Hyperspectral Distinction of Two Caribbean Shallow-Water Corals Based on Their Pigments and Corresponding Reflectance

1
NASA Postdoctoral Program, NASA Ames Research Center, MS 245-4, Bldg 245, Rm. 120, P.O. Box 1, Moffet Field, CA 94035, USA
2
Earth Science Division, NASA Ames Research Center, MS 245-4, Bldg 245, Rm. 120, P.O. Box 1, Moffet Field, CA 94035, USA
3
Bio-optical Oceanography Laboratory, Department of Marine Sciences, University of Puerto Rico, Mayaguez, PR 00680, USA
*
Author to whom correspondence should be addressed.
Remote Sens. 2012, 4(12), 3813-3832; https://doi.org/10.3390/rs4123813
Submission received: 30 September 2012
/
Revised: 20 November 2012
/
Accepted: 27 November 2012
/
Published: 30 November 2012
Abstract
:The coloration of tropical reef corals is mainly due to their association with photosynthetic dinoflagellates commonly known as zooxanthellae. Combining High Performance Liquid Chromatography (HPLC), spectroscopy and derivative analysis we provide a novel approach to discriminate between the Caribbean shallow-water corals Acropora cervicornis and Porites porites based on their associated pigments. To the best of our knowledge, this is the first time that the total array of pigments found within the coral holobiont is reported. A total of 20 different pigments were identified including chlorophylls, carotenes and xanthophylls. Of these, eleven pigments were common to both species, eight were present only in A. cervicornis, and three were present only in P. porites. Given that these corals are living in similar physical conditions, we hypothesize that this pigment composition difference is likely a consequence of harboring different zooxanthellae clades with a possible influence of endolithic green or brown algae. We tested the effect of this difference in pigments on the reflectance spectra of both species. An important outcome was the correlation of total pigment concentration with coral reflectance spectra up to a 97% confidence level. Derivative analysis of the reflectance curves showed particular differences between species at wavelengths where several chlorophylls, carotenes and xanthophylls absorb. Within species variability of spectral features was not significant while interspecies variability was highly significant. We recognize that the detection of such differences with actual airborne or satellite remote sensors is extremely difficult. Nonetheless, based on our results, the combination of these techniques (HPLC, spectroscopy and derivative analysis) can be used as a robust approach for the development of a site specific spectral library for the identification of shallow-water coral species. Studies (Torres-Pérez, NASA Postdoctoral Program) are currently underway to further apply this approach to other Caribbean benthic coral reef features. The data will be used with planned and future airborne and satellite studies of the site and for algorithm development to advance the use of future airborne and satellite instrument capabilities (NASA PRISM and HyspIRI) for discrimination of coral reef benthic composition.
1. Introduction
Tropical coral reefs are among the most important and complex marine ecosystems on Earth. These living systems provide the primary framework for the development, growth and survival of thousands of species in the tropics, mainly due to the skeletal buildup of Scleractinian corals. Due to their high biodiversity and complex benthic composition resulting in morphological heterogeneity, these important ecosystems present spectral challenges when mapped with remote sensing instruments.
Past efforts on the identification of reef components using remotely-sensed images or spectroscopy have mostly been concentrated at the ecological group level (i.e., corals vs. seagrass vs. algae vs. carbonate sands, etc.) [1–7]. Many coral species may look very similar and corals are usually lumped together in just one category, independent of their color. Some studies have discriminated by general coral color categories (e.g., brown vs. blue corals) [8]. Nonetheless, the presence of photosynthetic and photoprotective pigments within the tissues of reef corals and other Cnidarians, due to their symbiotic relationship with dinoflagellates commonly known as zooxanthellae, might vary and may present an advantage for the distinction of coral reef benthic types in remotely-sensed images. Dinoflagellates are known to contain characteristic pigments [9], particularly zooxanthellae which contain peridinin [10], and a different xanthophyll cycle than those of higher plants [11]. Variations in the concentration or proportions of photosynthetic or photoprotective pigments/compounds (i.e., chlorophylls, carotenes and xanthophylls) between different zooxanthellae clades harbored by different coral species can influence the spectral signal received by spectroradiometers.
In shallow waters (<3 m), the congenerate species Acropora cervicornis and Acropora palmata host indistinguishable Symbiodinium clades; while in deep waters (>15 m), where A. palmata generally does not occur, A. cervicornis hosts a different Symbiodinium clade that is only distantly related to those found in shallow waters [12,13]. This suggests that different algal symbionts enable coral hosts to live competitively in different photic habitats, and that the photic requirements of the algal taxa are conserved features of their biology. Although A. cervicornis hosts two different taxa of zooxanthellae, individual colonies do not contain them both simultaneously [14,15], as it is in the case of other Caribbean coral species like Montastraea faveolata and Porites astreoides[16].
Zooxanthellae were originally thought to be members of the single species Symbiodinium microadriaticum Freudenthal, but further evidence showed that they are a very diverse assemblage [15,17]. Possibly there are at least seven genera in four orders of dinoflagellates [17–20], which can be included into the generic name of “zooxanthellae” [17], and at least four of these inhabit Scleractinian corals [19]. In A. cervicornis, zooxanthellae type “C” (C12) appears only after a depth of approximately 10 m, while type “A” (A3) can be found between 0 m to 10 m deep [16]. A similar relationship can be found also in the mountainous corals Montastraea annularis and M. faveolata[11]. Porites porites, on the other hand, hosts only type “C” of zooxanthellae through its depth span [15], particularly C10. In fact, type “C” is probably the most diverse of the clades of zooxanthellae known [16] being dominant in the Caribbean and the Pacific as well [13,14,16,20].
This study investigates the possible influence of the differences in photosynthetic and photoprotective pigments composition of the shallow-water Caribbean corals A. cervicornis and P. porites on their respective reflectance as measured by a field spectroradiometer. The concentration of these pigments/compounds was obtained with High Performance Liquid Chromatography (HPLC) analysis. A derivative analysis was applied to the reflectance curves of each species to investigate whether particular absorption peaks associated with specific pigments could be identified. Derivative analysis, particularly using the 2nd derivative, is useful for identifying characteristic pigments absorption peaks influencing the reflectance spectra [21–23]. Further, the wavelength and magnitude of the derivative values for a given spectral signature of a particular species can be correlated with the characteristic absorbance peaks of the different pigments confirmed by HPLC analysis. The appealing results of the present study show a distinction between A. cervicornis and P. porites at wavelengths associated with the absorption of particular pigments. Using a combination of spectroscopy and pigment analysis, these techniques provide a possible novel approach to the improved characterization of high spectral resolution images to the species level, given the appropriate spatial resolution.
2. Experimental Section
2.1. Collection Site
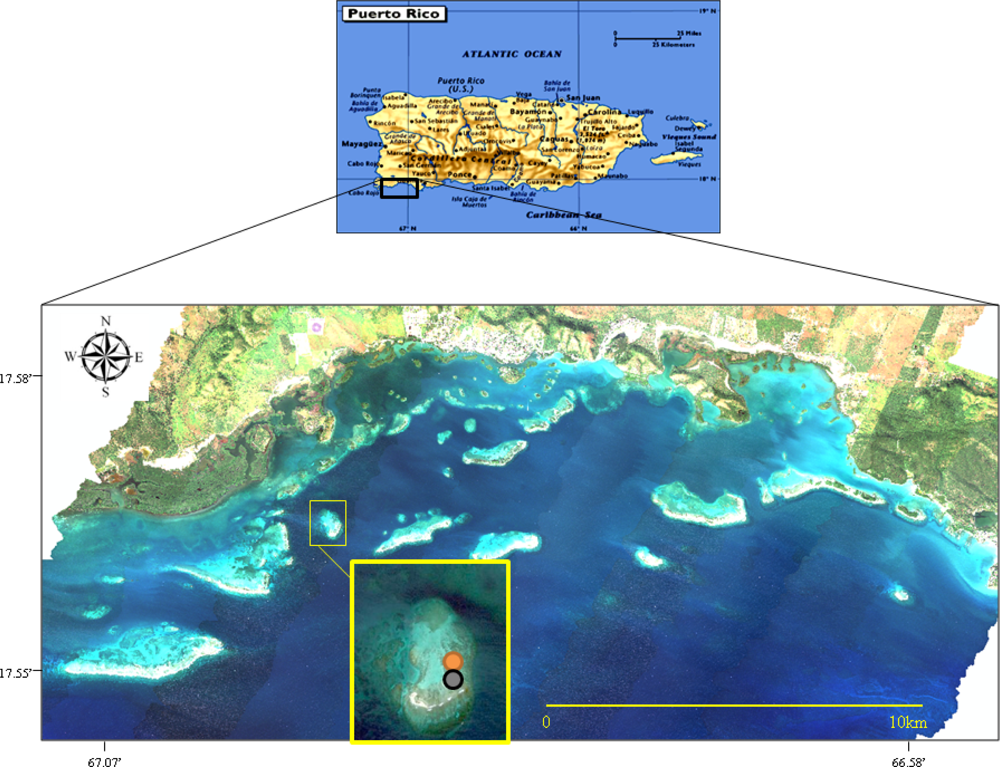
San Cristóbal Reef is a patch reef within the La Parguera reef platform, southwest Puerto Rico (17°56′41″N; 067°04′38″W) (Figure 1). La Parguera is one of the most studied coral reef platforms in the Caribbean. The reefs on this area are typically compose of a back-reef zone dominated by sand and isolated coral patches, a seagrass bed, a reef front dominated by fire and branching corals, and a fore-reef zone dominated by massive and plate corals. The isolated coral patches of the back-reef zone are mostly dominated by colonies of the threatened elkhorn and staghorn corals (Acropora palmata and Acropora cervicornis, respectively), finger corals (Porites porites and Porites furcata), and some massive coral species such as Montastraea annularis, Diploria strigosa and Siderastrea siderea.
