6.1. Biomass Models
Spatially explicit and timely biomass estimates for forests are of prime importance for biogeochemical models of forest productivity, but are also vital for the newly-developing carbon markets. Field-based estimates from inventories therefore need to be complemented by remote sensing-based measures in order to fulfill the spatio-temporal needs of many users from scientific and applied domains alike.
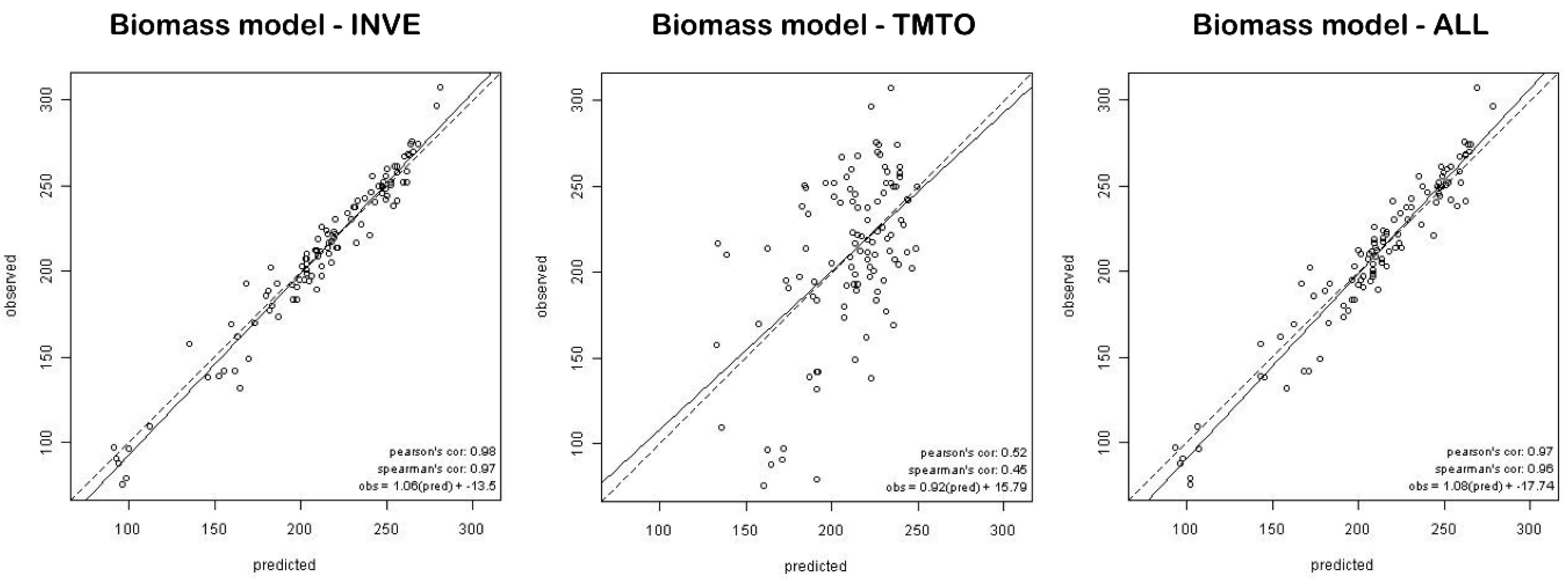
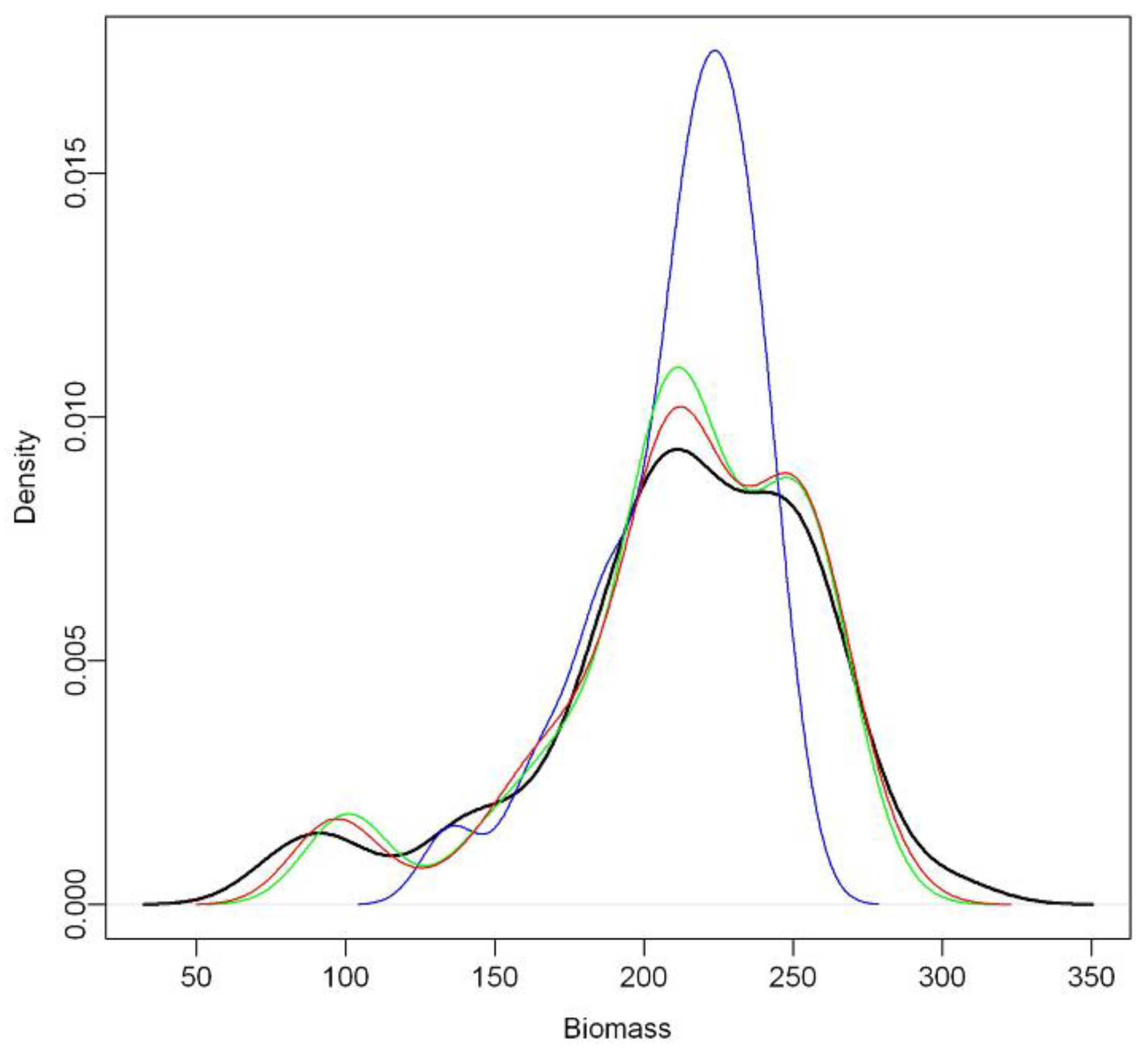
Our results showed a very strong relationship between inventory-based predictors and estimated biomass (
Table 1). The INVE model was the most predictive for biomass, and captured the complete observed biomass range from approximately 100 to 300 Mg ha
−1 (
Figure 3). These values are consistent with the typical range for boreal and temperate coniferous forests [
15,
52,
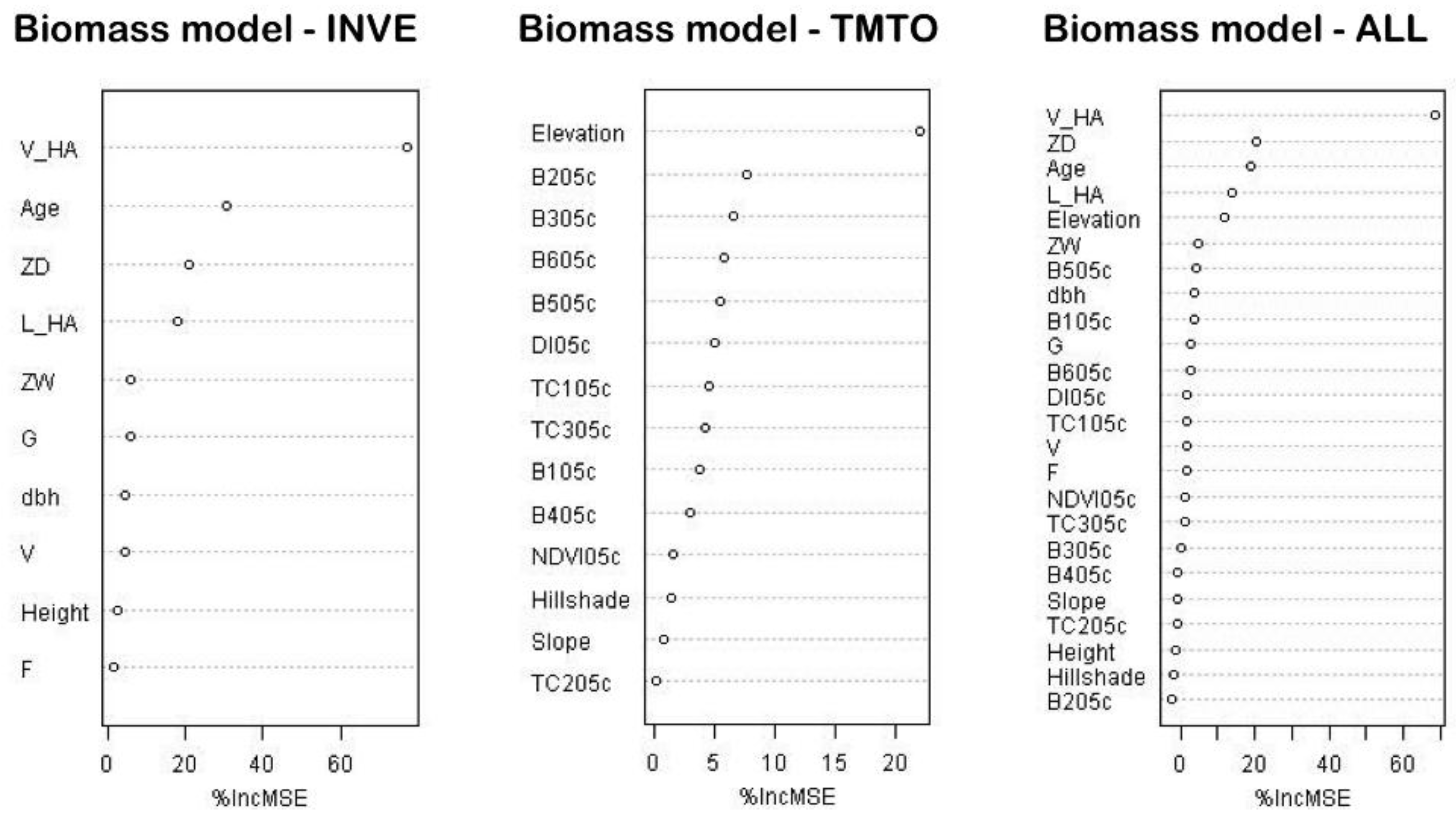
