
Label-Free Quantitative Proteomic Analysis of Chitosan Oligosaccharide-Treated Rice Infected with Southern Rice Black-Streaked Dwarf Virus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chitosan Oligosaccharide to Promote Rice Rooting and Germination
2.2. Plant Material and Samples of Proteomics
2.3. Extraction of Total Proteins and Polyacrylamide Gel Electrophoresis
2.4. In-Gel Digestion
2.5. LC-MS/MS, Database Searching, and Bioinformatics Analysis
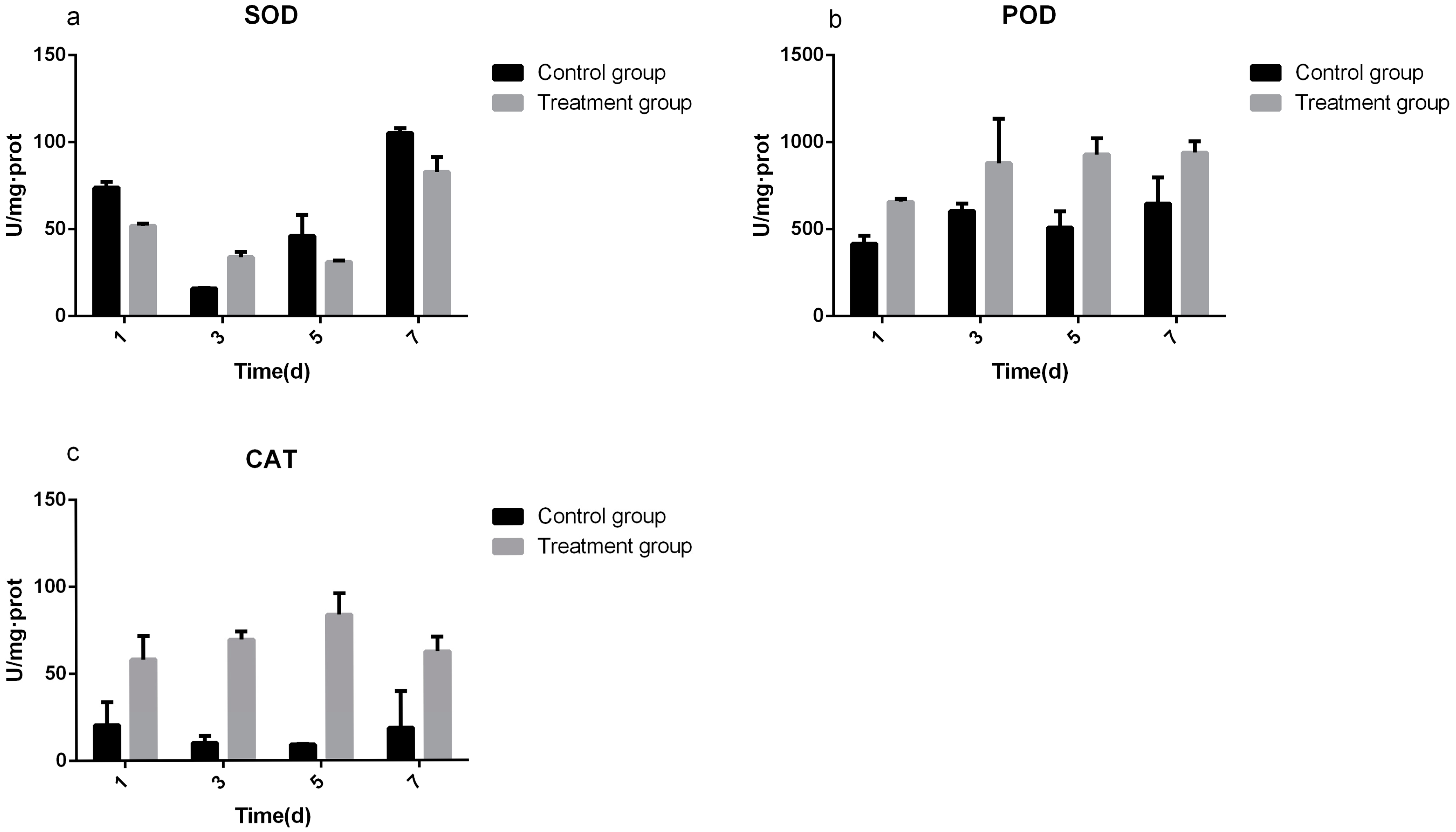
2.6. Defense Enzyme Activity Assay
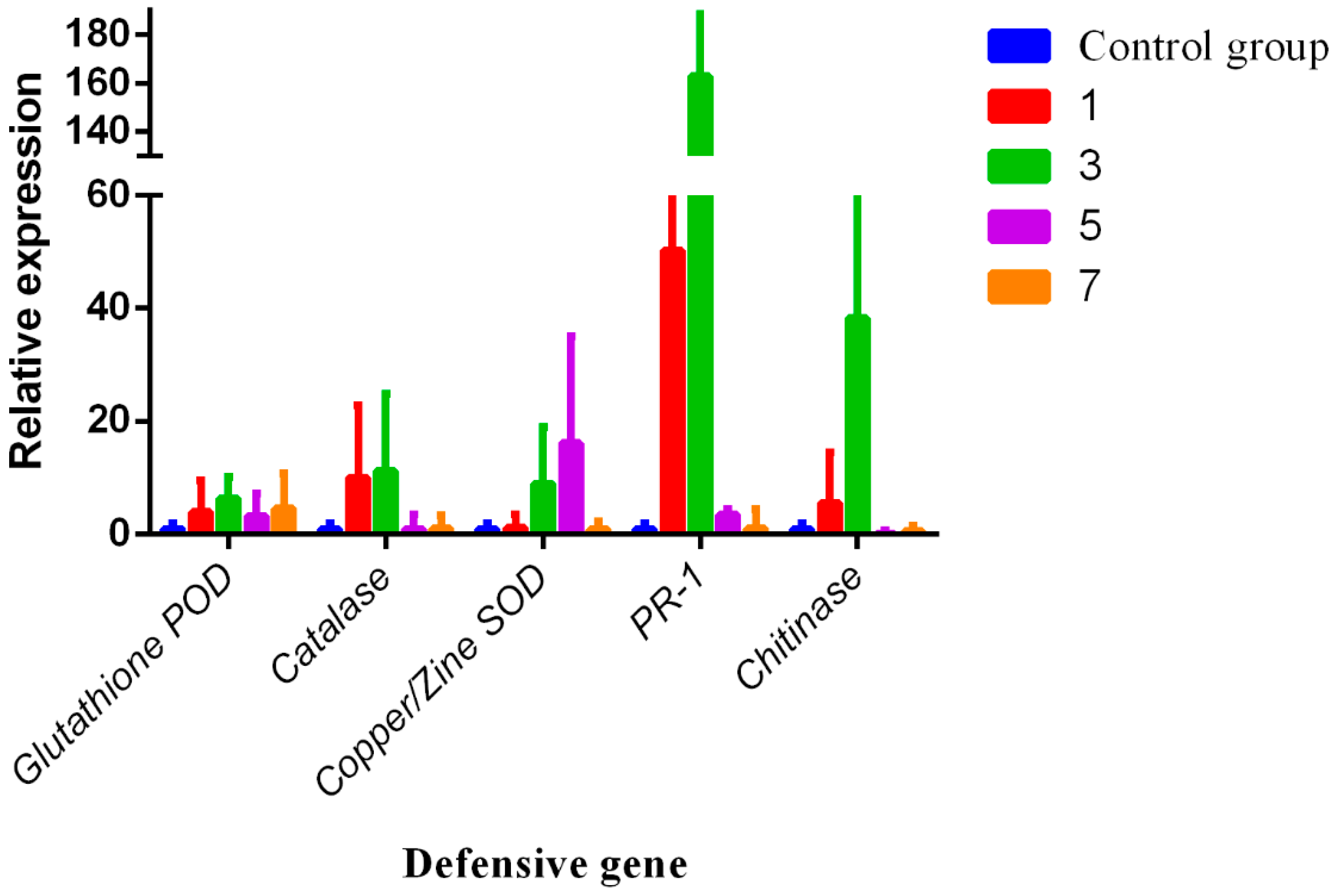
2.7. Gene Expression Analysis by Reverse Transcription-qPCR
3. Results
3.1. COS to Promote Growth Results
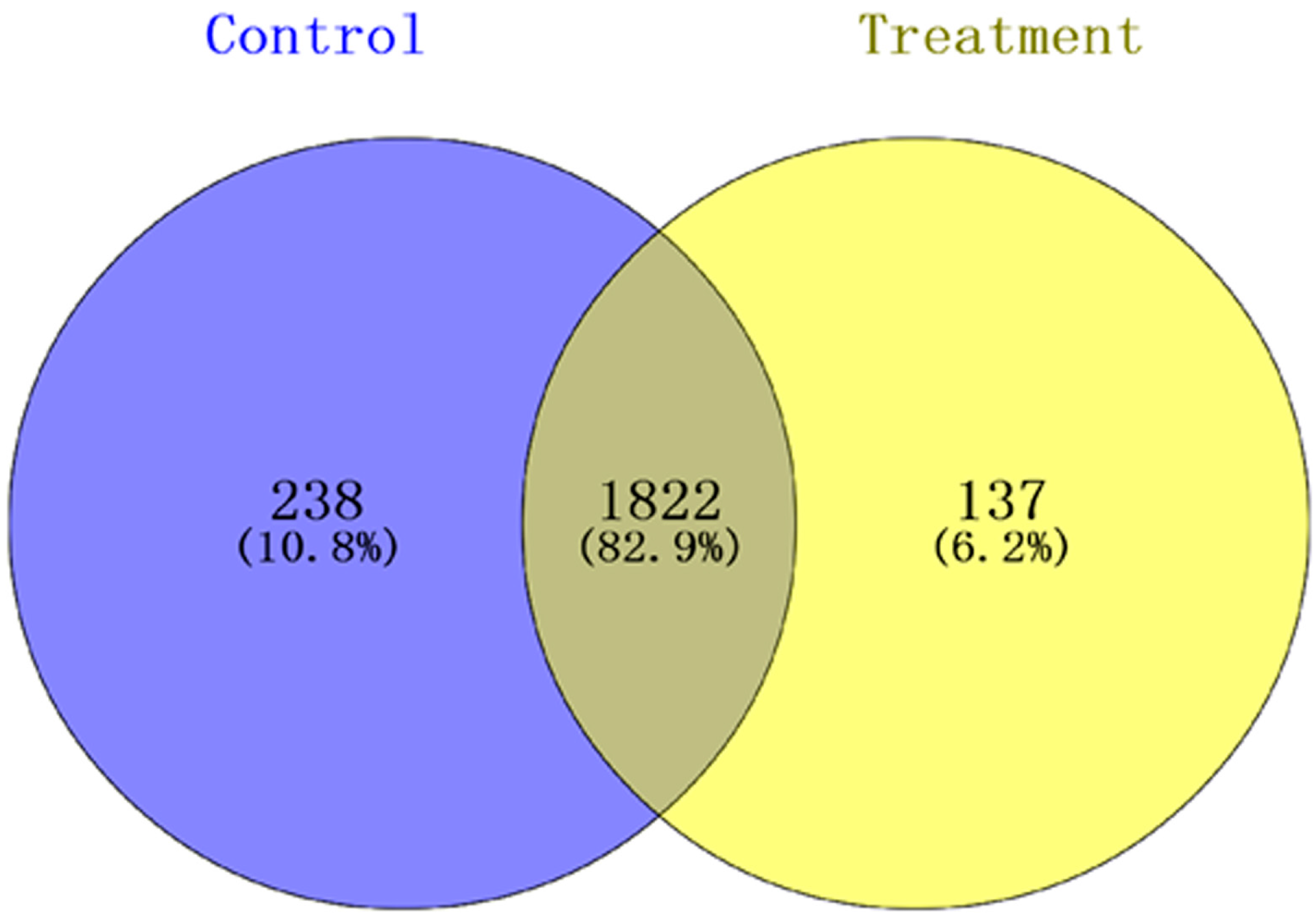
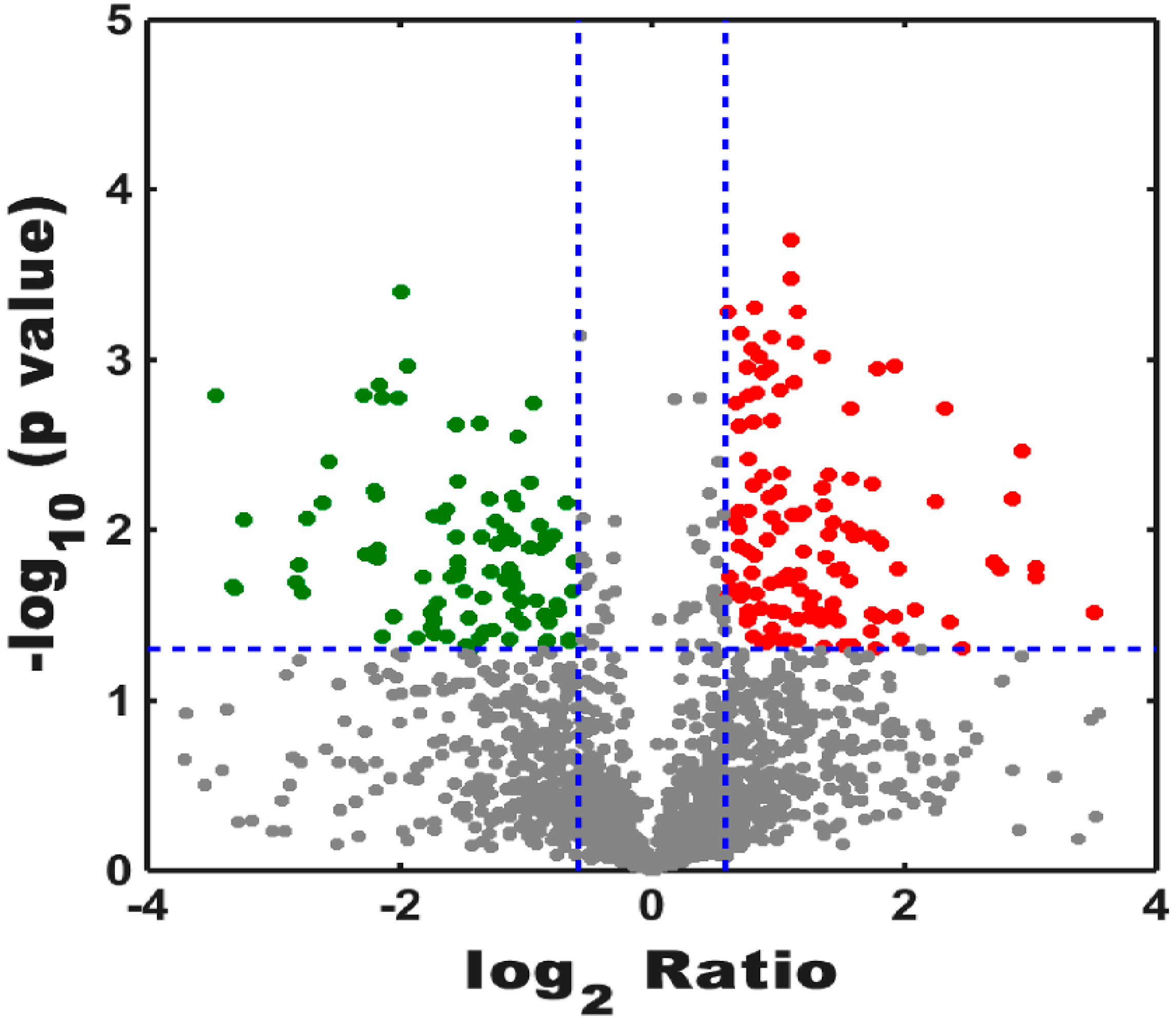
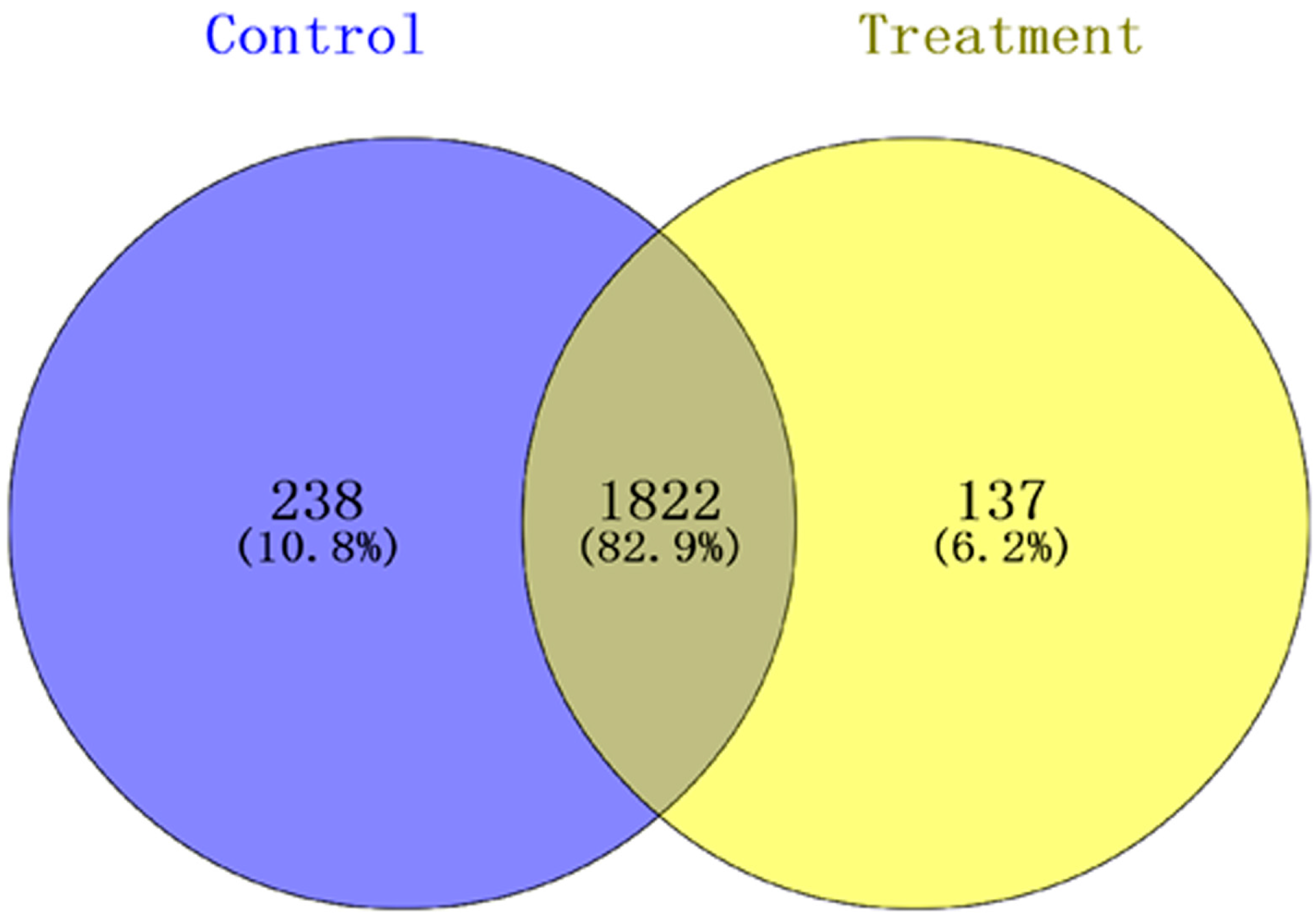
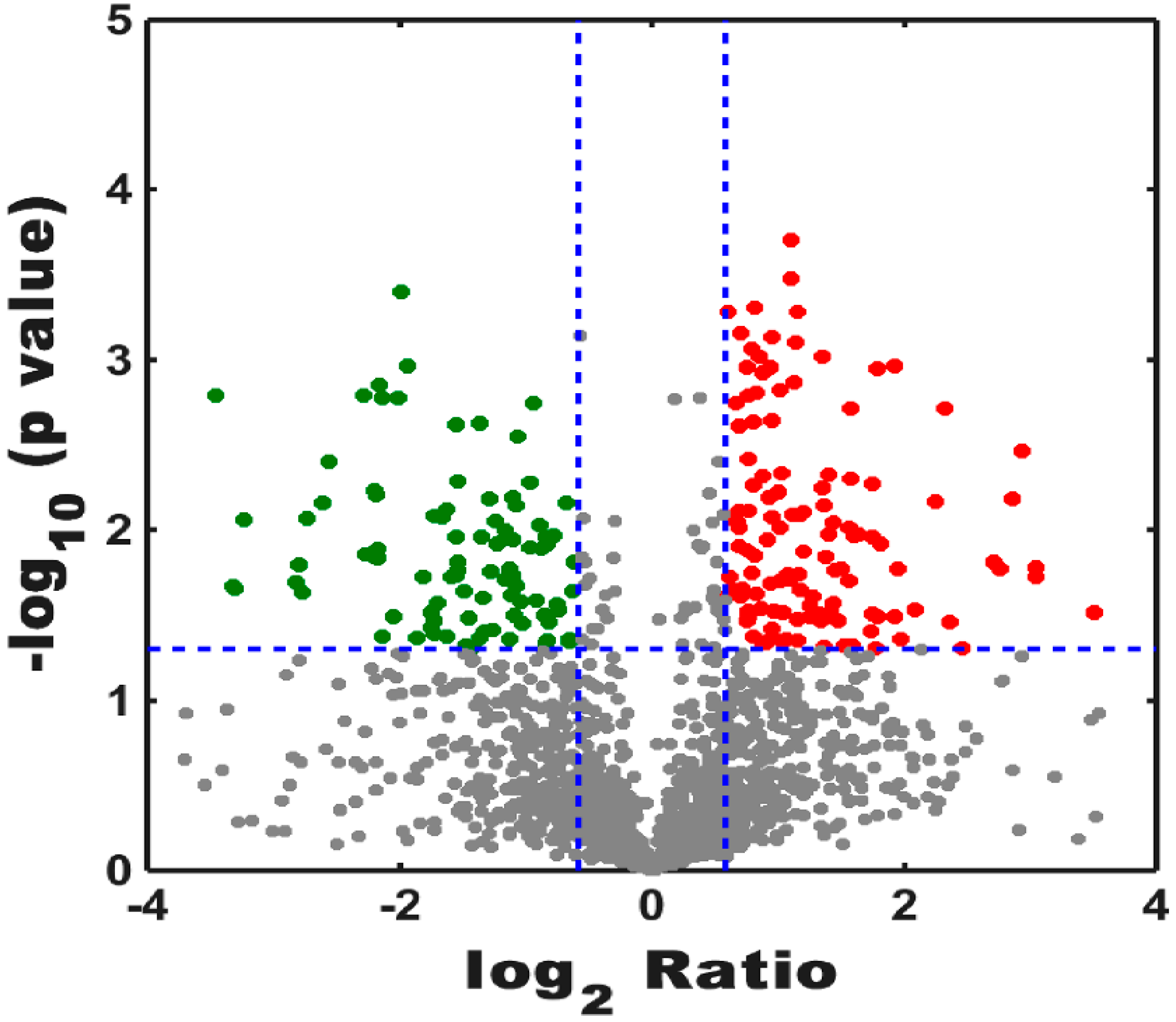
3.2. Label-Free ProteomicsComparative Analysis of Treatment Group and Control Group
3.2.1. Analysis of Proteomics
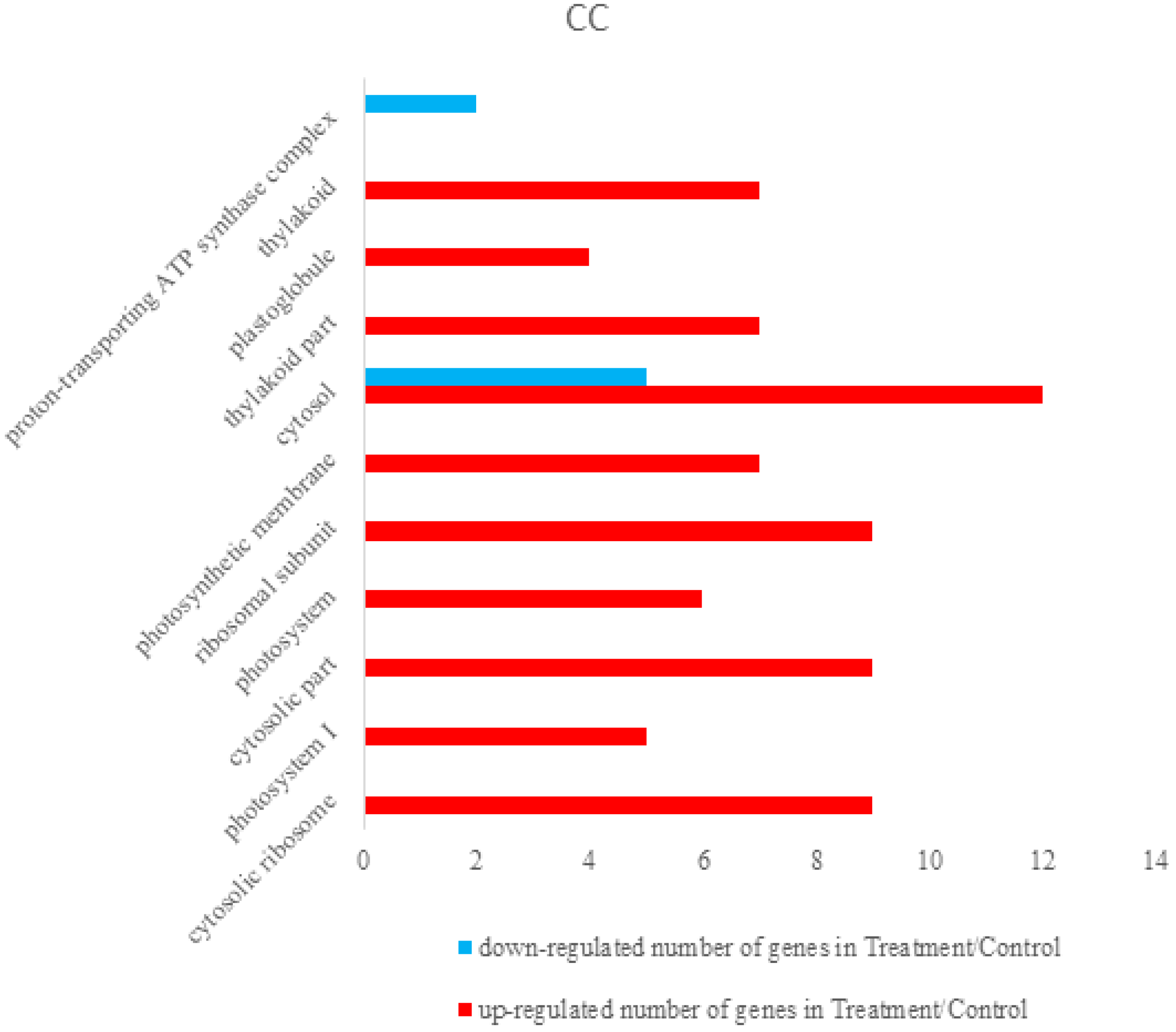
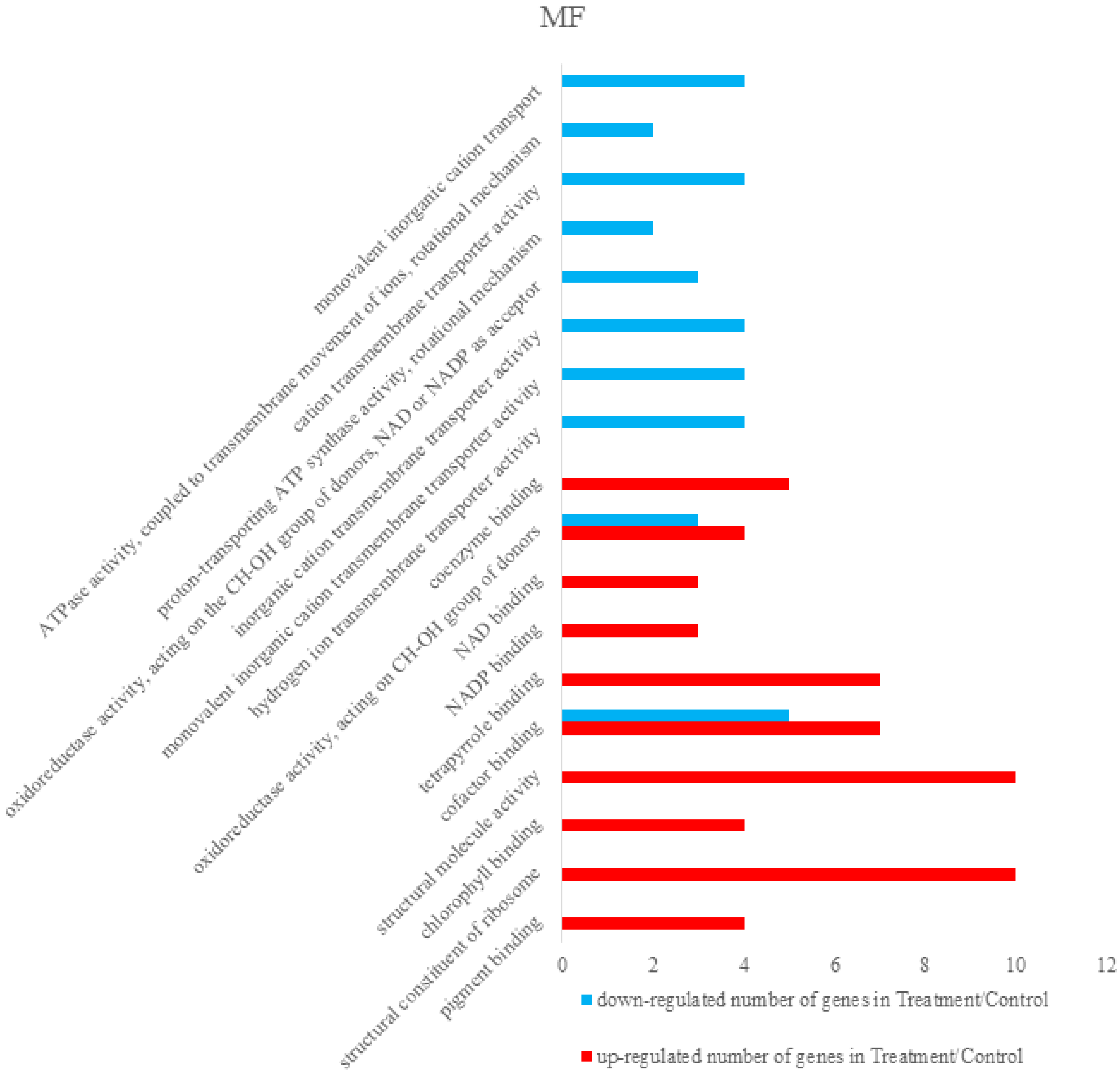
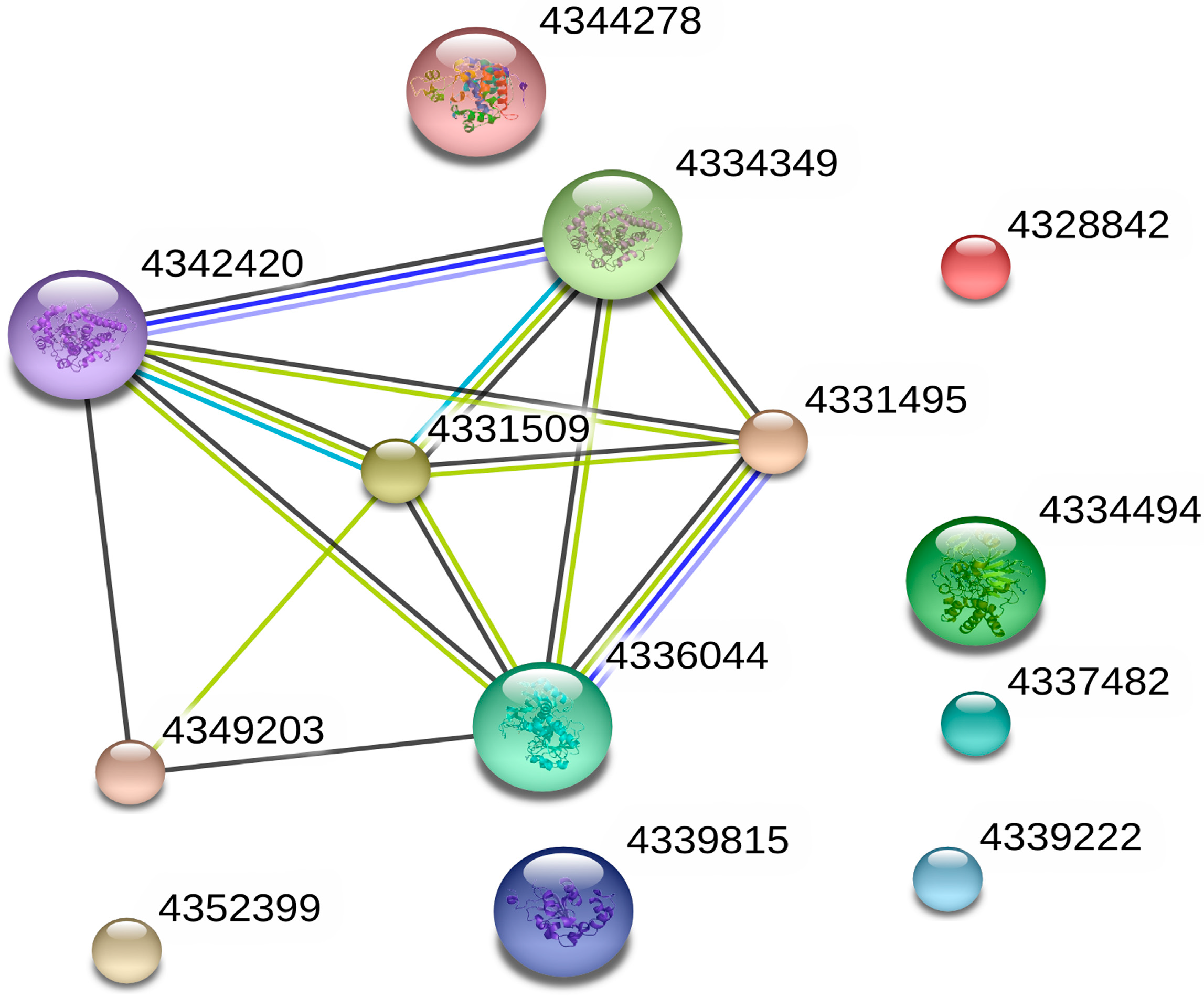
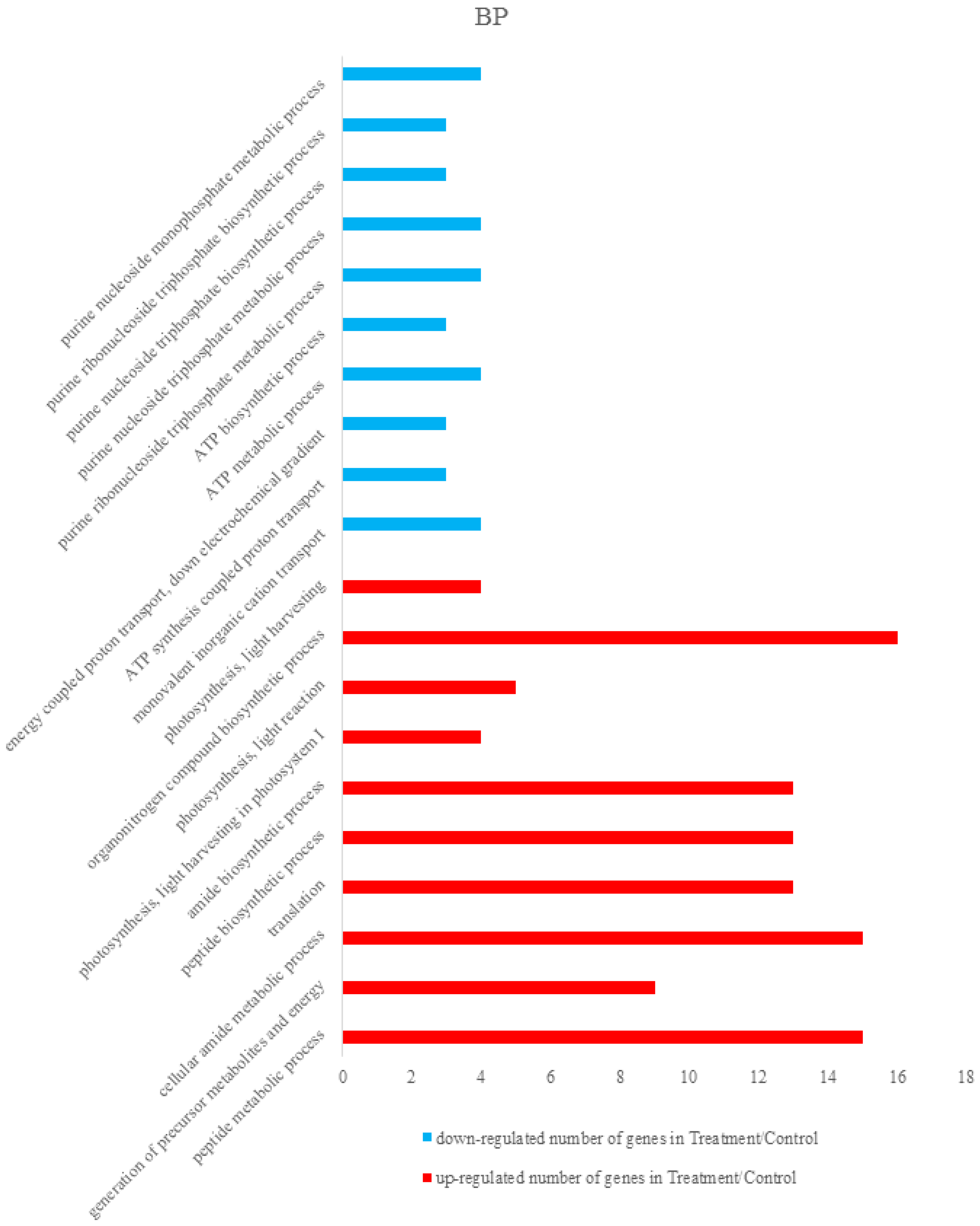
3.2.2. Bioinformatics Analysis of Control Groups and Treatment Groups
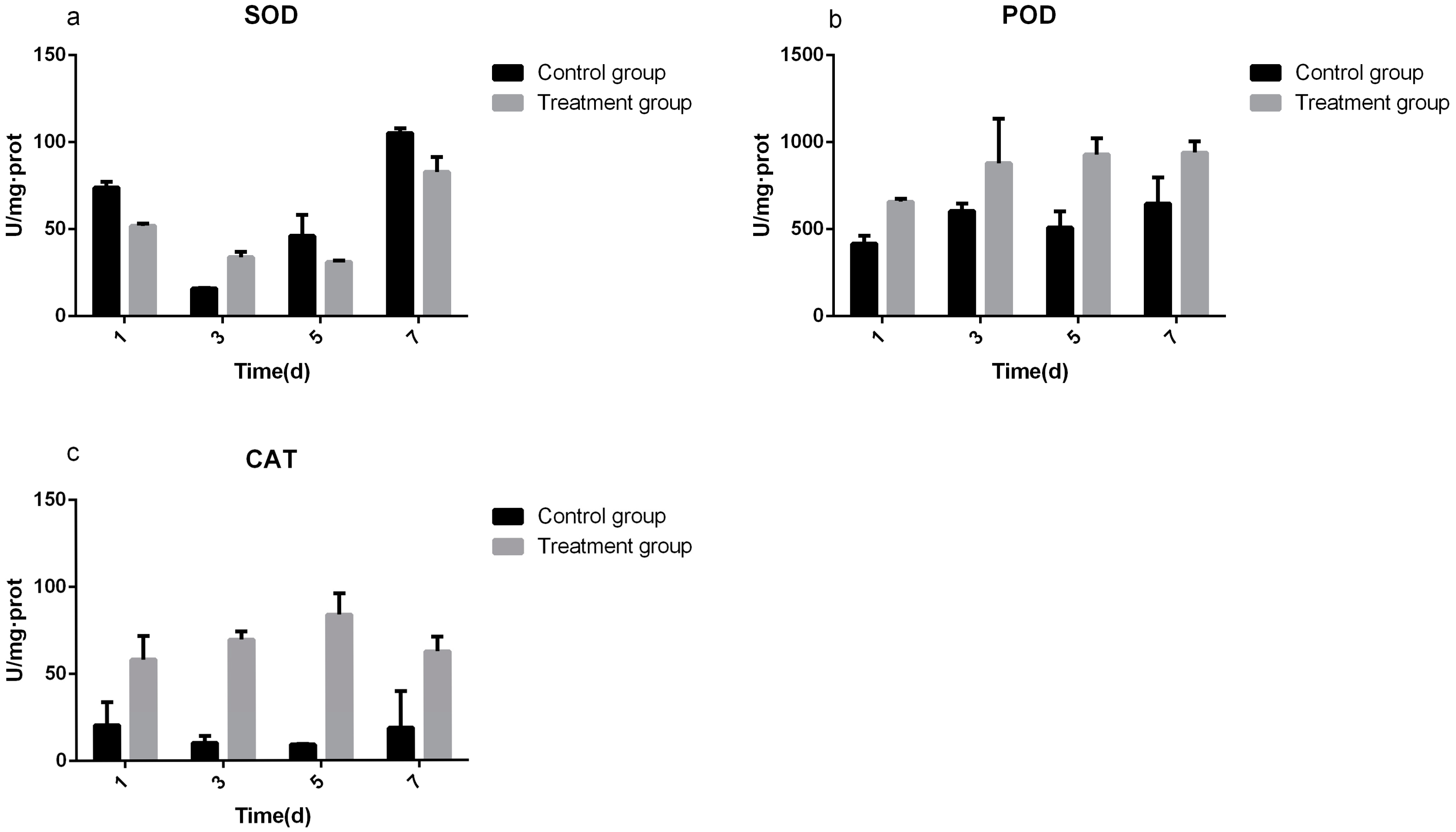
3.3. Determination of Defense Enzyme Activity
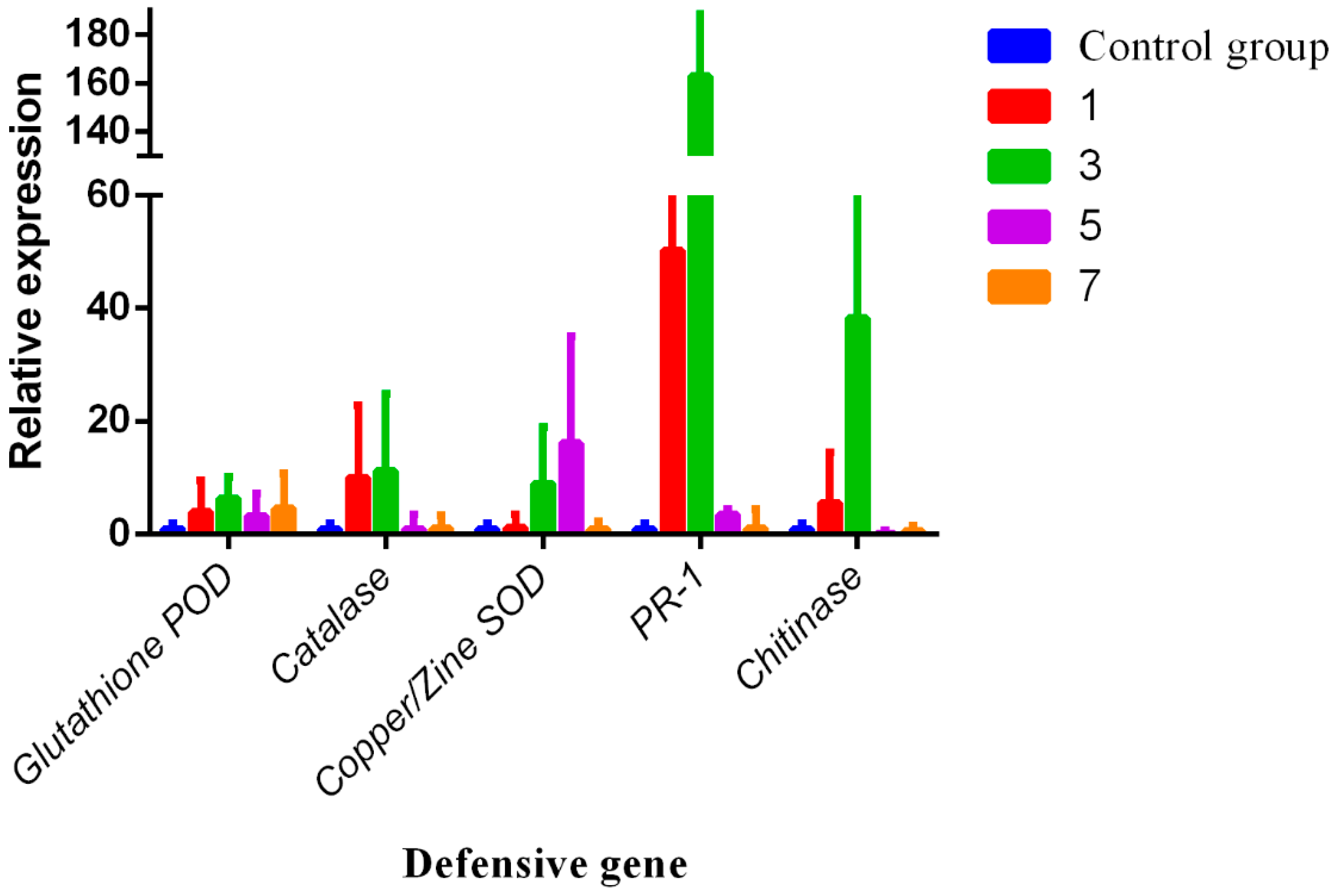
3.4. RNA Expression Levels of Chitinase, Catalase, Glutathione POD, Copper/Zine SOD, and PR-1
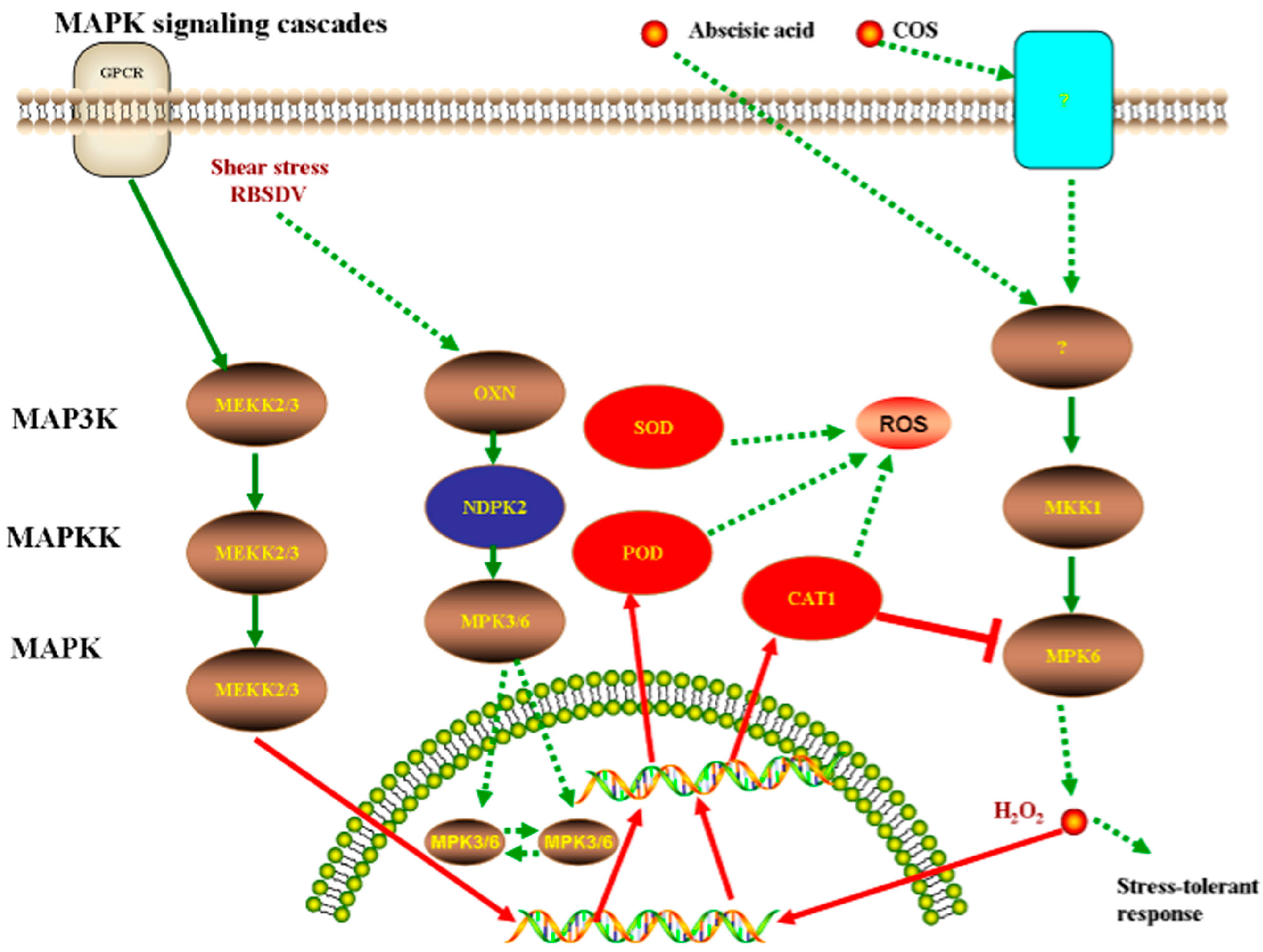
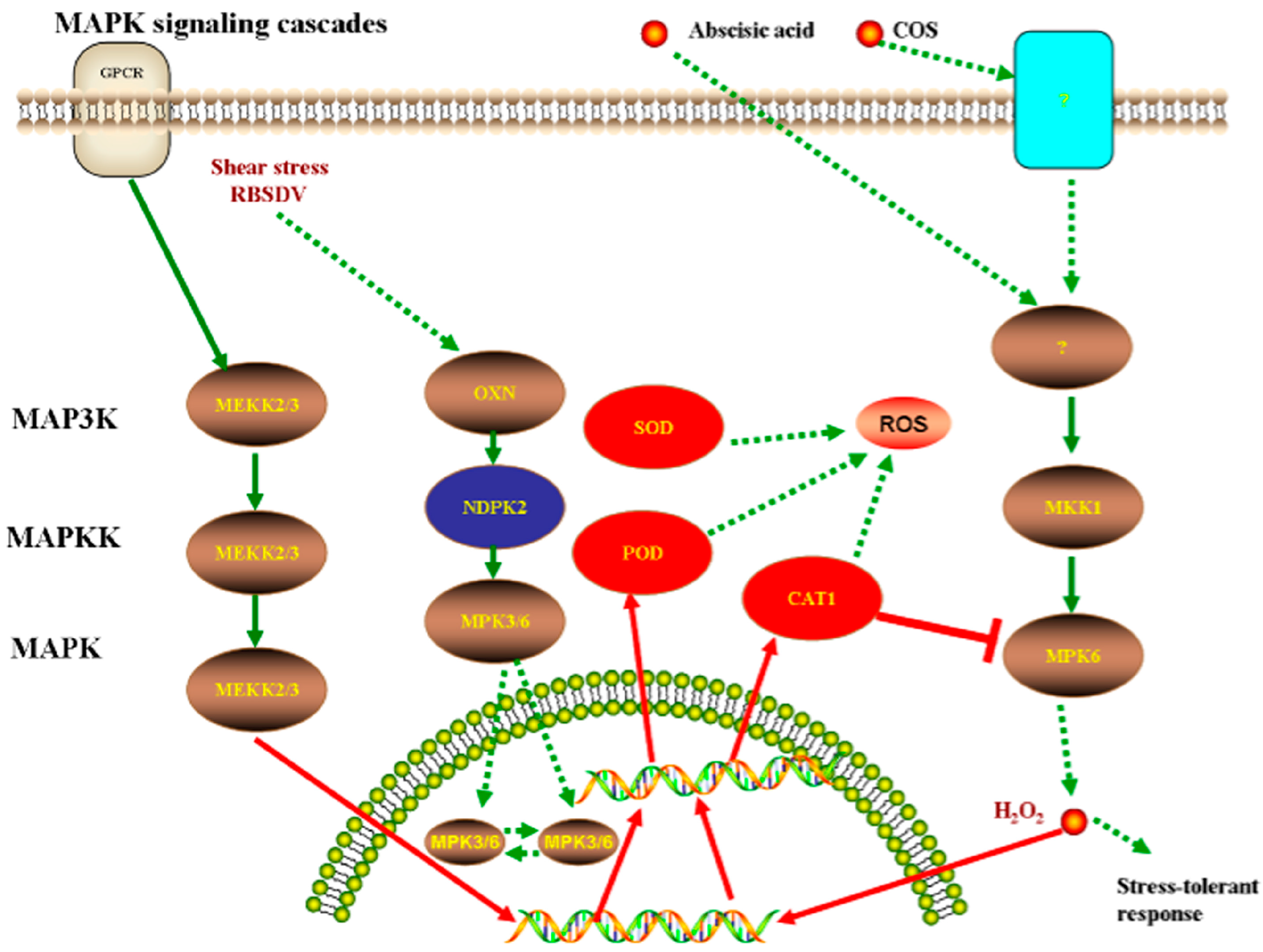
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, H.M.; Yang, J.; Chen, J.P.; Adams, M. A black-streaked dwarf disease on rice in China is caused by a novel fijivirus. Arch. Virol. 2008, 153, 1893–1898. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wen, J.; Cai, D.; Li, P.; Xu, D.; Zhang, S. Southern rice black-streaked dwarf virus: A new proposed Fijivirus species in the family Reoviridae. Chin. Sci. Bull. 2008, 53, 3677–3685. [Google Scholar] [CrossRef]
