1. Introduction
Marek’s disease (MD) is a lymphoproliferative disease in chickens that has caused considerable economic losses to the commercial poultry industry worldwide. The MD virus (MDV) is the etiological agent of MD. It is an oncogenic alpha-herpesvirus that belongs to the genus
Mardivirus and G
allid herpesviruses species, which includes G
allid herpesvirus type 2 (GaHV-2; represented as MDV in this manuscript), GaHV-3, and M
eleagrid herpesvirus type 1 (MeHV-1; represented as HVT in this manuscript) [
1]. However, GaHV-3 and MeHV-1 are non-oncogenic, with only MDV causing the disease in susceptible hosts. Moreover, MDV can be further classified into four pathotypes, including mild (m), virulent (v), very virulent (vv), and very virulent plus (vv+) MDV strains, based on the induction of MD lesions in unvaccinated and vaccinated chickens, in line with the MDV pathotyping assay developed at the Avian Disease and Oncology Laboratory (ADOL) of the United States Department of Agriculture [
2,
3]. The virulence and vaccine resistance of the MDV strains are generally associated, i.e., the more virulent MDV strains are more likely to induce more serious MD lesions in susceptible chickens, with the associated vaccines less likely to provide protection.
Vaccinations are the primary approach used for controlling MD in chickens, and three types of vaccines have been developed against MD, including HVT, nonpathogenic GaHV-3, and attenuated GaHV-2 [
4,
5,
6]. MD has been successfully controlled by vaccination, significantly reducing economic losses to the domestic poultry industry [
7,
8]. However, the vaccines used for MD control cannot induce sterile immunity and allow for replication and transmission of virulent MDV strains in a live host, leading to complicated interactions between the pathogens, vaccines, and hosts [
9,
10,
11,
12]. Thus, MDV has continued to evolve and obtain enhanced virulence over the last few decades owing to the widespread use of these vaccines. The relationship between increased MDV virulence and the introduction of different vaccines was summarized as a step-wise evolution, in which MDV strains have shown continuous evolution to maintain virulence, acquiring the ability to overcome immune responses induced by vaccines [
13]. Successive generations of MD vaccines have been introduced to protect birds from increasingly virulent MDV strains; however, the virus has countered each new vaccine.
In China, MDV was first reported in the 1970s. At present, laying and breeding chickens are vaccinated with vaccine CVI988 (an effective attenuated GaHV-2 vaccine used commercially worldwide), 814 (an effective attenuated GaHV-2 vaccine used widely in China), or a bivalent vaccine (CVI988 plus HVT) via subcutaneous or intramuscular injection at the time of hatching in the vast majority of Chinese poultry enterprises. The economical HVT vaccine is normally used to vaccinate commercial meat-producing chickens, which are typically raised for only 7–8 weeks and have a lower risk for MD. Although the use of vaccines has effectively controlled MDV, MD vaccination failure cases have occasionally occurred, although little is known about the causes of vaccination failure.
