Remote Sensing and Modeling of Mosquito Abundance and Habitats in Coastal Virginia, USA

Abstract
:1. Introduction
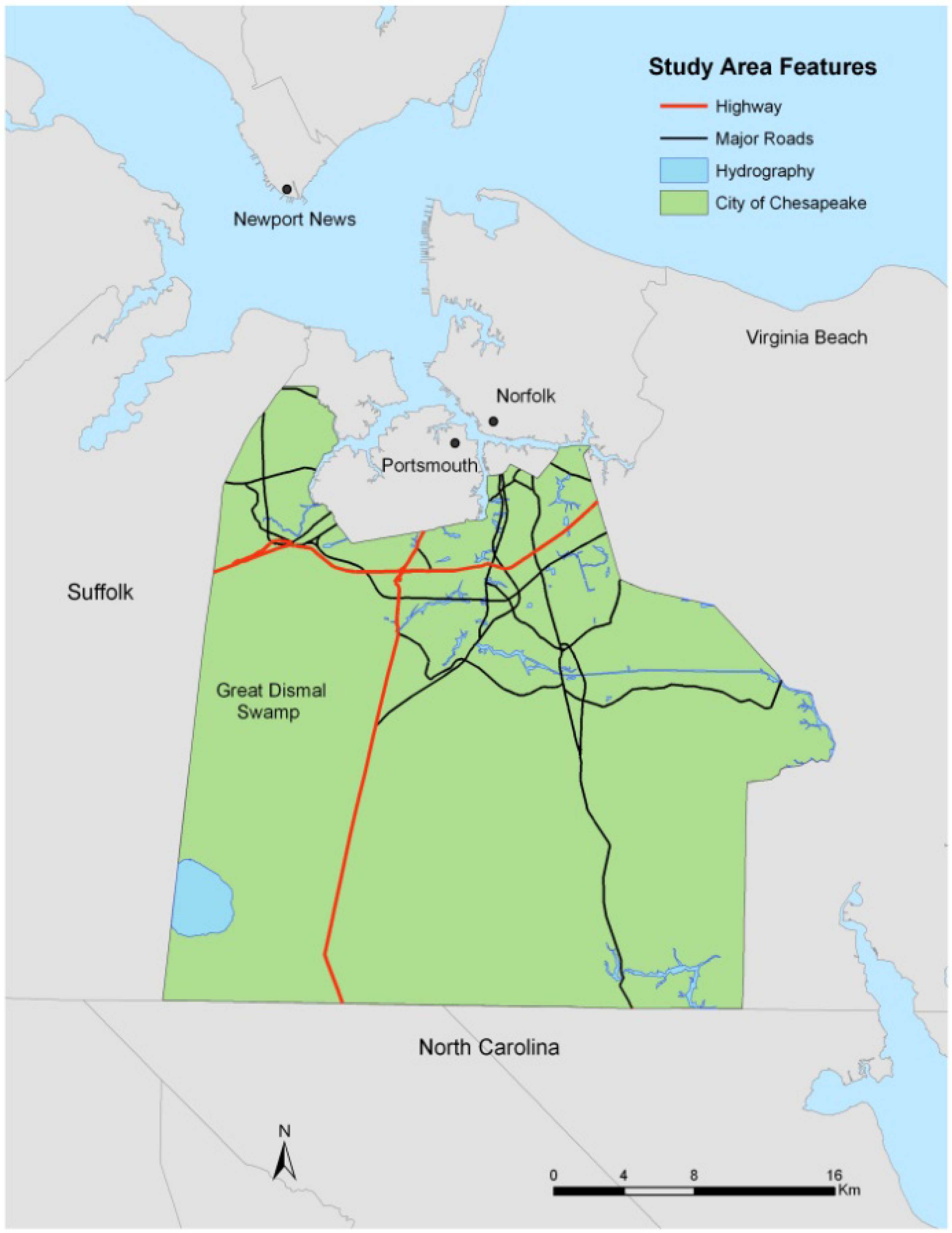
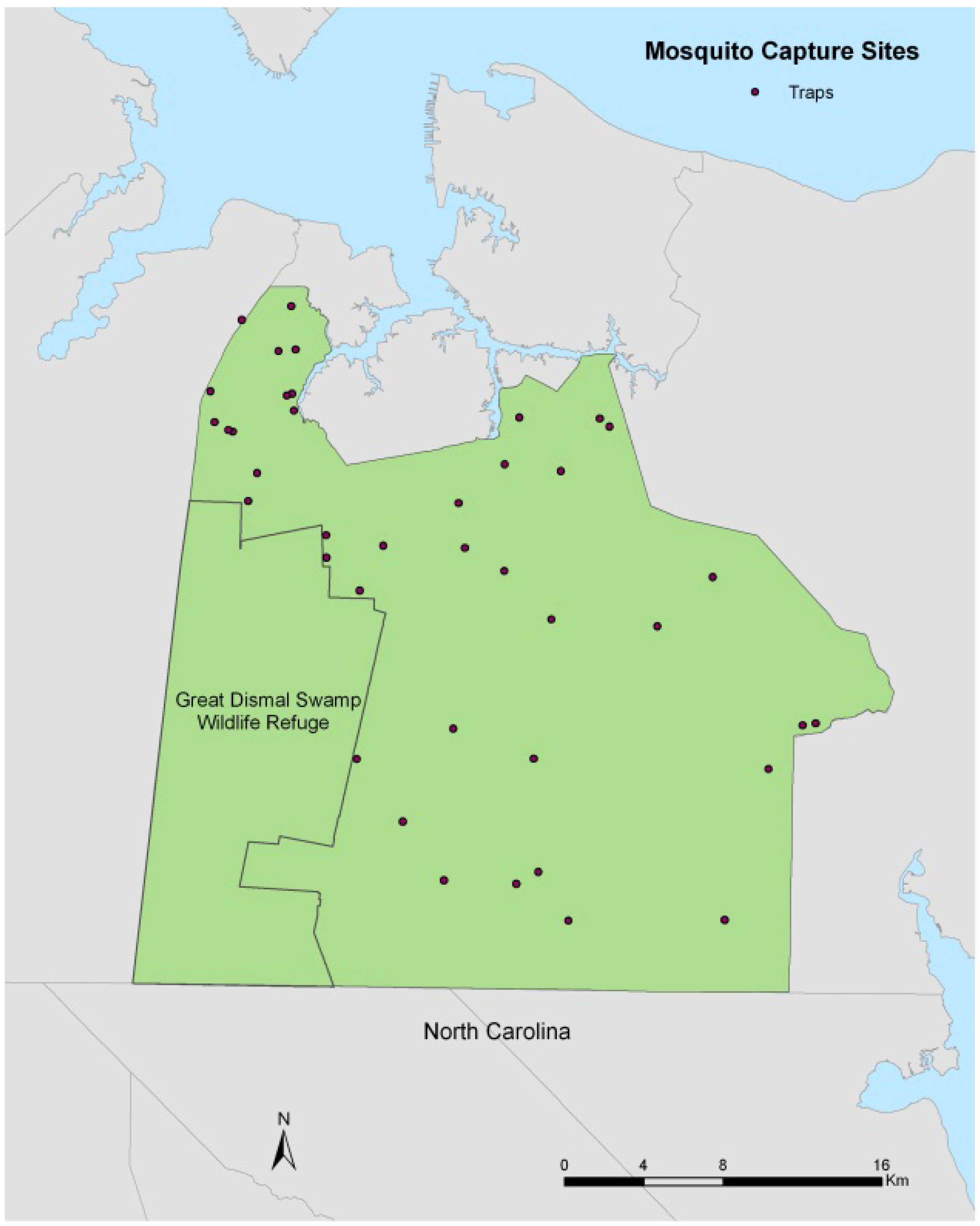
2. Study Area and Data


3. Methodology
3.1. Habitat Suitability Index (HSI)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Code | Data Type | Source | Description |
|---|---|---|---|---|
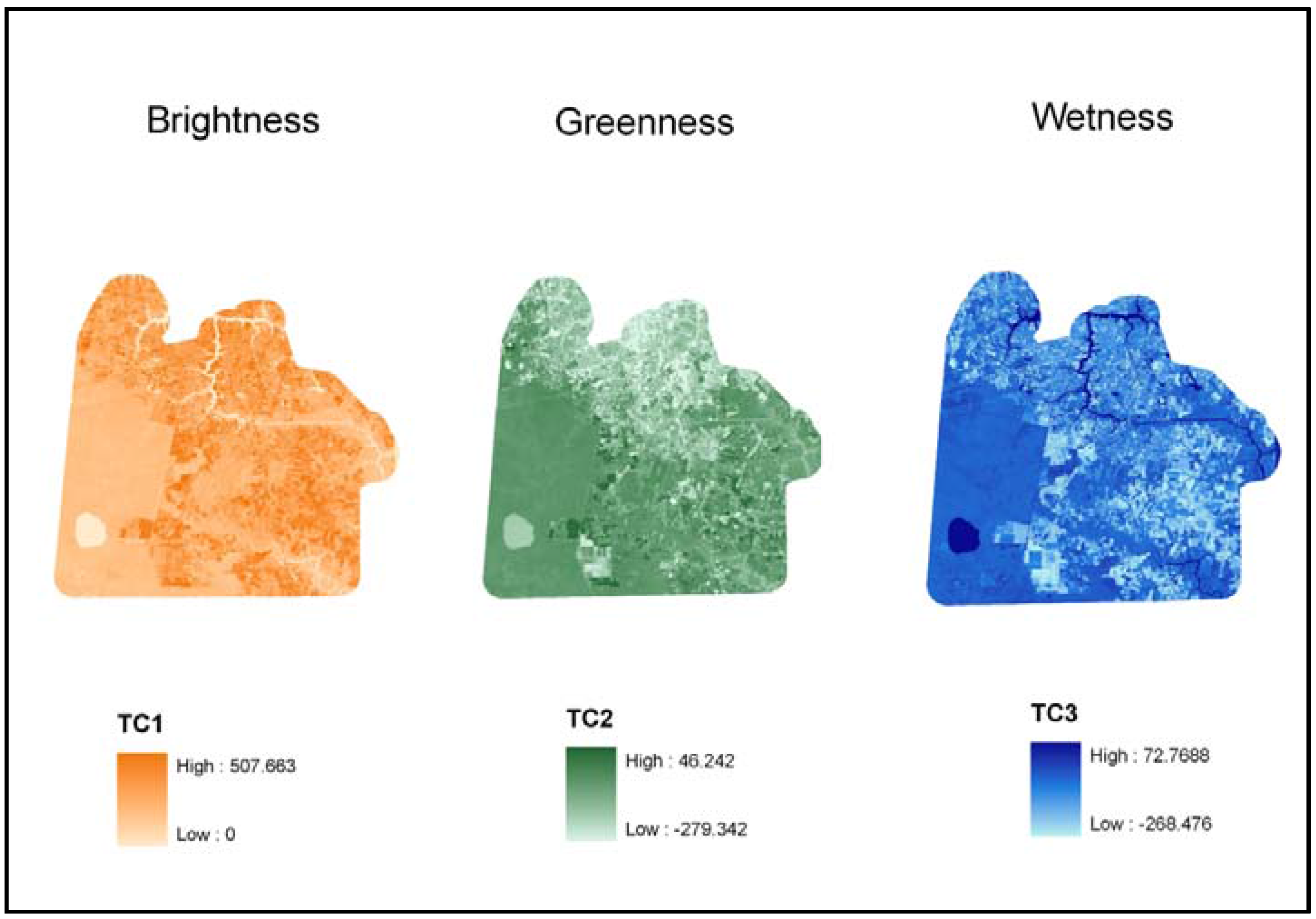
| Landsat Tasseled-cap indices, 2 July 2002 | TC1-TC3 | Raster: Landsat-7 ETM+ | USGS | TC1 (Brightness) TC2 (Greenness) TC3 (Wetness) |
| Hydrologic | HYD | Vector (polygon) | NRCS | Presence of water |