- Xu, H.X.; Zheng, X.S.; Yang, Y.J.; Tian, J.C.; Lu, Y.H.; Tan, K.H.; Heong, K.L.; Lu, Z.X. Methyl eugenol bioactivities as a new potential botanical insecticide against major insect pests and their natural enemies on rice (Oriza sativa). Crop Protect. 2015, 72, 144–149. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and Chitosan Preparation from Marine Sources. Structure, Properties and Applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed]
- Hadwiger, L.A.; Beckman, J.M. Chitosan as a component of pea-Fusarium solani interactions. Plant Physiol. 1980, 66, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhao, X.; Du, Y. Oligochitosan: A plant diseases vaccine—A review. Carbohydr. Polym. 2010, 82, 1–8. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, W.; Yin, H.; Zhao, X.; Du, Y. Oligochitosan induces programmed cell death in tobacco suspension cells. Carbohydr. Polym. 2012, 87, 2270–2278. [Google Scholar] [CrossRef]
- Wang, M.; Chen, Y.; Zhang, R.; Wang, W.; Zhao, X.; Du, Y.; Yin, H. Effects of chitosan oligosaccharides on the yield components and production quality of different wheat cultivars (Triticum aestivum L.) in Northwest China. Field Crops Res. 2015, 172, 11–20. [Google Scholar] [CrossRef]
- Li, S.-J.; Zhu, T.-H. Biochemical response and induced resistance against anthracnose (Colletotrichum camelliae) of camellia (Camellia pitardii) by chitosan oligosaccharide application. For. Pathol. 2013, 43, 67–76. [Google Scholar] [CrossRef]
- Yin, H.; Li, Y.; Zhang, H.Y.; Wang, W.X.; Lu, H.; Grevsen, K.; Zhao, X.; Du, Y. Chitosan Oligosaccharides–Triggered Innate Immunity Contributes to Oilseed Rape Resistance against Sclerotinia Sclerotiorum. Int. J. Plant Sci. 2013, 174, 722–732. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, K. Age-dependent variations of volatile emissions and inhibitory activity toward Botrytis cinerea and Fusarium oxysporum in tomato leaves treated with chitosan oligosaccharide. J. Plant Biol. 2009, 52, 332–339. [Google Scholar] [CrossRef]