2.2. Coral Colonies Sampling
One branch (5–8 cm long) from individual colonies of A. cervicornis and P. porites (n = 5) were collected at 1 m depth (here both species dominate the benthic coral cover) from the back-reef area of and transported to the University of Puerto Rico’s Department of Marine Sciences Magueyes Island Field Station for immediate processing. Collected colonies were spaced no less than 20 m apart. This spacing was set to avoid pseudo-replication or sampling of genetically similar ramets. Care was taken so only visibly “healthy” branches were collected (e.g., absence of any visible disease, discoloration or bleaching compared to the rest of the colony or its neighbors). All the colony branches were transported in aerated seawater in sterilized Whirl-Pak plastic bags and taken to an outdoor aquarium upon arrival. The whole transport operation was completed in less than 30 minutes. Upon arrival, the branches were once more visually inspected for any damage that might occur during transportation. Also, if detected, any epiphytes were removed from the skeleton with sterilized forceps before any analysis. Spectral data was obtained immediately upon arrival followed by pigment extraction.
2.3. Spectral Analysis
A GER-1500 field spectroradiometer (SpectraVista Corp.) equipped with a fiber optic cable was used to collect spectral data. This instrument has a spectral range of 280–1,100 nm (512 spectral bands) with a Full Width at Half Maximum (FWHM) equal to 2.8 nm. The tip of the cable was held less than one centimeter from the specimen in an outdoor seawater aquarium to ensure no signal contamination due to the presence of dissolved or particulate matter in the water column. Three radiance measurements were taken from different parts of each branch and averaged. These replicate measures were used to study possible differences in the reflectance along each individual branch. Also, radiance measurements from a 50% opaque diffuse barium sulfate (BaSO4) reference Spectralon® panel (Labsphere Inc.) were taken immediately after obtaining the sampled spectral data at the same distance to eliminate atmospheric effects. Coral-leaving radiance was converted to reflectance using:
where R is the reflectance, Lc is the coral-leaving radiance, Lp is the diffuse surface panel-leaving radiance and 0.5 is the reflectance factor of the diffuse panel. The spectra were smoothed with a low pass Savitzky-Golay filter [24] prior to derivative analysis to eliminate frequencies with periods shorter than 4 nm. A derivative curve was obtained with the Microcal’s Origin® analysis package by calculating differences between readings along specific wavelengths intervals. The 2nd derivative curve is the differentiation of the first curve; the 3rd derivative is the differentiation of the 2nd curve, and so on [25,26]. Only the 2nd derivative was used in this study to enhance spectral features. Differences in derivative peaks at particular wavelengths were compared to the presence/absence of pigments characterized by having absorption maxima at or near those wavelengths. Hypsochromic wavelength shifts [27] were considered during the analysis. A similar derivative analysis was applied to the absorption curves of each species obtained with the spectrophotometer.
2.4. Pigment Extraction and Analysis
Immediately following the spectral data collection, each branch was processed for pigment analysis. The white tip of the branches was discarded as it typically contains only a negligible amount of zooxanthellae and pigmentation [28]. Pigments were extracted with a methanol:tetrahydrofuran (80:20, v/v) solution, in the dark at 4 °C for 24 h to prevent photo-oxidation [11]. A second 20-min extraction in the dark at 4 °C was performed to ensure total pigment extraction. Photosynthetic pigments were separated from UV-absorbing compounds by injecting the extracts through a Sep-Pak C18 cartridge [29–31]. Absorbance measurements (400–700 nm) were obtained with a Hewlett-Packard 8452A diode-array spectrophotometer. These were used to calculate the overall chlorophyll and carotenoid concentration using the equations of Jeffrey and Humphrey [32]. Since these chromatic equations do not resolve for accessory photosynthetic pigments [33], reversed-phase HPLC was performed. Pigments were separated in a Shimadzu HPLC system using a gradient system of 80:20 methanol:ammonium acetate (pH 7.2, v/v), 90:10 acetonitrile:water, and 100% ethyl acetate with a Symmetry® C18, 25 cm × 3.9 mm-inner diameter, 5 μm particle size column at a flow rate of 1.0–1.8 ml·min−1 for 30 min. Eluting peaks were detected using the absorbance spectra at 436 nm for carotenes and chlorophylls following established protocols [34–37]. Peaks were integrated, and quantification of individual pigments was accomplished using peak areas and calibration factors determined by analysis of authentic standards of chlorophyll a and lycopene (Sigma Corp.). Individual pigments were identified using published signatures, retention times and their respective peak maxima [11,34,35,38,39]. Coral tissue area was determined following the aluminum foil technique of Marsh [40]. Concentrations of pigments/compounds were expressed in μg·cm−2.
2.5. Relationship between Reflectance and Pigment Concentration
A regression analysis was also performed to study a possible relationship between coral reflectance and the total pigment concentration. The area under each reflectance curve was integrated using:
where Rλ1, …, Rλn is the coral’s reflectance at λ1, …, λn. Total pigment concentration was obtained from the HPLC analysis described in Section 2.3.
2.6. Statistical Analysis
A two-sample t-test was used to test for differences in total pigments, chlorophylls, carotenes and xanthophylls concentration among the two studied species. A principal components analysis (PCA) was performed on the main pigment data with the PRIMER 6 statistical package (PRIMER-E Ltd.) to test for variations among both species main pigments (i.e., Chl a, Chl c2, Per) and pigment groups (chlorophylls, carotenes and xanthophylls). PCA has been used in the past as a data reduction technique and as means to identify different modes of data [1,41]. The 12 variables input into the PCA resulted in a matrix of the main individual pigment concentrations, relative individual pigment composition, pigments groups and relative pigment group composition (see Table 2 in Section 3.1) all by species (e.g., A. cervicornis and P. porites). Differences among the two species reflectance curves were tested using a contingency chi-square test. Differences in peak heights between species were tested with a Tukey test with pairwise comparisons. Statistical significance was set at α ≤ 0.05.