| Percent Hydric Composition | HYDRIC | Vector (polygon) | NRCS | Soil meets requirements for hydric soil |
| Drain Potential | DRAIN | Vector (polygon) | NRCS | Degree of hydraulic conductivity and low water-holding capacity |
| Runoff Potential | RUNOF | Vector (polygon) | NRCS | Degree of potential water loss by overland flow |
| Water Table Depth | WTD | Vector (polygon) | NRCS | Minimum value for the range in depth to the seasonally high water table (April-June) |
| Available Water Storage (25 cm) | AWS25 | Vector (polygon) | NRCS | Maximum value for the range of available water in plant root zones |

3.2. Mosquito Abundance Model
4. Results and Discussion
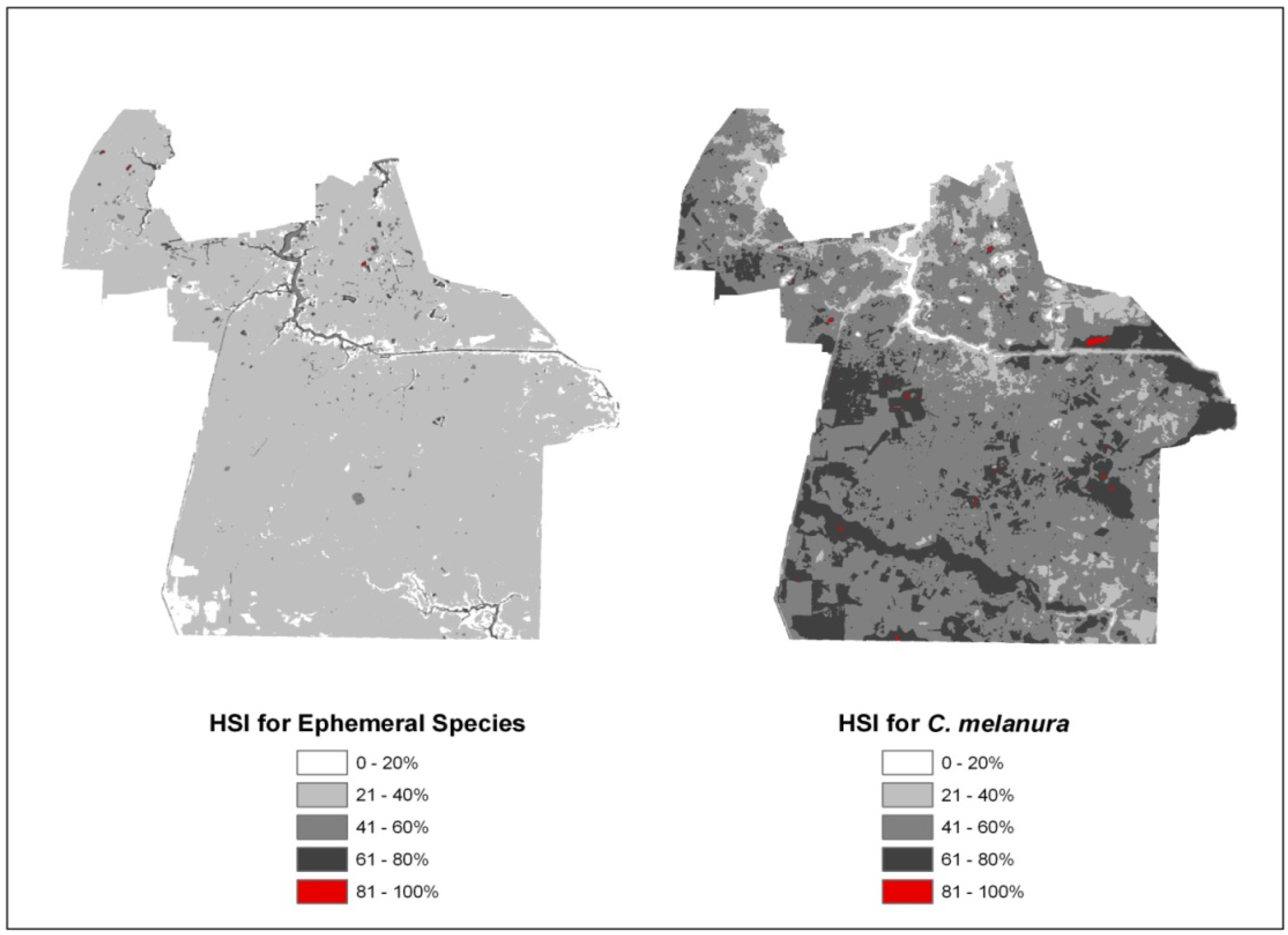
4.1. Habitat Suitability Index (HSI)

| Species | R2 | Adjusted R2 | F | Sig. |
|---|---|---|---|---|
| Ephemeral | 0.356 | 0.238 | 3.035 | 0.018 |
| C. melanura | 0.339 | 0.236 | 3.287 | 0.017 |
| Species | Variable | B | t | Sig. |
|---|---|---|---|---|
| Ephemeral | Constant | −111.719 | −1.629 | 0.113 |
| Ephemeral | TC1 | 1.065 | 3.106 | 0.004 |
| Ephemeral | TC2 | 0.517 | 1.805 | 0.080 |
| Ephemeral | HYD | 20.807 | 0.762 | 0.452 |
| Ephemeral | DRAIN | −7.925 | −1.212 | 0.234 |
| Ephemeral | RUNOFF | 20.730 | 0.904 | 0.373 |
| Ephemeral | WTD | −0.283 | −1.266 | 0.215 |
| C. melanura | Constant | −532.162 | −2.818 | 0.008 |
