A Satellite-Based Model for Simulating Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
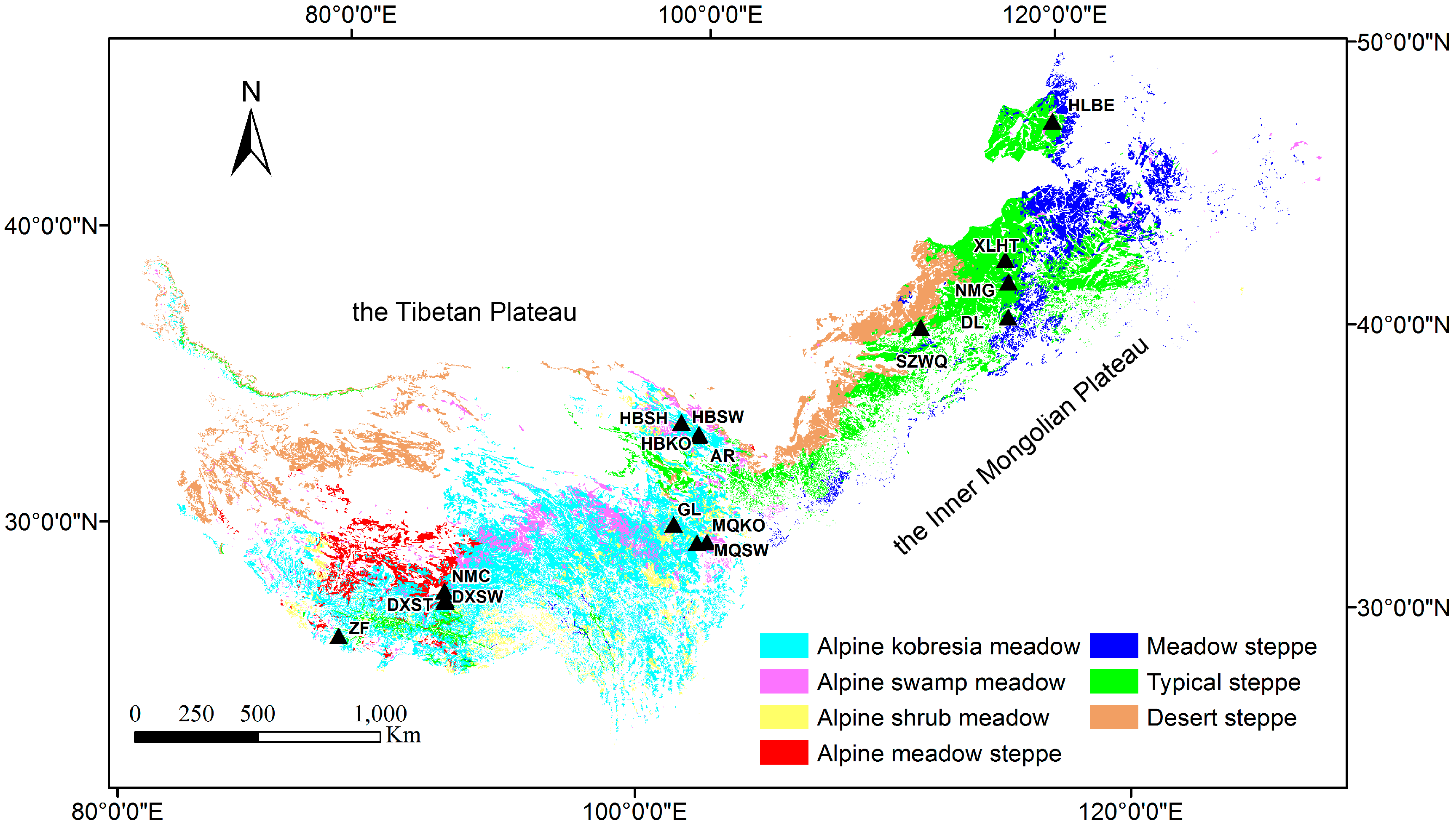
2.1. Study Area
2.2. Data
2.2.1. Flux-Tower Data
2.2.2. Remote-Sensing Data
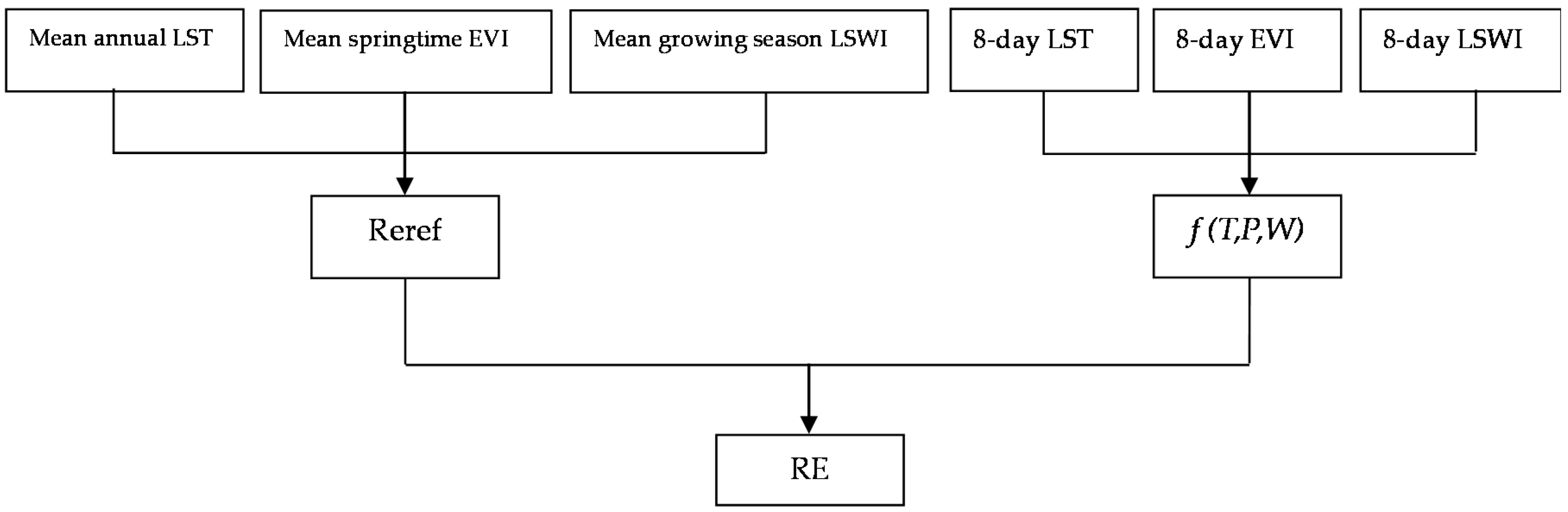
2.3. Model
2.3.1. Representation of Spatial Variability of RE
2.3.2. Representation of Temporal Variability of RE
2.4. Model Parameterization and Validation
3. Results
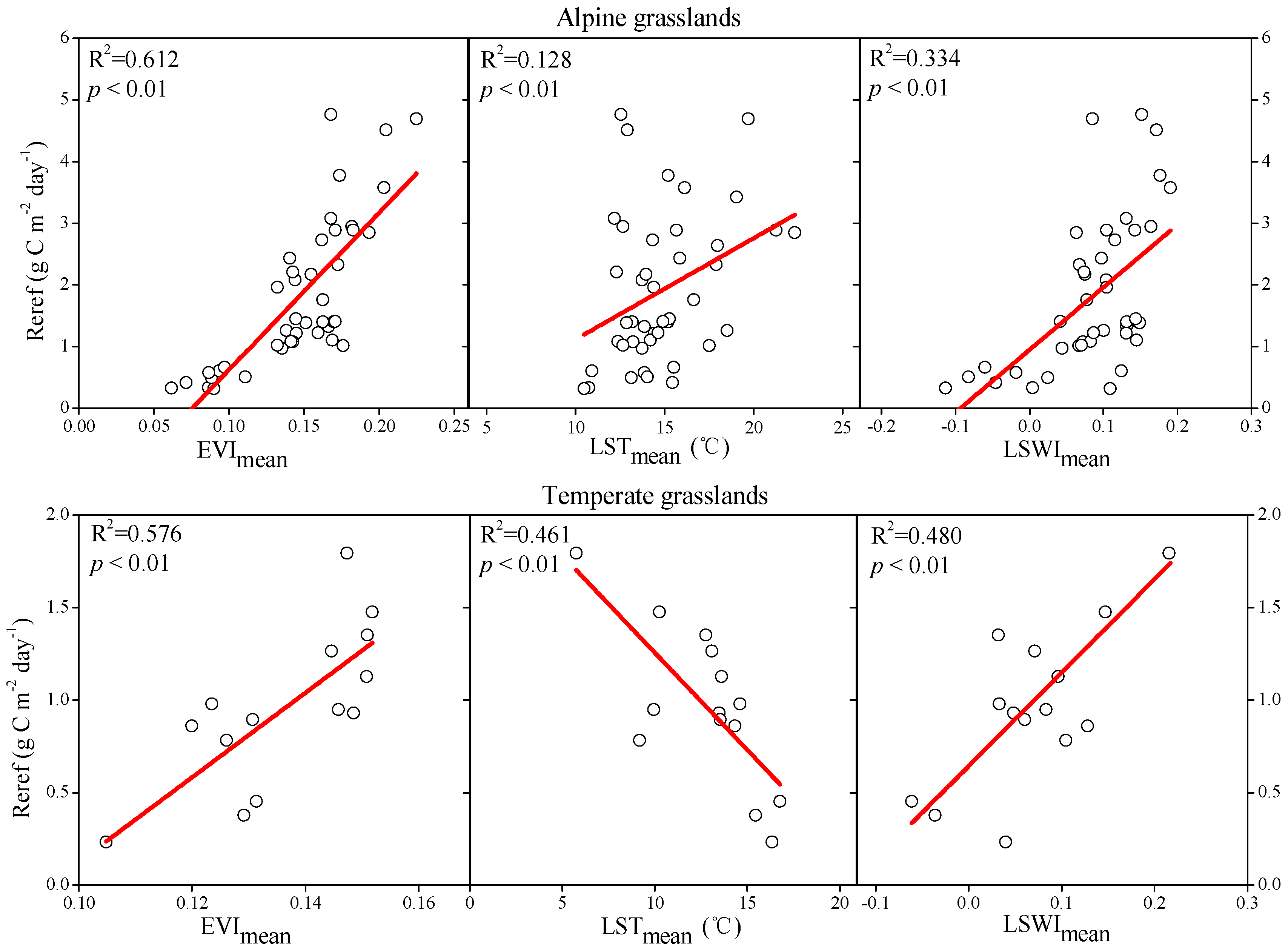
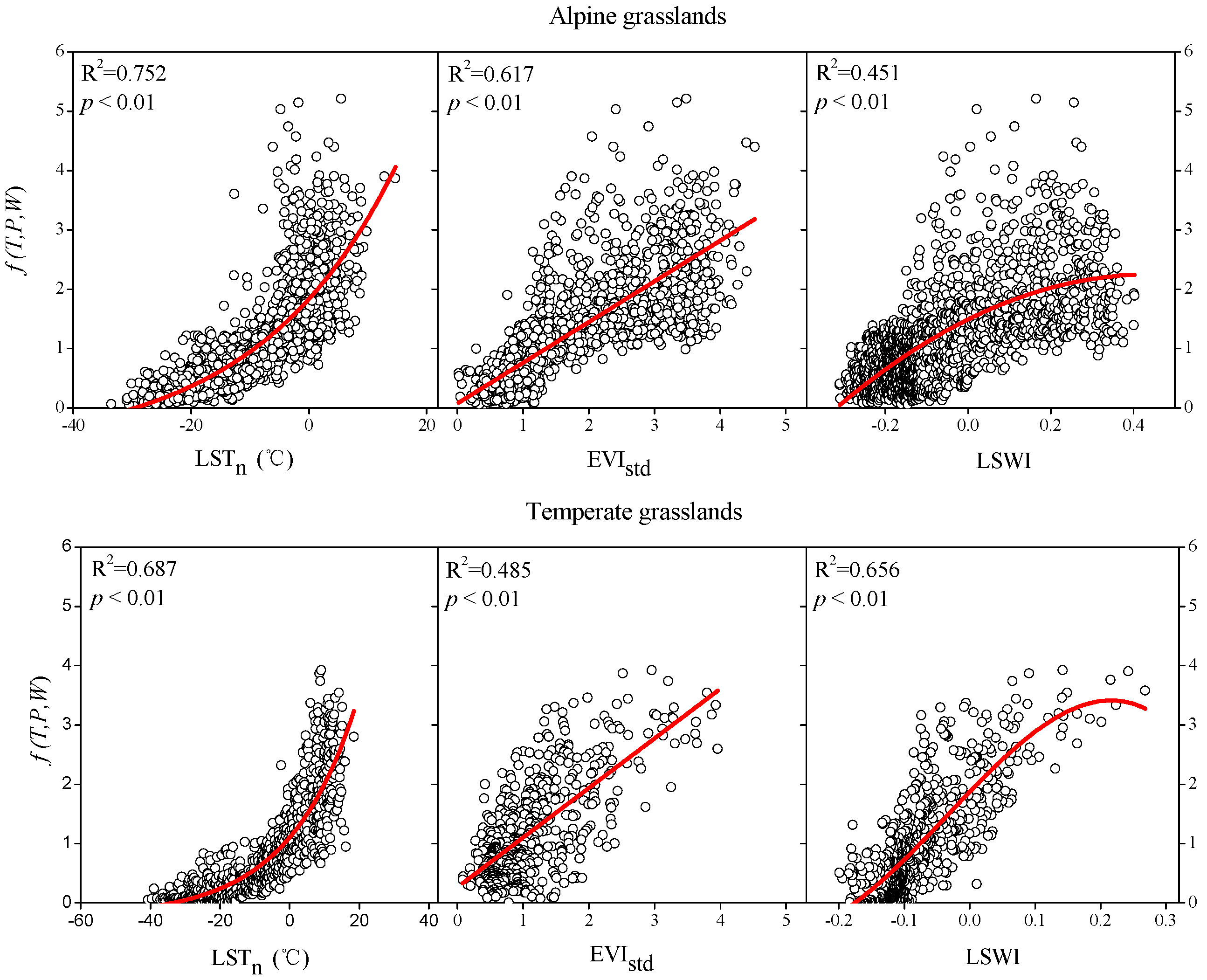
3.1. Quantitative Relationships between RE and Biotic and Climatic Factors
3.1.1. Quantitative Relationships between Reref and Biotic and Climatic Factors
3.1.2. Quantitative Relationships between and Biotic and Climatic Factors
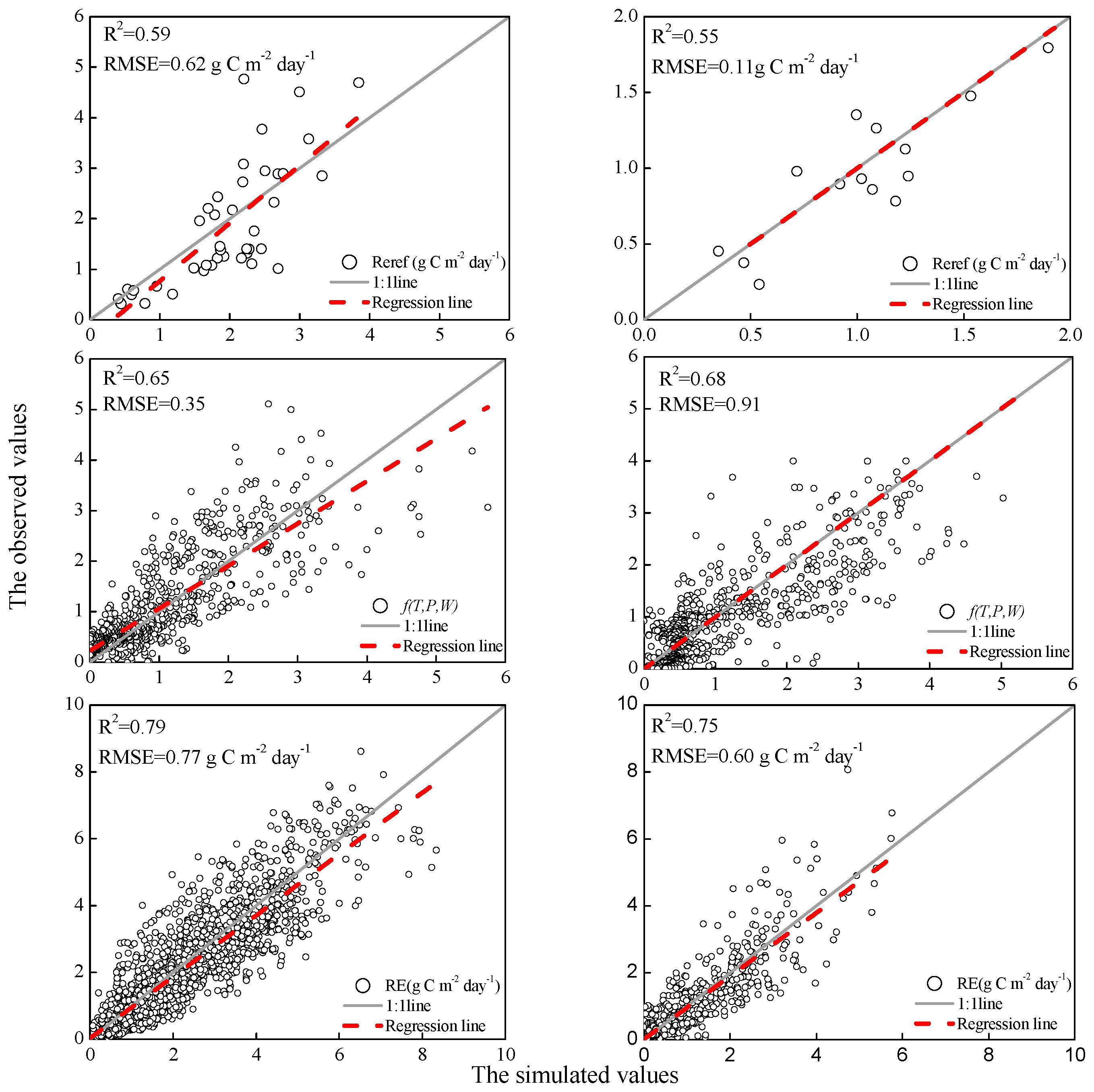
3.2. Model Parameterization and Validation
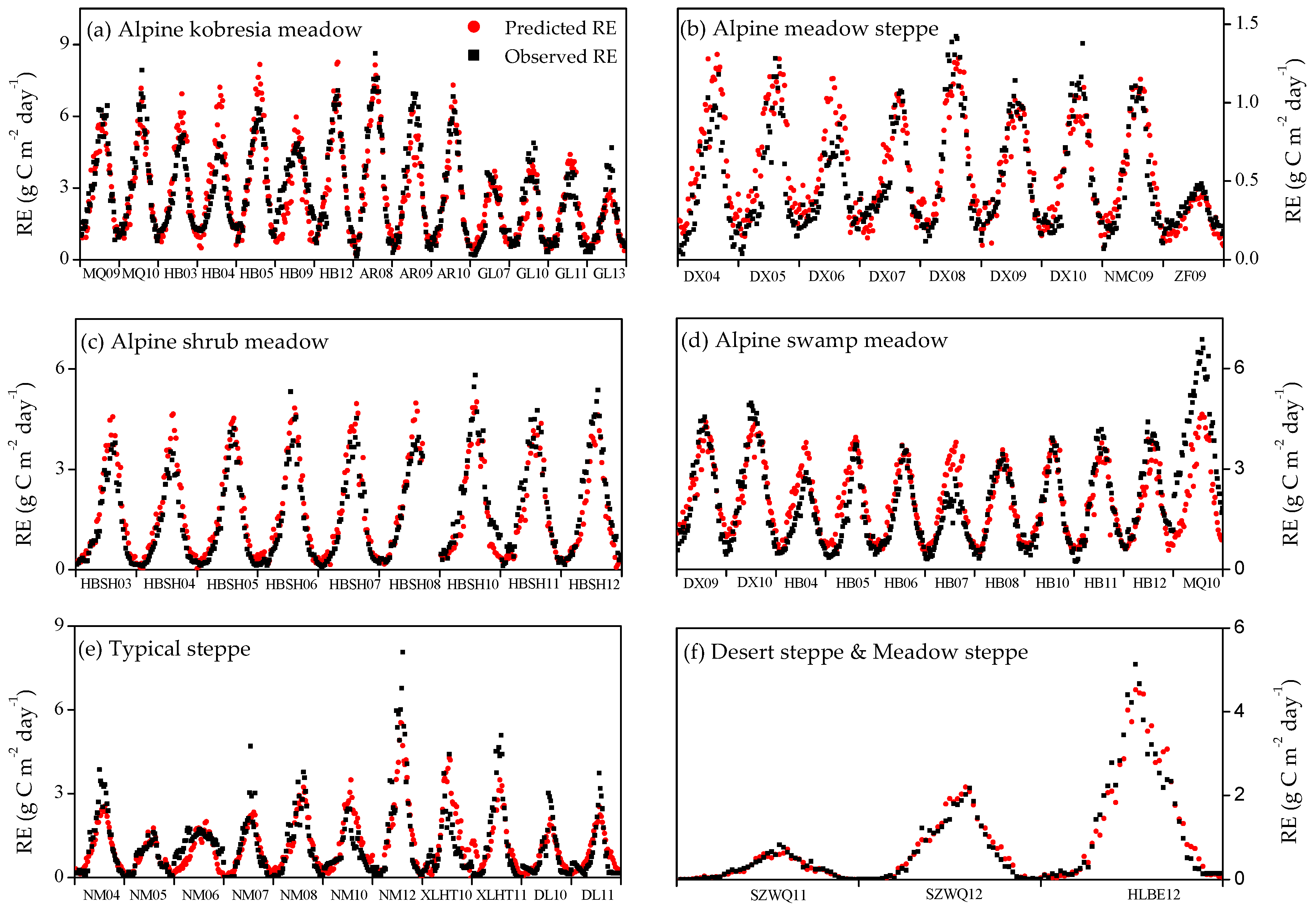
3.3. Model Simulation for Seasonal and Inter-Annual Dynamics of RE at the Site Scale
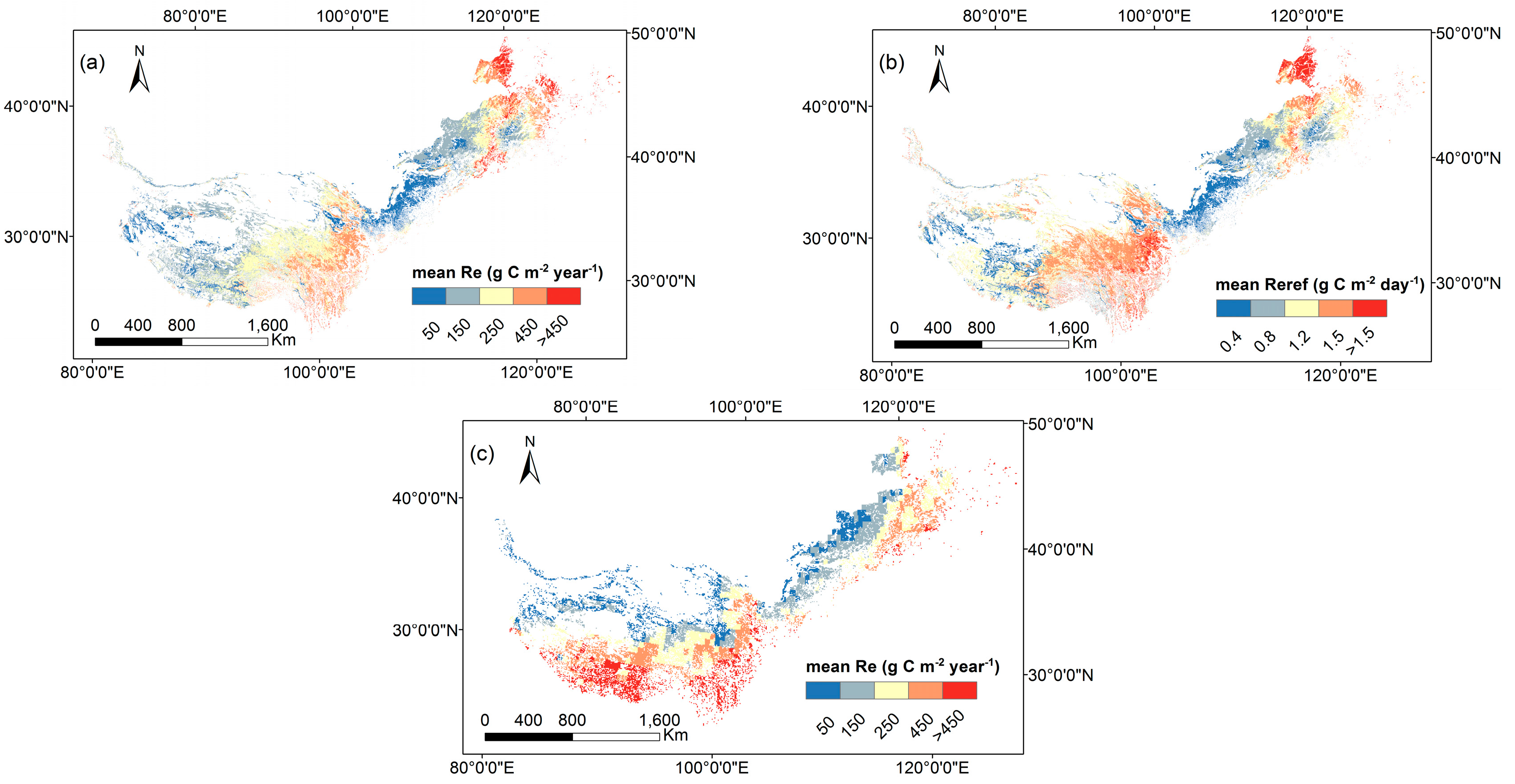
3.4. Model Simulation for Spatial Patterns of RE at the Regional Scale
4. Discussion
4.1. Biotic and Climatic Control over RE
4.2. Model Evaluation
4.3. Model Applications and Limitations
5. Conclusions and Implications
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Adams, J.M.; Faure, H.; Faure-Denard, L.; McGlade, J.; Woodward, F. Increases in terrestrial carbon storage from the last glacial maximum to the present. Nature 1990, 348, 711–714. [Google Scholar] [CrossRef]
- World Resources Institute. Taking stock of ecosystems-grassland ecosystems. In World Resources 2000–2001: People and Ecosystems—The Fraying Web of Life; World Resources Institute: Washington, DC, USA, 2000; pp. 119–131. [Google Scholar]
- Ni, J. Carbon storage in grasslands of china. J. Arid Environ. 2002, 50, 205–218. [Google Scholar] [CrossRef]
- Kang, L.; Han, X.; Zhang, Z.; Sun, O.J. Grassland ecosystems in china: Review of current knowledge and research advancement. Philos. Trans. R. Soc. B 2007, 362, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Fang, J.; Yang, Y.; Mohammat, A. Biomass carbon stocks and their changes in Northern China’s grasslands during 1982–2006. Sci. China Life Sci. 2010, 53, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Hu, Z.; Li, S.; Li, X.; Sun, X.; Yu, G. Spatial variations in aboveground net primary productivity along a climate gradient in Eurasian temperate grassland: Effects of mean annual precipitation and its seasonal distribution. Glob. Chang. Biol. 2012, 18, 3624–3631. [Google Scholar] [CrossRef]
- Chen, H.; Zhu, Q.; Peng, C.; Wu, N.; Wang, Y.; Fang, X.; Gao, Y.; Zhu, D.; Yang, G.; Tian, J. The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau. Glob. Chang. Biol. 2013, 19, 2940–2955. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Luo, Y.; Li, X.; Liu, S.; Yu, G.; Zhou, T.; Bahn, M.; Black, A.; Desai, A.R.; Cescatti, A. Redefinition and global estimation of basal ecosystem respiration rate. Glob. Biogeochem. Cycle 2011, 25. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A.; Luo, Y. On the variability of respiration in terrestrial ecosystems: Moving beyond q10. Glob. Chang. Biol. 2006, 12, 154–164. [Google Scholar] [CrossRef]