3. Results and Discussion
3.1. Influence of Coral Symbionts and Endolithic Algae on the Pigment Composition of Corals
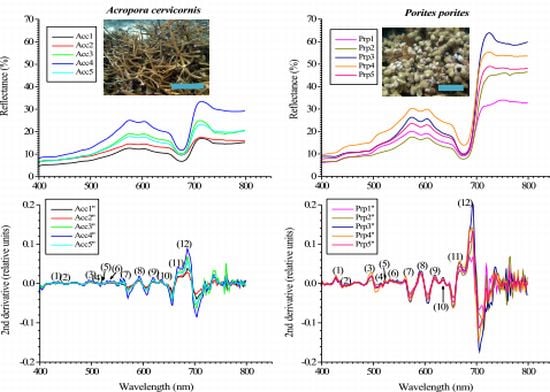
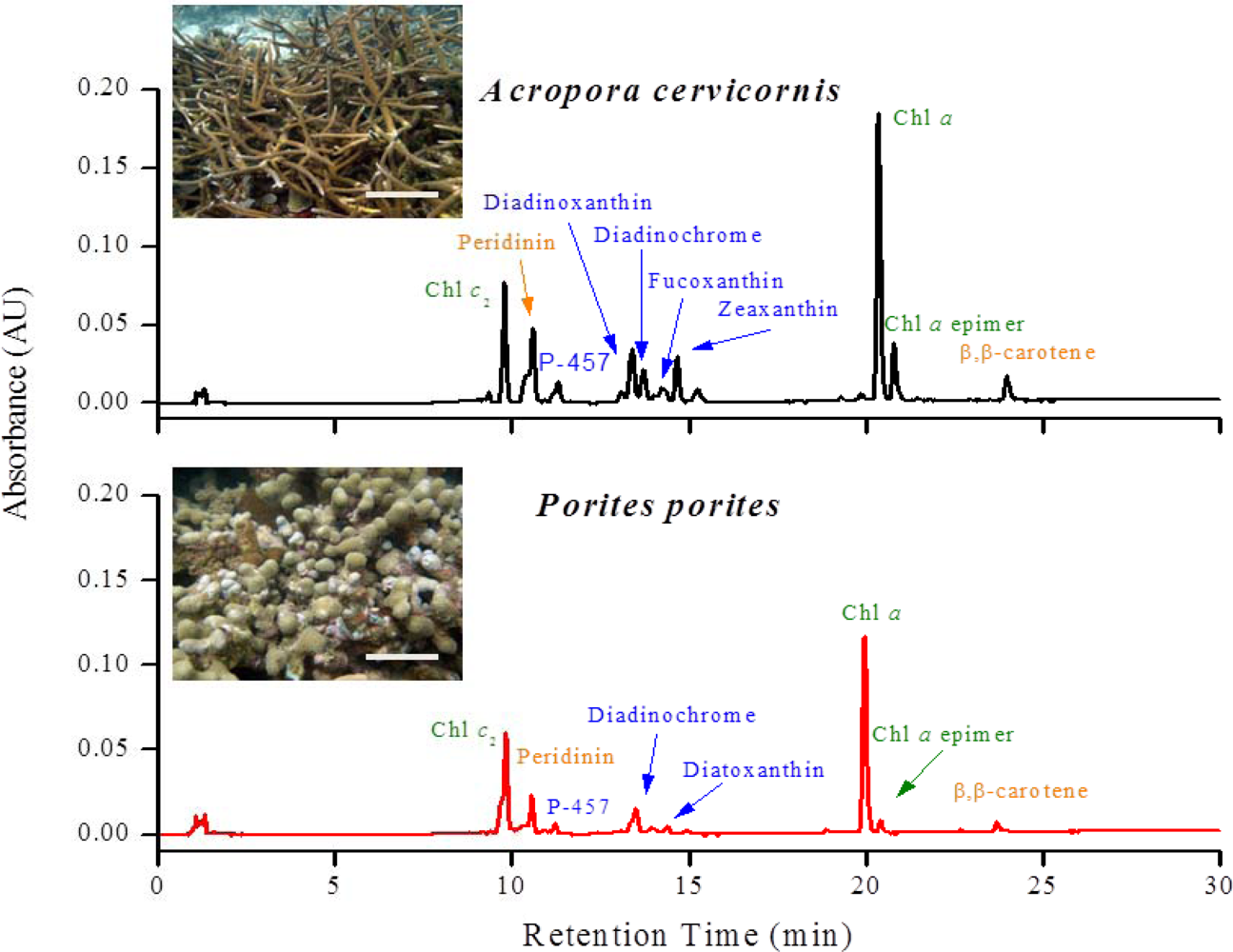
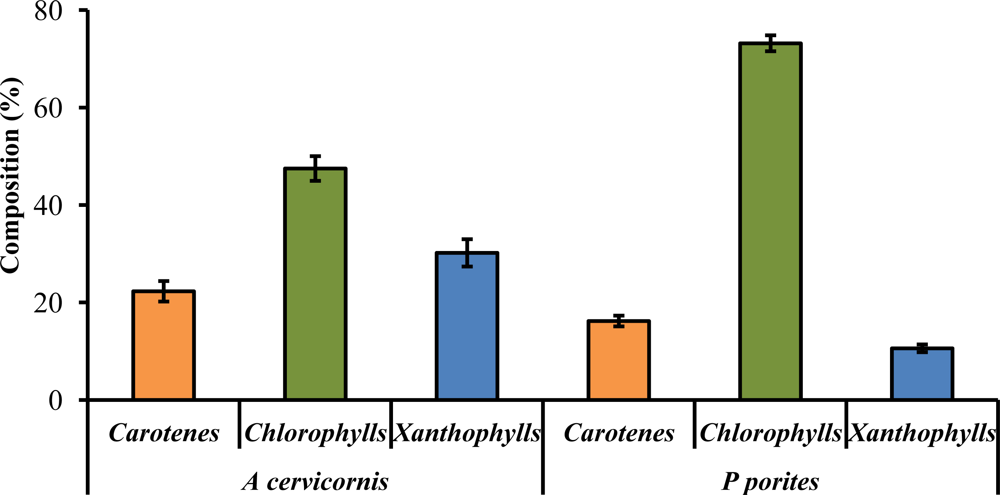
Twenty different pigments were identified between A. cervicornis and P. porites, including chlorophylls, carotenes, and xanthophylls (Table 1). So far, of the 20 pigments found, Per is the only pigment specific to zooxanthellae [39] (for abbreviations of pigment names, see Table 1). A characteristic chromatogram, for each species, resolving major pigments is shown in Figure 2. None of the P. porites colonies sampled contained Dd. Other pigments found (for example, Zea, Fc, An) are not taxonomic indicators since they can be found in other algal groups [38,39]. Nine pigments were common in all the samples of both studied species: Chl a, Chl a epimer, Chl c2, Per, P-457, Dc I and II, Nc, and β,β-car. Significant differences were found in total pigments concentration (Two-sample t-test, t = 5.58, P = 0.003) between species, and in total chlorophylls content (Two-sample t-test, t = −16.91, P < 0.00001), total carotenes content (Two-sample t-test, t = 16.98, P < 0.0001) and total xanthophylls content (Two-sample t-test, t = 10.54, P < 0.0001) between species (Figure 3). Chlorophylls in P. porites constitute approximately 71% of the total pigments, a 1.5x difference compared to those of A. cervicornis, whereas the latter contained a greater array of xanthophylls compared to the former. In fact, seven xanthophylls were only found in A. cervicornis and not in P. porites.
In the PCA, the 12 principal components were evaluated along with the PC eigenvectors, which are linear combinations for the PC axes rotation. The eigenvectors for the variables analyzed revealed which variables were most influential in the resultant differences between the two studied species. The PCA indicated that the first two principal components explain 86.7% of the variation in pigment between species. The first Principal Component (PC1—composed of the total concentration of chlorophylls and the percent of Chl a relative to the other pigments or pigment groups, namely carotenes and xanthophylls) is responsible of 69.4% of the variation. The second Principal Component (PC2—composed of the total concentration of chlorophylls and the percent of xanthophylls) is responsible to 17.3% of the variation. A third Principal Component (PC3—composed of the percent of Chl a and Per relative to the other pigments or pigment groups) only added an additional 10% to the explained variation. According to the PCA, other factors such as the percent of carotenes, and the concentration of Chl c2 and Per have a minimal effect on the differences in pigmentation between these two studied species (Table 2).
Early studies of Caribbean corals [10] found no significant differences in the relative concentrations of photosynthetic pigments (Chl a, Chl c2, Per, Dn and β,β-car), while the total pigments content did differ significantly. In the present study, we have found that, at least between A. cervicornis and P. porites, both the relative concentrations and the total pigment content did differ. Our results expand the range of Caribbean species analyzed by Gil-Turnes and Corredor [10]. The presence or absence of particular photosynthetic or photoprotective pigments depends on the group studied [39]. Typical pigments associated with reef coral symbionts include Chl c2, Per, Dd, Dt, and β,β-car [28,42]. Here we have presented not only the major pigments associated with reef coral dinoflagellate symbionts, but we also present the contribution of several minor pigments as well. These other minor pigments are not necessarily associated with dinoflagellates but other photosynthetic organisms that were probably present within the coral holobiont and could affect the spectral signal (e.g., reflectance) of a particular coral colony. For instance, Fc and Dd can also be found in diatoms, chrysomonads and silicoflagellates [38,39]. Further the xanthophyll cycle, which dissipates excess energy through non-photochemical quenching [43], differs depending on the photosynthetic organism. While in dinoflagellates and diatoms, the xanthophyll cycle depends on the conversion between Dd and Dt, in higher plants and green algae relies on the conversion between violaxanthin, An and Zea. Hence, the presence of An and Zea in A. cervicornis can most likely be due to the presence of intrusive microalgae within the coral skeleton. Likewise, finding Fc and Lu in the HPLC analysis can be indicative of the presence of different types of brown and green algae, respectively [3,44].