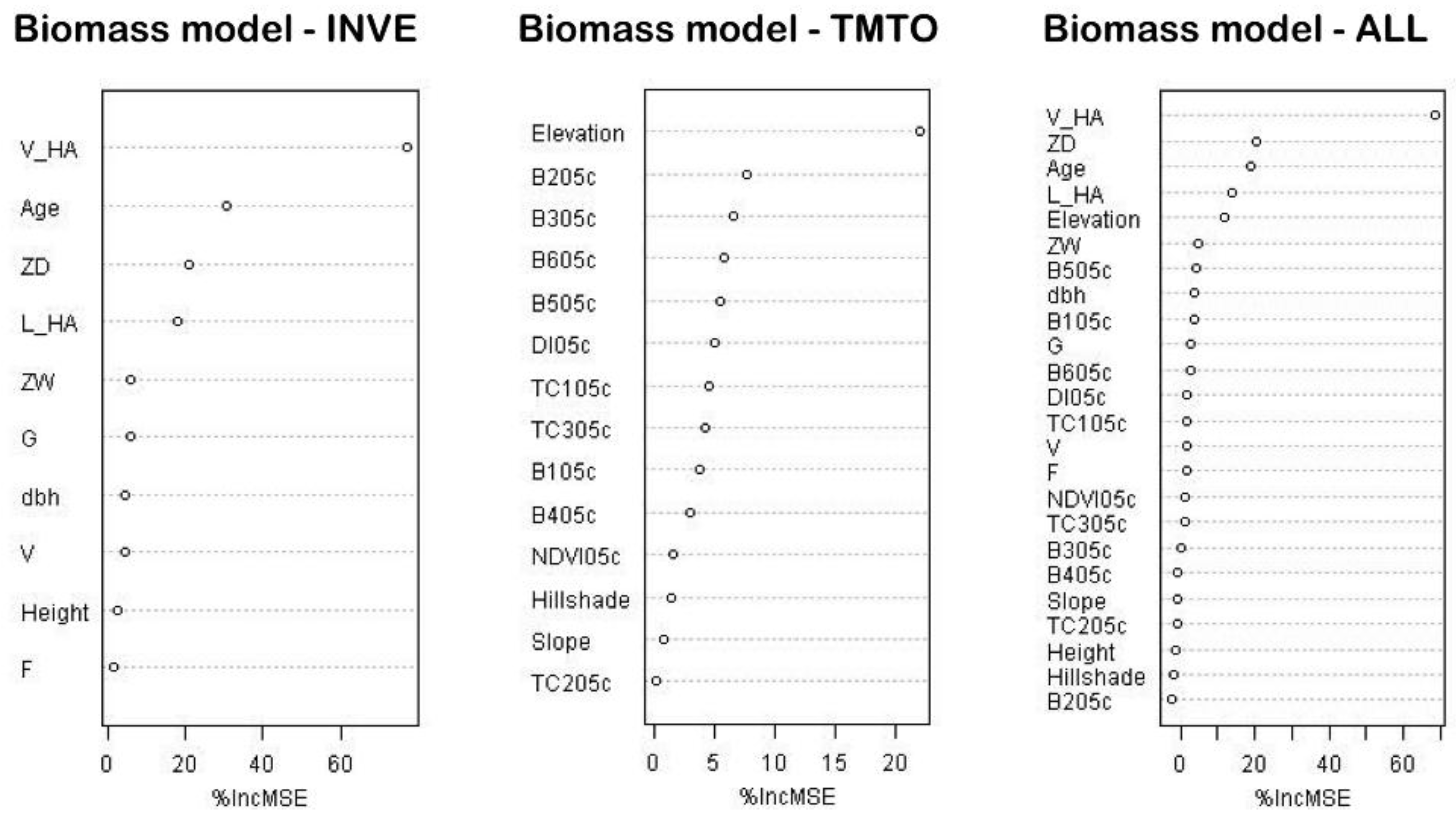
53]. Using our modeling techniques for areas with up-to-date SBI, we found stand volume, relative density, and dominant tree age (
Table 3) to be the strongest predictors for biomass estimation. This is consistent with allometric equations that often also incorporate these variables [
15,
17]. Generally, the predictive power of inventory-derived stand structure metrics is very high, especially for stand volume (
Table 3).
Biomass estimates for areas lacking forest inventory data (TMTO) led to considerably less precise results. None of the pixel-based variables, aside from elevation, significantly contributed to biomass prediction. Powell
et al. [
36] also reported similar results based on Landsat data and in areas with strong elevational gradients. In our case, elevation often correlates with the forest dieback experienced during the past decade, which may explain its relationship to biomass distribution [
40].
Tests on individual field-derived predictors which have improved TMTO-based biomass estimates (
Table 4 and
Table 5) provide promising results. We found that stand volume or even tree height—parameters easily taken in the field—significantly increased the correlation between observed and predicted biomass. Moreover, as tree height is easily measured in the field or can be measured from airborne Lidar data, we strongly recommend its inclusion to improve prediction results. To summarize, as might be expected, models based on SBI predicted biomass are better than models based on satellite data. However, by integrating TM data and stand volume or tree height information, our biomass estimate accuracy increased significantly. Our method therefore provides increased spatial detail within inventory polygons and fills the “knowledge gaps” in areas lacking SBI.