- Cabrera, J.C.; Messiaen, J.; Cambier, P.; van Cutsem, P. Size, acetylation and concentration of chitooligosaccharide elicitors determine the switch from defence involving PAL activation to cell death and water peroxide production in Arabidopsis cell suspensions. Physiol. Plant. 2006, 127, 44–56. [Google Scholar] [CrossRef]
- Falcón, A.B.; Cabrera, J.C.; Costales, D.; Ramírez, M.A.; Cabrera, G.; Toledo, V.; Martínez-Téllez, M.A. The effect of size and acetylation degree of chitosan derivatives on tobacco plant protection against Phytophthora parasitica nicotianae. World J. Microbiol. Biotechnol. 2008, 24, 103–112. [Google Scholar] [CrossRef]
- Liénart, Y.; Gautier, C.; Domard, A. Isolation from Rubus cell-suspension cultures of a lectin specific for glucosamine oligomers. Planta 1991, 184, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Choi, H.; Suh, S.; Doo, I.S.; Oh, K.Y.; Choi, E.J.; Taylor, A.T.S.; Low, P.S.; Lee, Y. Oligogalacturonic acid and chitosan reduce stomatal aperture by inducing the evolution of reactive oxygen species from guard cells of tomato and Commelinacommunis. Plant Physiol. 1999, 121, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Vasconsuelo, A.; Marí, A.; Picotto, G.; Rodriguez-Talou, J.; Boland, R. Involvement of the PLC/PKC pathway in chitosan-induced anthraquinone production by Rubiatinctorum L. cell cultures. Plant Sci. 2003, 165, 429–436. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Marí, A.; Boland, R. Signal transduction events mediating chitosan stimulation of anthraquinone synthesis in Rubiatinctorum. Plant Sci. 2004, 166, 405–413. [Google Scholar] [CrossRef]
