMed]
- Booton, G.C.; Visvesvara, G.S.; Byers, T.J.; Kelly, D.J.; Fuerst, P.A. Identification and distribution of Acanthamoeba species genotypes associated with nonkeratitis infections. J. Clin. Microbiol. 2005, 43, 1689–1693. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; Nerad, T.A.; Visvesvara, G.S. Characterization of a new pathogenic Acanthamoeba species, A. byersi n. sp., isolated from a human with fatal amoebic encephalitis. J. Eukaryot. Microbiol. 2013, 60, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Grün, A.-L.; Stemplewitz, B.; Scheid, P. First report of an Acanthamoeba genotype T13 isolate as etiological agent of a keratitis in humans. Parasitol. Res. 2014, 113, 2395–2400. [Google Scholar] [CrossRef] [PubMed]
- Walochnik, J.; Aspöck, H. Die Diagnostik von Infektionen mit freilebenden Amöben (FLA). Laboratoriums Medizin 2005, 29, 446–456. [Google Scholar] [CrossRef]
- Butt, C.G. Primary amebic meningoencephalitis. N. Engl. J. Med. 1966, 274, 1473–1476. [Google Scholar] [CrossRef] [PubMed]
- Gardner, H.A.; Martinez, A.J.; Visvesvara, G.S.; Sotrel, A. Granulomatous amebic encephalitis in an AIDS patient. Neurology 1991, 41, 1993–1995. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.; Zoller, L.; Pressmar, S.; Richard, G.; Michel, R. An extraordinary endocytobiont in Acanthamoeba sp. isolated from a patient with keratitis. Parasitol. Res. 2008, 102, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Wright, P. Acanthamoeba keratitis. In Proceedings of the 5th International Conference on the Biology and Pathogenicity of Free-Living Amoebae (FLAM), Brussels, Belgium, August 1989. [Google Scholar]
- Gupta, N.; Bhaskar, H.; Duggal, S.; Ghalaut, P.S.; Kundra, S.; Arora, D.R. Primary Amoebic Meningoencephalitis: First reported case from Rohtak, North India. Braz. J. Infect. Dis. 2009, 13, 236–237. [Google Scholar] [CrossRef] [PubMed]
- Capewell, L.G.; Harris, A.M.; Yoder, J.S.; Cope, J.R.; Eddy, B.A.; Roy, S.L.; Visvesvara, G.S.; Fox, L.M.; Beach, M.J. Diagnosis, clinical course, and treatment of Primary Amoebic Meningoencephalitis in the United States, 1937–2013. J. Pediat. Infect. Dis. 2015, 4, e68–75. [Google Scholar] [CrossRef] [PubMed]
- Martinez, A.J.; Visvesvara, G.S. Free-living, amphizoic and opportunistic amebas. Brain Pathol. 1997, 7, 583–598. [Google Scholar] [CrossRef] [PubMed]
- Marciano-Cabral, F.; Cabral, G.A. The immune response to Naegleria fowleri amebae and pathogenesis of infection. FEMS Immunol. Med. Microbiol. 2007, 51, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Visvesvara, G. Free-living amebae as opportunistic agents of human disease. J. Neuroparasitol. 2010, 1, N100802. [Google Scholar] [CrossRef]
- Scheid, P. Balamuthiose. In Handbuch der Infektionskrankheiten; Hofmann, F., Ed.; ecomed Verlag: Hamburg, Germany, 2012. [Google Scholar]
- Siddiqui, R.; Khan, N.A. Balamuthia amoebic encephalitis: An emerging disease with fatal consequences. Microb. Pathog. 2008, 44, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Balczun, C.; Scheid, P.L. Detection of Balamuthia mandrillaris DNA in the storage case of contact lenses in Germany. Parasitol. Res. 2016, 115, 2111–2114. [Google Scholar] [CrossRef] [PubMed]
- Gelman, B.B.; Popov, V.; Chaljub, G.; Nader, R.; Rauf, S.J.; Nauta, H.W.; Visvesvara, G.S. Neuropathological and ultrastructural features of amebic encephalitis caused by Sappinia diploidea. J. Neuropathol. Exp. Neurol. 2003, 62, 990–998. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, Y.; da Silva, A.J.; Schuster, F.L.; Gelman, B.B.; Visvesvara, G.S. Molecular confirmation of Sappinia pedata as a causative agent of amoebic encephalitis. J. Infect. Dis. 2009, 199, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- De Jonckheere, J.F.; Brown, S. Non-Acanthamoeba amoebic keratitis. Cornea 1999, 18, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkova, N.; Schild, M.; Panaiotov, S.; Kurdova-Mintcheva, R.; Gottstein, B.; Walochnik, J.; Aspock, H.; Lucas, M.S.; Muller, N. The identification of free-living environmental isolates of amoebae from Bulgaria. Parasitol. Res. 2004, 92, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Aitken, D.; Hay, J.; Kinnear, F.B.; Kirkness, C.M.; Lee, W.R.; Seal, D.V. Amebic keratitis in a wearer of disposable contact lenses due to a mixed Vahlkampfia and Hartmannella infection. Ophthalmology 1996, 103, 485–494. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Sriram, R.; Qvarnstrom, Y.; Bandyopadhyay, K.; da Silva, A.J.; Pieniazek, N.J.; Cabral, G.A. Paravahlkampfia francinae n. sp. masquerading as an agent of primary amoebic meningoencephalitis. J. Eukaryot. Microbiol. 2009, 56, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Tolba, M.E.M.; Huseein, E.A.M.; Farrag, H.M.M.; Mohamed, H.E.D.; Kobayashi, S.; Suzuki, J.; Ali, T.A.M.; Sugano, S. Allovahlkampfia spelaea causing keratitis in humans. PLoS Negl. Trop. Dis. 2016, 10, e0004841. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.K.; Balne, P.K.; Garg, P.; Sangwan, V.S.; Das, M.; Krishna, P.V.; Bagga, B.; Vemuganti, G.K. Dictyostelium polycephalum infection of human cornea. Emerg. Infect. Dis. 2010, 16, 1644–1645. [Google Scholar] [CrossRef] [PubMed]