In this study, we isolated a field MDV strain BS/15 from severely diseased chickens that were vaccinated with the commercial vaccine CVI988. Furthermore, the pathogenicity of BS/15 to specific pathogen-free (SPF) chickens was analyzed, and the protective efficacy of vaccine CVI988 and vaccine 814 against BS/15 was evaluated. Our study was created in order to increase understanding of the pathogenic characteristics of the newly isolated MDV strain, the causes of MD vaccination failure, and the evolution of MDV, as well as providing guidance for MD control.
2. Materials and Methods
2.1. Collection of Clinical Samples
A layer flock in Baishan City of the Jilin Province in China developed serious MD in 2015. Chickens had been vaccinated with the commercial vaccine CVI988 at 1 day of age, and clinical symptoms of MD were observed beginning at about 55 days of age. The incidence of MD in the chicken flocks quickly reached about 36% by 120 days of age. Visible tumors were frequently found in the visceral organs of dead or diseased chickens by postmortem examination. For molecular diagnosis and viral isolation, feather pulps were collected from the chickens with suspected MD.
2.2. Viral Isolation and Identification
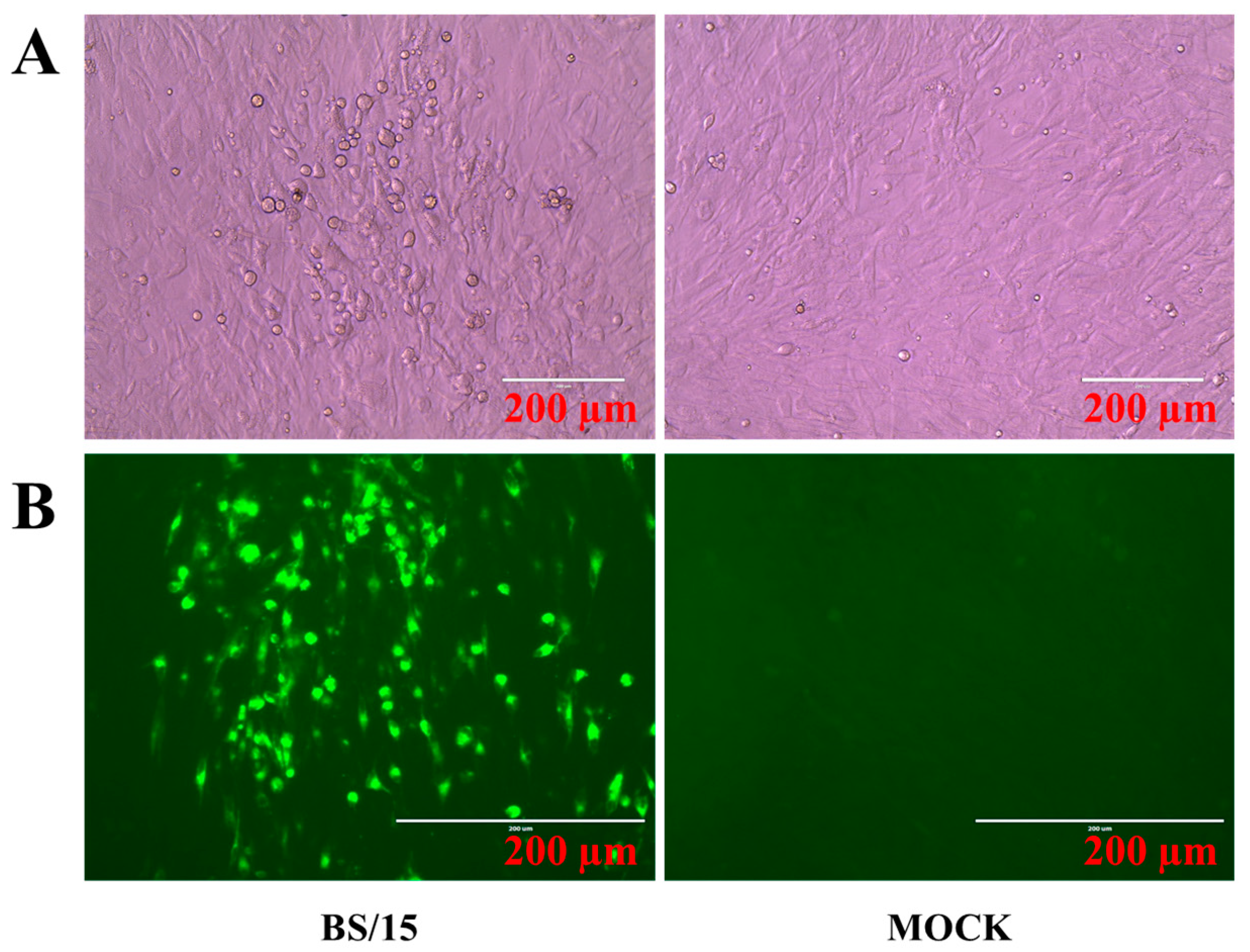
Feather pulps collected from the diseased chickens described above were used for viral isolation, as previously described [
14]. Briefly, SPF duck or chicken embryos were obtained from the Harbin Veterinary Research Institute (HVRI), Chinese Academy of Agricultural Sciences (CAAS) for the preparation of duck embryo fibroblasts (DEFs) or chicken embryo fibroblasts (CEFs). Feather pulp-inoculated primary DEFs were incubated at 37 °C in an atmosphere containing 5% CO
2, and blind passages were performed until cytopathogenic effects (CPEs) were observed. Viruses from infected cells with CPEs were plaque- purified in DEFs, and then the viruses were propagated and stored in liquid nitrogen.
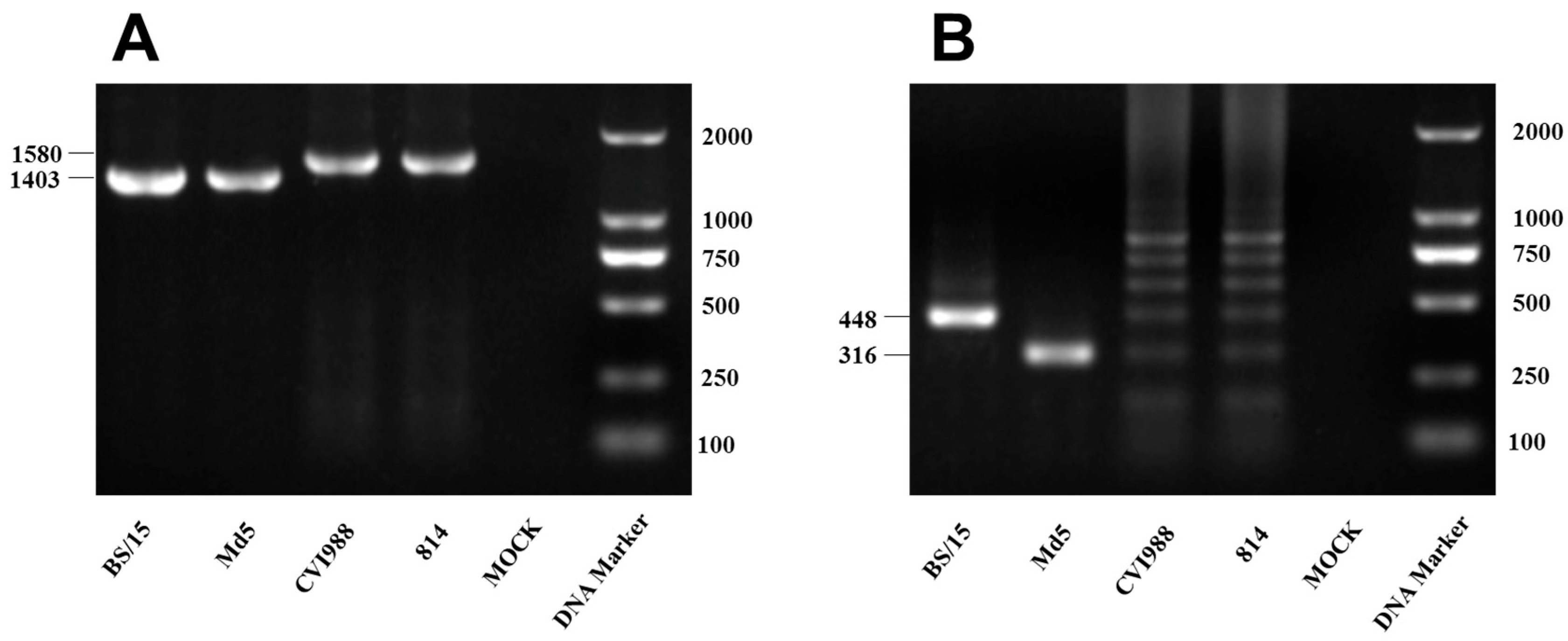
Viruses were identified by a polymerase chain reaction (PCR)-based method targeting the
meq gene and genomic 132-base pair repeat sequence (132bpr) of MDV, which we used to clearly distinguish between the vaccine and pathogenic wild-type MDV strains in a previous study [
15]. DNA samples from inoculated CEFs were used as PCR templates, and the PCR primers used for MDV strain identification are listed in
Table 1. Furthermore, an indirect immunofluorescence assay (IFA) was carried out to identify the viral plaques in CEFs using monoclonal antibodies, which were produced by our laboratory and are specific for the gI protein of MDV.