- Vasconsuelo, A.; Morelli, S.; Picotto, G.; Giulietti, A.M.; Boland, R. Intracellular calcium mobilization: A key step for chitosan-induced anthraquinone production in Rubiatinctorum L. Plant Sci. 2005, 169, 712–720. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Rakwal, R.; Tamogami, S.; Yonekura, M.; Kubo, A.; Saji, H. Chitosan activates defense/stress response(s) in the leaves of Oryza sativa seedlings. Plant Physiol. Biochem. 2002, 40, 1061–1069. [Google Scholar] [CrossRef]
- Boonlertnirun, S.; Boonraung, C.; Suvanasara, R. Application of chitosan in rice production. J. Met. Mater. Miner. 2008, 18, 47–52. [Google Scholar]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.P.; Tang, T.; Zhao, M.P. Control efficacy on southern rice black-streaked dwarf virus of oligosaccharins·plant activator protein and its effects on growth promotion and yield increase of rice. Agrochemicals 2015, 54, 606–609. [Google Scholar]
- Peters, R.J. Uncovering the complex metabolic network underlying diterpenoid phytoalexin biosynthesis in rice and other cereal crop plants. Phytochemistry 2006, 67, 2307–2317. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sengupta, D.; Kannan, M.; Reddy, A.R. A root proteomics-based insight reveals dynamic regulation of root proteins under progressive drought stress and recovery in Vigna radiata (L.) Wilczek. Planta 2011, 233, 1111–1127. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.O. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, D109–D114. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Thissen, D.; Steinberg, L.; Kuang, D. Quick and easy implementation of the Benjamini-Hochberg procedure for controlling the false positive rate in multiple comparisons. J. Educ. Behav. Stat. 2002, 27, 77–83. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. B 1995, 57, 289–300. [Google Scholar]