- Anav, A.; Friedlingstein, P.; Kidston, M.; Bopp, L.; Ciais, P.; Cox, P.; Jones, C.; Jung, M.; Myneni, R.; Zhu, Z. Evaluating the land and ocean components of the global carbon cycle in the cmip5 earth system models. J. Clim. 2013, 26, 6801–6843. [Google Scholar] [CrossRef]
- Olofsson, P.; Lagergren, F.; Lindroth, A.; Lindström, J.; Klemedtsson, L.; Eklundh, L. Towards operational remote sensing of forest carbon balance across Northern Europe. Biogeoscience 2007, 4, 3143–3193. [Google Scholar] [CrossRef]
- Yamaji, T.; Sakai, T.; Endo, T.; Baruah, P.J.; Akiyama, T.; Saigusa, N.; Nakai, Y.; Kitamura, K.; Ishizuka, M.; Yasuoka, Y. Scaling-up technique for net ecosystem productivity of deciduous broadleaved forests in japan using Modis data. Ecol. Res. 2008, 23, 765–775. [Google Scholar] [CrossRef]
- Schubert, P.; Eklundh, L.; Lund, M.; Nilsson, M. Estimating northern peatland CO2 exchange from Modis time series data. Remote Sens. Environ. 2010, 114, 1178–1189. [Google Scholar] [CrossRef]
- Gilmanov, T.G.; Tieszen, L.L.; Wylie, B.K.; Flanagan, L.B.; Frank, A.B.; Haferkamp, M.R.; Meyers, T.P.; Morgan, J.A. Integration of CO2 flux and remotely-sensed data for primary production and ecosystem respiration analyses in the northern great plains: Potential for quantitative spatial extrapolation. Glob. Ecol. Biogeogr. 2005, 14, 271–292. [Google Scholar] [CrossRef]
- Loranty, M.M.; Goetz, S.J.; Rastetter, E.B.; Rocha, A.V.; Shaver, G.R.; Humphreys, E.R.; Lafleur, P.M. Scaling an instantaneous model of tundra nee to the arctic landscape. Ecosystems 2011, 14, 76–93. [Google Scholar] [CrossRef]
- Huang, N.; He, J.-S.; Niu, Z. Estimating the spatial pattern of soil respiration in Tibetan alpine grasslands using landsat tm images and Modis data. Ecol. Indic. 2013, 26, 117–125. [Google Scholar] [CrossRef]
- Rahman, A.; Sims, D.; Cordova, V.; El-Masri, B. Potential of Modis Evi and surface temperature for directly estimating per-pixel ecosystem C fluxes. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Sims, D.A.; Rahman, A.F.; Cordova, V.D.; El-Masri, B.Z.; Baldocchi, D.D.; Bolstad, P.V.; Flanagan, L.B.; Goldstein, A.H.; Hollinger, D.Y.; Misson, L. A new model of gross primary productivity for North American ecosystems based solely on the enhanced vegetation index and land surface temperature from Modis. Remote Sens. Environ. 2008, 112, 1633–1646. [Google Scholar] [CrossRef]
- Wu, C.; Gaumont-Guay, D.; Black, T.A.; Jassal, R.S.; Xu, S.; Chen, J.M.; Gonsamo, A. Soil respiration mapped by exclusively use of Modis data for forest landscapes of Saskatchewan, Canada. ISPRS J. Photogramm. 2014, 94, 80–90. [Google Scholar] [CrossRef]