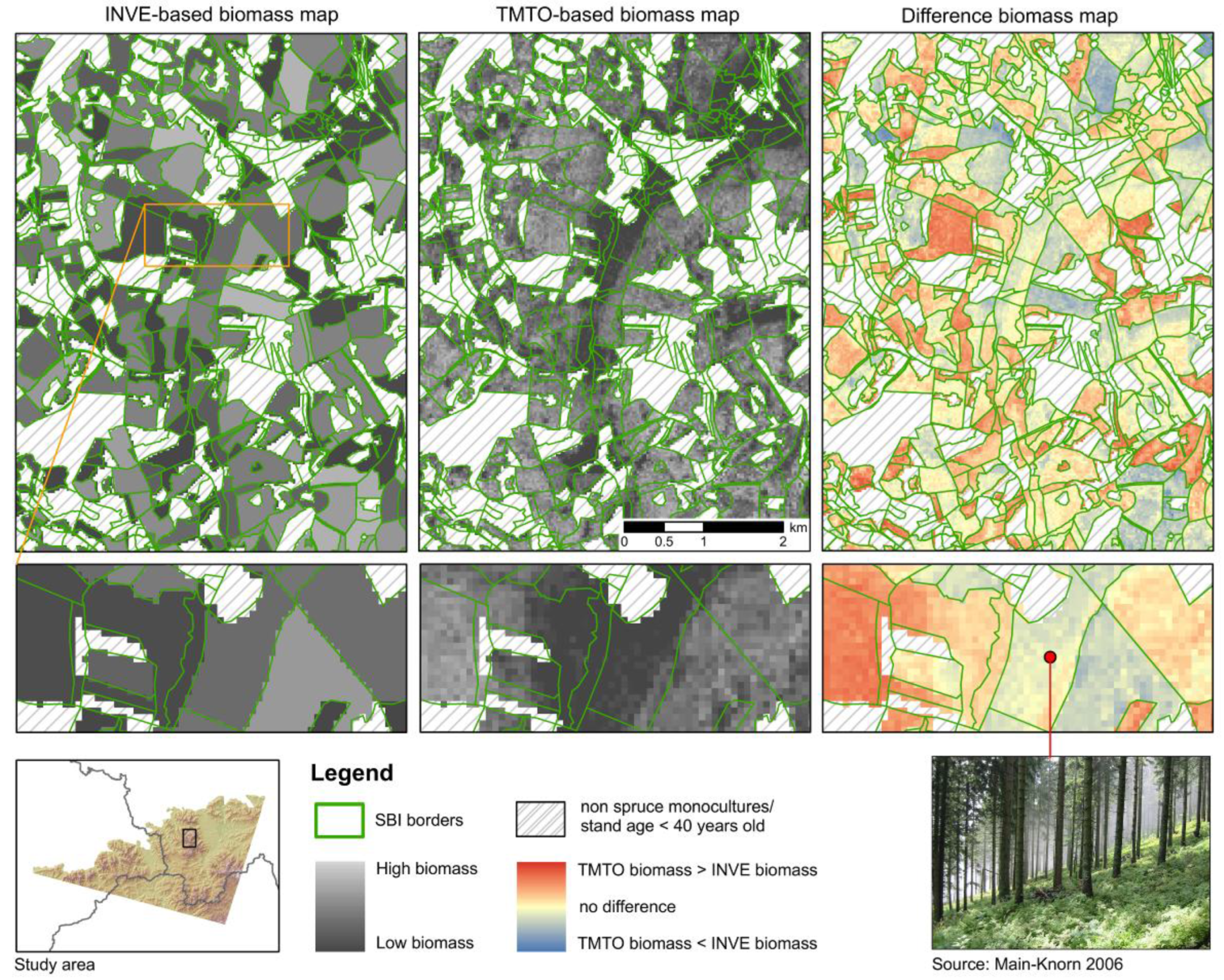
Beyond absolute model quality, we focused on the usefulness of remote sensing-based results in dealing with spatial variability within forest inventory stands. Our difference map highlights the potential of spectral data compared to polygon-based inventory layers that average stand-wise heterogeneity (
Figure 6). Even in homogenous areas such as those selected for this study, it is obvious that remote sensing-based mapping results support decision-making at a sub-stand level and reveal spatial detail that would otherwise be averaged out. In addition, remote sensing-based estimates of forest stand parameters is more time-efficient; it rather depends on the research or management question at hand if accuracy trade-offs between field-based and remote sensing-based results may be more important than spatial or temporal detail.
TMTO-based estimates better represent short-term dynamics that often do not show up in biomass estimates from INVE-models. Long intervals between field-based inventories lead to mismatches between TMTO- and INVE-based biomass estimates. Differences are particularly obvious along forest stand borders, where, e.g., windbreak frequently caused great dynamics. Furthermore, the INVE-based biomass estimates for the whole southern part of the study area were overestimated compared to the satellite-based estimates. This is mainly related to temporal inconsistencies in the reference data (see
Section 6.2).
Further differences between INVE- and TMTO-based biomass estimates relate to the spatial heterogeneity within forest stands. SBI-based estimates provide averaged biomass values for each stand, ignoring the spatial distribution of biomass and ecological complexity on the sub-stand level (
Figure 6). Thus, stand-based inventory details may be sufficient to accomplish forest management tasks, while the same information may not be sufficient to describe temporal and spatial dynamics in forest ecosystems, e.g., related to carbon sequestration potential.
6.2. Innovation and Limitations
The availability of ground-truthed data and a homogeneous forest structure in our study site facilitated the evaluation of different models’ predictive power. Nevertheless, a few uncertainties remained. Firstly, different explanatory variables might not perfectly match each other due to temporal differences in our reference data. SBI data from different forest districts are collected independently of one another and consequently also in different years. The latest SBI data updates were available for 2003, 2004 and 2007 in the different districts, respectively. Thus, the extent of windbreak after the destructive storms in November 2004 and following sanitary cutting in 2005 might be missed in some of our district’s SBI. Similarly, Landsat data from early September 2005 might not reflect the total extent of sanitary clearcut measures. Thus, inevitable time shifts between SBI, Landsat data and field measurements (2006, 2007) do not reflect exactly the same forest situation.
