
Comparative Study on the Antivirus Activity of Shuang–Huang–Lian Injectable Powder and Its Bioactive Compound Mixture against Human Adenovirus III In Vitro

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
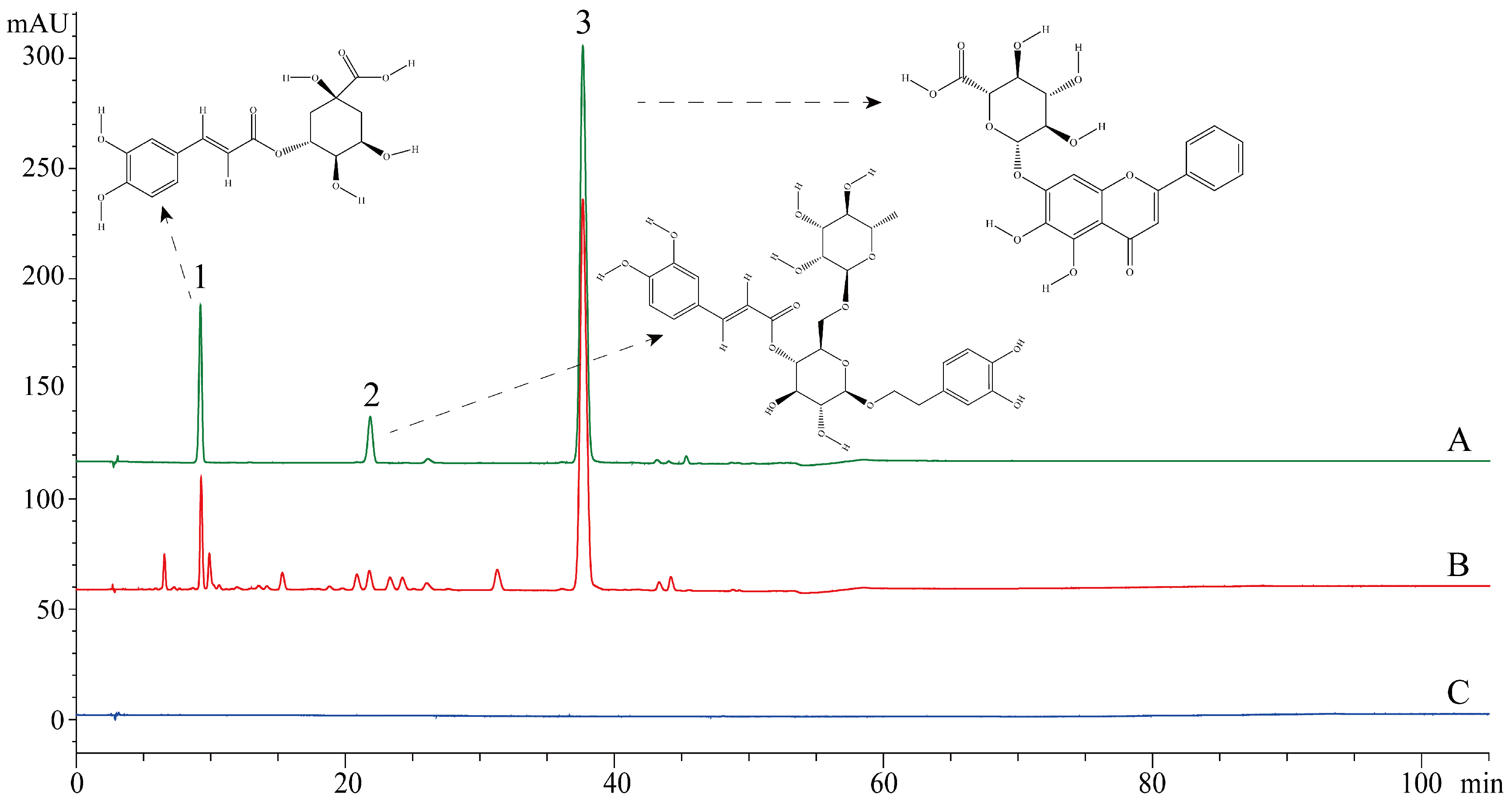
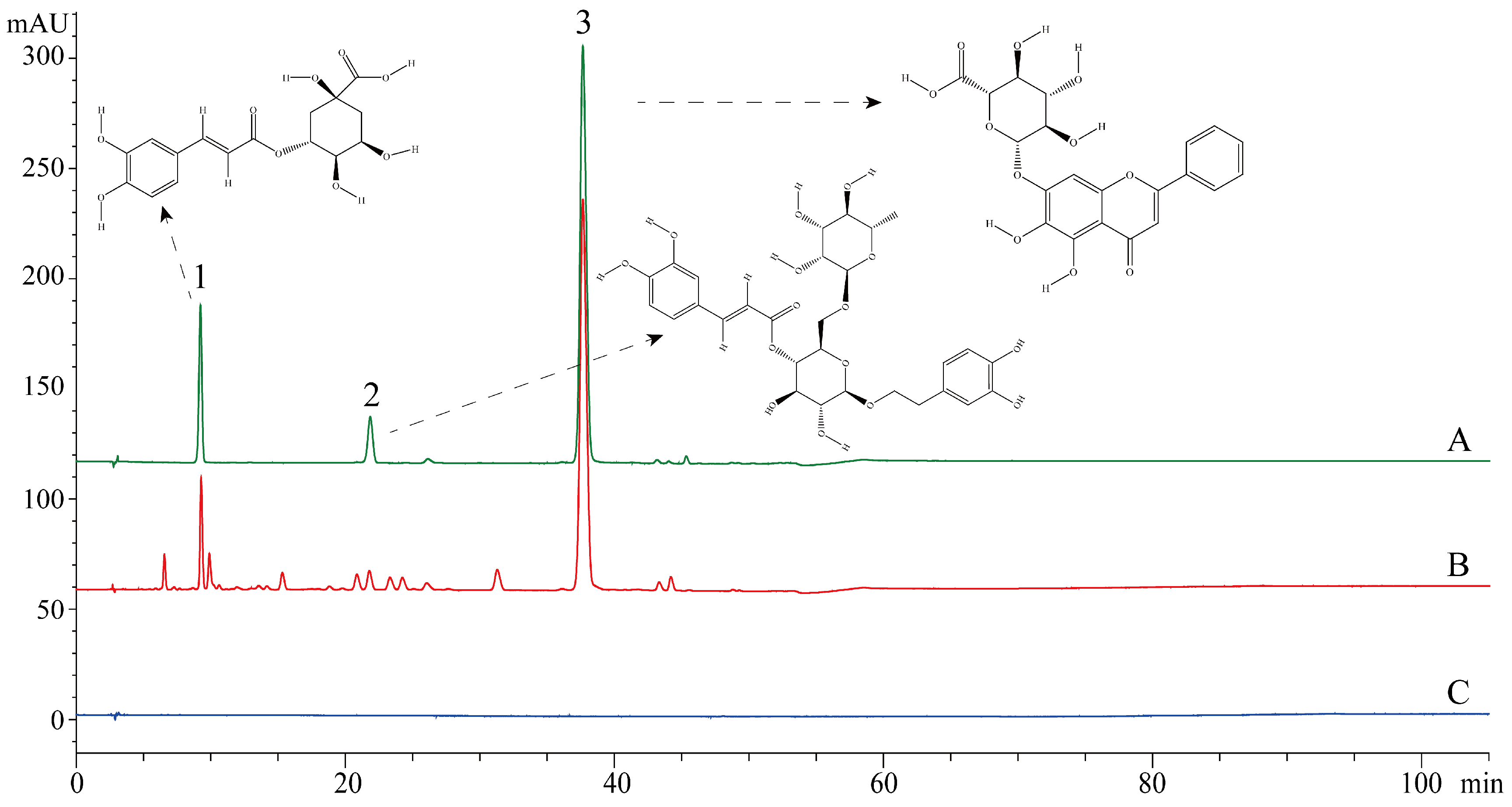
2.2. Chromatographic Conditions and Test Samples Preparation
2.3. Cells and Virus
2.4. Cytotoxicity Assay
2.5. Antiviral Effect Assay
2.6. Time of Addition Assay
2.7. Attachment Assay
2.8. Internalization Assay
2.9. Polymerase Chain Reaction (PCR) and Quantitative PCR (qPCR)
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Statistical Analysis
3. Results
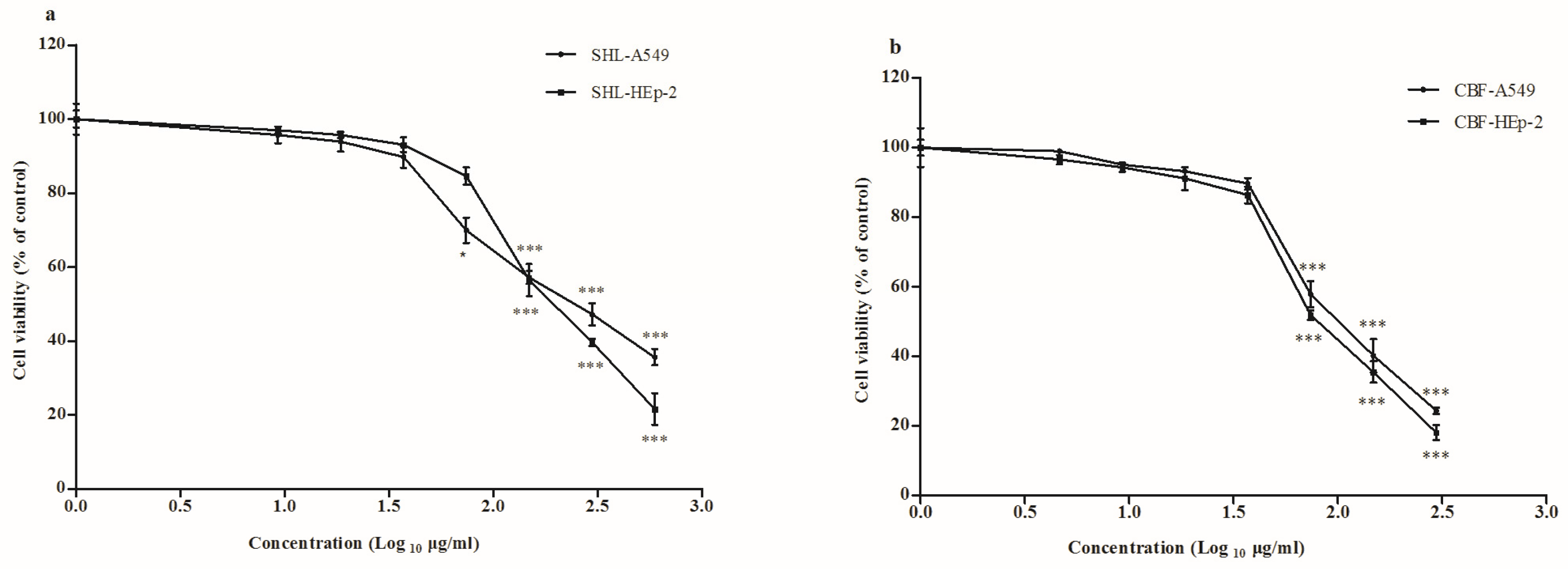
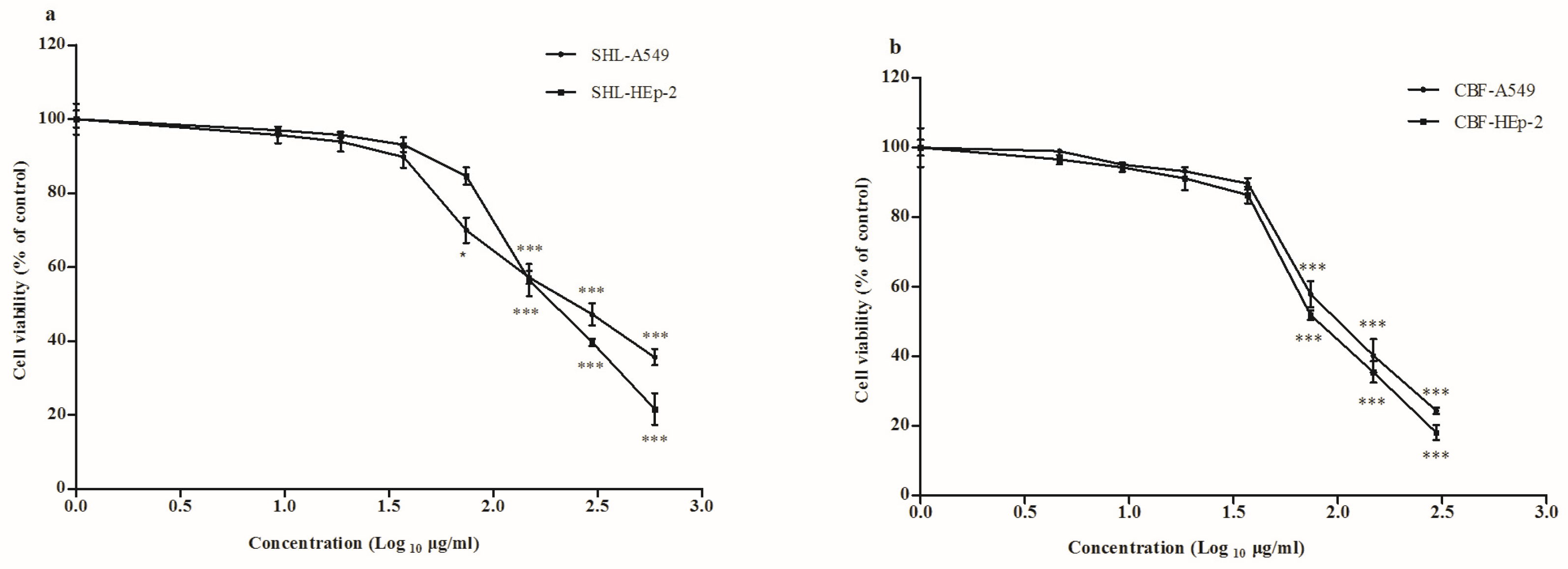
3.1. Cytotoxicity of SHL and CBF in A549 and HEp-2 Cells
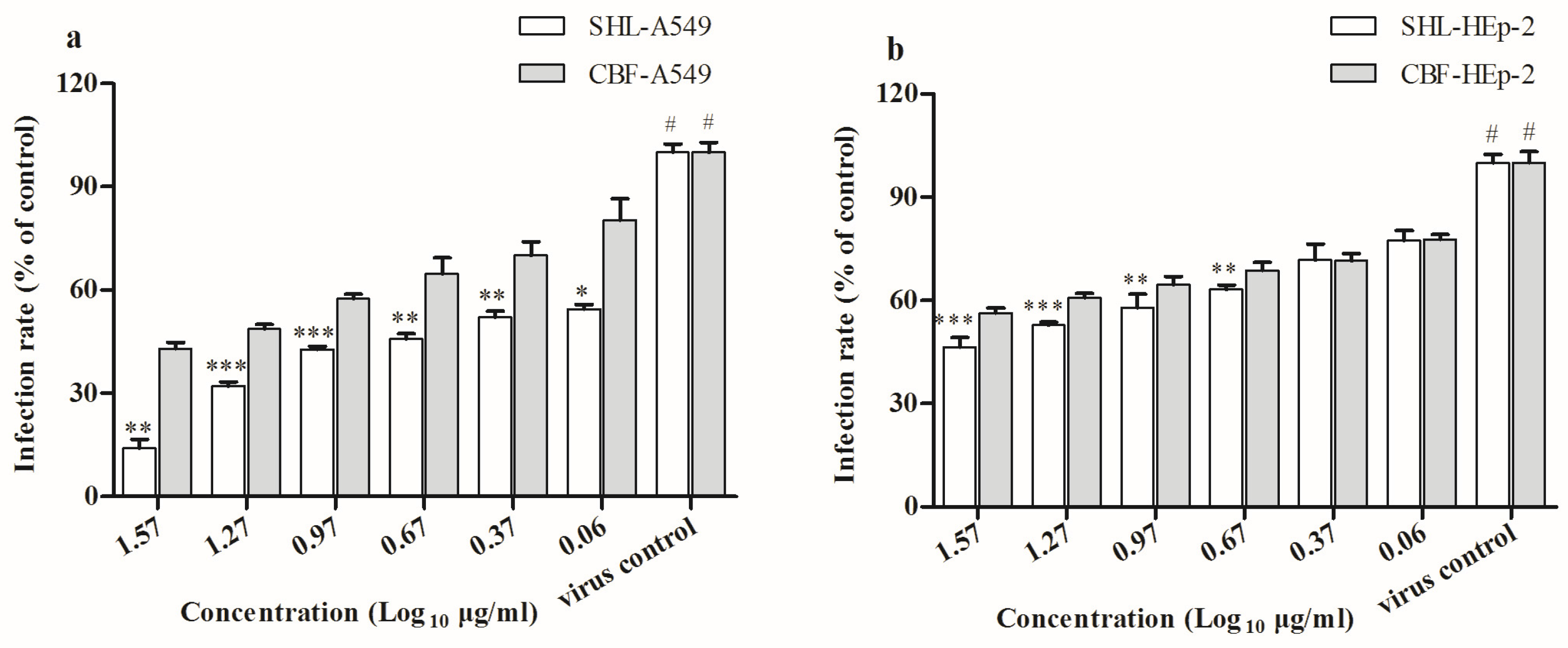
3.2. SHL Attenuated Virus Proliferation More Significantly Than CBF
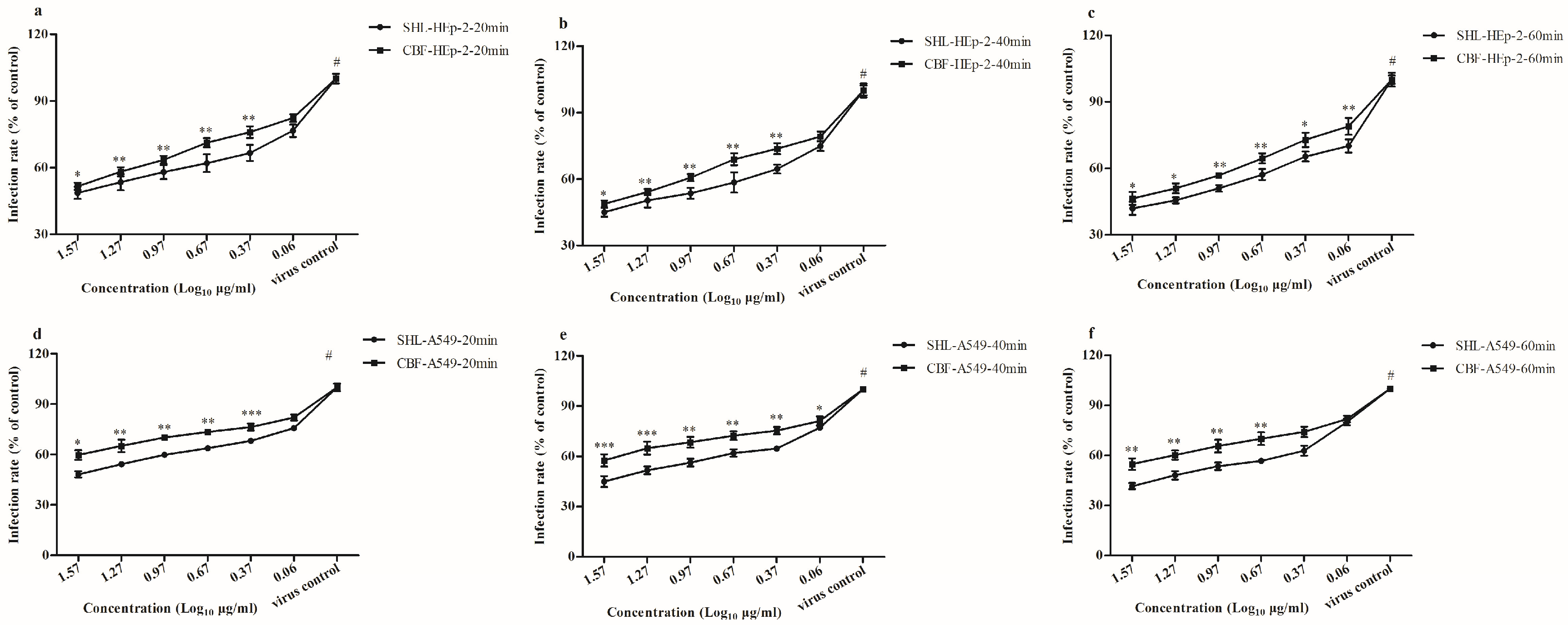
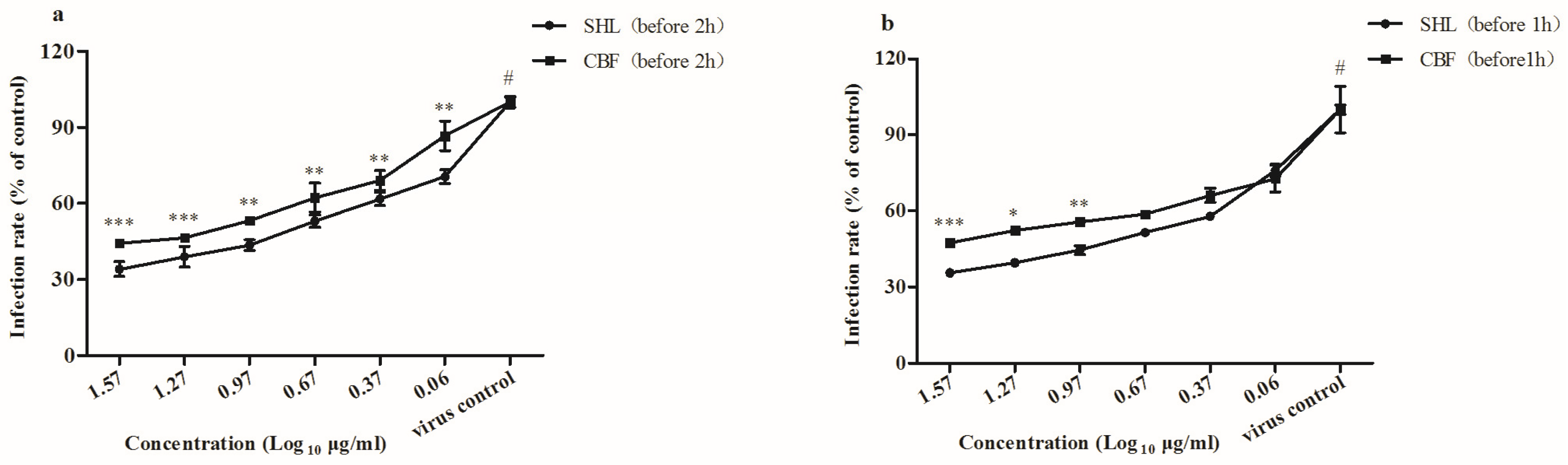
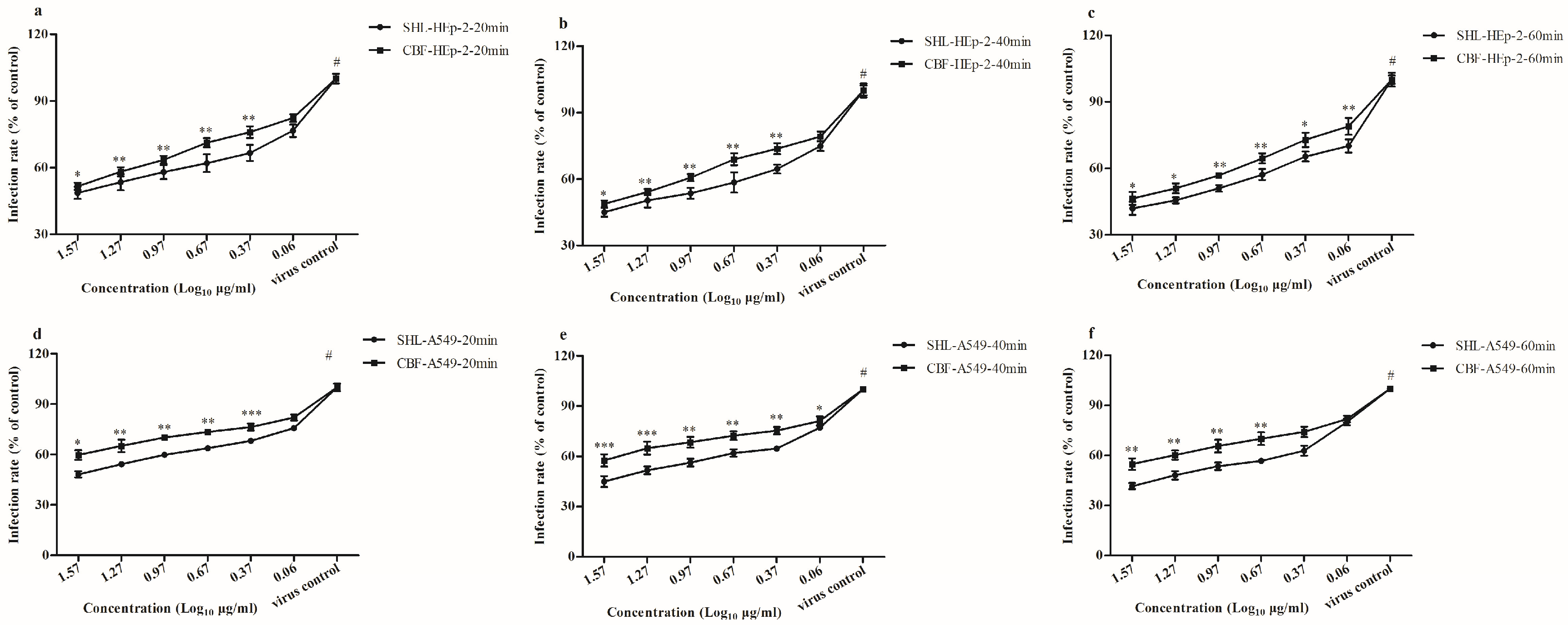
3.3. SHL Decreased Plaque Formation More Than CBF When Viral Inoculation Was Given in Different Working Points
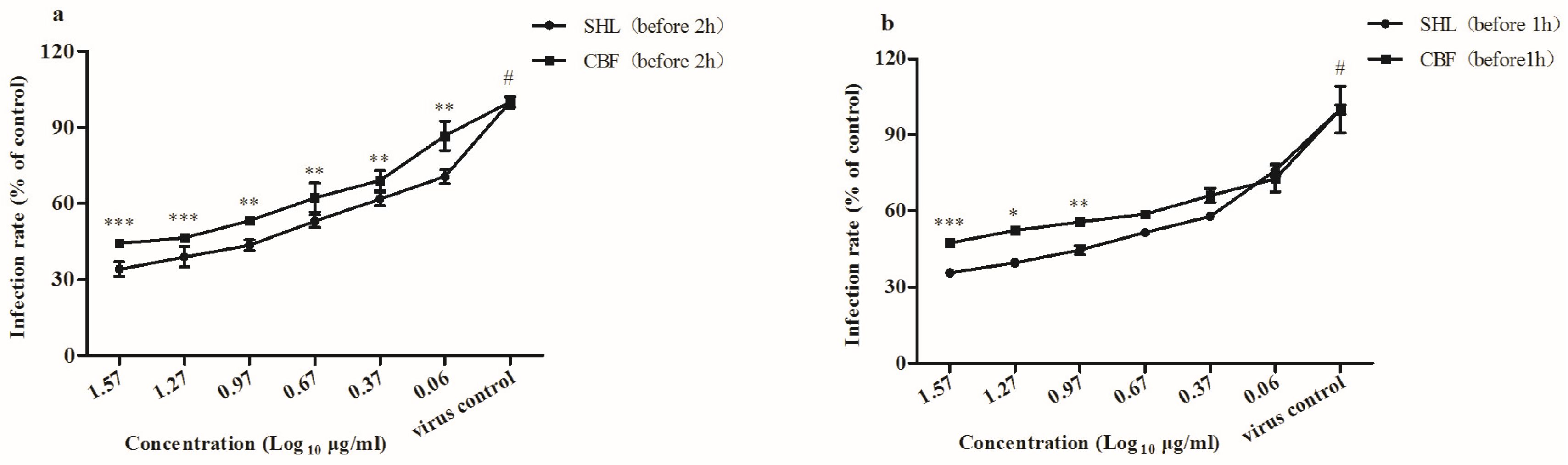
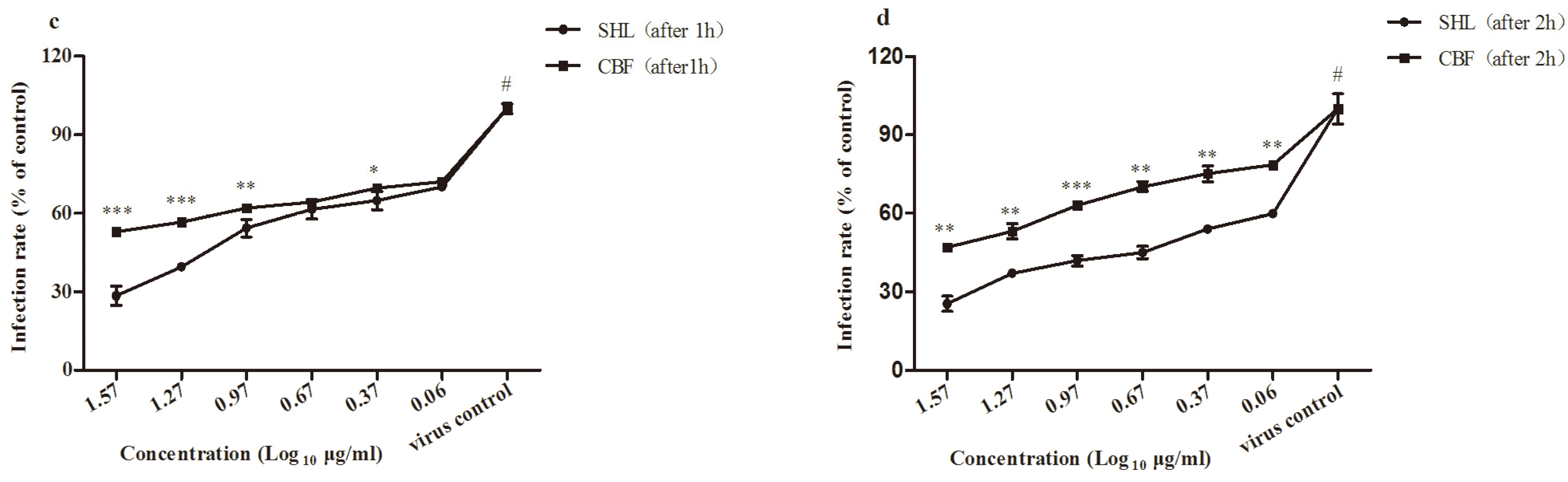
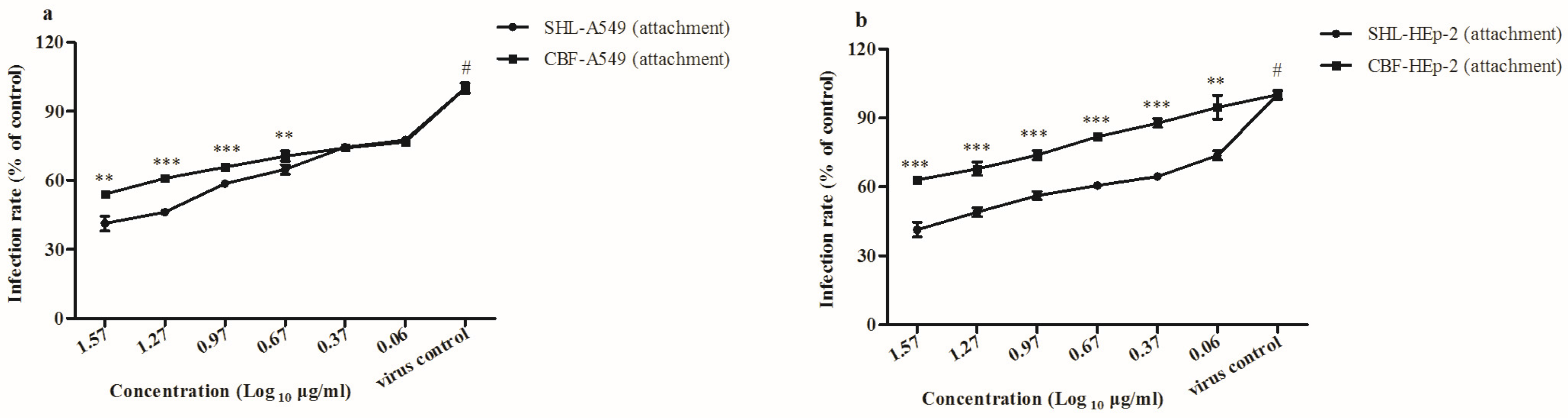