- Reichstein, M.; Rey, A.; Freibauer, A.; Tenhunen, J.; Valentini, R.; Banza, J.; Casals, P.; Cheng, Y.; Grünzweig, J.M.; Irvine, J. Modeling temporal and large-scale spatial variability of soil respiration from soil water availability, temperature and vegetation productivity indices. Glob. Biogeochem. Cycle 2003, 17. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogee, J.; Allard, V. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Migliavacca, M.; Reichstein, M.; Richardson, A.D.; Colombo, R.; Sutton, M.A.; Lasslop, G.; Tomelleri, E.; Wohlfahrt, G.; Carvalhais, N.; Cescatti, A. Semiempirical modeling of abiotic and biotic factors controlling ecosystem respiration across eddy covariance sites. Glob. Chang. Biol. 2011, 17, 390–409. [Google Scholar] [CrossRef] [Green Version]
- Wohlfahrt, G.; Anderson-Dunn, M.; Bahn, M.; Balzarolo, M.; Berninger, F.; Campbell, C.; Carrara, A.; Cescatti, A.; Christensen, T.; Dore, S. Biotic, abiotic, and management controls on the net ecosystem CO2 exchange of European mountain grassland ecosystems. Ecosystems 2008, 11, 1338–1351. [Google Scholar] [CrossRef]
- Gao, Y.; Yu, G.; Li, S.; Yan, H.; Zhu, X.; Wang, Q.; Shi, P.; Zhao, L.; Li, Y.; Zhang, F. A remote sensing model to estimate ecosystem respiration in northern china and the Tibetan plateau. Ecol. Model. 2015, 304, 34–43. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.L. Environmental Controls on Carbon Budgets in Typical Grassland Ecosystems on Chinese Grassland Transect. Ph.D. Thesis, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing, China, 2006. [Google Scholar]
- Jägermeyr, J.; Gerten, D.; Lucht, W.; Hostert, P.; Migliavacca, M.; Nemani, R. A high-resolution approach to estimating ecosystem respiration at continental scales using operational satellite data. Glob. Chang. Biol. 2014, 20, 1191–1210. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Gu, L.; Niu, Z. Estimating soil respiration using spatial data products: A case study in a deciduous broadleaf forest in the Midwest USA. J. Geophys. Res. Atmos. 2014, 119, 6393–6408. [Google Scholar] [CrossRef]
- Atkin, O.K.; Bloomfield, K.J.; Reich, P.B.; Tjoelker, M.G.; Asner, G.P.; Bonal, D.; Bönisch, G.; Bradford, M.G.; Cernusak, L.A.; Cosio, E.G. Global variability in leaf respiration in relation to climate, plant functional types and leaf traits. New Phytol. 2015, 206, 614–636. [Google Scholar] [CrossRef] [PubMed]
- Doughty, C.E.; Metcalfe, D.; Girardin, C.; Amézquita, F.F.; Cabrera, D.G.; Huasco, W.H.; Silva-Espejo, J.; Araujo-Murakami, A.; Da Costa, M.; Rocha, W. Drought impact on forest carbon dynamics and fluxes in Amazonia. Nature 2015, 519, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, J.; Clark, P.; Cazenave, A.; Gregory, J.; Jevrejeva, S.; Levermann, A.; Merrifield, M.; Milne, G.; Nerem, R.; Nunn, P. Contribution of working group i to the fifth assessment report of the intergovernmental panel on climate change. Clim. Chang. 2013, 1138–1191. [Google Scholar]