- Ashbolt, N.J. Environmental (saprozoic) pathogens of engineered water systems: Understanding their ecology for risk assessment and management. Pathogens 2015, 4, 390–405. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.; Hauroder, B.; Michel, R. Investigations of an extraordinary endocytobiont in Acanthamoeba sp.: Development and replication. Parasitol. Res. 2010, 106, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Fields, B.S.; Barbaree, J.M.; Shotts, E.B., Jr.; Feeley, J.C.; Morrill, W.E.; Sanden, G.N.; Dykstra, M.J. Comparison of guinea pig and protozoan models for determining virulence of Legionella species. Infect. Immun. 1986, 53, 553–559. [Google Scholar] [PubMed]
- Fenner, L.; Richet, H.; Raoult, D.; Papazian, L.; Martin, C.; La Scola, B. Are clinical isolates of Pseudomonas aeruginosa more virulent than hospital environmental isolates in amebal co-culture test? Crit. Care Med. 2006, 34, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Goy, G.; Thomas, V.; Rimann, K.; Jaton, K.; Prod’hom, G.; Greub, G. The Neff strain of Acanthamoeba castellanii, a tool for testing the virulence of Mycobacterium kansasii. Res. Microbiol. 2007, 158, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, K.M.; Levin, B.R. Grazing protozoa and the evolution of the Escherichia coli O157:H7 Shiga toxin-encoding prophage. Proc. Biol. Sci. 2007, 274, 1921–1929. [Google Scholar] [CrossRef] [PubMed]
- Greub, G.; Raoult, D. Microorganisms resistant to free-living amoebae. Clin. Microbiol. Rev. 2004, 17, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.; Lambert, P.A.; Brown, M.R. Influence of intra-amoebic and other growth conditions on the surface properties of Legionella pneumophila. Infect. Immun. 1993, 61, 3503–3510. [Google Scholar] [PubMed]
- Steinert, M.; Birkness, K.; White, E.; Fields, B.; Quinn, F. Mycobacterium avium bacilli grow saprozoically in coculture with Acanthamoeba polyphaga and survive within cyst walls. Appl. Environ. Microbiol. 1998, 64, 2256–2261. [Google Scholar] [PubMed]
- Fritsche, T.R.; Sobek, D.; Gautom, R.K. Enhancement of in vitro cytopathogenicity by Acanthamoeba spp. following acquisition of bacterial endosymbionts. FEMS Microbiol. Lett. 1998, 166, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Molmeret, M.; Horn, M.; Wagner, M.; Santic, M.; Abu Kwaik, Y. Amoebae as training grounds for intracellular bacterial pathogens. Appl. Environ. Microbiol. 2005, 71, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, J.D.; Falkow, S.; Tompkins, L.S.; Bermudez, L.E. Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infect. Immun. 1997, 65, 3759–3767. [Google Scholar] [PubMed]
- Steenbergen, J.N.; Shuman, H.A.; Casadevall, A. Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages. Proc. Natl. Acad. Sci. USA 2001, 98, 15245–15250. [Google Scholar] [CrossRef] [PubMed]
- Brieland, J.K.; Fantone, J.C.; Remick, D.G.; LeGendre, M.; McClain, M.; Engleberg, N.C. The role of Legionella pneumophila-infected Hartmannella vermiformis as an infectious particle in a murine model of Legionaires’ disease. Infect. Immun. 1997, 65, 5330–5333. [Google Scholar] [PubMed]
- Thomas, V.; McDonnell, G.; Denyer, S.P.; Maillard, J.-Y. Free-living amoebae and their intracellular pathogenic microorganisms: Risks for water quality. FEMS Microbiol. Rev. 2010, 34, 231–259. [Google Scholar] [CrossRef] [PubMed]
- Goñi, P.; Fernández, M.T.; Rubio, E. Identifying endosymbiont bacteria associated with free-living amoebae. Environ. Microbiol. 2014, 16, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Reteno, D.G.; Benamar, S.; Khalil, J.B.; Andreani, J.; Armstrong, N.; Klose, T.; Rossmann, M.; Colson, P.; Raoult, D.; La Scola, B. Faustovirus, an asfarvirus-related new lineage of giant viruses infecting amoebae. J. Virol. 2015, 89, 6585–6594. [Google Scholar] [CrossRef] [PubMed]
- Drozanski, W. Fatal bacterial infection in soil amoebae. Acta Microbiol. Pol. 1956, 5, 315–317. [Google Scholar]
- Drozanski, W.J. Sarcobium lyticum gen. nov., sp. nov., an obligate intracellular bacterial parasite of small free-living amoebae. Int. J. System. Bacteriol. 1991, 41, 82–87. [Google Scholar] [CrossRef]
- Adeleke, A.A.; Fields, B.S.; Benson, R.F.; Daneshvar, M.I.; Pruckler, J.M.; Ratcliff, R.M.; Harrison, T.G.; Weyant, R.S.; Birtles, R.J.; Raoult, D.; et al. Legionella drozanskii sp. nov., Legionella rowbothamii sp. nov. and Legionella fallonii sp. nov.: Three unusual new Legionella species. Int. J. Syst. Evol. Microbiol. 2001, 51, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Michel, R.; Muller, K.D.; Amann, R.; Schmid, E.N. Legionella-like slender rods multiplying within a strain of Acanthamoeba sp. isolated from drinking water. Parasitol. Res. 1998, 84, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Rowbotham, T.J. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J. Clin. Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.; Keevil, C.W. Immunogold and fluorescein immunolabelling of Legionella pneumophila within an aquatic biofilm visualized by using episcopic differential interference contrast microscopy. Appl. Environ. Microbiol. 1992, 58, 2326–2330. [Google Scholar] [PubMed]
- Boulanger, C.A.; Edelstein, P.H. Precision and accuracy of recovery of Legionella pneumophila from seeded tap water by filtration and centrifugation. Appl. Environ. Microbiol. 1995, 61, 1805–1809. [Google Scholar] [PubMed]