| C. melanura | TC2 | −5.357 | −2.599 | 0.014 |
| C. melanura | TC3 | 0.193 | 0.059 | 0.953 |
| C. melanura | DRAIN | 11.926 | 0.531 | 0.599 |
| C. melanura | RUNOFF | 510.400 | 3.096 | 0.004 |
| C. melanura | AWS25 | 51.574 | 1.924 | 0.063 |
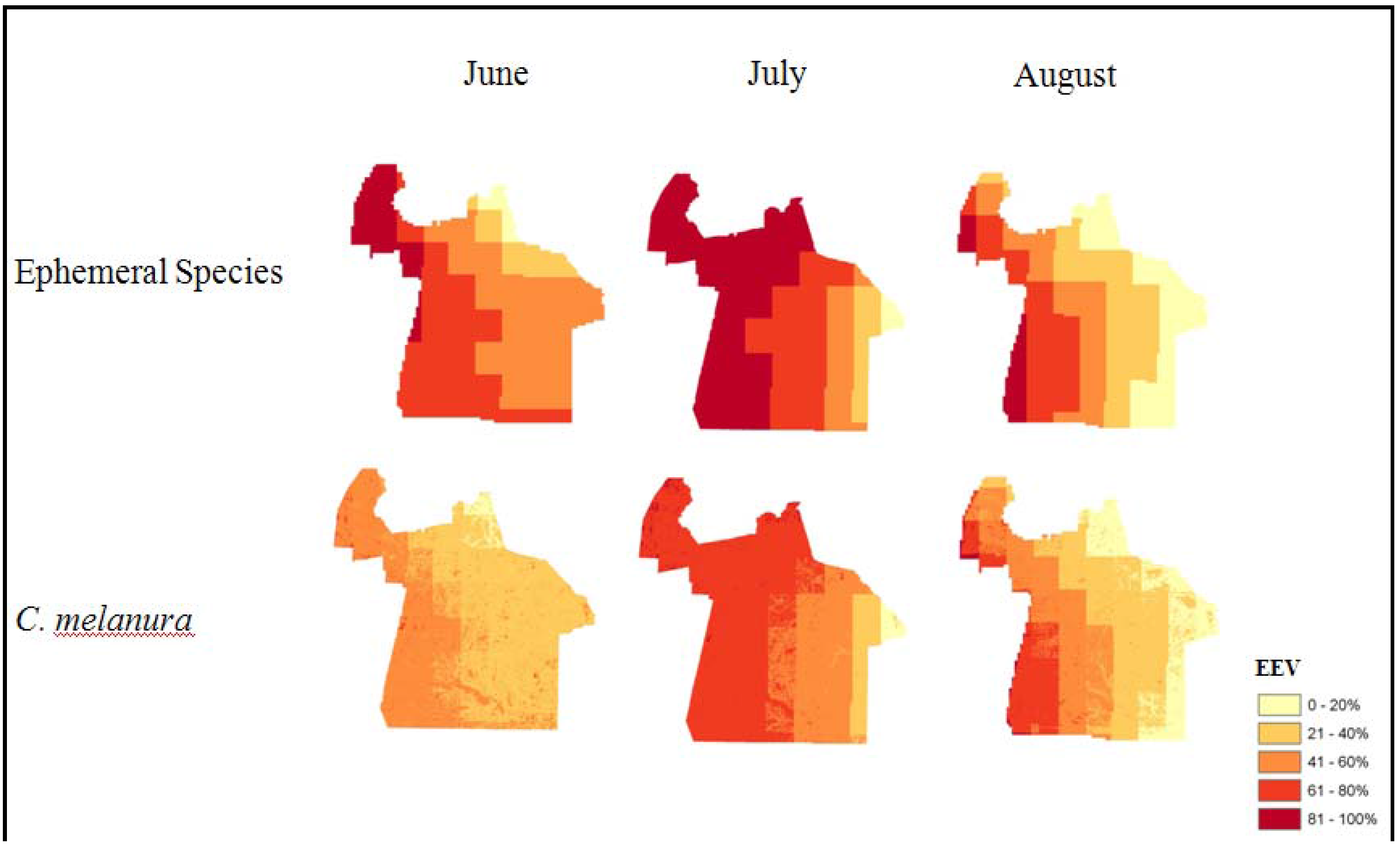
4.2. Effects of the Environmental Variables (EEV)
| Species | R2 | Adjusted R2 | F | Significance |
|---|---|---|---|---|
| Ephemeral | 0.270 | 0.235 | 7.846 | 0.000 |
| C. melanura | 0.405 | 0.377 | 14.793 | 0.000 |
| Species | Variable | B | t | Significance |
|---|---|---|---|---|
| Ephemeral | Constant | −104.888 | −2.875 | 0.005 |
| Ephemeral | Precipitation | 1.193 | 2.989 | 0.004 |