2.3. Screening for Causative Agents
Due to the multiple causes of oncosis in Chinese chicken flocks, it is necessary to detect and then cull chickens infected with one of several causative agents of oncogenicity. To detect avian leucosis virus (ALV), PCR and enzyme-linked immunosorbent assays (ELISAs) were performed, as previously described [
16,
17]. Additionally, PCR and IFA were used to detect reticuloendotheliosis virus (REV), as previously described [
16,
18]. Finally, a previously described PCR method was used to detect chicken infectious anemia virus (CIAV) [
19].
2.4. Animal Experiments
The MDV strain BS/15 was used as a challenge virus, and the standard vv MDV strain Md5 was used as a reference strain. The CVI988 vaccine and 814 vaccine purchased from commercial companies were used in animal experiments to assess vaccine efficacy.
In total, 105 one-day-old SPF White Leghorn chickens were obtained from the Experimental Animal Center (EAC) of HVRI. The birds were housed in negative-pressure-filtered air isolators and were randomly divided into seven groups (n = 15 birds each). Vaccination was performed on day 1 and both vaccines were used at a dose of 2000 plaque forming units (PFU) in 200 μL dedicated diluent provided by the manufacturers, being administered via the subcutaneous route (groups 3 and 6: CVI988; groups 4 and 7: 814). In addition, groups 1, 2, 3, 5, and 6 were inoculated with 200 μL diluent in the same manner. On day 7 post-vaccination, MDV challenge was performed via the intra-abdominal route with 1000 PFU of MDV in 200-μL diluent (groups 2, 3, and 4: Md5; groups 5, 6, and 7: BS/15). Chickens of group 1 received the same amount of diluent in the same manner and served as controls.
In this study, the birds were observed daily for clinical signs of MD, and feather pulps were randomly plucked from five birds at 4, 7, 14, 21, 28, 35, 42, and 90 days post-challenge (dpc;
n = 2 in the BS/15-challenged group, and
n = 3 in the Md5-challenged control group at 90 dpc). In addition, the body weights and immune organ (bursa, thymus, and spleen) weights of the surviving chickens at the end-point of the experiment (90 dpc) were recorded for further analysis. The MD status of the experimental animals was estimated mainly by monitoring for early mortality syndrome, immune organ damage, and tumor formation, as described previously [
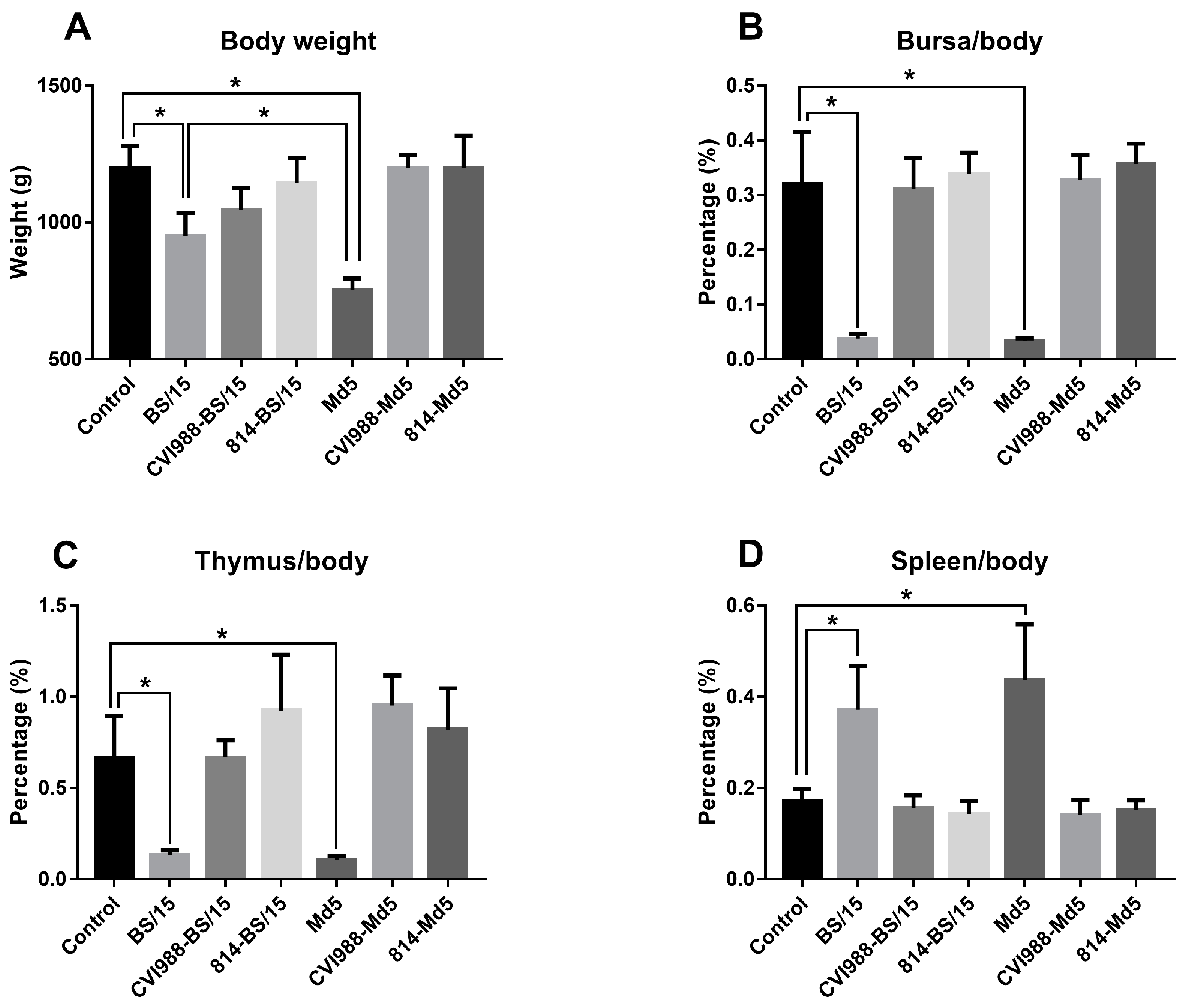
14]. The days that various chickens died during the experimental period in each group were recorded for survival analysis.