- Hägele, S.; Kohler, R.; Merkert, H.; Schleicher, M.; Hacker, J.; Steinert, M. Dictyostelium discoideum: A new host model system for intracellular pathogens of the genus Legionella. Cell. Microbiol. 2000, 2, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Hookey, J.V.; Saunders, N.A.; Fry, N.K.; Birtles, R.J.; Harrison, T.G. Phylogeny of Legionellaceae based on small-subunit ribosomal DNA sequences and proposal of Legionella lytica comb. nov. for Legionella-like amoebal pathogens. Int. J. System. Bacteriol. 1996, 46, 526–531. [Google Scholar] [CrossRef]
- Akya, A.; Pointon, A.; Thomas, C. Mechanism involved in phagocytosis and killing of Listeria monocytogenes by Acanthamoeba polyphaga. Parasitol. Res. 2009, 105, 1375–1383. [Google Scholar] [CrossRef] [PubMed]
- Ly, T.M.; Muller, H.E. Ingested Listeria monocytogenes survive and multiply in protozoa. J. Med. Microbiol. 1990, 33, 51–54. [Google Scholar] [CrossRef] [PubMed]
- White, C.I.; Birtles, R.J.; Wigley, P.; Jones, P.H. Mycobacterium avium subspecies paratuberculosis in free-living amoebae isolated from fields not used for grazing. Vet. Rec. 2010, 166, 401–402. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Raoult, D. Afipia felis in hospital water supply in association with free-living amoebae. Lancet 1999, 353, 1330. [Google Scholar] [CrossRef]
- Hoffmann, R.; Michel, R. Distribution of free-living amoebae (FLA) during preparation and supply of drinking water. Int. J. Hyg. Environ. Health. 2001, 203, 215–219. [Google Scholar] [CrossRef]
- Michel, R.; Müller, K.-D.; Schmid, E. Ehrlichia-like organismus (KSL1) observed as obligate intracellular parasites of Saccamoeba species. Endocytobio. Cell Res. 1995, 11, 69–80. [Google Scholar]
- Michel, R. Freilebende Amöben als Wirte und Vehikel von Mikroorganismen. Mitteilungen der Österreichischen Gesellschaft für Tropenmedizin und Parasitologie 1997, 19, 11–20. [Google Scholar]
- Corsaro, D.; Muller, K.-D.; Michel, R. Molecular characterization and ultrastructure of a new amoeba endoparasite belonging to the Stenotrophomonas maltophilia complex. Exp. Parasitol. 2013, 133, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Kahane, S.; Dvoskin, B.; Mathias, M.; Friedman, M.G. Infection of Acanthamoeba polyphaga with Simkania negevensis and S. negevensis survival within amoebal cysts. Appl. Environ. Microbiol. 2001, 67, 4789–4795. [Google Scholar] [CrossRef] [PubMed]
- Michel, R.; Muller, K.-D.; Zöller, L.; Walochnik, J.; Hartmann, M.; Schmid, E. Free living amoebae serve as host for the Chlamydia-like bacterium Simkania nevegensis. Acta Protozool. 2005, 44, 113–121. [Google Scholar]
- Hennig, K.; Zöller, L.; Hauröder, B.; Hotzel, H.; Michel, R. Hartmannella vermiformis (Hartmannellidae) harboured a hidden Chlamydia-like endosymbiont. Endocytobio. Cell Res. 2007, 18, 1–10. [Google Scholar]
- Horn, M.; Wagner, M.; Muller, K.D.; Schmid, E.N.; Fritsche, T.R.; Schleifer, K.H.; Michel, R. Neochlamydia hartmannellae gen. nov., sp. nov. (Parachlamydiaceae), an endoparasite of the amoeba Hartmannella vermiformis. Microbiology 2000, 146, 1231–1239. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, D.; Müller, K.D.; Wingender, J.; Michel, R. “Candidatus Mesochlamydia elodeae” (Chlamydiae: Parachlamydiaceae), a novel chlamydia parasite of free-living amoebae. Parasitol. Res. 2013, 112, 829–1838. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Fritsche, T.R.; Gautom, R.K.; Schleifer, K.H.; Wagner, M. Novel bacterial endosymbionts of Acanthamoeba spp. related to the Paramecium caudatum symbiont Caedibacter caryophilus. Environ. Microbiol. 1999, 1, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Fritsche, T.R.; Linner, T.; Gautom, R.K.; Harzenetter, M.D.; Wagner, M. Obligate bacterial endosymbionts of Acanthamoeba spp. related to the beta-Proteobacteria: Proposal of ‘Candidatus Procabacter acanthamoebae’ gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Horn, M.; Harzenetter, M.D.; Linner, T.; Schmid, E.N.; Muller, K.D.; Michel, R.; Wagner, M. Members of the Cytophaga-Flavobacterium-Bacteroides phylum as intracellular bacteria of acanthamoebae: Proposal of ‘Candidatus Amoebophilus asiaticus’. Environ. Microbiol. 2001, 3, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Gaze, W.H.; Burroughs, N.; Gallagher, M.P.; Wellington, E.M.H. Interactions between Salmonella typhimurium and Acanthamoeba polyphaga, and observation of a new mode of intracellular growth within contractile vacuoles. Microb. Ecol. 2003, 46, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Tezcan-Merdol, D.; Ljungstrom, M.; Winiecka-Krusnell, J.; Linder, E.; Engstrand, L.; Rhen, M. Uptake and replication of Salmonella enterica in Acanthamoeba rhysodes. Appl. Environ. Microbiol. 2004, 70, 3706–3714. [Google Scholar] [CrossRef] [PubMed]
- King, C.H.; Shotts, E.B., Jr.; Wooley, R.E.; Porter, K.G. Survival of coliforms and bacterial pathogens within protozoa during chlorination. Appl. Environ. Microbiol. 1988, 54, 3023–3033. [Google Scholar] [PubMed]
- Snelling, W.J.; McKenna, J.P.; Lecky, D.M.; Dooley, J.S.G. Survival of Campylobacter jejuni in waterborne protozoa. Appl. Environ. Microbiol. 2005, 71, 5560–5571. [Google Scholar] [CrossRef] [PubMed]