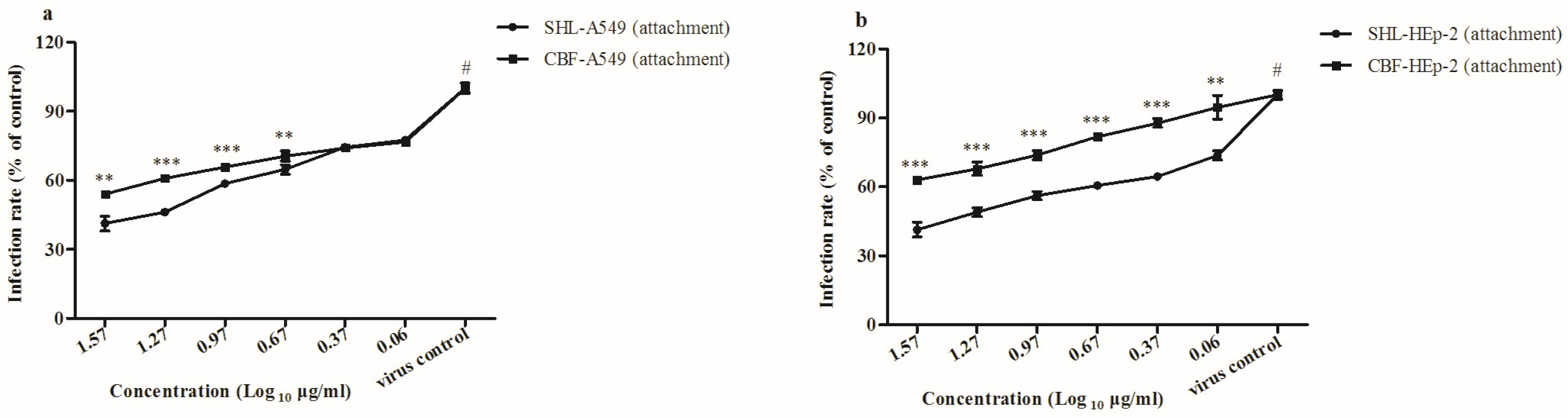
3.4. SHL Inhibited Viral Attachment Better Than CBF in A549 and HEp-2 Cells
3.5. SHL Affected Viral Internalization More Than CBF
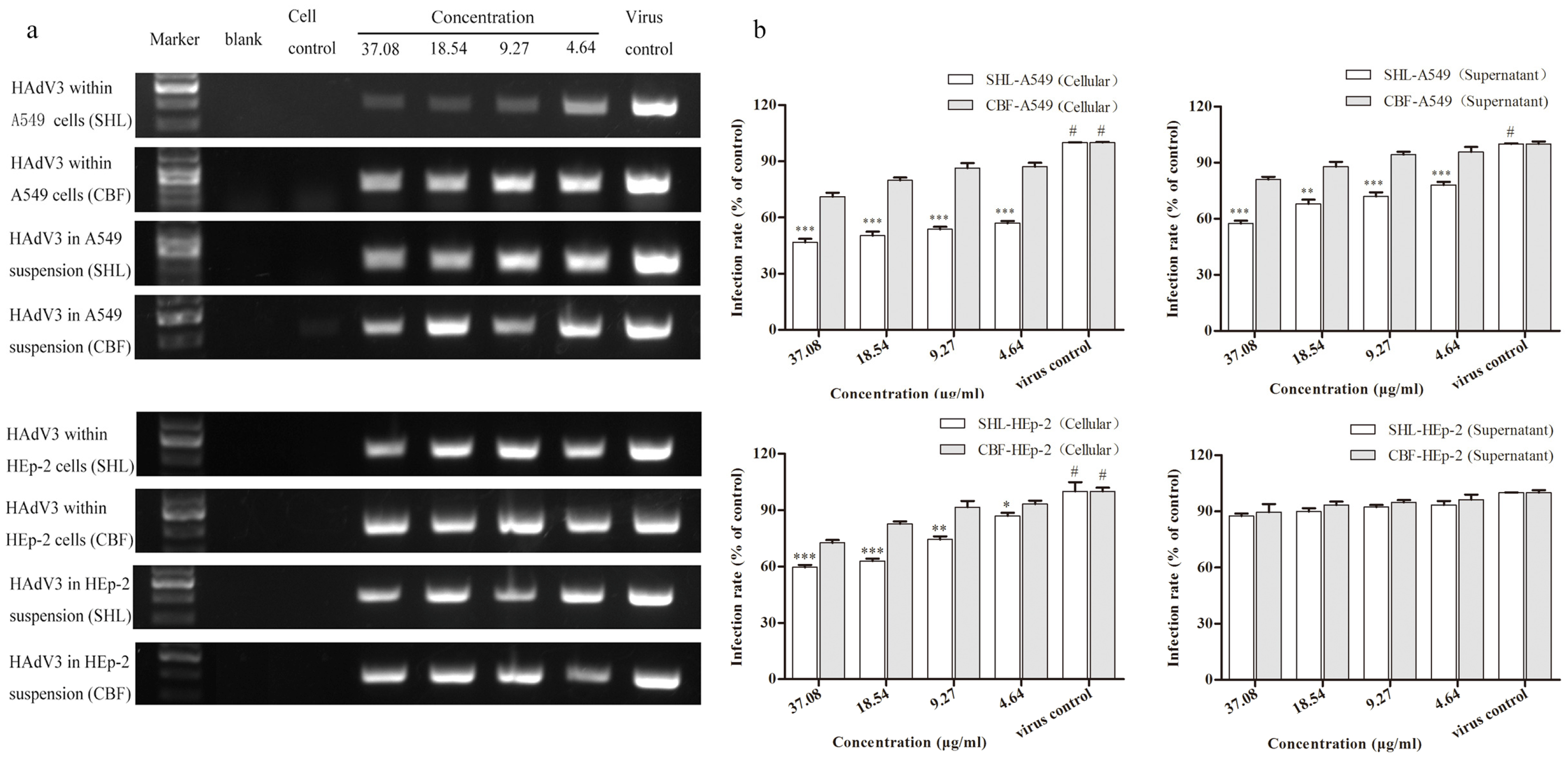
3.6. SHL Significantly Suppressed Viral DNA Replication in A549 and HEp-2 Cells Compared to CBF
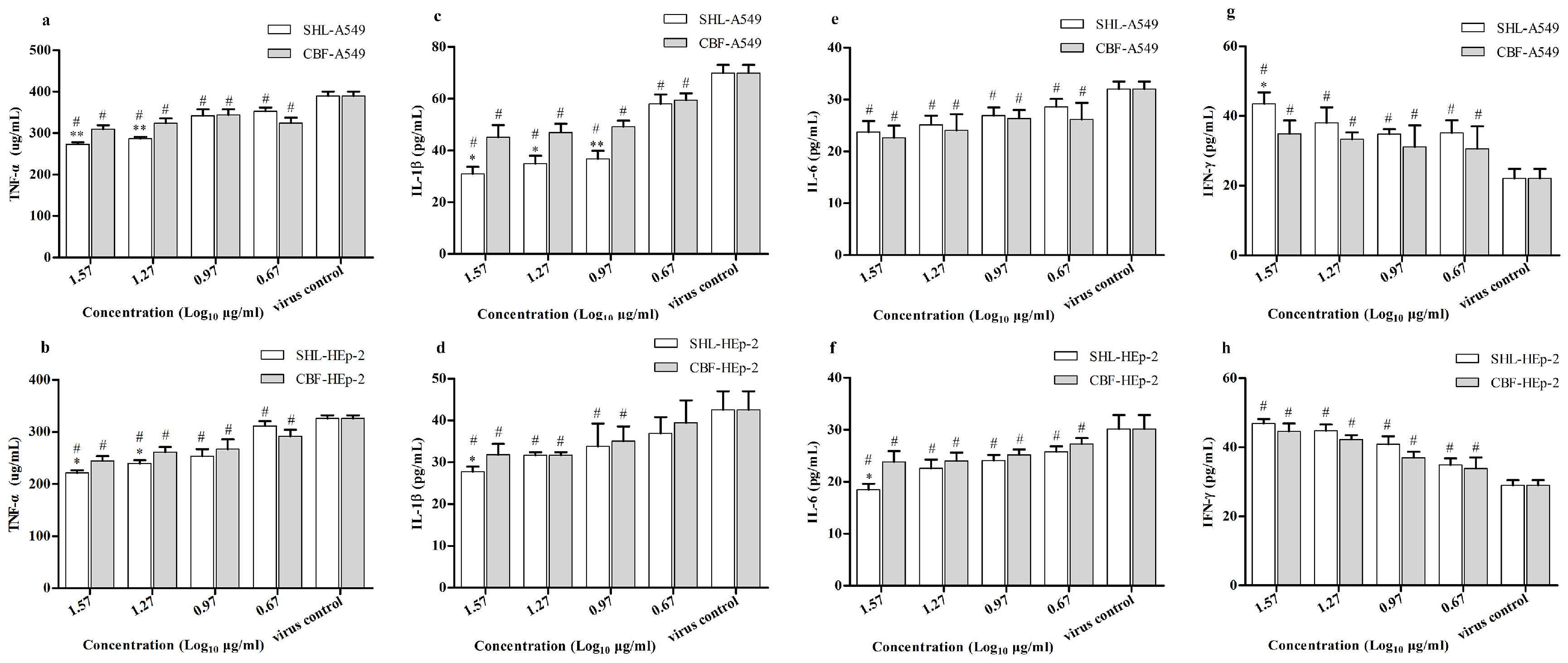
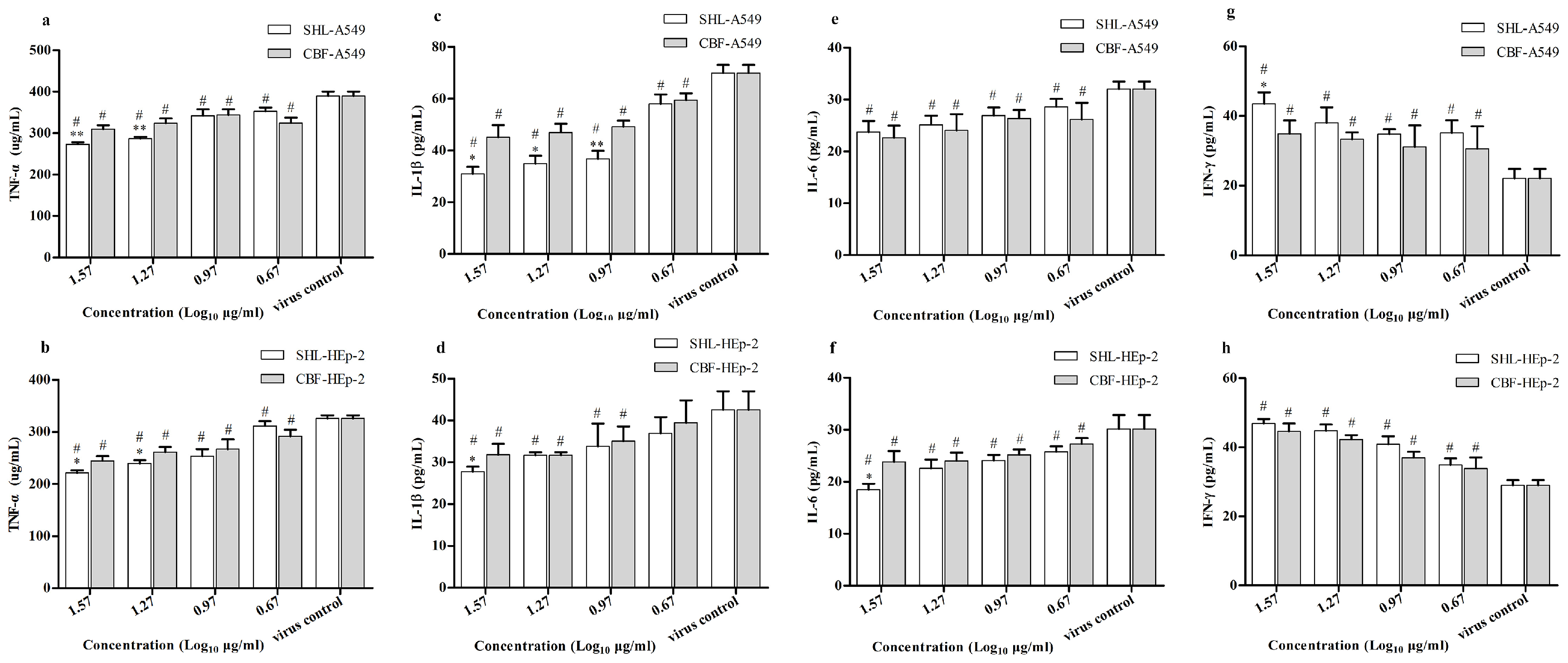
3.7. SHL Attenuated Inflammatory Effects Significantly More Than CBF in HAdV3-Stimulated A549 and HEp-2 Cells
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| CBF | chlorogenic acid, baicalin, forsythia glycosides A |
| CC50 | 50% cytotoxic concentrations |
| CCK-8 | cell counting kit-8 |
| DAD | diode array detector |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ELISA | enzyme-linked immunosorbent assay |
| FBS | fetal bovine serum |
| HAdV | human adenovirus |
| HIV | human immunodeficiency virus |
| HPLC | high performance liquid chromatography |
| HSD | honestly significant difference |
| IC50 | 50% inhibitory concentration |
| IFN | interferon |
| IL | interleukin |
| PCR | polymerase chain reaction |
| qPCR | quantitative polymerase chain reaction |
| S.D. | standard deviation |
| SHL | Shuang–Huang–Lian injectable powder |