- Zhang, J.-W. Vegetation of Xizang (Tibet); Science Press: Beijing, China, 1988. [Google Scholar]
- Bai, Y.; Wu, J.; Xing, Q.; Pan, Q.; Huang, J.; Yang, D.; Han, X. Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 2008, 89, 2140–2153. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.-R.; Wen, X.-F.; Sun, X.-M.; Tanner, B.D.; Lee, X.; Chen, J.-Y. Overview of Chinaflux and evaluation of its eddy covariance measurement. Agric. For. Meteorol. 2006, 137, 125–137. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Li, C.; He, H.; Liu, M.; Su, W.; Fu, Y.; Zhang, L.; Wen, X.; Yu, G. The design and application of CO2 flux data processing system at Chinaflux. Geo-Inf. Sci. 2008, 10, 557–565. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the Modis vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. Timesat—A program for analyzing time-series of satellite sensor data. Comput. Geosci.-UK 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Reichstein, M.; Beer, C. Soil respiration across scales: The importance of a model–data integration framework for data interpretation. J. Plant Nutr. Soil Sci. 2008, 171, 344–354. [Google Scholar] [CrossRef]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Raich, J.W.; Potter, C.S. Global patterns of carbon dioxide emissions from soils. Glob. Biogeochem. Cycle 1995, 9, 23–36. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, G.; Ge, J.; Wang, Q.; Zhu, X.; Xu, Z. Roles of climate, vegetation and soil in regulating the spatial variations in ecosystem carbon dioxide fluxes in the northern hemisphere. PLoS ONE 2015, 10, e0125265. [Google Scholar] [CrossRef] [PubMed]
- Janssens, I.; Lankreijer, H.; Matteucci, G.; Kowalski, A.; Buchmann, N.; Epron, D.; Pilegaard, K.; Kutsch, W.; Longdoz, B.; Grünwald, T. Productivity overshadows temperature in determining soil and ecosystem respiration across European forests. Glob. Chang. Biol. 2001, 7, 269–278. [Google Scholar] [CrossRef]
- Sampson, D.; Janssens, I.; Curiel Yuste, J.; Ceulemans, R. Basal rates of soil respiration are correlated with photosynthesis in a mixed temperate forest. Glob. Chang. Biol. 2007, 13, 2008–2017. [Google Scholar] [CrossRef]
- Epstein, H.E.; Burke, I.C.; Lauenroth, W.K. Regional patterns of decomposition and primary production rates in the US great plains. Ecology 2002, 83, 320–327. [Google Scholar]
- McCulley, R.L.; Burke, I.C.; Nelson, J.A.; Lauenroth, W.K.; Knapp, A.K.; Kelly, E.F. Regional patterns in carbon cycling across the great plains of North America. Ecosystems 2005, 8, 106–121. [Google Scholar] [CrossRef]
- Noormets, A.; Desai, A.; Cook, B.; Euskirchen, E.; Ricciuto, D.; Davis, K.; Bolstad, P.; Schmid, H.; Vogel, C.; Carey, E. Moisture sensitivity of ecosystem respiration: Comparison of 14 forest ecosystems in the upper great lakes region, USA. Agric. For. Meteorol. 2008, 148, 216–230. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Raich, J.W.; Tufekciogul, A. Vegetation and soil respiration: Correlations and controls. Biogeochemistry 2000, 48, 71–90. [Google Scholar] [CrossRef]