- Medina, G.; Flores-Martin, S.; Fonseca, B.; Otth, C.; Fernandez, H. Mechanisms associated with phagocytosis of Arcobacter butzleri by Acanthamoeba castellanii. Parasitol. Res. 2014, 113, 1933–1942. [Google Scholar] [CrossRef] [PubMed]
- Fernández, H.; Villanueva, M.P.; Medina, G. Endosimbiosis de Arcobacter butzleri en Acanthamoeba castellanii. Rev. Argent. Microbiol. 2012, 44, 133. [Google Scholar] [PubMed]
- Mella, C.; Medina, G.; Flores-Martin, S.; Toledo, Z.; Simaluiza, R.J.; Péréz-Péréz, G.; Fernández, H. Interaction between zoonotic bacteria and free living amoebas. A new angle of an epidemiological polyhedron of public health importance? Archivos de Medicina Veterinaria 2016, 48, 1–10. [Google Scholar] [CrossRef]
- Jadin, J.; Francois, J.; Bisoux, M.; Languillon, J.; Moris, R. Dévelopment intranucléaire de Mycobacterium leprae dans les cellules histiocytaires chez l‘animal. Bull. Acad. Natl. Med. 1968, 152, 89–91. [Google Scholar] [PubMed]
- Krishna-Prasad, B.; Gupta, S. Preliminary report on engulfment and retention of mycobacteria by trophozoites of axenically grown Acanthamoeba castellanii Douglas. Curr. Sci. 1978, 47, 245–247. [Google Scholar]
- Alsam, S.; Khan, N. (Eds.) Methicillin resistant Staphylococcus aureus interactions with Acanthamoeba. In Proceedings of the Booklet Free Living Amoebae (FLAM) Meeting, Tenerife Spain, May 2009. [Google Scholar]
- Tomov, A.T.; Tsvetkova, E.D.; Tomova, I.A.; Michailova, L.I.; Kassovski, V.K. Persistence and multiplication of obligate anaerobe bacteria in amebae under aerobic conditions. Anaerobe 1999, 5, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Drancourt, M.; Adekambi, T.; Raoult, D. Interactions between Mycobacterium xenopi, amoeba and human cells. J. Hosp. Infect. 2007, 65, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Dey, R.; Hoffman, P.S.; Glomski, I.J. Germination and amplification of anthrax spores by soil-dwelling amoebas. Appl. Environ. Microbiol. 2012, 78, 8075–8081. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.; Loret, J.-F.; Jousset, M.; Greub, G. Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant. Environ. Microbiol. 2008, 10, 2728–2745. [Google Scholar] [CrossRef] [PubMed]
- Landers, P.; Kerr, K.G.; Rowbotham, T.J.; Tipper, J.L.; Keig, P.M.; Ingham, E.; Denton, M. Survival and growth of Burkholderia cepacia within the free-living amoeba Acanthamoeba polyphaga. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Barker, J.; Humphrey, T.J.; Brown, M.W. Survival of Escherichia coli O157 in a soil protozoan: Implications for disease. FEMS Microbiol. Lett. 1999, 173, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Winiecka-Krusnell, J.; Wreiber, K.; von Euler, A.; Engstrand, L.; Linder, E. Free-living amoebae promote growth and survival of Helicobacter pylori. Scand. J. Infect. Dis. 2002, 34, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Wagner, Y.; Noack, B.; Hoffmann, T.; Jacobs, E.; Christian Luck, P. Periodontopathogenic bacteria multiply in the environmental amoeba Acanthamoeba castellani. Int. J. Hyg. Environ. Health. 2006, 209, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Essig, A.; Heinemann, M.; Simnacher, U.; Marre, R. Infection of Acanthamoeba castellanii by Chlamydia pneumoniae. Appl. Environ. Microbiol. 1997, 63, 1396–1399. [Google Scholar] [PubMed]
- Michel, R.; Hauröder-Philippczyk, B.; Müller, K.-D.; Weishaar, I. Acanthamoeba from human nasal mucosa infected with an obligate intracellular parasite. Europ. J. Protistol. 1994, 30, 104–110. [Google Scholar] [CrossRef]
- Amann, R.; Springer, N.; Schonhuber, W.; Ludwig, W.; Schmid, E.N.; Muller, K.D.; Michel, R. Obligate intracellular bacterial parasites of acanthamoebae related to Chlamydia spp. Appl. Environ. Microbiol. 1997, 63, 115–121. [Google Scholar] [PubMed]
- Everett, K.D.; Bush, R.M.; Andersen, A.A. Emended description of the order Chlamydiales, proposal of Parachlamydiaceae fam. nov. and Simkaniaceae fam. nov., each containing one monotypic genus, revised taxonomy of the family Chlamydiaceae, including a new genus and five new species, and standards for the identification of organisms. Int. J. System. Bacteriol. 1999, 49 Pt 2, 415–440. [Google Scholar]
- Michel, R.; Muller, K.D.; Hoffmann, R. Enlarged Chlamydia-like organisms as spontaneous infection of Acanthamoeba castellanii. Parasitol. Res. 2001, 87, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Michel, R.; Steinert, M.; Zöller, L.; Hauröder, B.; Hennig, K. Cocultivation of protozoa and the Chlamydia-like bacterium Waddlia chondrophila isolated from an aborted bovine foetus in Germany. Acta Protozool. 2004, 43, 37–42. [Google Scholar]