- Shah, K.; Nahakpam, S. Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol. Biochem. 2012, 57, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, G. Effect of Drought Stress on Activities of Cell Defense Enzymes and Lipid Peroxidation in Glycyrrhiza uralensis Seedings. Acta Ecol. Sin. 2001, 22, 503–507. [Google Scholar]
- Zhang, C.; Liu, Y.; Liu, L.; Lou, Z.; Zhang, H.; Miao, H.; Hu, X.; Pang, Y.; Qiu, B. Rice black streaked dwarf virus P9–1, an α-helical protein, self-interacts and forms viroplasms in vivo. J. Gen. Virol. 2008, 89, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Yang, J.; Xie, L.; Li, J.; Song, X.J.; Chen, J.P.; Zhang, H.M. P5–2 of rice black-streaked dwarf virus is a non-structural protein targeted to chloroplasts. Arch. Virol. 2015, 160, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xue, J.; Zhang, H.-M.; Yang, J.; Xie, L.; Chen, J.-P. Characterization of homologous and heterologous interactions between viroplasm proteins P6 and P9–1 of the fijivirus southern rice black-streaked dwarf virus. Arch. Virol. 2015, 160, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xie, L.; Andika, I.B.; Tan, Z.; Chen, J. Non-structural protein P6 encoded by rice black-streaked dwarf virus is recruited to viral inclusion bodies by binding to the viroplasm matrix protein P9–1. J. Gen. Virol. 2013, 94, 1908–1916. [Google Scholar] [CrossRef] [PubMed]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.M.; Shin, S.H.; Kim, K.S.; Kim, Y.C.; Eun, M.Y.; Cho, B.H. Enhanced expression of a gene encoding a nucleoside diphosphate kinase 1 (OsNDPK1) in rice plants upon infection with bacterial pathogens. Mol. Cells 2004, 18, 390–395. [Google Scholar] [PubMed]