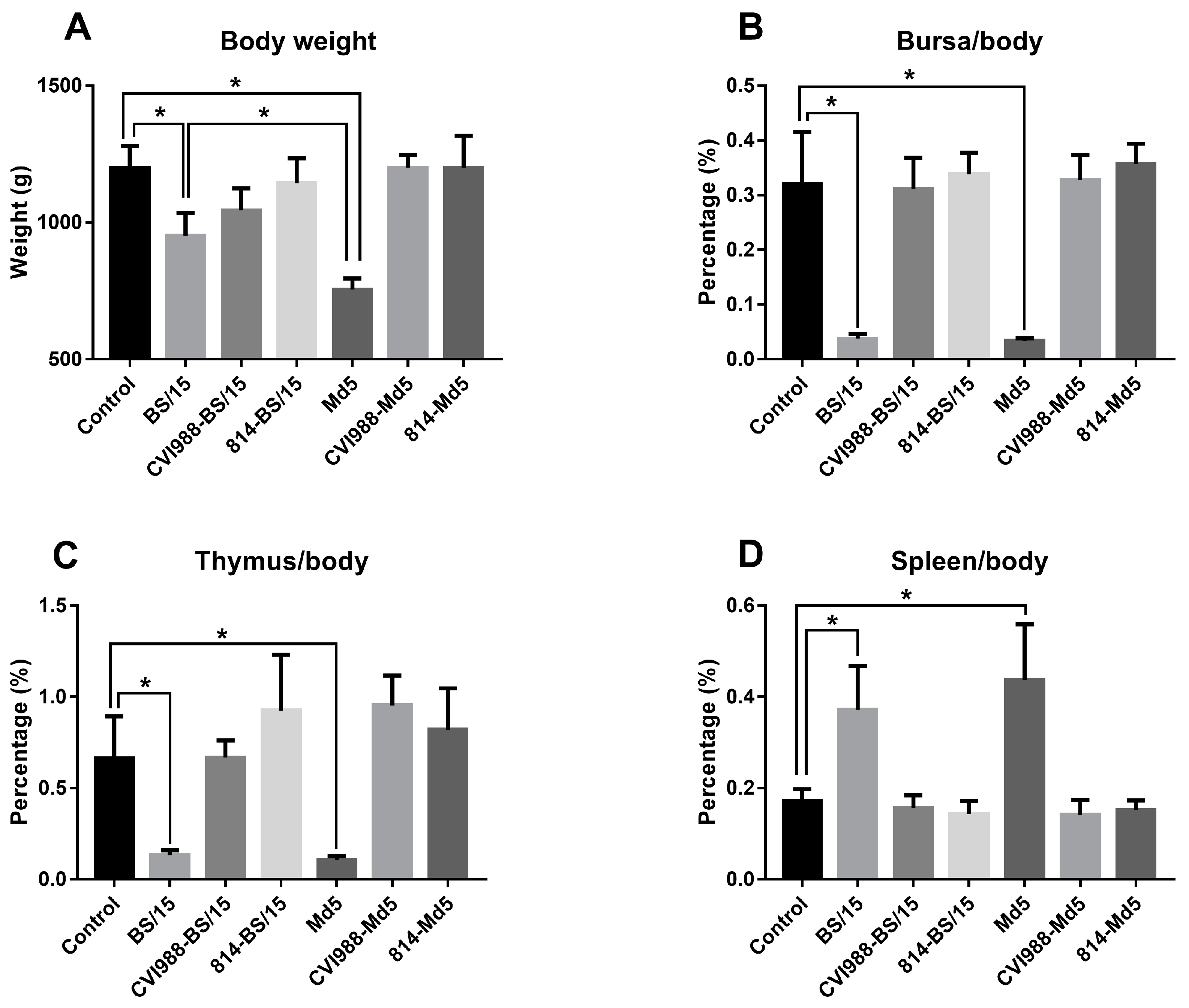
The birds were randomly assigned to each group and numbered by workers at the EAC, HVRI. To ensure a blinded study, the group number was only known by the workers who dealt directly with the researchers. All animal experiments were approved by the ethical review board of HVRI, CAAS, and performed in accordance with approved animal care guidelines and protocols (approval number: SYXK (Heilongjiang) 2011022). These standard procedures included using as few animals as possible (n = 15 in each group); ensuring that the workers of the EAC of HVRI were trained strictly, in accordance with animal policy, and complied with all standard operating procedures; the feeding environment was clean, spacious, and comfortable, allowing the animals to move and feed freely; the poultry feed was nutritionally balanced, and the drinking water was fresh and clean; during the experimental operation, the treatments were applied using gentle movements, to avoid frightening the animals; and at the end of the experiment, the animals were euthanized immediately.