- Collingro, A.; Poppert, S.; Heinz, E.; Schmitz-Esser, S.; Essig, A.; Schweikert, M.; Wagner, M.; Horn, M. Recovery of an environmental Chlamydia strain from activated sludge by co-cultivation with Acanthamoeba sp. Microbiology 2005, 151, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Heinz, E.; Kolarov, I.; Kastner, C.; Toenshoff, E.R.; Wagner, M.; Horn, M. An Acanthamoeba sp. containing two phylogenetically different bacterial endosymbionts. Environ. Microbiol. 2007, 9, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Raoult, D. Survival of Coxiella burnetii within free-living amoeba Acanthamoeba castellanii. Clin. Microbiol. Infect. 2001, 7, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, K. Growth and survival of four strains of Francisella tularensis in a rich medium preconditioned with Acanthamoeba palestinensis. Can. J. Microbiol. 1989, 35, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.; Schwarzenberger, R.; Schacht, E.; Splettstoesser, W. Free-living amoebae as hosts and vehicles of Francisella sp. In Proceedings of the Medical Biodefense conference, Medical Corps International Forum 2011. Munich, Germany, 2011. [Google Scholar]
- Abd, H.; Johansson, T.; Golovliov, I.; Sandstrom, G.; Forsman, M. Survival and growth of Francisella tularensis in Acanthamoeba castellanii. Appl. Environ. Microbiol. 2003, 69, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Abd, H. Interaction between waterborne pathogenic bacteria and Acanthamoeba castellanii; Dissertation: Stockholm, Sweden, 2006. [Google Scholar]
- Corsaro, D.; Walochnik, J.; Venditti, D.; Steinmann, J.; Muller, K.-D.; Michel, R. Microsporidia-like parasites of amoebae belong to the early fungal lineage Rozellomycota. Parasitol. Res. 2014, 113, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Koehsler, M.; Walochnik, J.; Michel, R.; Lugauer, J.; Wylezich, C. Molecular identification and classification of Cochlonema euryblastum, a zoopagalean parasite of Thecamoeba quadrilineata. Mycologia 2007, 99, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.L.; Schwarzenberger, R. Free-living amoebae as vectors of cryptosporidia. Parasitol. Res. 2011, 109, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Zahedi, A.; Paparini, A. Cryptosporidium in humans and animals-a one health approach to prophylaxis. Parasite Immunol. 2016, 38, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Couso, H.; Paniagua-Crespo, E.; Ares-Mazas, E. Acanthamoeba as a temporal vehicle of Cryptosporidium. Parasitol. Res. 2007, 100, 1151–1154. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; Coronado-Alvarez, N.; Martinez-Carretero, E.; Maciver, S.K.; Valladares, B. Detection of four adenovirus serotypes within water-isolated strains of Acanthamoeba in the Canary Islands, Spain. Am. J. Trop. Med. Hyg. 2007, 77, 753–756. [Google Scholar] [PubMed]
- La Scola, B.; Audic, S.; Robert, C.; Jungang, L.; de Lamballerie, X.; Drancourt, M.; Birtles, R.; Claverie, J.-M.; Raoult, D. A giant virus in amoebae. Science 2003, 299, 2033. [Google Scholar] [PubMed]
- Hoffmann, R.; Michel, R.; Muller, K.D.; Schmid, E. Archaea-like endocytobiotic organisms isolated from Acanthamoeba sp. (Gr II). Endocytobio. Cell Res. 1998, 12, 185–188. [Google Scholar]
- Scheid, P.; Schwarzenberger, R. Acanthamoeba spp. as vehicle and reservoir of adenoviruses. Parasitol. Res. 2012, 111, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Mattana, A.; Serra, C.; Mariotti, E.; Delogu, G.; Fiori, P.L.; Cappuccinelli, P. Acanthamoeba castellanii promotion of in vitro survival and transmission of coxsackie b3 viruses. Eukaryotic Cell 2006, 5, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Boyer, M.; Yutin, N.; Pagnier, I.; Barrassi, L.; Fournous, G.; Espinosa, L.; Robert, C.; Azza, S.; Sun, S.; Rossmann, M.G.; et al. Giant Marseillevirus highlights the role of amoebae as a melting pot in emergence of chimeric microorganisms. Proc. Natl. Acad. Sci. USA 2009, 106, 21848–21853. [Google Scholar] [CrossRef] [PubMed]
- Arslan, D.; Legendre, M.; Seltzer, V.; Abergel, C.; Claverie, J.-M. Distant Mimivirus relative with a larger genome highlights the fundamental features of Megaviridae. Proc. Natl. Acad. Sci. USA 2011, 108, 17486–17491. [Google Scholar] [CrossRef] [PubMed]
- Philippe, N.; Legendre, M.; Doutre, G.; Coute, Y.; Poirot, O.; Lescot, M.; Arslan, D.; Seltzer, V.; Bertaux, L.; Bruley, C.; et al. Pandoraviruses: Amoeba viruses with genomes up to 2.5 Mb reaching that of parasitic eukaryotes. Science 2013, 341, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Legendre, M.; Bartoli, J.; Shmakova, L.; Jeudy, S.; Labadie, K.; Adrait, A.; Lescot, M.; Poirot, O.; Bertaux, L.; Bruley, C.; et al. Thirty-thousand-year-old distant relative of giant icosahedral DNA viruses with a pandoravirus morphology. Proc. Natl. Acad. Sci. USA 2014, 111, 4274–4279. [Google Scholar] [CrossRef] [PubMed]
- Legendre, M.; Lartigue, A.; Bertaux, L.; Jeudy, S.; Bartoli, J.; Lescot, M.; Alempic, J.-M.; Ramus, C.; Bruley, C.; Labadie, K.; et al. In-depth study of Mollivirus sibericum, a new 30,000-y-old giant virus infecting Acanthamoeba. Proc. Natl. Acad. Sci. USA 2015, 112, E5327–E5335. [Google Scholar] [CrossRef] [PubMed]
- Ecke, D.J.; Sampath, R.; Willett, P.; Samant, V.; Massire, C.; Hall, T.A.; Hari, K.; McNeil, J.A.; Buchen-Osmond, C.; Budowle, B. The Microbial Rosetta Stone database: A common structure for microbial biosecurity threat agents. J. Forensic Sci. 2005, 50, 1380–1385. [Google Scholar] [PubMed]
- Adékambi, T.; Ben Salah, S.; Khlif, M.; Raoult, D.; Drancourt, M. Survival of environmental mycobacteria in Acanthamoeba polyphaga. Appl. Environ. Microbiol. 2006, 72, 5974–5981. [Google Scholar] [CrossRef] [PubMed]