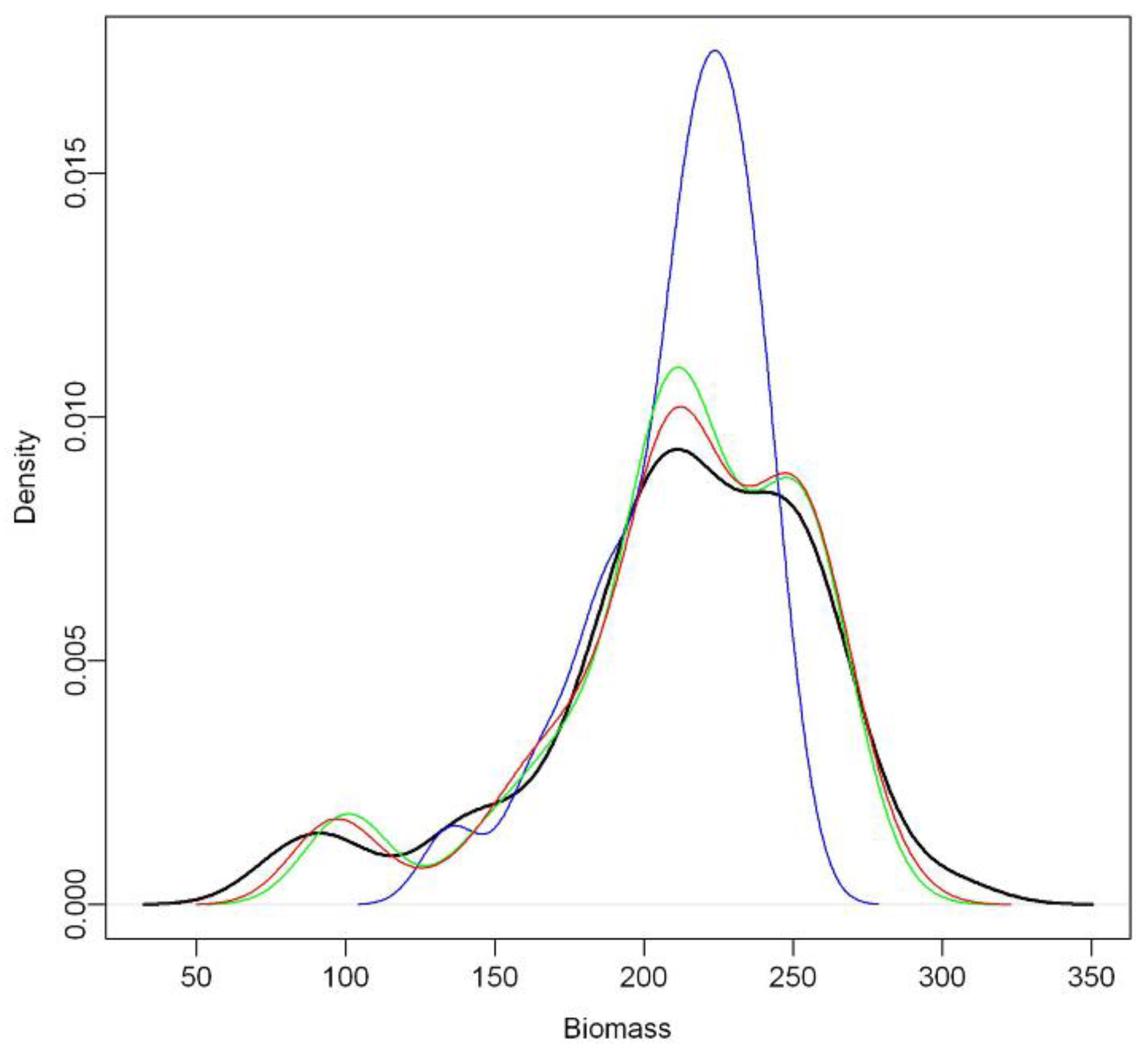
Secondly, the model estimates tend to bias for low and high target values. We found slight over- and under-estimates for lower and higher biomass values, respectively. The tendency to over-predict low biomass and under-predict high biomass conditions (
Figure 3) has also been identified in previous studies, for instance where regression tree techniques [
54] or kNN algorithms [
55] were applied. This tendency is most likely related to two factors: optical sensors have a limited sensitivity for reproducing the canopy structure in dense forests, where biomass can lead to saturated measurements. There is also the confounding effect of understory vegetation in sparse and/or young forests, where forest biomass is generally low. We illustrated these effects in the example of AGB by comparing relative stand density with observed and predicted biomass values (
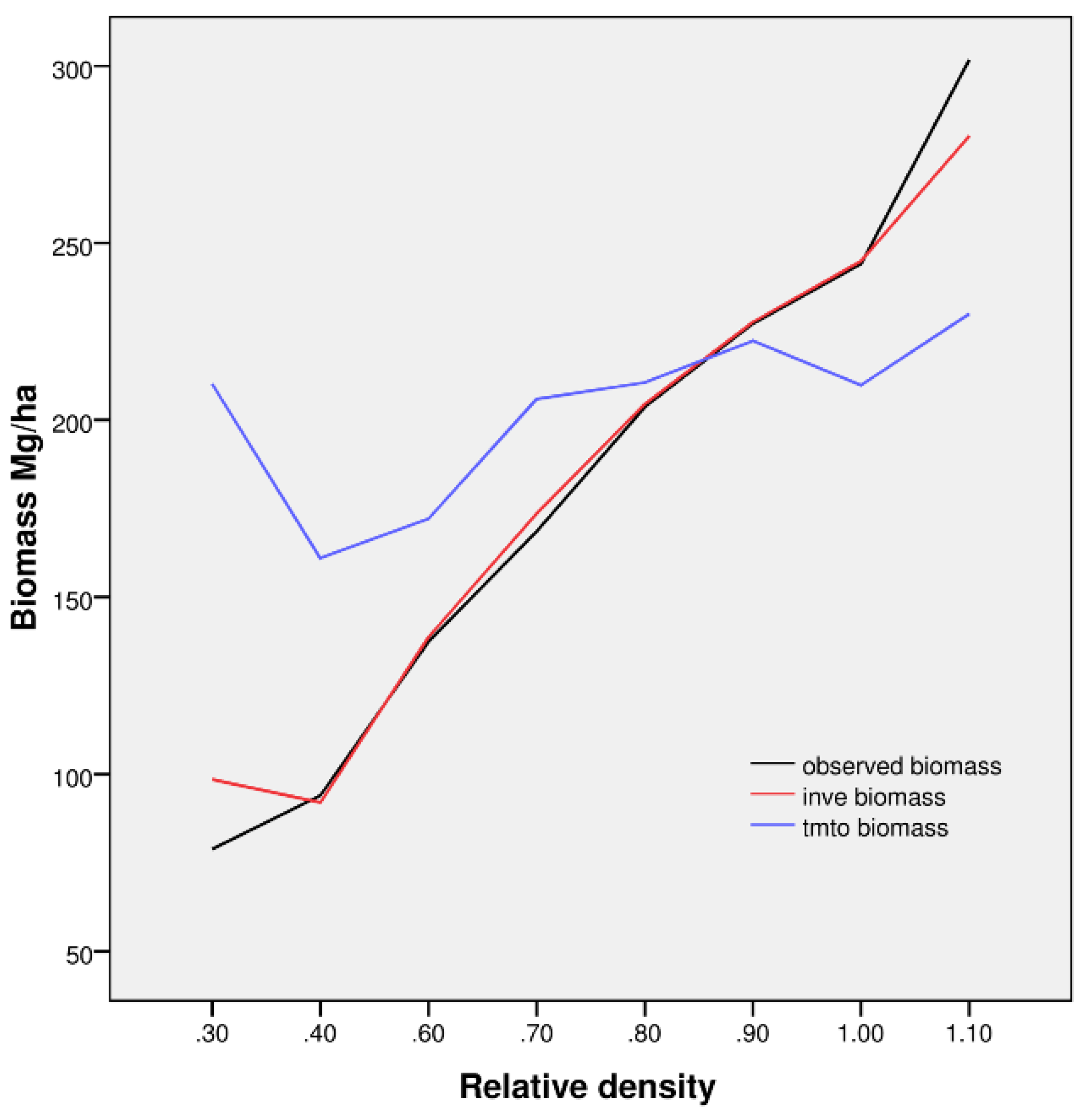
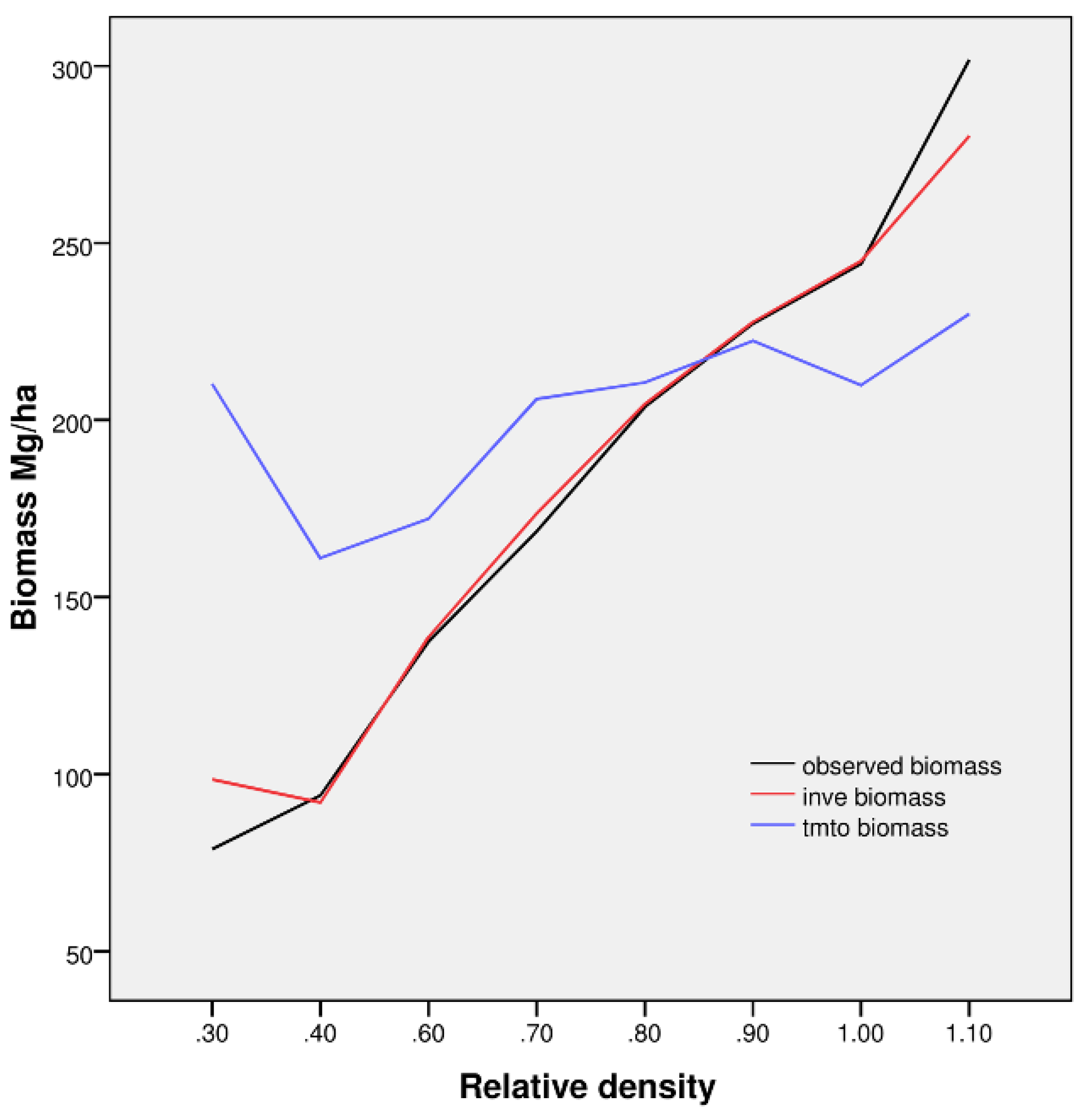