| Ephemeral | Temperature | 1.462 | 2.987 | 0.004 |
| Ephemeral | Precip_Temp | −0.015 | −3.108 | 0.003 |
| Ephemeral | TMI | 0.000 | −0.006 | 0.995 |
| C. melanura | Constant | −138.191 | −3.298 | 0.001 |
| C. melanura | Precipitation | 1.444 | 3.295 | 0.001 |
| C. melanura | Temperature | 1.881 | 3.508 | 0.001 |
| C. melanura | Precip_Temp | −0.020 | −3.498 | 0.001 |
| C. melanura | TMI | 0.016 | 1.224 | 0.224 |

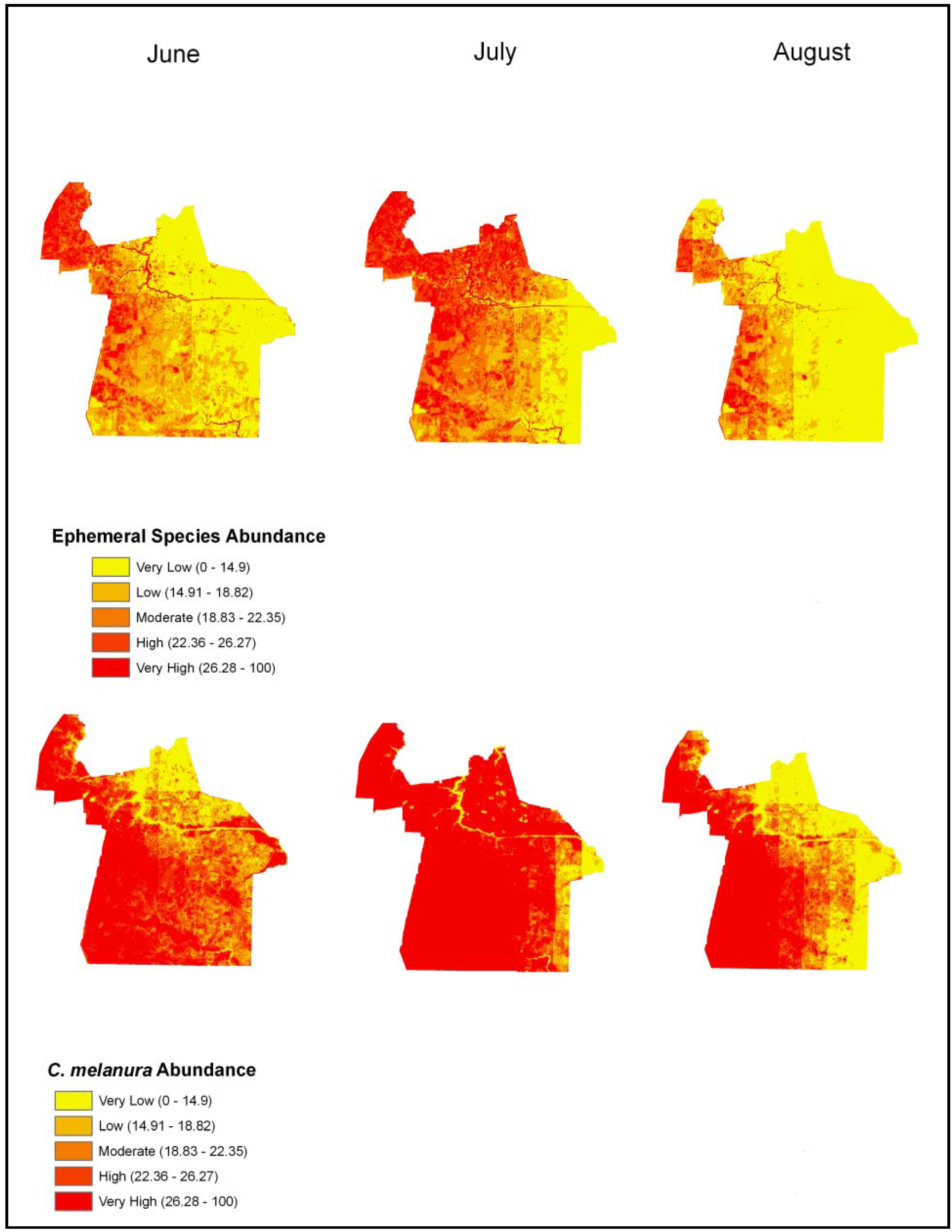
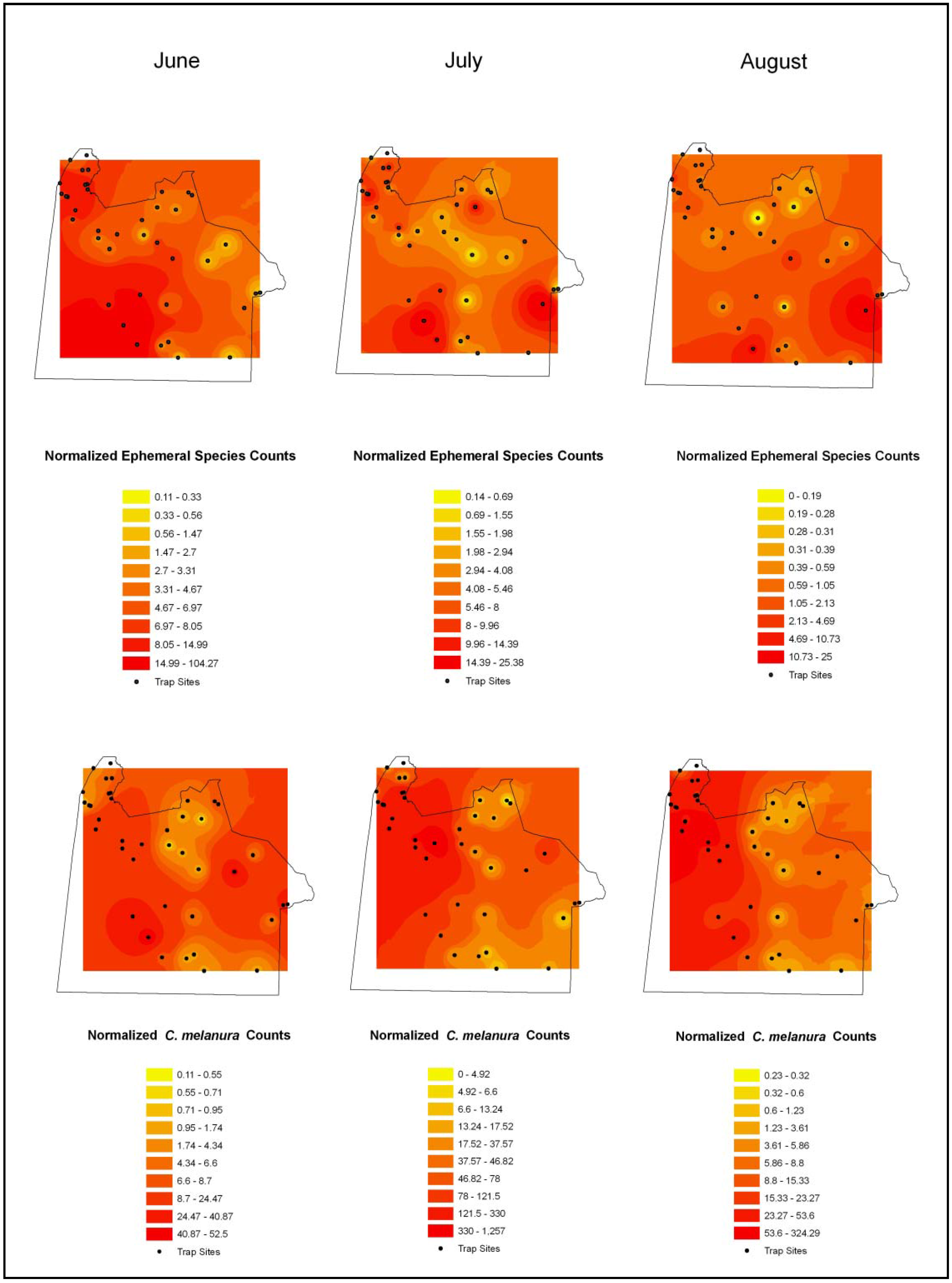
4.3. Mosquito Abundance


5. Conclusions
Acknowledgments
References