Dc I and II, on the other hand, are alteration products of Dd [38] in dinoflagellates as well as Nc is of neoxanthin. The presence of such pigments in all the samples may either be indicative of alterations in the mechanism of the photosynthetic apparatus or a rearrangement during the extraction processes. As explained in the methodology section, care was taken in selecting the colonies to be sampled. Colonies showing signs of some sort of stress were not collected. Nonetheless, this inspection was only performed visually; hence, while less likely, we cannot categorically discard that an alteration of the photosynthetic apparatus happened. A possible rearrangement of pigments during the extraction is more probable. Our previous experiences with coral samples showed incomplete extraction of pigments even after several hours. As such, we chose a 24 h extraction time in the dark at 4 °C based on previously established protocols [36,37,45]. Therefore, the finding of pigments like Dc I, Dc II and Nc in the extracted samples should not necessarily be indicative of their presence as part of the in vivo array of pigments typical of the coral holobiont’s photosynthetic apparatus. Similarly, pigments associated with endolithic algae, such as Fc, Zea and An, might not be present in every single colony of a particular coral species as it will depend on the presence or absence of these algae. Nonetheless if present, these pigments should be taken into account in similar studies as they contribute to the total absorption of visible light and, hence, have an impact on the reflectance of a particular coral colony containing them.
The concentration of xanthophylls that are involved in the dissipation of excess energy by non-photochemical quenching vary enormously depending on the incident radiation and the previous light history of the algae [28,42,45] as well as the variation on the incident radiation. The samples from both of the species studied here, while living at the same depth, were collected between 900 to 1,000 h, and this might have influenced the results, particularly of the xanthophyll pool. Therefore, a possible conversion of Dd to Dt at other hours in the day in A. cervicornis as well as from Dt to Dd in P. porites, cannot be discarded.
3.2. Reflectance Spectra and Derivative Analysis
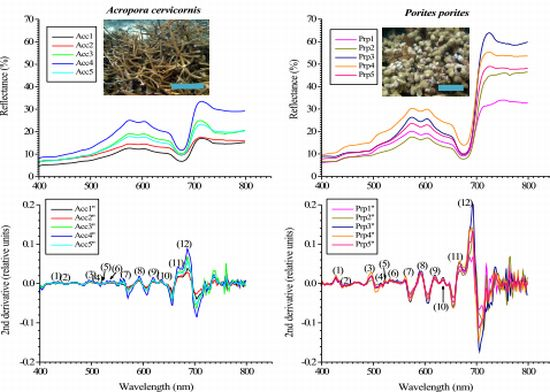
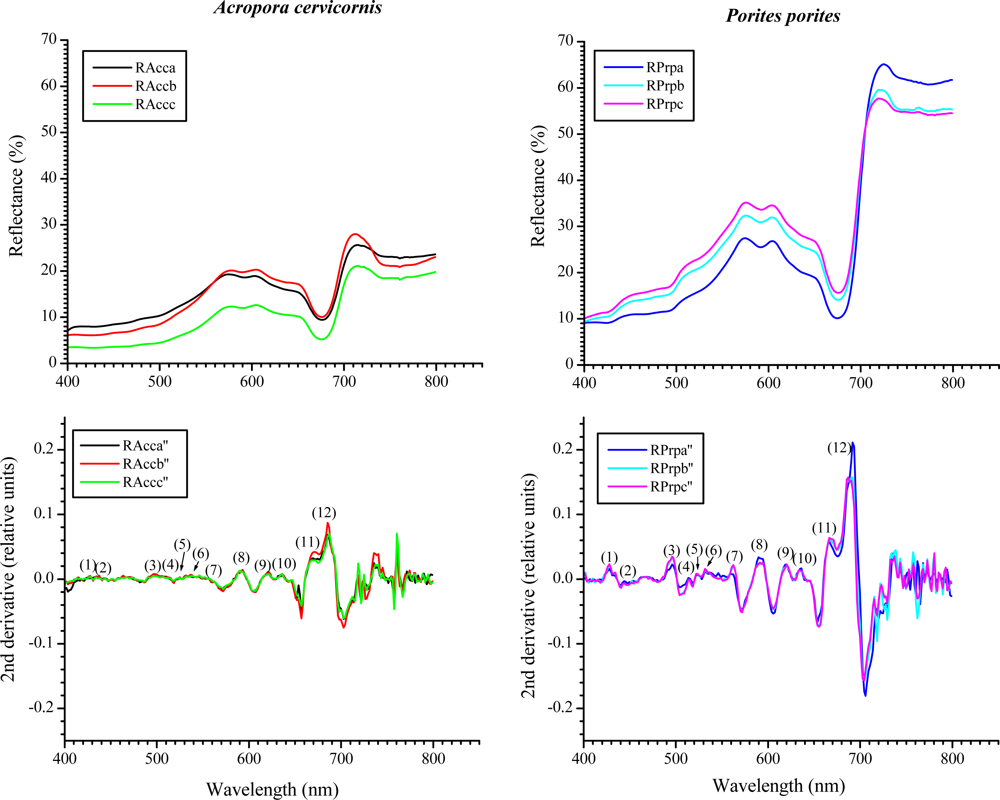
Within the visible range (400–700 nm) most of the features were very similar between the studied species’ reflectance (Figure 4). Nonetheless, a distinctive increase-in-reflectance feature can be seen in P. porites between approximately 426–462 nm, not present in A. cervicornis (Figures 4 and 5). The 2nd derivative of the reflectance curves shows differences between the two species through the visible spectrum with significant different peaks in the blue and red regions particularly around 427, 442 and 495 nm, 634, 665 and 685 nm, respectively. Some of these wavelengths coincide within ±4 nm of the maximum absorbance peaks of various chlorophylls and xanthophylls. For example, the Dd and Dt concentration and the 2nd derivative peak heights at 442 nm were highly correlated (r2 = 0.79 in A. cervicornis and 0.92 in P. porites, respectively). Similarly, the Chl c2 concentration was correlated with the peak heights at 634 nm (r2 = 0.76 in A. cervicornis and 0.98 in P. porites). At 685 nm the height of the 2nd derivative peaks correlated with the concentration of Chl a (r2 = 0.92 in A. cervicornis and 0.94 in P. porites) showing the influence of the Chl a fluorescence on both species spectra. The use of the 4th derivative is somewhat controversial for spectral analyses as there are potentially several other factors influencing the complex features of the 4th derivative result not relevant to this analysis. The 4th derivative also increases the amount of noise in the data. Hence, we decided not to include any 4th derivative analysis in the present study.
A constant maximum around the 427 nm in the 2nd derivative coincides with the absorption of Dc I and II, whose percentages relative to the total pigments pool in P. porites, double those of A. cervicornis. Nonetheless, Dc I and II are alteration products of Dd and could have developed during the extraction process; hence they were probably not present when the reflectance spectra were obtained in vivo. Therefore, the peak at 427 nm is most indicative of the differences in Chl a between the species complemented as well as with the signatures from some of the carotenes and xanthophylls. In fact, the peak heights at 427 nm correlated with Chl a concentration (r2 = 0.94 for A. cervicornis and 0.89 for P. porites). Similarly, the difference in the 685 nm maxima in both species in the 2nd derivative can also be indicative of the difference in the input of Chl a and its derivates, as they compose 71% of the total pigments pool in P. porites compared to 47% in A. cervicornis. Also, the PCA identified the total chlorophylls concentration and Chl a percentage as the main variables responsible for the pigment differences between these species. The more complex array of xanthophylls and carotenes in A. cervicornis, and hence higher absorption within the blue region, is reflected in typically smoother derivatives compared to P. porites. Again, while their influence is apparently minor, the percent of xanthophylls accounted for part of the variability associated with the PC2 in the PCA. Another feature that distinguishes the reflectance spectra of both species appears within the near-infrared region (700–800 nm). This is highly dependent on the amount of scattering within the coral tissue, colony morphology and skeletal characteristics [46]. The reflectance curves of P. porites showed a higher variability than those of A. cervicornis most indicative of higher scattering occurring within the tissue and a higher skeletal density of the former.
Typically the reflectance spectra of corals having similar and distinct spectral features are not common. Nevertheless, absorption bands that are too close together to be resolved as individual peaks in an absorption or reflectance spectrum may be resolved in a derivative spectrum. The 2nd derivative is very useful for qualitative identification of pigments, while the magnitude does not provide a reliable measure of the concentration of pigments since the absorption contributed by overlapping pigments affects the value of the 2nd derivative of the pigment of interest [25,26]. Variations in the magnitude of the reflectance among different specimens is to be expected as minute variations in the exposure to light field may lead to different concentrations of photosynthetic and photoprotective pigments-compounds. Our results show that while there is variation in the magnitude of each specimen reflectance (Figure 4) the general features along each curve are maintained. On the other hand, the reflectance curves of both species were statistically different (Chi-square test, χ2 = 94.74, df = 4, p < 0.0001). We tested whether there were differences in the reflectance along different parts of the same branch of each studied species. Reflectance was very similar along different sites of the branch with spectral features maintained (an example is shown in Figure 5). A One-Way ANOVA showed no statistical differences in 2nd derivative peak heights among specimens of the same species (p = 0.598 and 0.836 for A. cervicornis and P. porites, respectively). This statement is maintained as long as the measurements from the spectroradiometer are taken at the same distance and angle. Measures taken on opposite sides of the branch (i.e., top vs. under side) may lead to different results as they would be exposed to different light regimes.