- Thomas, V.; McDonnell, G. Relationship between mycobacteria and amoebae: Ecological and epidemiological concerns. Lett. Appl. Microbiol. 2007, 45, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Kilvington, S.; Price, J. Survival of Legionella pneumophila within cysts of Acanthamoeba polyphaga following chlorine exposure. J. Appl. Bacteriol. 1990, 68, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Fallon, R.J.; Rowbotham, T.J. Microbiological investigations into an outbreak of Pontiac fever due to Legionella micdadei associated with use of a whirlpool. J. Clin. Pathol. 1990, 43, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Thom, S.; Warhurst, D.; Drasar, B.S. Association of Vibrio cholerae with fresh water amoebae. J. Med. Microbiol. 1992, 36, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Howard, K.; Inglis, T.J.J. Disinfection of Burkholderia pseudomallei in potable water. Water Res. 2005, 39, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Bruggemann, H.; Hagman, A.; Jules, M.; Sismeiro, O.; Dillies, M.-A.; Gouyette, C.; Kunst, F.; Steinert, M.; Heuner, K.; Coppee, J.-Y.; et al. Virulence strategies for infecting phagocytes deduced from the in vivo transcriptional program of Legionella pneumophila. Cell. Microbiol. 2006, 8, 1228–1240. [Google Scholar] [CrossRef] [PubMed]
- Steinert, M.; Emody, L.; Amann, R.; Hacker, J. Resuscitation of viable but nonculturable Legionella pneumophila Philadelphia JR32 by Acanthamoeba castellanii. Appl. Environ. Microbiol. 1997, 63, 2047–2053. [Google Scholar] [PubMed]
- Barker, J.; Brown, M.R.; Collier, P.J.; Farrell, I.; Gilbert, P. Relationship between Legionella pneumophila and Acanthamoeba polyphaga: Physiological status and susceptibility to chemical inactivation. Appl. Environ. Microbiol. 1992, 58, 2420–2425. [Google Scholar] [PubMed]
- Axelsson-Olsson, D.; Waldenstrom, J.; Broman, T.; Olsen, B.; Holmberg, M. Protozoan Acanthamoeba polyphaga as a potential reservoir for Campylobacter jejuni. Appl. Environ. Microbiol. 2005, 71, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Axelsson-Olsson, D.; Svensson, L.; Olofsson, J.; Salomon, P.; Waldenstrom, J.; Ellstrom, P.; Olsen, B. Increase in acid tolerance of Campylobacter jejuni through coincubation with amoebae. Appl. Environ. Microbiol. 2010, 76, 4194–4200. [Google Scholar] [CrossRef] [PubMed]
- Snelling, W.J.; Stern, N.J.; Lowery, C.J.; Moore, J.E.; Gibbons, E.; Baker, C.; Dooley, J.S.G. Colonization of broilers by Campylobacter jejuni internalized within Acanthamoeba castellanii. Arch. Microbiol. 2008, 189, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Henkle, E.; Winthrop, K.L. Nontuberculous mycobacteria infections in immunosuppressed hosts. Clin. Chest. Med. 2015, 36, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, M.P.; Medina, G.; Fernandez, H. Arcobacter butzleri survives within trophozoite of Acanthamoeba castellanii. Rev. Argent. Microbiol. 2016, 48, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, E.; Bare, J.; van Damme, I.; Bert, W.; Sabbe, K.; Houf, K. Behavior of Yersinia enterocolitica in the presence of the bacterivorous Acanthamoeba castellanii. Appl. Environ. Microbiol. 2013, 79, 6407–6413. [Google Scholar] [CrossRef] [PubMed]
- Bleasdale, B.; Lott, P.J.; Jagannathan, A.; Stevens, M.P.; Birtles, R.J.; Wigley, P. The Salmonella pathogenicity island 2-encoded type III secretion system is essential for the survival of Salmonella enterica serovar Typhimurium in free-living amoebae. Appl. Environ. Microbiol. 2009, 75, 1793–1795. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.A.; Sharma, V.K.; McCuddin, Z.P.; Rasmussen, M.A.; Franklin, S.K. Involvement of a Salmonella genomic island 1 gene in the rumen protozoan-mediated enhancement of invasion for multiple-antibiotic-resistant Salmonella enterica serovar Typhimurium. Infect. Immun. 2007, 75, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.T.; Xiong, N.; Dier, J.D.; Anderson, K.L.; Rasmussen, M.A.; Franklin, S.K.; Carlson, S.A. Comparisons of Salmonella conjugation and virulence gene hyperexpression mediated by rumen protozoa from domestic and exotic ruminants. Vet. Microbiol. 2011, 151, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.A.; Carlson, S.A.; Franklin, S.K.; McCuddin, Z.P.; Wu, M.T.; Sharma, V.K. Exposure to rumen protozoa leads to enhancement of pathogenicity of and invasion by multiple-antibiotic-resistant Salmonella enterica bearing SGI1. Infect. Immun. 2005, 73, 4668–4675. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Raoult, D. Gene repertoire of amoeba-associated giant viruses. Intervirology 2010, 53, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; La Scola, B.; Birtles, R. The discovery and characterization of Mimivirus, the largest known virus and putative pneumonia agent. Clin. Infect. Dis. 2007, 45, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Schuster, F.L.; Dunnebacke, T.H. Virus-like particles and an unassociated infectious agent in amoebae of the genus Naegleria. Ann. Soc. Belge Méd. trop. 1974, 54, 359–370. [Google Scholar]