2.5. DNA Extraction and TaqMan Real-Time Polymerase Chain Teaction (qPCR)
The collected feather pulps were homogenized in phosphate buffer solution (PBS), and DNA was extracted using the AxyPrep Body Fluid Viral DNA/RNA Miniprep Kit (Corning Life Sciences Co., Ltd., Suzhou, China) according to the manufacturer’s instructions. The MDV meq gene and the chicken ovotransferrin gene were used as a real-time PCR (qPCR) target gene in the MDV genome and an internal reference gene in the host cell genome, respectively, and qPCR detection was performed as previously described [
20,
21].
2.6. Statistical Analysis
The vaccine protective index (PI) was calculated as previously described [
22], using the following formula: PI = ((%MD in unvaccinated chickens; %MD in vaccinated chickens))/%MD in unvaccinated chickens) × 100. The absolute numbers of the MDV genome per million cells from the collected feather pulps were calculated based on the standard curves generated and were normalized to the viral load. The normalized viral load was calculated using the formula: normalized viral load = log
10 ((MDV genome copy number/chicken genome copy number) × 10
6). Viral load, body weight, immune organ index, and survival analysis data were analyzed using GraphPad Prism (Version 7.02; GraphPad Software, Inc., San Diego, CA, USA). Comparisons of the viral load between each group at each time point were determined using Multiple
t tests (Holm–Sidak method, with alpha = 0.05, by GraphPad Prism), and the statistical significance of body weights and immune organ indexes between each group was evaluated using the same method. Survival patterns between each two groups were compared by Log-rank (Mantel–Cox) test. Differences were considered to be statistically significant at
p < 0.05.
4. Discussion
Although MDV has been continually evolving and has obtained enhanced virulence over the last few decades, no report has described MDV stains with more virulence than vv+, and no trend of a large-scale MD outbreak has been shown. However, vaccination failure of MD has often occurred in China in recent years [
14,
23,
24,
25,
26]. Here, a field MDV strain BS/15 was isolated from a Chinese chicken flock in 2015 that had been vaccinated with the commercial vaccine CVI988 and had developed severe MD. Determination of MDV pathotypes is useful for investigating the cause of excessive MD losses in vaccinated flocks [