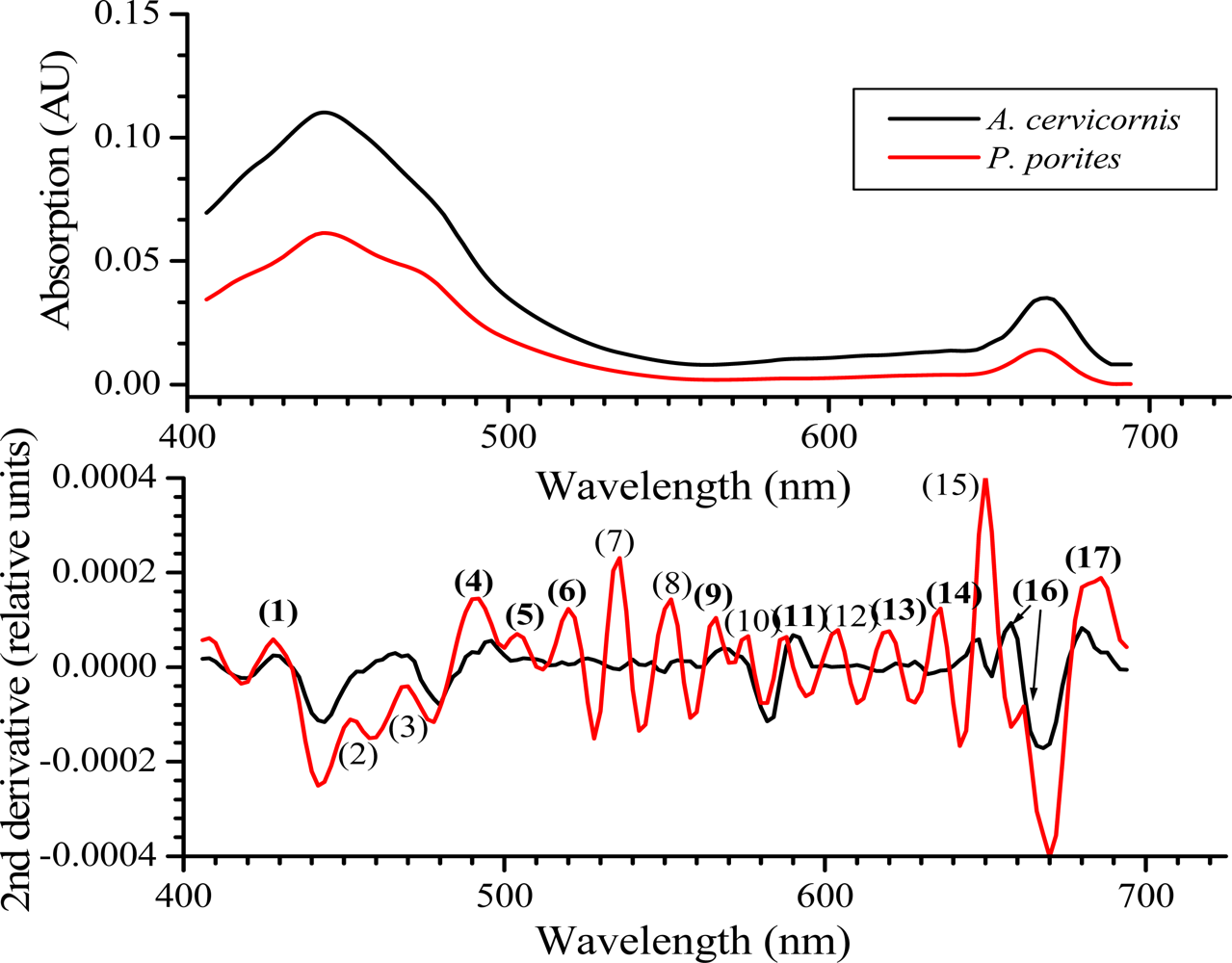
Hypsochromic shift (decrease in wavelength for the maximum absorption wavelengths), typically occurs in pigment absorbance peaks as they are extracted with organic solvents [27]. This result from the dissociation of pigments from the proteins they are associated with in vivo once they become in contact with the solvent. Ths hypsochromic shift should be taken into account when comparing in vivo vs. extracted spectra [22]. For example in the present study, the peaks at 427 and 495 nm in the 2nd derivative of the reflectance spectrum (see Figure 3) appear at 424 nm and 490 nm, respectively, in the 2nd derivative of the absorbance spectrum (Figure 6), and so on.
An additional factor influencing the differences in the reflectance curves is the presence/absence of Green Fluorescent Proteins (GFPs). GFP’s are potential contributors to the coloration of reef corals as it has been demonstrated in the Caribbean coral Montastraea cavernosa[47–49]. The 495 to 589 nm peaks on the 2nd derivative of both species (see Figures 4 and 5) might be a consequence of the presence of GFPs on this species as it has been demonstrated in the past [47]. For instance, recently D’Angelo et al found GFP’s emitting at 490 nm and 584 nm in Porites lobata and Acropora pulchra, respectively [50]. While a possible contribution of GFPs to the reflectance signals of both species cannot be discarded, and no attempts were made to quantify this, we postulate this contribution of GFPs within the blue region, particularly below 450 nm, might be minimal as the emission peaks of GFPs are usually within the green and red regions of the visible spectrum [49].
3.3. Reflectance and Total Pigment Concentration
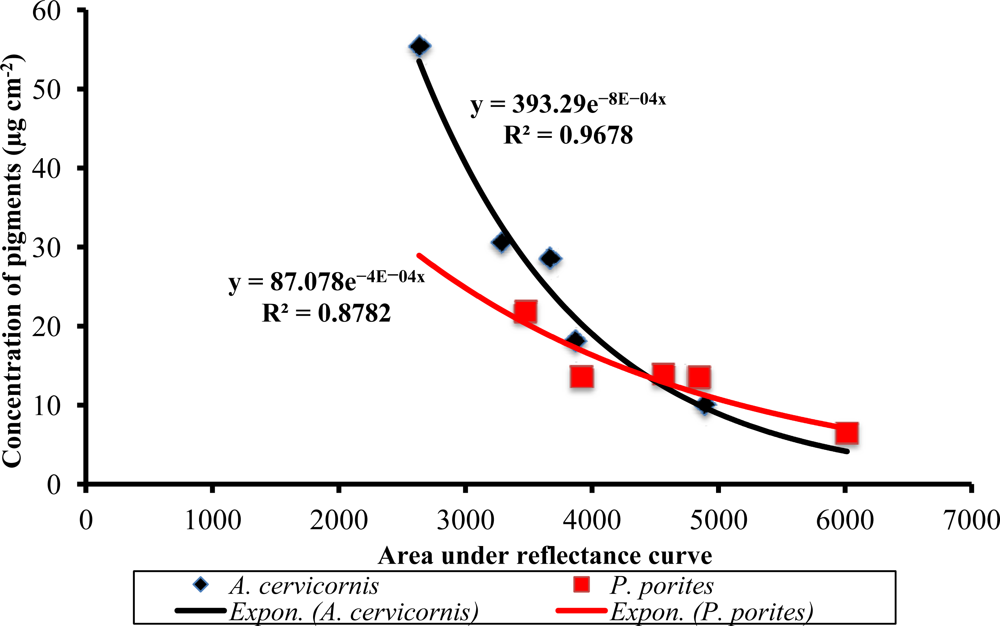
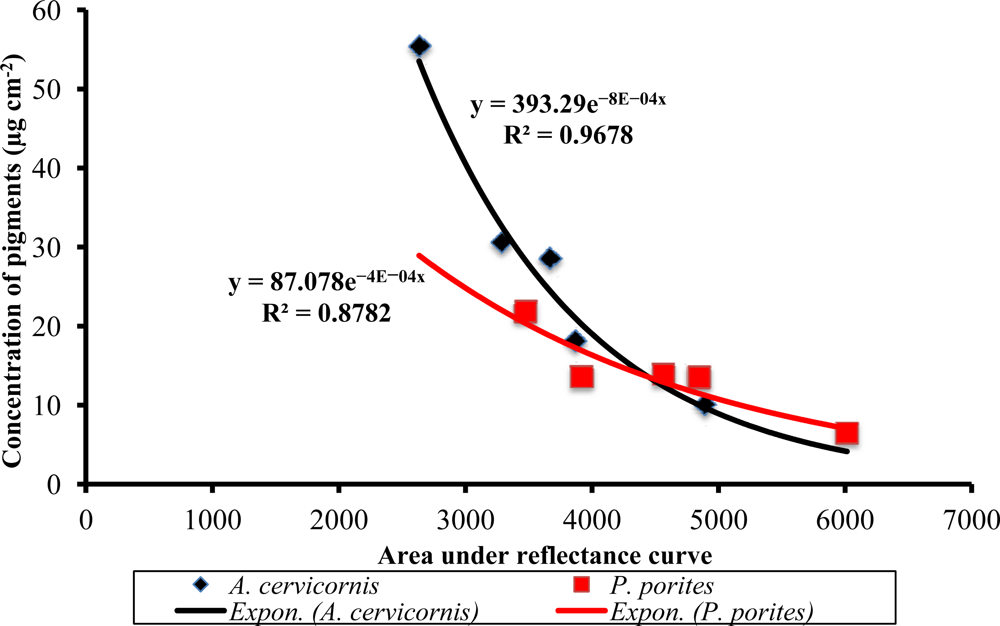
An integration of the reflectance curves was compared to the total pigment concentrations measured by the HPLC. In both species, an inverse exponential relationship was found with an 88%–97% predictability of total pigment concentration (Figure 7), establishing evidence for the use of reflectance curves as a first-approach non-invasive technique for estimating total coral pigment concentrations. Further work in progress by the authors will investigate whether this technique can be used as an application for field spectral assessment studies relevant to the impacts of climate change and coral bleaching/diseases on corals without the need of invasive (or destructive) sampling.