| TCID50 | 50% tissue culture infection dose |
| TCM | traditional Chinese medicine |
| TNF-α | tumor necrosis factor alpha |
| DNA | deoxyribonucleic acid |
References
- Dehghan, S.; Liu, E.B.; Seto, J.; Torres, S.F.; Hudson, N.R.; Kajon, A.E.; Metzgar, D.; Dyer, D.W.; Chodosh, J.; Jones, M.S.; et al. Five genome sequences of subspecies B1 human adenoviruses associated with acute respiratory disease. J. Virol. 2012, 86, 635–636. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.M.; Zhou, X.; Rajaiya, J.; Yousuf, M.A.; Singh, G.; DeSerres, J.J.; Walsh, M.P.; Wong, S.; Seto, D.; Dyer, D.W.; et al. Predicting the next eye pathogen: Analysis of a novel adenovirus. MBio 2013, 4, e00595:1–e00595:12. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.C.; Fugate, W.H.; Murray, C.K.; Cropper, T.L.; Lott, L.; McDonald, J.M. Pandemic influenza virus 2009 H1N1 and adenovirus in a high risk population of young adults: Epidemiology, comparison of clinical presentations, and coinfection. PLoS ONE 2014, 9, e85094. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.B.; Tong, Y.G.; Wo, Y.; Wang, H.Y.; Liu, E.M.; Gray, G.C.; Liu, W.; Cao, W.C. Epidemiology of human adenovirus and molecular characterization of human adenovirus 55 in