27]. The ADOL method of MDV pathotyping is widely recognized throughout the world. However, because of the limitations of ADOL (requirements for specific chicken types, large numbers of birds, and objective statistical methods to measure lesion responses), it is hardly used in other laboratories. Thus, researchers at the ADOL proposed that the pathotypes of MDV could be evaluated by comparing the pathogenicity and vaccine efficacy between the MDV isolates and the reference MDV strains in local SPF chickens [
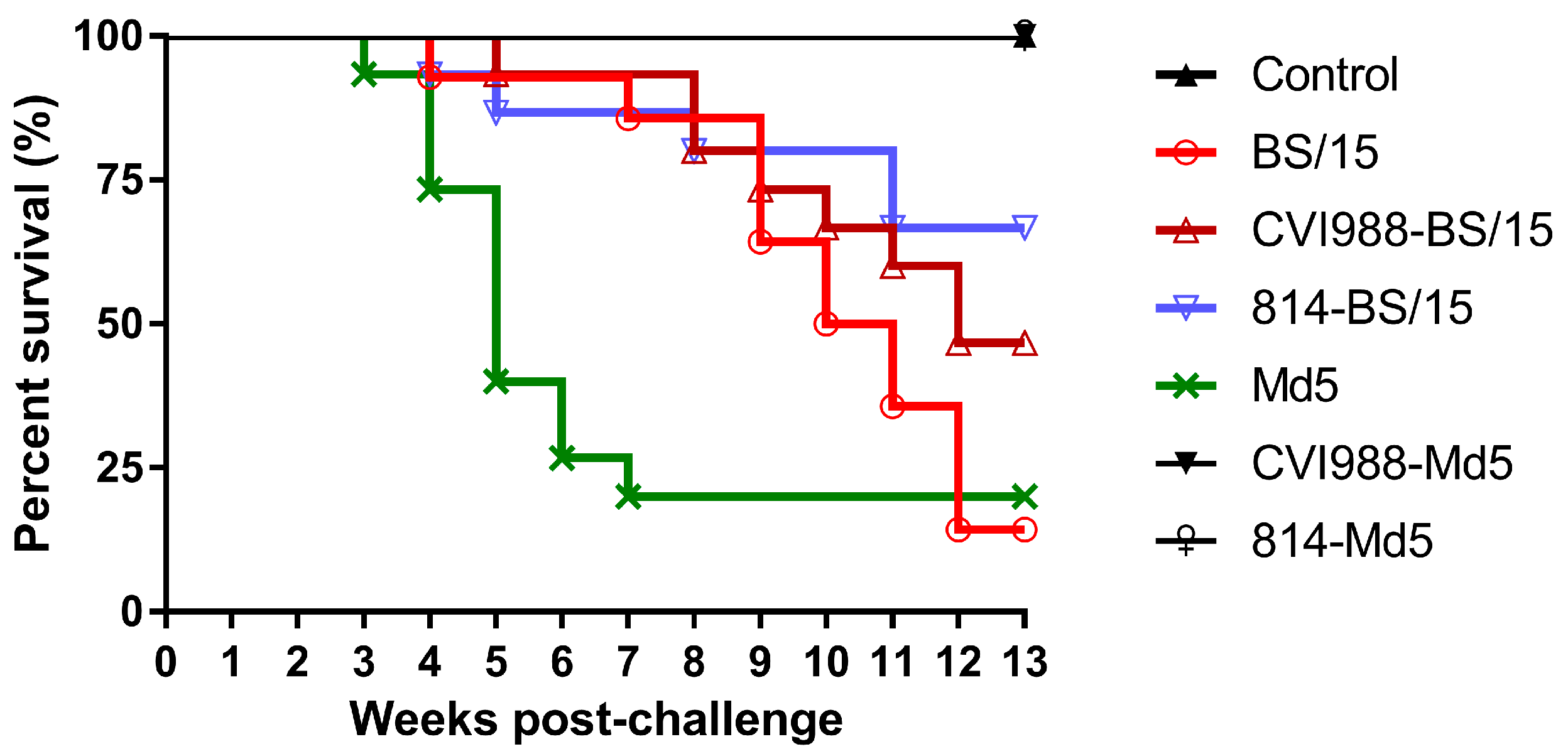
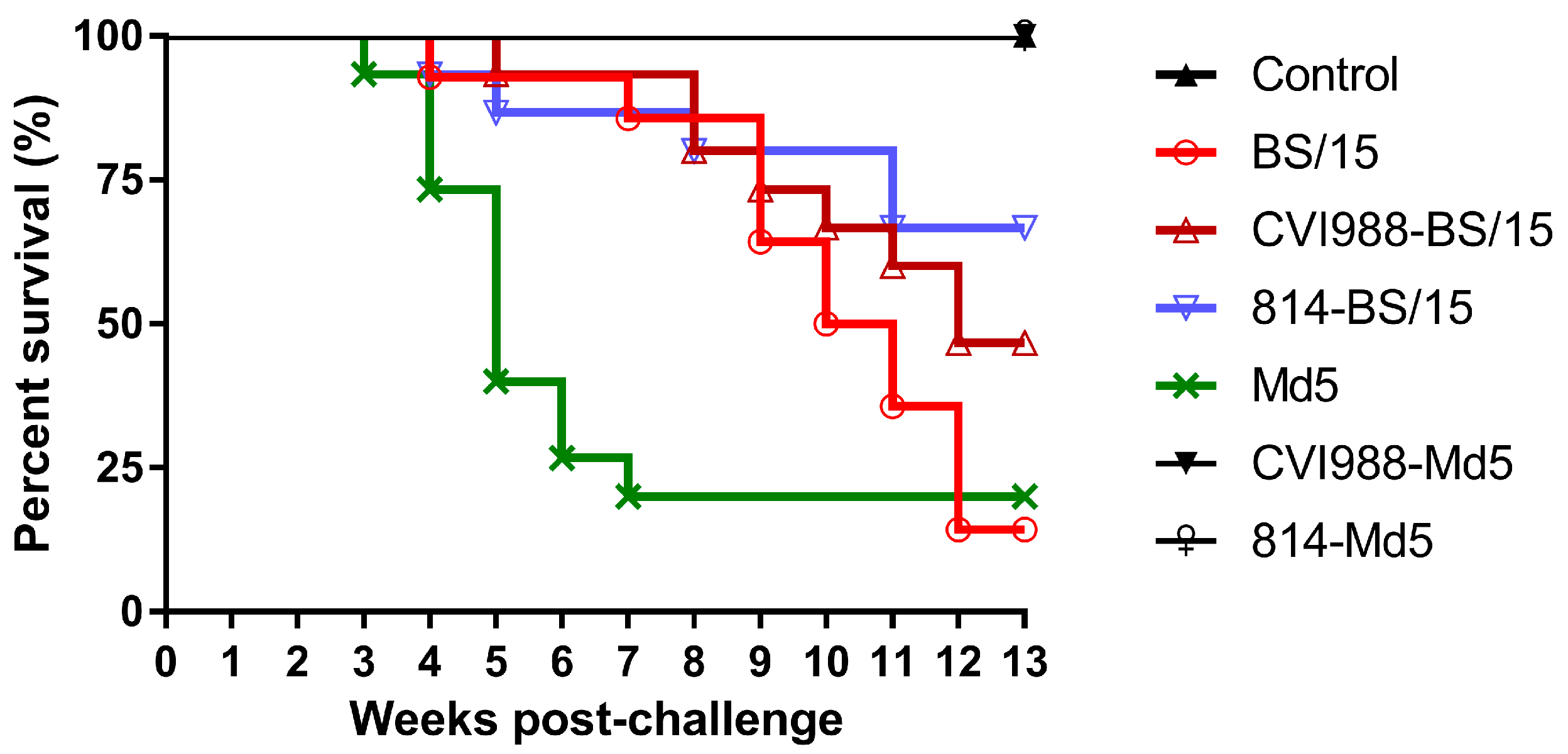
27]. In this study, to investigate the causes of vaccine immune failure, unvaccinated and CVI988- or 814-vaccinated SPF White Leghorn chickens were challenged with BS/15 and the standard vv MDV strain Md5 at day 7 post-vaccination, and the birds were observed for 90 dpc.
Animal experimental results showed that BS/15 and Md5 challenge both induced 100% MD incidence with similar mortality (85.7% and 80.0%, respectively) in unvaccinated SPF chickens. Unexpectedly, BS/15 induced much higher tumor rates than did Md5 in unvaccinated chickens (64.3% and 40.0%, respectively). In addition, although the number of surviving animals in the BS/15- and Md5-challenged groups at 90 dpc (
n = 2 and 3, respectively) was less than that in the other groups (
n = 5), which might allow for some statistical error, our data, to some extent, showed that BS/15 infection caused fewer serious effects on chicken growth and similar immune organ damages in unvaccinated chickens compared to Md5 (
Figure 4). In vaccine protection tests, the vaccines CVI988 and 814 only provided PIs of 33.3 and 66.7 to SPF chickens against BS/15, while the PIs of vaccine CVI988 and 814 against Md5 were 92.9 and 100, respectively. Moreover, these results suggested that the virulence of BS/15 was similar to that of the vv MDV strain Md5 with different pathogenic characteristics, but vaccines could not provide effective protection against BS/15, especially the vaccine CVI988. These results confirmed that the virulence and vaccine resistance of MDV are different traits, as previously discussed [
14,
28]. Thus, we could not clearly classify the pathotype of MDV BS/15 by the ADOL pathotyping method. However, this did not affect our understanding of the pathogenicity and the potential threat of MDV BS/15. Data generated in this study demonstrated the existence of variant Chinese strains with known virulence with altered pathogenic characteristics, which were highly resistant to existing vaccines. Thus, the presence of such strains in the field may be a cause of MD vaccination failure in China.