Current satellite and airborne remote sensors (including imaging spectrometers) along with already developed algorithms can broadly differentiate between reef corals, seagrass and sandy areas [4,6,7]. Our results provide some first novel steps towards the differentiation between coral species living in relatively clear shallow waters based on their reflectance and corresponding 2nd derivatives. Moreover, these methods can be expanded to study differences between the species studied here and other coral species. As such, research is currently underway by the authors to explore these possible differences with the aim of fully exploiting the available spectral data set for discrimination of coral reef benthic components in La Parguera patch reefs. This will help to exploit improved analyses of airborne imaging spectrometer data with associated field spectroscopy data available from 2004, 2005, 2007 and 2010 missions.
4. Conclusions
Remote sensing has proven to be a valuable tool to study the status and composition of coral reefs worldwide. Nonetheless, while much work has been done to identify the dominant benthic components of shallow reefs, very few studies have worked on the identification of corals to the species level with remotely-sensed data. This is a novel approach for identifying different coral species using their respective reflectance spectra and the underlying signatures of their corresponding pigments. First, we have reported not only those pigments typically associated with the zooxanthellae of the common Caribbean shallow-water primary framework species such as Acropora cervicornis and Porites porites but also those that occur due to the presence of other microflora associated with the coral holobiont and which can influence the spectral response of a particular coral colony. These different pigment assemblages in turn influence the reflectance of these species. Furthermore, these assemblages are the causative agents of the different inflections typically seen in their respective reflectance curves. Second, derivative analysis of the reflectance curves showed highly significant differences between species in the 2nd derivative peak heights at 472, 442, 495, 530, 589, 618, 634, 665 and 685 nm. The concentration of major pigments were positively correlated with some of these peaks highlighting those at 427, 665 and 685 nm (all three with Chl a), 442 nm (with Dd or Dt) and 634 nm (with Chl c2). Peaks at 495 and 589 nm could be related to the presence of host-associated pigments such as green fluorescent proteins. Third, the High Performance Liquid Chromatography (HPLC) and principal components analysis (PCA) results showed that the pigment differences between these two coral species are mainly due to the relative contribution of chlorophylls and xanthophylls. The first three principal components (PC) explained that 96.7% of the pigment variation between species. For PC 1 (69.4%), the eigenvectors showed that percent Chl a and total chlorophylls combined were the most influential of the variables analyzed. For PC2 (17.3%), the most influential were total chlorophylls combined and percent of xanthophylls. For PC 3 percent of peridinin and percent of Chl a contributed to the 10% variation explained. As such, our results show that although these species live at the same depth, their pigment array is very different most probably due to the presence of different zooxanthellae clades and the harboring of other invasive microalgal species as well. In terms of reflectance features, significant differences were found only when comparing between samples from different species (p < 0.0001). Finally, the area under the reflectance curve correlated with total pigment concentration with an 88% confidence level in P. porites and a 97% in A. cervicornis. Hence, we propose this approach as a very good estimator of total pigment concentration in reef corals. As such, this spectroscopic analysis can possibly be applied to studies of the health status of coral species without the need of invasive techniques. Nevertheless, regarding the applicability of this technique to actual remote sensors, we recognize that due to the difficulties assessing remotely-detected signals from airborne or satellite imagery, much higher spectral resolution is needed to detect the observed signal differences particularly in the blue region of the spectrum. Yet, we believe that this work provides a complementary field-truthing tool for spectral unmixing of coral reef features at fine scales and can be taken into consideration to specify sensor requirements to meet the scientific community’s needs to study changes in the benthic communities of coastal and marine ecosystems. To advance this approach, research is currently underway to include additional coral species and other reef benthic photosynthetic components. This may provide quantifiable measures of reef biodiversity using remote sensing capabilities in the future.
Acknowledgments
We would like to thank Jorge Corredor, Director of the Marine Chemistry Laboratory of the UPR-DMS, and Rocío García and Oswaldo Cárdenas for helping with the spectrophotometric and HPLC analysis. Anabella Zuluaga helped with the statistical analysis of the data. The comments of four anonymous reviewers significantly improved the final version of this manuscript. JL Torres-Pérez is currently supported by the NASA Postdoctoral Program at the NASA Ames Research Center in California.
References
- Holden, H.; LeDrew, E. Hyperspectral identification of coral reef features. Int. J. Remote Sens. 1999, 20, 2545–2563. [Google Scholar]
- Lubin, D.; Li, W.; Dustan, P.; Mazel, C.H.; Stamnes, K. Spectral signatures of coral reefs: features from space. Remote Sens. Environ. 2001, 75, 127–137. [Google Scholar]
- Hedley, J.D.; Mumby, P.J. Biological and remote sensing perspectives of pigmentation in coral reef organisms. Adv. Mar. Biol. 2002, 43, 277–317. [Google Scholar]
- Hochberg, E.J.; Atkinson, M.J. Capabilities of remote sensors to classify coral, algae and sand as pure and mixed spectra. Remote Sens. Environ. 2003, 85, 174–189. [Google Scholar]
- Hochberg, E.J.; Atkinson, M.J.; Apprill, A.; Andréfouët, S. Spectral reflectance of coral. Coral Reefs 2004, 23, 84–95. [Google Scholar]
- Hedley, J.D.; Mumby, P.J.; Joyce, K.E.; Phinn, S.R. Spectral unmixing of coral reef benthos under ideal conditions. Coral Reefs 2004, 23, 60–73. [Google Scholar]
- Hedley, J.D.; Roelfsema, C.M.; Phinn, S.R.; Mumby, P.J. Environmental and sensor limitations in optical remote sensing of coral reefs: implications for monitoring and sensor design. Remote Sens. 2012, 4, 271–302. [Google Scholar]
- Hochberg, E.J.; Atkinson, M.J.; Andréfouët, S. Spectral reflectance of coral reef bottom-types worldwide and implications for coral reef remote sensing. Remote Sen. Environ. 2003, 85, 159–173. [Google Scholar]
- Johansen, J.E.; Svec, W.A.; Liaaen-Jensen, S. Carotenes of the Dinophyceae. Phytochemistry 1974, 13, 2261–2271. [Google Scholar]
- Gil-Turnes, S.; Corredor, J. Studies of Photosynthetic Pigments of Zooxanthellae in Caribbean Hermatypic Corals. Proceedings of the 4th International Coral Reef Symposium, Manila, Philippines, 18–22 May 1981; pp. 51–54.
- Jeffrey, S.W.; Mantoura, R.F.C.; Wright, S.W. Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; UNESCO Publishing; Paris, France, 1997. [Google Scholar]
- Rowan, R.; Knowlton, N. Intraspecific diversity and ecological zonation in coral-algal symbiosis. Proc. Natl Acad. Sci. USA 1995, 92, 2850–2853. [Google Scholar]
- La Jeunesse, T.C.; Loh, W.K.W.; van Woesick, R.; Hoegh-Guldberg, O.; Schmidt, G.W.; Fitt, W.K. Low symbiont diversity in southern Great Barrier Reef corals, relative to those of the Caribbean. Limnol. Oceanogr. 2003, 48, 2046–2054. [Google Scholar]
- Baker, A.C.; Rowan, R. Diversity of Symbiotic Dinoflagellates (Zooxanthellae) in Scleractinian Corals of the Caribbean and the Eastern Pacific. Proceedings of the 8th International Coral Reef Symposium, Panama, 24–29 June 1997; pp. 1301–1306.