- Yao, S.; Wang, Y.; Yang, T.; Kou, F.; Lu, W.; Xiao, K. Expression pattern and function of wheat mitogen-activated protein kinase (MPK) cascade genes under micronutrient-deprived conditions. Acta Physiol. Plant. 2016, 39, 40. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, J.; Li, Y.; Rong, X.; Sun, J.; Sun, T.; Li, M.; Wang, L.; Feng, Y.; Chai, R.; et al. Expression of TaWRKY44, a wheat WRKY gene, in transgenic tobacco confers multiple abiotic stress tolerances. Front. Plant Sci. 2015, 6, 615. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.Q.; Hu, J.B.; Liu, J.H. Cloning and characterization of FcWRKY40, A WRKY transcription factor from Fortunella crassifolia linked to oxidative stress tolerance. Plant Cell Tissue Organ Cult. 2014, 119, 197–210. [Google Scholar] [CrossRef]
- Zhang, X.H.; Rao, X.L.; Shi, H.T.; Li, R.J.; Lu, Y.T. Overexpression of a cytosolic glyceraldehyde-3-phosphate dehydrogenase gene OsGAPC3 confers salt tolerance in rice. Plant Cell Tissue Organ Cult. 2011, 107, 1. [Google Scholar] [CrossRef]
- Zhao, F.; Zhang, H. Salt and paraquat stress tolerance results from co-expression of the Suaeda salsa glutathione S-transferase and catalase in transgenic rice. Plant Cell Tissue Organ Cult. 2006, 86, 349–358. [Google Scholar] [CrossRef]
- Jia, X.; Meng, Q.; Zeng, H.; Wang, W.; Yin, H. Chitosan oligosaccharide induces resistance to Tobacco mosaic virus in Arabidopsis via the salicylic acid-mediated signalling pathway. Sci. Rep. 2016, 6, 26144. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| β-actin | 5′-TCCTCCGTGGAGAAGAGCTA-3′ | 5′-GCAATGCCAGGGAACATAGT-3′ |
| Chitinase | 5′-CTACACGTACGACGCCTTCA-3′ | 5′-GAAGCAGTAACCCCACTGGA-3′ |
| Catalase | 5′-AAGCCGAGCATGTAAGGAGA-3′ | 5′-ACACGAATTGTGCGGTGATA-3′ |
| Ascorbate POD | 5′-CAAGGAGGAGATACCCACCA-3′ | 5′-TAGGTGGTCAGAACCCTTGG-3′ |
| PR-1 | 5′-CAGTGGTACGACCACGACAG-3′ | 5′-GGCGAGTAGTTGCAGGTGAT-3′ |
| Copper/Zine SOD | 5′-TTTTCCAGTCCCCTTCCTCT-3′ | 5′-AGCCGTGAAGTCCAGGAGTA-3′ |
| Buds Long | ||||||||
| Repeated Numbers | 1 | 2 | 3 | 4 | 5 | 6 | 7 | Average Value |
| CK | 1.6 | 1.5 | 1.4 | 1.3 | 0.9 | 0.9 | 0.9 | 1.21 ± 0.39 |