- Migliavacca, M.; Reichstein, M.; Richardson, A.D.; Mahecha, M.D.; Cremonese, E.; Delpierre, N.; Galvagno, M.; Law, B.E.; Wohlfahrt, G.; Andrew Black, T. Influence of physiological phenology on the seasonal pattern of ecosystem respiration in deciduous forests. Glob. Chang. Biol. 2015, 21, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.A.; Verchot, L.V.; Cattanio, J.H.; Ackerman, I.L.; Carvalho, J. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry 2000, 48, 53–69. [Google Scholar] [CrossRef]
- Salimon, C.; Davidson, E.; Victoria, R.; Melo, A. CO2 flux from soil in pastures and forests in Southwestern Amazonia. Glob. Chang. Biol. 2004, 10, 833–843. [Google Scholar] [CrossRef]
- Raich, J.W.; Potter, C.S.; Bhagawati, D. Interannual variability in global soil respiration, 1980–94. Glob. Chang. Biol. 2002, 8, 800–812. [Google Scholar] [CrossRef]
- Wang, X.; Ma, M.; Li, X.; Song, Y.; Tan, J.; Huang, G.; Yu, W. Comparison of remote sensing based GPP models at an alpine meadow site. J. Remote Sens. 2012, 16. [Google Scholar]
- Yang, Y.; Fang, J.; Ji, C.; Han, W. Above-and belowground biomass allocation in Tibetan grasslands. J. Veg. Sci. 2009, 20, 177–184. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S. Detection and attribution of vegetation greening trend in china over the last 30 years. Glob. Chang. Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Luyssaert, S.; Ciais, P.; Janssens, I.A.; Chen, A.; Cao, C.; Fang, J.; Friedlingstein, P.; Luo, Y.; Wang, S. Forest annual carbon cost: A global-scale analysis of autotrophic respiration. Ecology 2010, 91, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science & Business Media: Luxemburg, 2011. [Google Scholar]
- Craine, J.; Tilman, D.; Wedin, D.; Reich, P.; Tjoelker, M.; Knops, J. Functional traits, productivity and effects on nitrogen cycling of 33 grassland species. Funct. Ecol. 2002, 16, 563–574. [Google Scholar] [CrossRef]
- Mahecha, M.D.; Reichstein, M.; Carvalhais, N.; Lasslop, G.; Lange, H.; Seneviratne, S.I.; Vargas, R.; Ammann, C.; Arain, M.A.; Cescatti, A. Global convergence in the temperature sensitivity of respiration at ecosystem level. Science 2010, 329, 838–840. [Google Scholar] [CrossRef] [PubMed]
- Hirota, M.; Zhang, P.; Gu, S.; Du, M.; Shimono, A.; Shen, H.; Li, Y.; Tang, Y. Altitudinal variation of ecosystem CO2 fluxes in an alpine grassland from 3600 to 4200 m. J. Plant Ecol. 2009, 2, 197–205. [Google Scholar] [CrossRef]