- Baker, A.C.; Rowan, R.; Knowlton, N. Symbiosis Ecology of two Caribbean Acroporid Corals. Proceedings of the 8th International Coral Reef Symposium, Panama, 24–29 June 1997; pp. 1295–1300.
- LaJeunesse, T.C. Diversity and community structure of symbiotic dinoflagellates from Caribbean coral reefs. Mar. Biol. 2002, 141, 378–400. [Google Scholar]
- Banaszak, A.T.; Iglesias-Prieto, R.; Trench, R.K. Scrippsiella velellae sp. nov. (Peridiniales) and Gloedinium viscum sp. nov. (Phytodiniales), dinoflagellate symbionts of two hydrozoans (Cnidaria). J. Phycol. 1993, 29, 517–528. [Google Scholar]
- Blank, R.J.; Trench, R.K. Nomenclature of endosymbiotic dinoflagellates. Taxon 1986, 35, 286–294. [Google Scholar]
- Rowan, R. Diversity and ecology of zooxanthellae on coral reefs. J. Phycol. 1998, 34, 407–417. [Google Scholar]
- Rowan, R.; Powers, D.A. Molecular genetic identification of symbiotic dinoflagellates (zooxanthellae). Mar. Ecol. Prog. Ser. 1991, 71, 65–73. [Google Scholar]
- Bidigare, R.R.; Morrow, J.H.; Kiefer, D.A. Derivative analysis of spectral absorption by photosynthetic pigments in the Western Sargasso Sea. J. Mar. Res. 1989, 47, 323–341. [Google Scholar]
- Andréfouët, S.; Payri, C.; Hochberg, E.J.; Che, L.M.; Atkinson, M.J. Airbornes hyperspectral detection of microbial mat pigmentation in Rangiroa atoll (French Polynesia). Limnol. Oceanogr. 2003, 48, 426–430. [Google Scholar]
- Polerecky, L.; Bissett, A.; Al-Najjar, M.; Faerber, P.; Osmers, H.; Suci, P.A.; Stoodley, P.; de Beer, D. Modular spectral imaging system for discrimination of pigments in cells and microbial communities. Appl. Environ. Microbiol. 2009, 75, 758–771. [Google Scholar]
- Savitzky, A.; Golay, M.J.E. Smoothing and differentiation of data by simplified least squares procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar]
- Butler, W.L.; Hopkins, D.W. Higher derivative analysis of complex absorption spectra. Photochem. Photobiol 1970, 12, 439–450. [Google Scholar]
- Tsai, F.; Philpot, W. Derivative analysis of hyperspectral data. Remote Sens. Environ. 1998, 66, 41–51. [Google Scholar]
- Smith, C.M.; Alberte, R.S. Characterization of in vivo absorption features of chlorophyte, phaeophyte and rhodophyte algal species. Mar. Biol. 1994, 118, 511–521. [Google Scholar]
- Hochberg, E.J.; Apprill, A.; Atkinson, M.J.; Bidigare, R.R. Bio-optical modeling of photosynthetic pigments in corals. Coral Reefs 2006, 25, 99–109. [Google Scholar]
- Torres-Pérez, J.L.; Armstrong, R.A. Effects of UV radiation on the growth, photosynthetic and photoprotective components, and reproduction of the Caribbean shallow-water coral Porites furcata. Coral Reefs 2012, 31, 1077–1091. [Google Scholar]
- Torres, J.L.; Armstrong, R.A.; Corredor, J.E.; Gilbes, F. Physiological responses of Acropora cervicornis to increased solar irradiance. Photochem. Photobiol. 2007, 83, 839–851. [Google Scholar]
- Shick, J.M.; Lesser, M.P.; Dunlap, W.C.; Stochaj, W.R.; Chalker, B.E.; Wu Won, J. Depth-dependent responses to solar ultraviolet radiation and oxidative stress in the zooxanthellate coral Acropora microphthalma. Mar. Biol. 1995, 122, 41–51. [Google Scholar]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants and natural phytoplankton. Biochem. Physiol. Pfl. 1975, 167, 191–194. [Google Scholar]
- Chalker, B.E.; Dunlap, W.C. Extraction and Quantification of Endosymbiotic Algal Pigments from Reef-Building Corals. Proceedings of the 4th International Coral Reef Symposium, Manila, Philippines, 18–22 May 1981; pp. 45–50.
- Wright, S.W.; Shearer, J.D. Rapid extraction and high-performance liquid chromatography of chlorophylls and carotenes from marine phytoplankton. J. Chromatogr. 1984, 294, 281–295. [Google Scholar]
- Wright, S.W.; Jeffrey, S.W.; Mantoura, R.F.C.; Llewellyn, C.A.; Bjþrnland, T.; Repeta, D.; Welschmeyer, N. Improved HPLC method for the analysis of chlorophylls and carotenes from marine phytoplankton. Mar. Ecol. Prog. Ser. 1991, 77, 183–196. [Google Scholar]
- Bidigare, R.R. Analysis of Algal Chlorophylls and Carotenes. In Marine Particles: Analysis and Characterization; Hurd, D.C., Spencer, D.W., Eds.; American Geophysical Union: Washington DC, USA, 1991; pp. 119–123. [Google Scholar]
- Bidigare, R.R.; van Heukelem, L.; Trees, C.C. Analysis of Algal Pigments by High Performance Liquid Chromatography. In Culturing Methods and Growth Measurements; Andersen, R.A., Ed.; Academic Press: New York, NY, USA, 2005; pp. 327–345. [Google Scholar]
- Egeland, E.S.; Garrido, J.L.; Clementson, L.; Andresen, K.; Thomas, C.S.; Zapata, M.; Airs, R.; Llewellyn, C.A.; Newman, G.L.; Rodríguez, F.; et al. Data Sheets Aiding Identification of Phytoplankton Carotenes and Chlorophylls. In Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Roy, S., Llewelyn, C.A., Egeland, E.S., Johnsen, G., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 665–811. [Google Scholar]
- Jeffrey, S.W.; Wright, S.W.; Zapata, M. Microalgal Classes and their Signature Pigments. In Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Roy, S., Llewelyn, C.A., Egeland, E.S., Johnsen, G., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 3–77. [Google Scholar]
- Marsh, J.A. Primary productivity of reef building calcareous red algae. Ecology. 1970, 51, 255–263. [Google Scholar]
- Fung, T.; LeDrew, E. Application of principal components analysis to change detection. Photogramm. Eng. Rem. S. 1987, 53, 1649–1658. [Google Scholar]
- Apprill, A.; Bidigare, R.R.; Gates, R.D. Visibly healthy corals exhibit variable pigment concentrations and symbiont phenotypes. Coral Reefs 2007, 26, 387–397. [Google Scholar]
- Warner, M.E.; Berry-Lowe, S. Differential xanthophyll cycling and photochemical activity in symbiotic dinoflagellates in multiple locations of three species of Caribbean coral. J. Exp. Mar. Biol. Ecol. 2006, 339, 86–95. [Google Scholar]
- Ugalde, J.A.; Chang, B.S.; Matz, M.V. Evolution of coral pigments recreated. Science 2004, 305, 1433. [Google Scholar]
- Corredor, J.E.; Bruckner, A.W.; Muszynski, F.Z.; Armstrong, R.A.; García, R.; Morell, J.M. UV-absorbing compounds in three species of Caribbean zooxanthellate corals: depth distribution and spectral response. Bull. Mar. Sci. 2000, 67, 821–830. [Google Scholar]
- Enríquez, S.; Méndez, E.R.; Iglesias-Prieto, R. Multiple scattering on coral skeletons enhances light absorption by symbiotic algae. Limnol. Oceanogr. 2005, 50, 1025–1032. [Google Scholar]
- Mazel, C.H. Spectral measurements of fluorescence emission in Caribbean cnidarians. Mar. Ecol. Prog. Ser. 1995, 120, 185–191. [Google Scholar]
- Mazel, C.H.; Lesser, M.P.; Gorbunov, M.Y.; Barry, T.M.; Farell, J.H.; Wyman, K.D.; Falkowski, P.G. Green-fluorescent proteins in Caribbean corals. Limnol. Oceanogr. 2003, 48, 402–411. [Google Scholar]
- Oswald, F.; Schmitt, F.; Leutenegger, A.; Ivanchenko, S.; D’Angelo, C.; Salih, A.; Maslakova, S.; Bulina, M.; Schirmbeck, R.; Nienhaus, G.U.; et al. Contributions of host and symbiont pigments to the coloration of reef corals. FEBS J. 2007, 274, 1102–1109. [Google Scholar]
- D’Angelo, C.; Smith, E.G.; Oswald, F.; Burt, J.; Tchernov, D.; Wiedenmann, J. Locally accelerated growth is part of the innate immune response and repair mechanisms in reef-building corals as detected by green fluorescent protein (GFP)-like pigments. Coral Reefs 2012, 31, 1045–1056. [Google Scholar]
Figure 1.