- Toole, M.J. Communicable diseases and disease control. In The Public Health Consequences of Disasters; Noji, E.K., Ed.; Oxford University Press: New York, NY, USA, 1997; p. 81. [Google Scholar]
- Waring, S.; Zakos-Feliberti, A.; Wood, R.; Stone, M.; Padgett, P.; Arafat, R. The utility of geographic information systems (GIS) in rapid epidemiological assessments following weather-related disasters: Methodological issues based on the Tropical Storm Allison experience. Int. J. Hyg. Environ. Health 2005, 208, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, F.; Crans, W.J. Effect of temperature on the development of Culiseta melanura (Diptera:Culicidae) and its impact on amplification of eastern equine encephalitis virus in birds. J. Med. Entomol. 1998, 35, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Crans, W.G. A classification system for mosquito life cycle: Life cycle types for mosquitoes of the northeastern United States. J. Vector Ecol. 2004, 29, 1–10. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention (CDC). Eastern Equine Encephalitis; 2009. Available online: http://www.cdc.gov/EasternEquineEncephalitis/Epi.html (accessed on 29 October 2009).
- Centers for Disease Control and Prevention (CDC). West Nile Virus: What You Need To Know; 2006. Available online: http://www.cdc.gov/ncidod/dvbid/westnile/wnv_factsheet.htm (accessed on 2 February 2010).