- Geng, Y.; Wang, Y.; Yang, K.; Wang, S.; Zeng, H.; Baumann, F.; Kuehn, P.; Scholten, T.; He, J.-S. Soil respiration in Tibetan alpine grasslands: Belowground biomass and soil moisture, but not soil temperature, best explain the large-scale patterns. PLoS ONE 2012, 7, e34968. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Shi, P.; Zong, N.; Fu, G.; Shen, Z.; Zhang, X.; Song, M. Climatic patterns modulate ecosystem and soil respiration responses to fertilization in an alpine meadow on the Tibetan Plateau, China. Ecol. Res. 2015, 30, 3–13. [Google Scholar] [CrossRef]
- Zhao, J.; Luo, T.; Li, R.; Li, X.; Tian, L. Grazing effect on growing season ecosystem respiration and its temperature sensitivity in alpine grasslands along a large altitudinal gradient on the central Tibetan Plateau. Agric. For. Meteorol. 2016, 218, 114–121. [Google Scholar] [CrossRef]
- Wagle, P.; Kakani, V.G. Confounding effects of soil moisture on the relationship between ecosystem respiration and soil temperature in switchgrass. Bioenergy Res. 2014, 7, 789–798. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Q.; Han, X.; Wan, S.; Li, L. Temporal and spatial variability and controls of soil respiration in a temperate steppe in Northern China. Glob. Biogeochem. Cycle 2010, 24. [Google Scholar] [CrossRef]
- Dornbush, M.E.; Raich, J.W. Soil temperature, not aboveground plant productivity, best predicts intra-annual variations of soil respiration in central Iowa grasslands. Ecosystems 2006, 9, 909–920. [Google Scholar] [CrossRef]
- Wan, S.; Norby, R.J.; Ledford, J.; Weltzin, J.F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Glob. Chang. Biol. 2007, 13, 2411–2424. [Google Scholar] [CrossRef]
- Davidson, E.; Belk, E.; Boone, R.D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Glob. Chang. Biol. 1998, 4, 217–227. [Google Scholar] [CrossRef]
- HoÈgberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.; Ekblad, A.; HoÈgberg, M.N.; Nyberg, G.; Ottosson-LoÈfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fang, J.; Ma, W.; Guo, D.; Mohammat, A. Large-scale pattern of biomass partitioning across China’s grasslands. Glob. Ecol. Biogeogr. 2010, 19, 268–277. [Google Scholar] [CrossRef]
- Oleson, K.W.; Lawrence, D.M.; Gordon, B.; Flanner, M.G.; Kluzek, E.; Peter, J.; Levis, S.; Swenson, S.C.; Thornton, E.; Feddema, J. Technical Description of Version 4.0 of the Community land Model (CLM); National Center for Atmospheric Research: Boulder, CO, USA, 2010.
- Schimel, D.S.; Participants, V.; Braswell, B. Continental scale variability in ecosystem processes: Models, data, and the role of disturbance. Ecol. Monogr. 1997, 67, 251–271. [Google Scholar] [CrossRef]
- Parton, W.J.; Hartman, M.; Ojima, D.; Schimel, D. Daycent and its land surface Submodel: Description and testing. Glob. Planet. Chang. 1998, 19, 35–48. [Google Scholar] [CrossRef]
- Cramer, W.; Bondeau, A.; Woodward, F.I.; Prentice, I.C.; Betts, R.A.; Brovkin, V.; Cox, P.M.; Fisher, V.; Foley, J.A.; Friend, A.D. Global response of terrestrial ecosystem structure and function to CO2 and climate change: Results from six dynamic global vegetation models. Glob. Chang. Biol. 2001, 7, 357–373. [Google Scholar] [CrossRef]