| COS(50µg/mL) | 2.5 | 2.4 | 3.0 | 2.4 | 2.0 | 2.0 | 2.7 | 2.43 ± 0.33 |
| COS (100µg/mL) | 1.8 | 1.7 | 1.5 | 2.0 | 1.8 | 1.5 | 1.5 | 1.69 ± 0.18 |
| COS (200µg/mL) | 3.5 | 3.0 | 4.0 | 2.5 | 2.5 | 2.0 | 2.5 | 2.86 ± 0.64 |
| Root Long | ||||||||
| Repeated Numbers | 1 | 2 | 3 | 4 | 5 | 6 | 7 | Average Value |
| CK | 1.4 | 1.7 | 2.0 | 2.3 | 2.0 | 1.7 | 2.2 | 1.90 ± 0.29 |
| COS (50µg/mL) | 2.5 | 2.6 | 4.0 | 3.0 | 3.1 | 3.6 | 3.5 | 3.19 ± 0.51 |
| COS (100µg/mL) | 4.0 | 2.5 | 2.5 | 4.0 | 2.0 | 3.5 | 2.5 | 3.00 ± 0.76 |
| COS (200µg/mL) | 1.5 | 1.8 | 2.0 | 1.8 | 2.0 | 2.0 | 2.0 | 1.87 ± 0.17 |
| Protein ID | Protein Names | Organism | Length | LogRatio | LogP | Sig |
|---|---|---|---|---|---|---|
| PP2A2_ORYSJ | Serine/threonine-protein phosphatase PP2A-2 catalytic subunit (EC 3.1.3.16) | Oryza sativa subsp. Japónica (Rice) | 307 | 2.71344 | 1.809716 | 1 |
| GLO1_ORYSJ | Peroxisomal (S)-2-hydroxy-acid oxidase GLO1 (EC 1.1.3.15) (Glycolate oxidase 1) (GOX 1) (OsGLO1) (Short chain alpha-hydroxy acid oxidase GLO1) | Oryza sativa subsp. japonica (Rice) | 369 | 1.100617 | 3.704594 | 1 |
| GLO5_ORYSJ | Peroxisomal (S)-2-hydroxy-acid oxidase GLO5 (EC 1.1.3.15) (Glycolate oxidase 5) (GOX 5) (OsGLO5) (Short chain alpha-hydroxy acid oxidase GLO5) | Oryza sativa subsp. Japónica (Rice) | 369 | 0.68682 | 2.115814 | 1 |
| PER1_ORYSJ | Peroxidase 1 (EC 1.11.1.7) | Oryza sativa subsp. Japónica (Rice) | 326 | 1.197095 | 2.103695 | 1 |
| V5LEK5_9REOV | P9-1 | Southern rice black-streaked dwarf virus | 347 | −3.45753 | 2.791139 | −1 |
| Q9ZRI9_ORYSJ | Catalase (EC 1.11.1.6) | Oryza sativa subsp. Japónica (Rice) | 492 | 0.883992 | 2.928169 | 1 |
| Q7XSV2_ORYSJ | Peroxidase (EC 1.11.1.7) | Oryza sativa subsp. Japónica (Rice) | 346 | 1.18305 | 1.649601 | 1 |
| Q5U1F5_ORYSJ | Peroxidase (EC 1.11.1.7) | Oryza sativa subsp. Japónica (Rice) | 344 | −1.7243 | 1.38769 | −1 |