- Suzan-Monti, M.; La Scola, B.; Raoult, D. Genomic and evolutionary aspects of Mimivirus. Virus Res. 2006, 117, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Aherfi, S.; Colson, P.; La Scola, B.; Raoult, D. Giant viruses of amoebas: An update. Front. Microbiol. 2016, 7, 349. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; Pagnier, I.; Yoosuf, N.; Fournous, G.; La Scola, B.; Raoult, D. “Marseilleviridae”, a new family of giant viruses infecting amoebae. Arch. Virol. 2013, 158, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; de Lamballerie, X.; Fournous, G.; Raoult, D. Reclassification of giant viruses composing a fourth domain of life in the new order Megavirales. Intervirology 2012, 55, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P. A strange endocytobiont revealed as largest virus. Curr. Opin. Microbiol. 2016, 31, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P. Viruses in close associations with free-living amoebae. Parasitol. Res. 2015, 114, 3959–3967. [Google Scholar] [CrossRef] [PubMed]
- Scheid, P.; Balczun, C.; Schaub, G.A. Some secrets are revealed: Parasitic keratitis amoebae as vectors of the scarcely described pandoraviruses to humans. Parasitol. Res. 2014, 113, 3759–3764. [Google Scholar] [CrossRef] [PubMed]
- Antwerpen, M.H.; Georgi, E.; Zoeller, L.; Woelfel, R.; Stoecker, K.; Scheid, P. Whole-genome sequencing of a Pandoravirus isolated from keratitis-inducing acanthamoeba. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Abergel, C.; Legendre, M.; Claverie, J.-M. The rapidly expanding universe of giant viruses: Mimivirus, Pandoravirus, Pithovirus and Mollivirus. FEMS Microbiol. Rev. 2015, 39, 779–796. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Colson, P.; Chabrol, O.; Pontarotti, P.; Raoult, D. Pithovirus sibericum, a new bona fide member of the “Fourth TRUC” club. Front. Microbiol. 2015, 6, 722. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.C.F.; Almeida, G.M.F.; Oliveira, D.B.; Dornas, F.P.; Campos, R.K.; La Scola, B.; Ferreira, P.C.P.; Kroon, E.G.; Abrahao, J.S. A resourceful giant: APMV is able to interfere with the human type I interferon system. Microbes Infect. 2014, 16, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Colson, P.; La Scola, B.; Raoult, D. Giant viruses of amoebae as potential human pathogens. Intervirology 2013, 56, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Margulis, L.; Schwartz, K.V. Die fünf Reiche der Organismen: Ein Leitfaden; Spektrum d. Wiss: Heidelberg, Germany, 1989. [Google Scholar]
- Desnues, C.; La Scola, B.; Yutin, N.; Fournous, G.; Robert, C.; Azza, S.; Jardot, P.; Monteil, S.; Campocasso, A.; Koonin, E.V.; et al. Provirophages and transpovirons as the diverse mobilome of giant viruses. Proc. Natl. Acad. Sci. USA 2012, 109, 18078–18083. [Google Scholar] [CrossRef] [PubMed]
- Boyer, M.; Azza, S.; Barrassi, L.; Klose, T.; Campocasso, A.; Pagnier, I.; Fournous, G.; Borg, A.; Robert, C.; Zhang, X.; et al. Mimivirus shows dramatic genome reduction after intraamoebal culture. Proc. Natl. Acad. Sci. USA 2011, 108, 10296–10301. [Google Scholar] [CrossRef] [PubMed]
- Maumus, F.; Blanc, G. Study of gene trafficking between Acanthamoeba and giant viruses suggests an undiscovered family of amoeba-infecting viruses. Genome Biol. Evol. 2016, 8, 3351–3363. [Google Scholar] [CrossRef] [PubMed]
- Verneau, J.; Levasseur, A.; Raoult, D.; La Scola, B.; Colson, P. MG-Digger: An automated pipeline to search for giant virus-related sequences in metagenomes. Front. Microbiol. 2016, 7, 428. [Google Scholar] [CrossRef] [PubMed]
- Winiecka-Krusnell, J.; Dellacasa-Lindberg, I.; Dubey, J.P.; Barragan, A. Toxoplasma gondii: Uptake and survival of oocysts in free-living amoebae. Exp. Parasitol. 2009, 121, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Smith, L.M.; May, R.C. New weapons in the Cryptococcus infection toolkit. Curr. Opin. Microbiol. 2016, 34, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, J.N.; Nosanchuk, J.D.; Malliaris, S.D.; Casadevall, A. Cryptococcus neoformans virulence is enhanced after growth in the genetically malleable host Dictyostelium discoideum. Infect. Immun. 2003, 71, 4862–4872. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Casadevall, A.; Ausubel, F.M. Exploiting amoeboid and non-vertebrate animal model systems to study the virulence of human pathogenic fungi. PLoS Pathog. 2007, 3, e101. [Google Scholar] [CrossRef] [PubMed]
- Cursons, R.T.; Brown, T.J.; Keys, E.A. Effect of disinfectants on pathogenic free-living amoebae: In axenic conditions. Appl. Environ. Microbiol. 1980, 40, 62–66. [Google Scholar] [PubMed]
- Hiti, K.; Walochnik, J.; Haller-Schober, E.M.; Faschinger, C.; Aspock, H. Viability of Acanthamoeba after exposure to a multipurpose disinfecting contact lens solution and two hydrogen peroxide systems. Br. J. Ophthalmol. 2002, 86, 144–146. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Transmission electron microscopic picture of Pandoravirus inopinatum by negative staining contrast. The bar in (A): 100 nm, and the bar in (B): 250 nm (staining performed by Dimitrios Frangoulidis and Claudia Kahlhofer; Bundeswehr Institute of Microbiology, Munich).
Figure 1.
Transmission electron microscopic picture of Pandoravirus inopinatum by negative staining contrast. The bar in (A): 100 nm, and the bar in (B): 250 nm (staining performed by Dimitrios Frangoulidis and Claudia Kahlhofer; Bundeswehr Institute of Microbiology, Munich).

{kind=link}
Table 1.