Figure 7). AGB is highly positively correlated with relative density, as shown for observed biomass (reference) and INVE-based estimates. TMTO-based estimates revealed only a moderate relationship to stand density. Regarding dense forests, we observed considerable underestimates of TMTO-based biomass for relative forest densities above 0.9. Additionally, overestimated TMTO-based biomass values, as found in the case of low relative stand density and in the presence of understory as well as underwood, confirms their confounding effect in spectral data. Moreover, overestimation of biomass for spruce stands with tree age below 50 years may also be related to a low sample representation (about 3%) of young forest stands in the modeling process.
Figure 7.
Relative stand density, observed biomass (black line) and predicted values of biomass for models INVE (red line) and TMTO (blue line) for all of 105 stands we sampled in summer 2006 and 2007.
Figure 7.
Relative stand density, observed biomass (black line) and predicted values of biomass for models INVE (red line) and TMTO (blue line) for all of 105 stands we sampled in summer 2006 and 2007.
Thirdly, while spruce stands in our study area and Ukraine are very similar, differences due to regional climate variability and genetic origin exist. Minor inaccuracies in model parameterization and hence biomass estimates can therefore not be completely ruled out.
An integrated modeling approach combining SBI and satellite data has obvious advantages for determining spatial variability in forest biomass and structure. Our findings demonstrated that the integrated use of satellite data compliments stand level inventories, though the latter had the greater predictive strength for aboveground biomass if SBI information is updated regularly. Nevertheless, field-based information alone will, due to time- and work-intensive updating procedures, not deliver all the necessary information in such a dynamic region within a reasonable time frame.