| H9BJU7_9REOV | P5 | Southern rice black-streaked dwarf virus | 939 | −4.47428 | 2.610644 | −1 |
| Q6ER49_ORYSJ | Peroxidase (EC 1.11.1.7) | Oryza sativa subsp. japonica (Rice) | 321 | 0.899285 | 1.336709 | 1 |
| Q7F1U0_ORYSJ | Peroxidase (EC 1.11.1.7) | Oryza sativa subsp. japonica (Rice) | 317 | −0.97083 | 2.274778 | −1 |
| Q9SNK3_ORYSJ | Glyceraldehyde-3-phosphate dehydrogenase (EC 1.2.1.-) | Oryza sativa subsp. Japónica (Rice) | 444 | 0.75745 | 2.954426 | 1 |
| Q7X8A1_ORYSJ | Glyceraldehyde-3-phosphate dehydrogenase (EC 1.2.1.-) | Oryza sativa subsp. Japónica (Rice) | 402 | 0.94783 | 2.074914 | 1 |
| Q945W5_ORYSJ | Glutathione S-transferase GSTU6, putative, expressed (Os10g0530500 protein) (Putative glutathione S-transferase) (Putative glutathione S-transferase OsGSTU13) | Oryza sativa subsp. Japónica (Rice) | 233 | 1.129885 | 2.872945 | 1 |
| Q5TKF4_ORYSJ | Nucleoside diphosphate kinase (EC 2.7.4.6) | Oryza sativa subsp. Japónica (Rice) | 239 | - | - | 0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, A.; Yu, L.; Chen, Z.; Zhang, S.; Shi, J.; Zhao, X.; Yang, Y.; Hu, D.; Song, B. Label-Free Quantitative Proteomic Analysis of Chitosan Oligosaccharide-Treated Rice Infected with Southern Rice Black-Streaked Dwarf Virus. Viruses 2017, 9, 115. https://doi.org/10.3390/v9050115
Yang A, Yu L, Chen Z, Zhang S, Shi J, Zhao X, Yang Y, Hu D, Song B. Label-Free Quantitative Proteomic Analysis of Chitosan Oligosaccharide-Treated Rice Infected with Southern Rice Black-Streaked Dwarf Virus. Viruses. 2017; 9(5):115. https://doi.org/10.3390/v9050115
Chicago/Turabian StyleYang, Anming, Lu Yu, Zhuo Chen, Shanxue Zhang, Jing Shi, Xiaozhen Zhao, Yuanyou Yang, Deyu Hu, and Baoan Song. 2017. "Label-Free Quantitative Proteomic Analysis of Chitosan Oligosaccharide-Treated Rice Infected with Southern Rice Black-Streaked Dwarf Virus" Viruses 9, no. 5: 115. https://doi.org/10.3390/v9050115