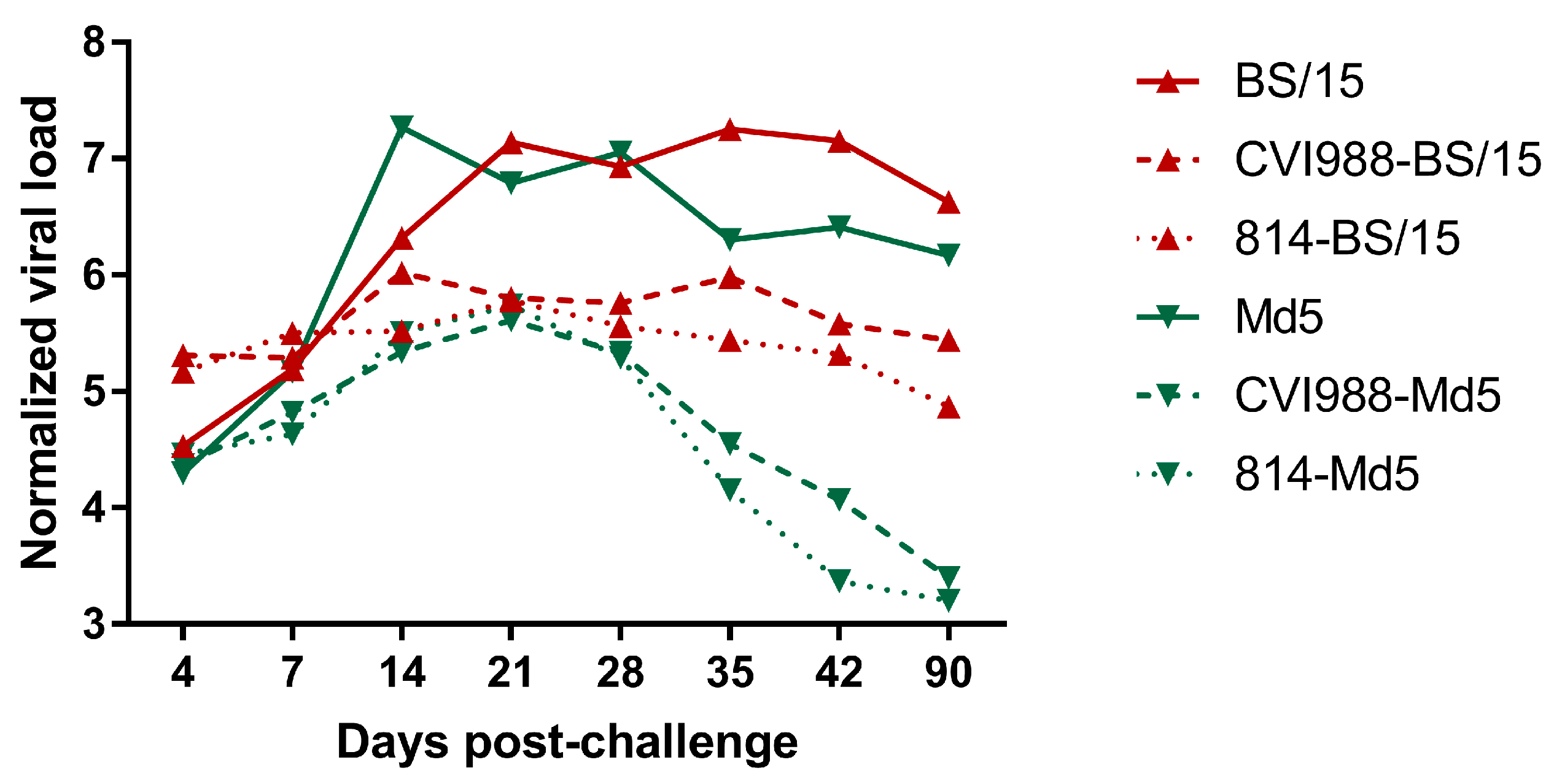
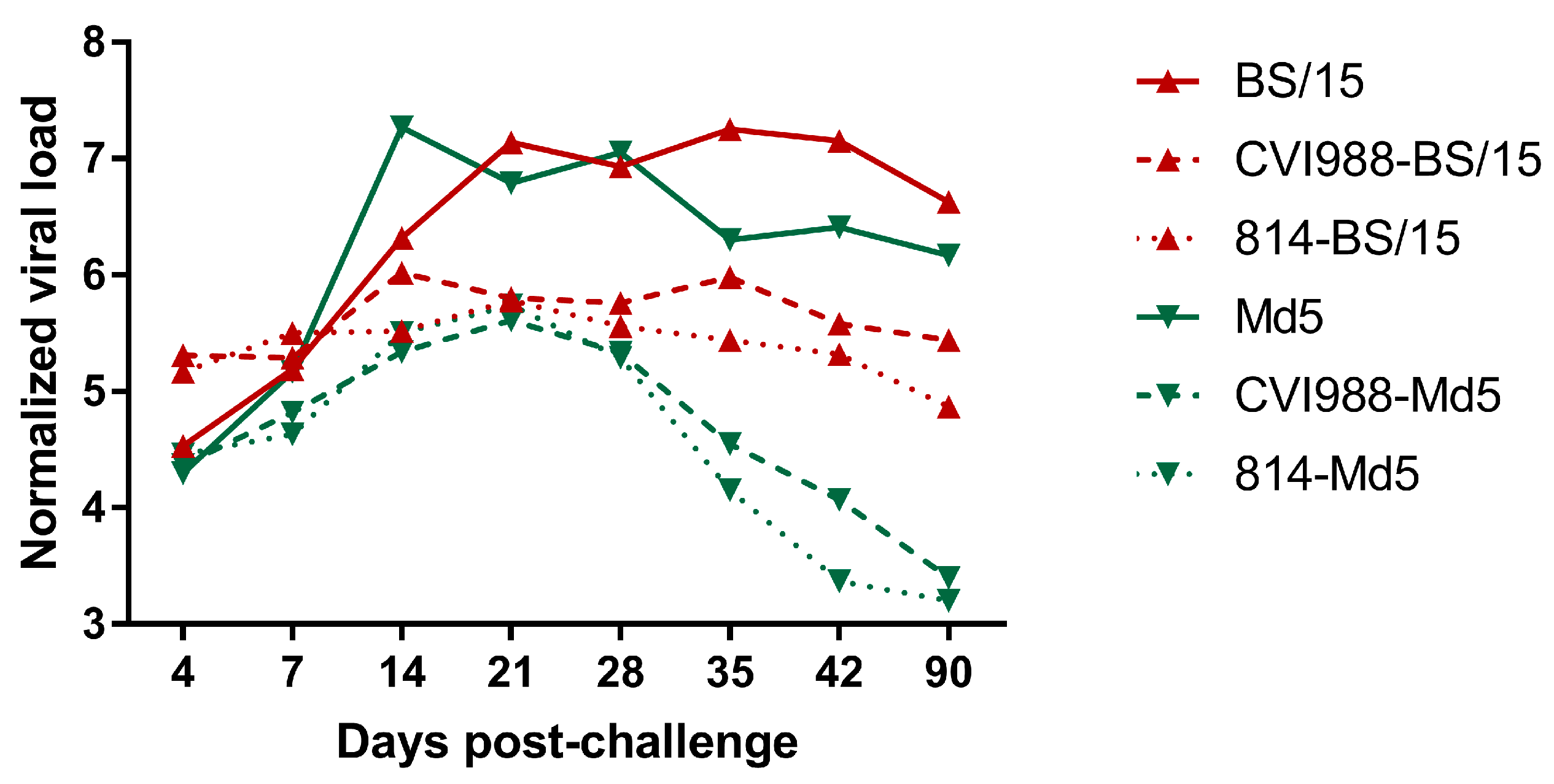
The MDV genome load in the feather pulps of chickens plays an important role in monitoring the MD status and vaccination performance [
21]. In this study, we quantified the viral load in chicken feather pulps (
Figure 5). The results showed that the viral loads of BS/15-challenged unvaccinated chickens were lower than those of Md5-challenged unvaccinated chickens at 14 dpc (
p < 0.05), but were higher than those of Md5-challenged unvaccinated chickens at 35 and 42 dpc (
p < 0.05 for each comparison). The lower viral loads observed in BS/15-challenged unvaccinated chickens during an early experimental period (about 7–21 dpc) may be associated with the delayed pathogenesis of BS/15. As shown in
Figure 3, increasing deaths occurred in the BS/15-challenged group beginning at 9 wpc, while none of the chickens died in the Md5-challenged control group after 7 wpc. These data indicated that chickens in the BS/15-challenged group were suffering from MD, while some chickens in the Md5-challenged control group were already resistant to MD at 35 and 42 dpc, as consistently revealed by measuring the viral load in the two groups at the same time point. Furthermore, the vaccines could not effectively inhibit the replication of BS/15 compared to Md5 after 28 dpc (
p < 0.05 for each comparison), which may explain the different protective effects of vaccines against BS/15 and Md5 to a certain extent.
Previous epidemiological studies have demonstrated the lack of efficacy of vaccines against Chinese MDV isolates with virulence of no obvious enhancement. SD2012-1 has a long latent period and causes subsequent death with a mortality rate of 70.3% in unvaccinated SPF chickens; however, SD2012-1 can break through the protection provided by vaccine HVT (FC126 strain) or bivalent vaccine HVT plus SB-1, causing mortality rates of over 60% in vaccinated SPF chickens [