Selection of microorganisms interacting with free-living amoebae (FLA), including pathogens with public health importance.
Table 1.
Selection of microorganisms interacting with free-living amoebae (FLA), including pathogens with public health importance.
| Species | Host Amoeba | References (Selection) |
|---|---|---|
| BACTERIA | ||
| In Situ Interaction | ||
| Legionella-like, Legionella lytica (Sarcobium lyticum) | Acanthamoebae, Naegleriae, Hartmanellae | [47,48,49,50] |
| Legionella pneumophila | Several FLA, e.g., Acanthamoebae | [51,52,53,54,55] |
| Listeria monocytogenes | Acanthamoebae | [56,57] |
| Mycobacterium avium | Acanthamoebae | [41,58] |
| Afipia felis | Acanthamoebae | [59] |
| Pseudomonas saccharophilia | Acanthamoebae | [60] |
| Pseudomonas aeruginosa | Echinamoebae; Acanthamoebae; Hartmannellae | [61,62] |
| Ralstonia (Burkholderia) picketti | Acanthamoebae | [62] |
| Stenotrophomonas maltophilia-Komplex | Acanthamoebae; Naegleriae | [63] |
| Rickettsia-like | Acanthamoebae | [39] |
| Simkania nevegensis (Chlamydia-like) | Acanthamoebae, Naegleriae, Hartmannellae; Balamuthia mandrillaris | [64,65,66] |
| Neochlamydia hartmannellae | Hartmannellae; Hartmannella (Vermamoeba) vermiformis | [67] |
| Candidatus Mesochlamydia elodeae | Vannellae, Saccamoebae | [68] |
| Ehrlichia-like | Saccamoebae | [61] |
| Candidatus Paracaedibacter symbiosus | Acanthamoebae | [69] |
| Procabacter acanthamoebae (Betaproteobacterium) | Acanthamoebae | [70] |
| Amoebophilus asiaticus | Acanthamoebae | [71] |
| In Vitro Interaction | ||
| Salmonella enterica, S. thyphimurium (and other Salmonellae) | Acanthamoebae | [72,73] |
| Yersinia enterocolitica | Acanthamoebae | [74] |
| Campylobacter jejuni | Acanthamoebae (Acanthamoeba polyphaga) | [75] |
| Arcobacter butzleri | Acanthamoebae (Acanthamoeba castellanii) | [76,77,78] |
| Mycobacterium spp., M. leprae | Acanthamoebae | [38,79,80] |
| Staphylococcus aureus (incl.MRSA) | Acanthamoebae | [81] |
| Mobiluncus curtisii | Acanthamoebae | [82] |
| Mycobacterium leprae | Acanthamoebae | [79] |
| Mycobacterium xenopi | Acanthamoebae | [83] |
| Mycobacterium spp; Mycobacterium avium | Acanthamoebae | [38,79,80] |
| Opportunistic Mycobacteria | Acanthamoebae | [41,80] |
| Bacillus anthracis | Acanthamoeba castellanii Hartmannella (Vermamoeba) vermiformis | [84] |
| Acidovorax temperans | Naegleriae | [85] |
| Burkholderia cepacia and Burkholderia pickettii | Acanthamoebae | [86] |
| Caulobacter vibrioides | Echinamoebae | [85] |
| Flavobacterium johnsoniae | Naegleriae | [85] |
| Escherichia coli O157 | Acanthamoebae | [87] |
| Helicobacter pylori | Acanthamoebae | [88] |
| Porphyromonas gingivalis | Acanthamoebae | [89] |
| Prevotella intermedia | Acanthamoebae | [89] |
| Chlamydophila pneumoniae | Acanthamoebae | [90] |
| Parachlamydia acanthamoebae | Acanthamoebae | [91,92,93] |
| Waddlia sp., Waddlia chondrophila, other Chlamydia-like endocytobionts | Acanthamoebae, Naegleriae, Hartmannellae, Hyperamoebae, Vahlkampfiae, Dictyostelium discoideum | [85,94,95] |
| Other Parachlamydia-like endocytobionts | Acanthamoebae | [96,97] |
| Coxiella burnetii | Acanthamoebae | [98] |
| Francisella tularensis | Acanthamoebae | [99,100,101,102] |
| PROTISTS AND FUNGI | ||
| In Situ Interaction | ||
| Paramicrosporidium gen. nov | Vannella sp. | [103] |
| Cochlonema euryblastum | Thecamoebae; Thecamoeba quadrilineata | [104] |
| In Vitro Interaction | ||
| Cryptosporidium parvum | Acanthamoebae | [105,106,107] |
| Cryptococcus neoformans | Acanthamoebae | [42] |
| VIRUSES | ||
| In Situ Interaction | ||
| Adenoviridae | Acanthamoebae | [108] |
| Acanthamoeba polyphaga Mimivirus | Acanthamoebae | [109] |
| Pandoravirus inopinatum | Acanthamoebae | [12,31] |
| Pithovirus | Acanthamoebae | [110] |
| In Vitro Interaction | ||
| Adenoviridae | Acanthamoebae | [111] |
| Coxackie virus | Acanthamoeba | [112] |
| Marseillevirus | Acanthamoebae | [113] |
| Megavirus chilensis | Acanthamoebae | [114] |
| Pandoravirus salinus, Pandoravirus dulcis | Acanthamoebae | [115] |
| Pithovirus sibericum | Acanthamoebae | [116] |
| Faustovirus | Vermamoeba vermiformis | [46] |
| Mollivirus sibericum | Acanthamoebae | [117] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balczun, C.; Scheid, P.L. Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance. Viruses 2017, 9, 65. https://doi.org/10.3390/v9040065
AMA Style
Balczun C, Scheid PL. Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance. Viruses. 2017; 9(4):65. https://doi.org/10.3390/v9040065
Chicago/Turabian StyleBalczun, Carsten, and Patrick L. Scheid. 2017. "Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance" Viruses 9, no. 4: 65. https://doi.org/10.3390/v9040065
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.