- Kitron, U. Risk maps: Transmission and burden of vector-borne diseases. Parasitol. Today 2000, 16, 324–325. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Glass, G.E.; Keesing, F. Spatial epidemiology: An emerging (or re-emerging) discipline. Trend. Ecol. Evol. 2005, 20, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Bellows, A.S. Modeling Habitat and Environmental Factors Affecting Mosquito Abundance in Chesapeake, Virginia. Ph.D. Dissertation, Old Dominion University, Norfolk, VA, USA, 2007. [Google Scholar]
- Hutchinson, G.E. Population studies: Animal ecology and demography. In Cold Spring Harbor Symposia on Quantitative Biology; 1957; Volume 22, pp. 415–427. [Google Scholar]
- Hardesty, D.L. The niche concept: Suggestions for its use in human ecology. Human Ecol. 1975, 3, 71–85. [Google Scholar] [CrossRef]
- Crist, E.P.; Cicone, R.C. Application of the Tasseled Cap concept to simulated Thematic Mapper data. Photogramm. Engin. Remote Sensing 1984, 50, 343–352. [Google Scholar]
- Guerra, M.; Walker, E.; Jones, C.; Paskewitz, S.; Cortinas, M.R.; Stancil, A.; Beck, L.; Bobo, M.; Kitron, U. Predicting the risk of Lyme disease: Habitat suitability for Ixodes scapularis in the north central United States. Emerg. Infect. Disease 2002, 8, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Crist, E.P.; Laurin, R.; Ciccone, R.C. Vegetation and Soils Information Contained in Transformed Thematic Mapper Data. In Proceedings of IGARSS ’86 Symposium, Zurich, Switzerland, 8–11 September 1986; pp. 1465–1470.
- Crist, E.P.; Cicone, R.C. A physically-based transformation of Thematic Mapper Data—The TM Tasseled Cap. GeoSci. Remote Sens. 1984, 3, 256–263. [Google Scholar] [CrossRef]
- Tanser, F.C.; Sharp, B.; Le Sueur, D. Potential effect of climate change on malaria in Africa. Lancet 2003, 362, 1792–1798. [Google Scholar] [CrossRef]
- Turner, M.G. Landscape ecology: The effect of pattern on process. Ann. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar] [CrossRef]
- Beven, K.J. Topmodel: A critique. Hydrol. Process. 1997, 11, 1069–1085. [Google Scholar] [CrossRef]
- PRISM Climate Group. PRISM Data. 2011. Available online http://prism.oregonstate.edu (accessed on 1 August 2011).
- Smith, C.M.; Graffeo, C.S. Regional impact of Hurricane Isabel on emergency departments in coastal southeastern Virginia. Vet. Res. Commun. 2005, 12, 1201–1205. [Google Scholar] [CrossRef]
- Pettie, S.T. A classification system for mosquito life cycle: Life cycle types for mosquitoes of the northeastern United States. J. Forest Hist. 1976, 20, 28–33. [Google Scholar]
- Ward, M.P.; Wittich, C.A.; Fosgate, G.; Srinivasan, R. Environmental risk factors for equine West Nile Virus disease cases in Texas. Acad. Emerg. Med. 2009, 33, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Gage, K.L.; Burkot, T.R.; Eisen, R.R.; Hayes, E.B. Climate and vector-borne diseases. Am. J. Prevent. Med. 2008, 35, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Service, M.W. Effects of wind on the behaviour and distribution of mosquitoes and blackflies. Int. J. Biometeorol. 1980, 24, 347–353. [Google Scholar] [CrossRef]
- Hartfield, K.A.; Landau, K.I.; van Leeuwen, W.J.D. Fusion of high resolution aerial Multispectral and LiDAR data: Land cover in the context of urban mosquito habitat. Remote Sens. 2011, 3, 2364–2383. [Google Scholar] [CrossRef]
- Maxwell, S.K. Downscaling pesticide use data to the crop field level in California using Landsat satellite imagery: Paraquat case study. Remote Sens. 2011, 3, 1805–1816. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cleckner, H.L.; Allen, T.R.; Bellows, A.S. Remote Sensing and Modeling of Mosquito Abundance and Habitats in Coastal Virginia, USA. Remote Sens. 2011, 3, 2663-2681. https://doi.org/10.3390/rs3122663
Cleckner HL, Allen TR, Bellows AS. Remote Sensing and Modeling of Mosquito Abundance and Habitats in Coastal Virginia, USA. Remote Sensing. 2011; 3(12):2663-2681. https://doi.org/10.3390/rs3122663
Chicago/Turabian StyleCleckner, Haley L., Thomas R. Allen, and A. Scott Bellows. 2011. "Remote Sensing and Modeling of Mosquito Abundance and Habitats in Coastal Virginia, USA" Remote Sensing 3, no. 12: 2663-2681. https://doi.org/10.3390/rs3122663