China, 2009–2012. Influenza Other Respir. Viruses 2014, 8, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Barrero, P.R.; Valinotto, L.E.; Tittarelli, E.; Mistchenko, A.S. Molecular typing of adenoviruses in pediatric respiratory infections in Buenos Aires, Argentina (1999–2010). J. Clin. Virol. 2012, 53, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Alkhalaf, M.A.; Guiver, M.; Cooper, R.J. Genome stability of adenovirus types 3 and 7 during a simultaneous outbreak in Greater Manchester, UK. J. Med. Virol. 2015, 87, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Ampuero, J.S.; Ocana, V.; Gomez, J.; Gamero, M.E.; Garcia, J.; Halsey, E.S.; Laguan-Torres, V.A. Adenovirus respiratory tract infections in Peru. PLoS ONE 2012, 7, e46898. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Gonzalez, R.; Zhou, H.; Wu, C.; Vernet, G.; Wang, Z.; Wang, J. Detection of three human adenovirus species in adults with acute respiratory infection in China. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.Y.L.C. Adenovirus serotype 3 and 7 infection with acute respiratory failure in children inTaiwan, 2010–2011. PLoS ONE 2013, 8, e53614. [Google Scholar]
- Lee, W.J.; Jung, H.D.; Cheong, H.M.; Kim, K. Molecular epidemiology of a post-influenza pandemic outbreak of acute respiratory infections in Korea caused by human adenovirus type 3. J. Med. Virol. 2015, 87, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.S.; Cheng, F.; Shi, Y.Q.; Li, Z.G.; Qin, H.D.; Liu, Z.P. Hypotensive response in rats and toxicological mechanisms induced by Shuanghuanglian, an herbal extract mixture. Drug Discov. Ther. 2010, 4, 13–18. [Google Scholar] [PubMed]
- McDougall, B.; King, P.J.; Wu, B.W.; Hostomsky, Z.; Reinecke, M.G.; Robinson, W.J. Dicaffeoylquinic and dicaffeoyltartaric acids are selective inhibitors of human immunodeficiency virus type 1 integrase. Antimicrob. Agents Chemother. 1998, 42, 140–146. [Google Scholar] [PubMed]
- Chiang, L.C.; Chiang, W.; Chang, M.Y.; Ng, L.T.; Lin, C.C. Antiviral activity of Plantago major extracts and related compounds in vitro. Antivir. Res. 2002, 55, 53–62. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, J.; Li, W.; Shen, L.; Huang, S.; Tang, J.; Duan, J.; Fang, F.; Huang, Y.; Chang, H.; et al. Computational screen and experimental validation of anti-influenza effects of quercetin and chlorogenic acid from traditional Chinese medicine. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Gamaleldin, E.K.M.; Matei, M.F.; Jaiswal, R.; Illenberger, S.; Kuhnert, N. Neuraminidase inhibition of Dietary chlorogenic acids and derivatives—Potential antivirals from dietary sources. Food Funct. 2016, 7, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, Y.; Hou, X.; Peng, H.; Zhang, L.; Jiang, Q.; Shi, M.; Ji, Y.; Wang, Y.; Shi, W. Chlorogenic acid inhibits the replication and viability of enterovirus 71 in vitro. PLoS ONE 2016, 7, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.J.; Gu, L.G.; Yu, H.T. Antagonism of baicalin on cell cyclical distribution and cell apoptosis in A549 cells infected with influenza A (H1N1) virus. Bing Du Xue Bao 2011, 27, 108–116. (In Chinese) [Google Scholar] [PubMed]
- Moghaddam, E.; Teoh, B.T.; Sam, S.S.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a metabolite of baicalein with antiviral activity against dengue virus. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Wu, Z.; Gao, B.; Xu, J. Analysis of influence of baicalin joint resveratrol retention enema on the TNF-alpha, SIgA, IL-2, IFN-gamma of rats with respiratory syncytial virus infection. Cell Biochem. Biophys. 2014, 70, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, J.; Zhang, Z.; Ma, Y.; Liao, F.; Zhang, Y.; Wu, G. Forsythoside A inhibits the avian infectious bronchitis virus in cell culture. Phytother. Res. 2011, 25, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.; Liu, J.; Wei, L.; Song, B.; Shao, L. Exposure of the murine RAW 264.7 macrophage cell line to dicalcium silicate coating: Assessment of cytotoxicity and pro-inflammatory effects. J. Mater. Sci. Mater. Med. 2016, 27. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.S.; Wang, K.C.; Shieh, D.E.; Hsu, F.F.; Chiang, L.C. Ge-Gen-Tang has anti-viral activity against human respiratory syncytial virus in human respiratory tract cell lines. J. Ethnopharmacol. 2012, 139, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Bartee, E.; Mohamed, M.R.; McFadden, G. Tumor necrosis factor and interferon: Cytokines in harmony. Curr. Opin. Microbiol. 2008, 11, 378–383. [Google Scholar] [CrossRef] [PubMed]
- McFadden, G.; Mohamed, M.R.; Rahman, M.M.; Bartee, E. Cytokine determinants of viral tropism. Nat. Rev. Immunol. 2009, 9, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Cheng, Q. Etiological mechanisms of post-stroke depression: A review. Neurol. Res. 2009, 31, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Trefler, J.; Paradowska-Gorycka, A.; Lacki, J.K. Influence of genetic factors on development and severity of rheumatoid arthritis—Part II. Pol. Merkur. Lekarski 2009, 27, 161–165. [Google Scholar] [PubMed]