Combining SBI and satellite data (ALL) did not improve the accuracy of biomass estimates. However, an integrated approach facilitates better detection of horizontal structural heterogeneity in forest systems [
56], thereby guiding ground-based sampling intensity and distribution. In addition, it would not just reveal important information about mean carbon stocks, but also about the spatial distribution in stand structure and ecological complexity with which biomass is correlated (
Figure 6). The capacity to map spatial co-variation among multiple ecosystem attributes has important implications for sustainable forest management and planning where the objective is to optimize the provision of multiple ecosystem services, including carbon storage, timber harvest, and biodiversity [
57,
58].



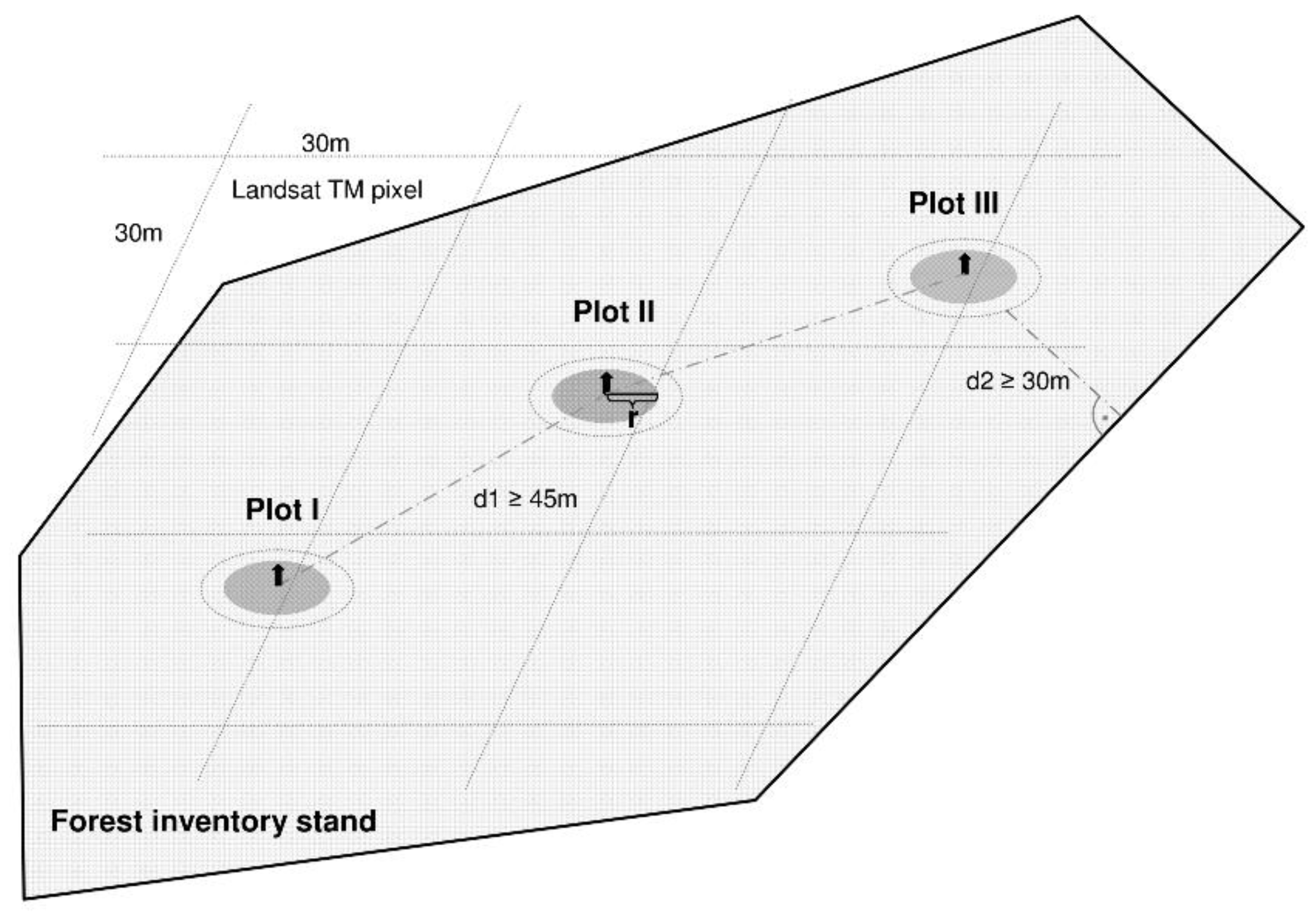


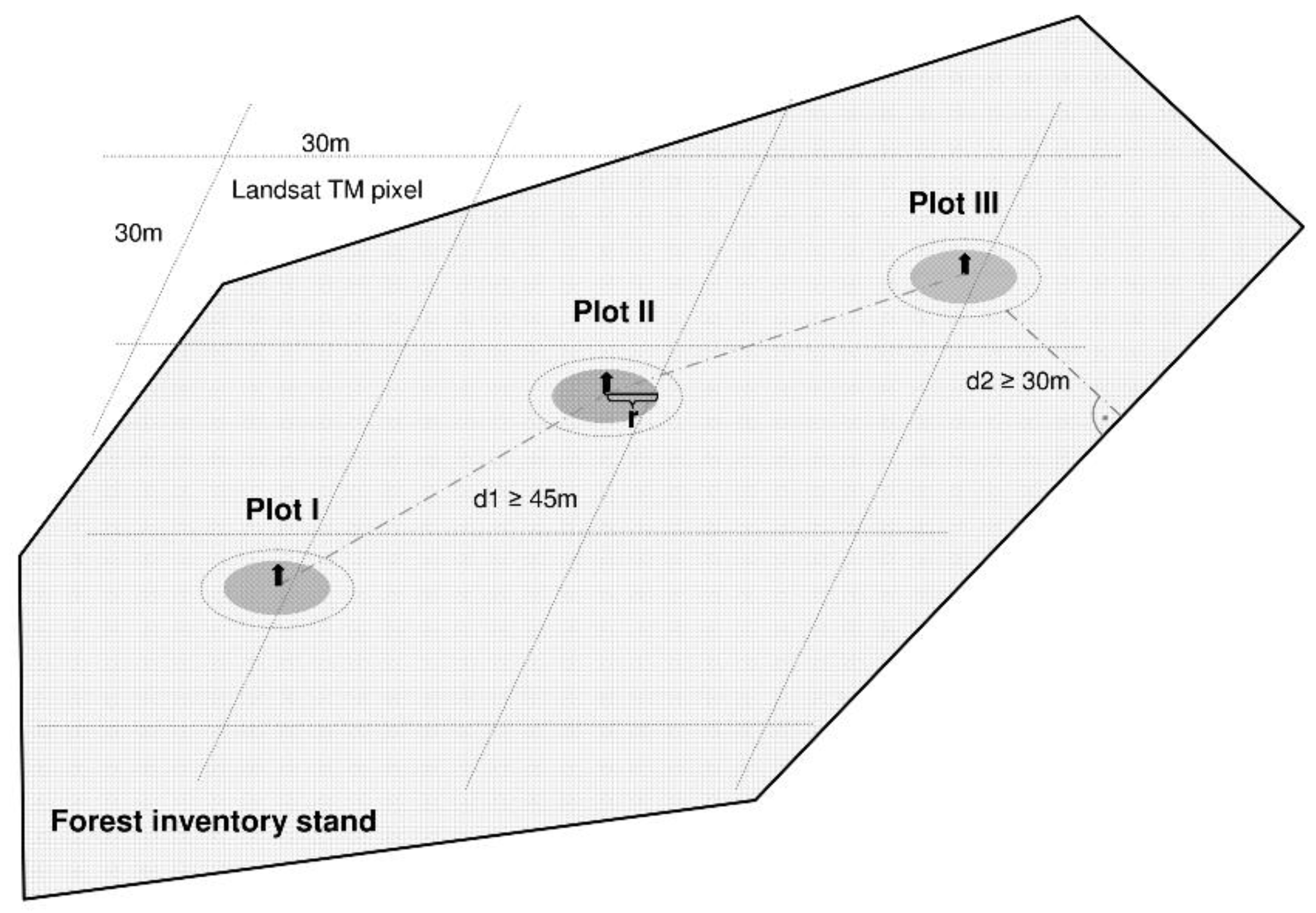
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}