- Wang, T.; Ciais, P.; Piao, S.; Ottle, C.; Brender, P.; Maignan, F.; Arain, A.; Gianelle, D.; Gu, L.; Lafleur, P. Controls on winter ecosystem respiration at mid-and high-latitudes. Biogeoscience 2010, 7. [Google Scholar] [CrossRef]
- Mielnick, P.C.; Dugas, W.A. Soil CO2 flux in a tallgrass prairie. Soil Biol. Biochem. 2000, 32, 221–228. [Google Scholar] [CrossRef]
- Liu, X.; Wan, S.; Su, B.; Hui, D.; Luo, Y. Response of soil CO2 efflux to water manipulation in a tallgrass prairie ecosystem. Plant Soil 2002, 240, 213–223. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Campioli, M. Nutrient availability as the key regulator of global forest carbon balance. Nat. Clim. Chang. 2014, 4, 471–476. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore, B. Satellite-based modeling of gross primary production in an evergreen Needleleaf forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Wu, C.; Niu, Z.; Gao, S. Gross primary production estimation from Modis data with vegetation index and photosynthetically active radiation in maize. J. Geophys. Res. Atmos. 2010, 115. [Google Scholar] [CrossRef]
- Yuste, J.C.; Janssens, I.; Carrara, A.; Meiresonne, L.; Ceulemans, R. Interactive effects of temperature and precipitation on soil respiration in a temperate maritime pine forest. Tree Physiol. 2003, 23, 1263–1270. [Google Scholar] [CrossRef]
- Amos, B.; Arkebauer, T.J.; Doran, J.W. Soil surface fluxes of greenhouse gases in an irrigated maize-based agroecosystem. Soil Sci. Soc. Am. J. 2005, 69, 387–395. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grassland Type | Site Name | Location | Elevation (m) | Canopy Height (m) | Tower Height (m) | Operation Period |
|---|---|---|---|---|---|---|
| Alpine shrub meadow | HBSH | 37.67°N 101.33°E | 3293 | 0.6–0.7 | 2.2 | 2003–2008, 2010–2012 |
| Alpine Kobresia meadow | HBKO | 37.61°N 101.31°E | 3148 | 0.2–0.3 | 2.2 | 2003–2005, 2009, 2012 |
| GL | 34.35°N 100.56°E | 3980 | 0.2–0.3 | 2.2 | 2007, 2010–2011, 2013 | |
| AR | 38.04°N 100.46°E | 3033 | 0.2–0.3 | 3.15 | 2008–2010 | |
| MQKO | 33.88°N 102.15°E | 3533 | 0.2–0.3 | 3.15 | 2009–2010 | |
| Alpine swamp meadow | HBSW | 37.61°N 101.33°E | 3160 | 0.2–0.5 | 2.2 | 2004–2008, 2010–2012 |
| DXSW | 30.47°N 91.06°E | 4286 | 0.2–0.5 | 2.1 | 2009–2010 | |
| MQSW | 33.76°N 101.68°E | 3503 | 0.3–0.5 | 3.2 | 2010 | |
| Alpine meadow steppe | DXST | 30.50°N 91.06°E | 4333 | <0.2 | 2.2 | 2004–2010 |
| ZF | 28.36°N 86.95°E | 4293 | <0.2 | 3.1 | 2009 | |
| NMC | 30.77°N 90.96°E | 4730 | <0.2 | 3.1 | 2009 | |
| Typical steppe | NMG | 43.53°N 116.67°E | 1200 | 0.2–0.3 | 4 | 2004–2008, 2010, 2012 |
| DL | 42.05°N 116.28°E | 1324 | 0.3–0.5 | 5 | 2010–2011 | |
| XLHT | 44.13°N 116.32°E | 1187 | 0.1–0.3 | 5 | 2010–2011 | |
| Desert steppe | SZWQ | 41.8°N 111.9°E | 1438 | 0.1–0.2 | 3 | 2011–2012 |
| Meadow steppe | HLBE | 49.06°N 119.4°E | 628 | 0.3–0.5 | 3 | 2012 |
| Grassland Type | Alpine Grasslands | Temperate Grasslands | |||