- Masters, S.L.; Simon, A.; Aksentijevich, I.; Kastner, D.L. Horror autoinflammaticus: The molecular pathophysiology of autoinflammatory disease. Annu. Rev. Immunol. 2009, 27, 621–668. [Google Scholar] [CrossRef] [PubMed]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Liang, D.; Song, S.; Yu, Q.; Shi, C.; Xing, X.; Luo, J.-B. Comparative Study on the Antivirus Activity of Shuang–Huang–Lian Injectable Powder and Its Bioactive Compound Mixture against Human Adenovirus III In Vitro. Viruses 2017, 9, 79. https://doi.org/10.3390/v9040079
Ma Q, Liang D, Song S, Yu Q, Shi C, Xing X, Luo J-B. Comparative Study on the Antivirus Activity of Shuang–Huang–Lian Injectable Powder and Its Bioactive Compound Mixture against Human Adenovirus III In Vitro. Viruses. 2017; 9(4):79. https://doi.org/10.3390/v9040079
Chicago/Turabian StyleMa, Qinhai, Dedong Liang, Shuai Song, Qintian Yu, Chunyu Shi, Xuefeng Xing, and Jia-Bo Luo. 2017. "Comparative Study on the Antivirus Activity of Shuang–Huang–Lian Injectable Powder and Its Bioactive Compound Mixture against Human Adenovirus III In Vitro" Viruses 9, no. 4: 79. https://doi.org/10.3390/v9040079