23]. In another study, the CVI988 vaccine could only protect 83.0% of SPF chickens from SX1301 infection, despite the low mortality rate of 57.0% caused by SX1301 in unvaccinated SPF chickens [
24]. In our previous study, the field MDV strain LTS induced a very low mortality (23.1% by 60 dpc) in unvaccinated chickens, but the vaccine CVI988 could not provide efficient protection (PI: 66.7) against LTS [
14]. In addition, MDV isolates ZY/1203 and WC/1110 induced very low MD mortality in unvaccinated chickens (20.5% and 21.1% by 60 dpc, respectively) and were similar to that of the standard v MDV strain GA (23.5% by 60 dpc). Although vaccine CVI988 could provide good protection against ZY/1203 and WC/1110 (PI as 82.4 and 83.3, respectively), vaccine CVI988 could not provide complete protection against them compared to that of vaccine CVI988 against GA (PI: 100) [
25].
These studies suggested that the virulence of field MDV strains prevalent in China may not have increased in recent years, but several newly isolated MDV strains had obtained the ability to counter the vaccines, such as strain LTS isolated previously and strain BS/15 isolated in this study. We further inferred that the evolution patterns of MDV strains prevalent in China might have changed and no longer synchronously enhanced virulence and vaccine resistance. Thus, the newly isolated Chinese MDV strains often did not show stronger pathogenicity, but their survivability in vaccinated hosts was stronger, leading to increased vaccination failure in recent years. Therefore, the emergence of variant strains and their potential harm to the poultry industry need to be constantly monitored. Given the delayed pathogenesis of these variant MDV strains, their popularity in the field may be a greater threat to laying and breeding chickens than to meat-producing chickens. Thus, we are considering the necessity and feasibility of classifying these variant MDV strains as a new pathotype (such as “late virulence”), to facilitate the analysis and study of these variant strains in the future.

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}