|---|---|---|---|---|---|
| Reref | p1 | −2.646 ± 0.456 | −0.019 ± 0.023 | ||
| p2 | 15.100 ± 2.062 | 8.347 ± 1.068 | |||
| p3 | 0.173 ± 0.024 | −0.029 ± 0.009 | |||
| p4 | 18.828 ± 0.939 | 3.952 ± 0.322 | |||
| f(T,P,W) | p5 | 1.787 ± 0.081 | (1.421 ± 0.136) * | 0.658 ± 0.119 | (0.941 ± 0.147) * |
| E0 | 279.764 ± 53.87 | (164.507 ± 31.874) * | 166.327 ± 12.402 | (154.305 ± 18.729) * | |
| p6 | 0.250 ± 0.024 | (0.193 ± 0.046) * | 0.944 ± 0.039 | (0.085 ± 0.061) * | |
| p7 | 0.094 ± 0.039 | (0.108 ± 0.046) * | −0.363 ± 0.081 | (0.190 ± 0.050) * | |
| k | 0.021 ± 0.064 | 0.207 ± 0.0.082 | |||
| Vegetation Type | Types of Parameter Estimation | Types of Cross-Validation | R2 | RMSE | p Value |
|---|---|---|---|---|---|
| Alpine Grasslands | SH_SW_ST | KO | 0.844 | 0.841 | <0.01 |
| KO_SW_ST | SH | 0.756 | 1.047 | <0.01 | |
| KO_SH_ST | SW | 0.917 | 0.927 | <0.01 | |
| KO_SH_SW | ST | 0.784 | 1.544 | <0.01 | |
| Joint-sites (AR_GL_HBKO_GL) | / | 0.778 | 0.774 | <0.01 | |
| Temperate Grasslands | TS_DS | MS | 0.803 | 0.405 | <0.01 |
| MS_DS | TS | 0.575 | 0.919 | <0.01 | |
| TS_MS | DS | 0.643 | 1.021 | <0.01 | |
| Joint-sites (TS_MS_DS) | / | 0.749 | 0.605 | <0.01 |
| Grassland Type | EVImean | EVImean & LSWImean | EVImean & LSWImean & LSTmean | |||
|---|---|---|---|---|---|---|
| R2 | RMSE (g C m−2 day−1) | R2 | RMSE (g C m−2 day−1) | R2 | RMSE (g C m−2 day−1) | |
| Temperate grasslands | 0.424 | 0.147 | 0.541 | 0.125 | 0.554 | 0.113 |
| Alpine grasslands | 0.539 | 0.714 | 0.577 | 0.653 | 0.588 | 0.617 |
| Vegetation Type | Model without fw | Model with fw | ||
|---|---|---|---|---|
| R2 | RMSE (g C m−2 day−1) | R2 | RMSE (g C m−2 day−1) | |
| Typical steppe | 0.68 | 0.70 | 0.74 | 0.61 |
| Desert steppe | 0.78 | 0.22 | 0.97 | 0.13 |
| Meadow steppe | 0.94 | 0.35 | 0.95 | 0.34 |
| Alpine Kobresia meadow | 0.75 | 1.09 | 0.78 | 0.93 |
| Alpine meadow steppe | 0.71 | 0.21 | 0.81 | 0.16 |
| Alpine swamp meadow | 0.69 | 0.77 | 0.72 | 0.74 |
| Alpine shrub meadow | 0.86 | 0.74 | 0.88 | 0.72 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, R.; He, H.; Ren, X.; Zhang, L.; Li, P.; Zeng, N.; Yu, G.; Zhang, L.; Yu, S.-Y.; Zhang, F.; et al. A Satellite-Based Model for Simulating Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands. Remote Sens. 2018, 10, 149. https://doi.org/10.3390/rs10010149
Ge R, He H, Ren X, Zhang L, Li P, Zeng N, Yu G, Zhang L, Yu S-Y, Zhang F, et al. A Satellite-Based Model for Simulating Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands. Remote Sensing. 2018; 10(1):149. https://doi.org/10.3390/rs10010149
Chicago/Turabian StyleGe, Rong, Honglin He, Xiaoli Ren, Li Zhang, Pan Li, Na Zeng, Guirui Yu, Liyun Zhang, Shi-Yong Yu, Fawei Zhang, and et al. 2018. "A Satellite-Based Model for Simulating Ecosystem Respiration in the Tibetan and Inner Mongolian Grasslands" Remote Sensing 10, no. 1: 149. https://doi.org/10.3390/rs10010149