Collection site in La Parguera, southwestern Puerto Rico. San Cristóbal Reef is enlarged in the image. The circles indicate the areas were each species was collected: orange—A. cervicornis and black—P. porites.
Figure 1.
Collection site in La Parguera, southwestern Puerto Rico. San Cristóbal Reef is enlarged in the image. The circles indicate the areas were each species was collected: orange—A. cervicornis and black—P. porites.
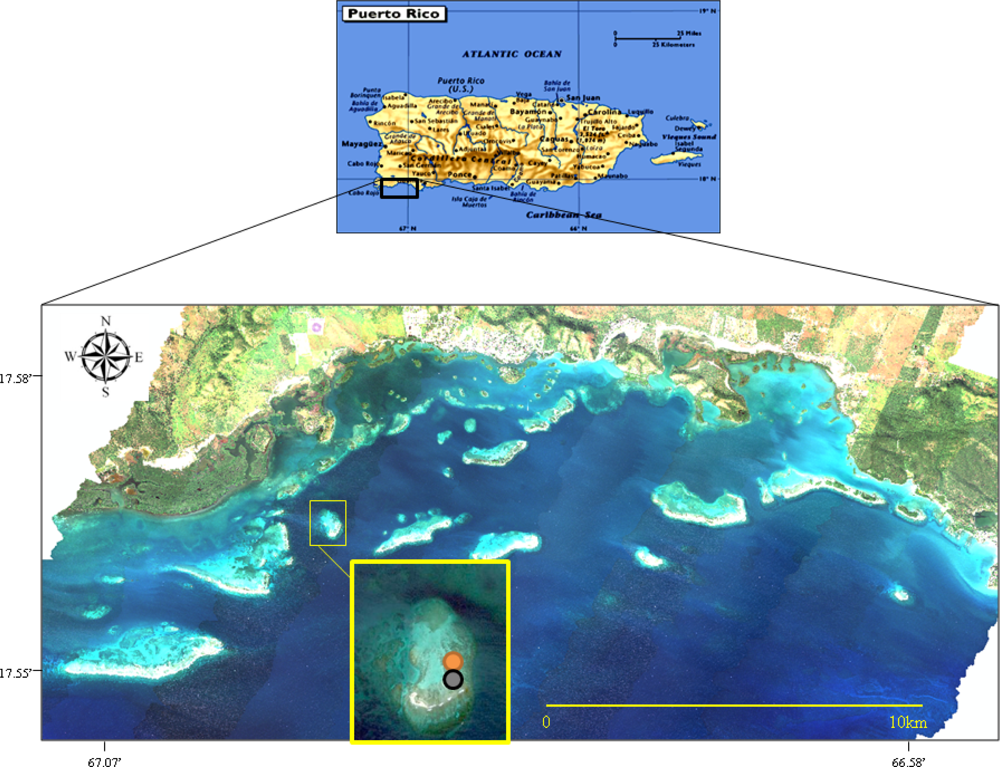
Figure 2.
Representative chromatograms of A. cervicornis and P. porites showing the major pigments resolved. Note the differences in pigments that elute between 13 to 16 minutes. Only the most prominent peaks are identified in the graph. For the complete list of all the pigments found in both species, see Table 1 above. Scale bars in the photos = 30 cm in A. cervicornis and 20 cm in P. porites.
Figure 2.
Representative chromatograms of A. cervicornis and P. porites showing the major pigments resolved. Note the differences in pigments that elute between 13 to 16 minutes. Only the most prominent peaks are identified in the graph. For the complete list of all the pigments found in both species, see Table 1 above. Scale bars in the photos = 30 cm in A. cervicornis and 20 cm in P. porites.
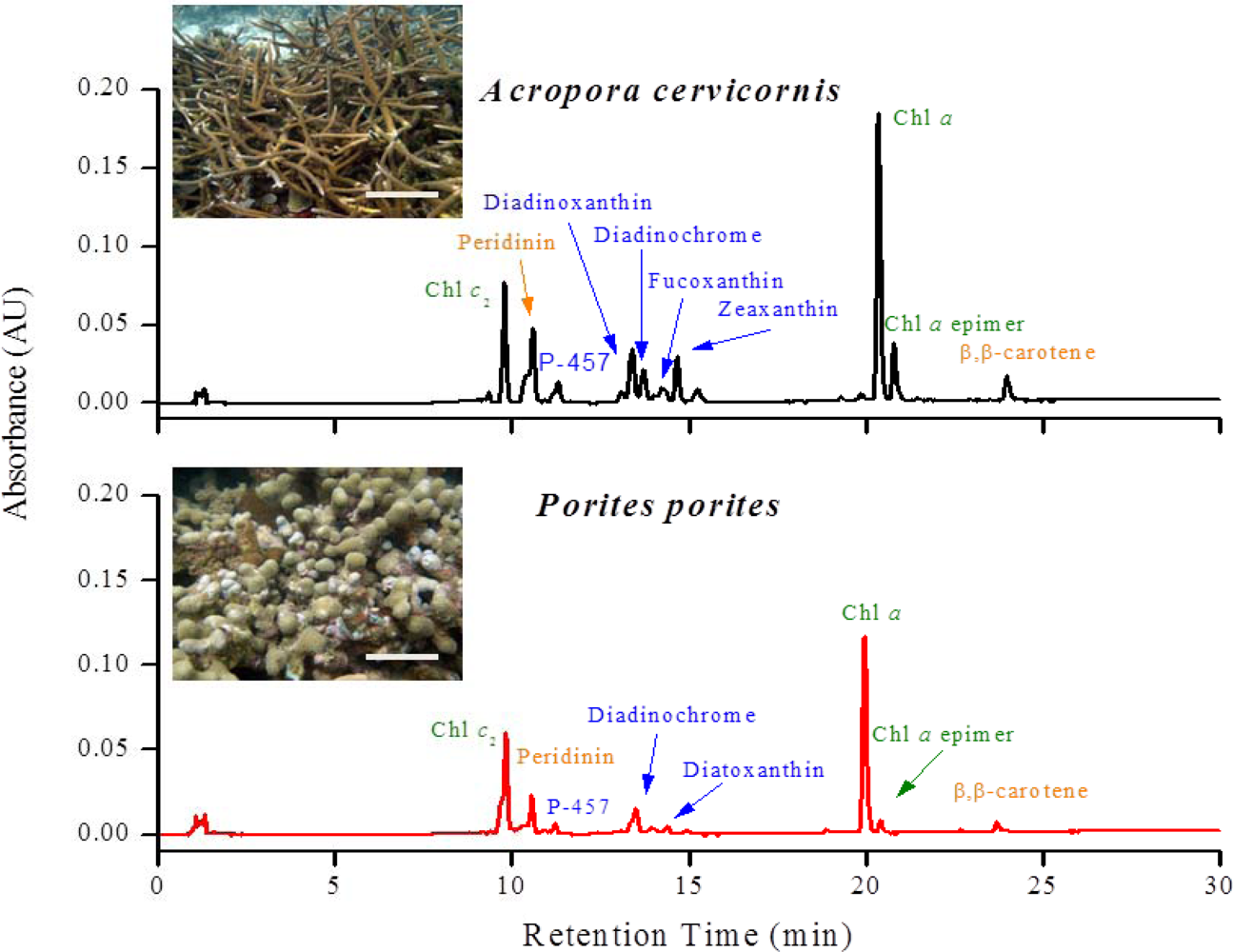
Figure 3.
General composition of photosynthetic pigments in A. cervicornis and P. porites. Error bars represent ± 1 SD.
Figure 3.
General composition of photosynthetic pigments in A. cervicornis and P. porites. Error bars represent ± 1 SD.

Figure 4.
Reflectance curves for A. cervicornis and P. porites (n = 5) with their respective 2nd derivative. The numbers in parenthesis in the derivative graphs represent peaks at specific wavelengths (nm): (1) 427, (2) 442, (3) 495, (4), 514, (5) 522, (6) 530, (7) 563, (8) 589, (9) 618, (10) 634, (11) 665 and (12) 685. The heights of nine out of twelve peaks were significantly different between species (peaks at 514, 522 and 563 nm were not significantly different at α ≤ 0.05).
Figure 4.
Reflectance curves for A. cervicornis and P. porites (n = 5) with their respective 2nd derivative. The numbers in parenthesis in the derivative graphs represent peaks at specific wavelengths (nm): (1) 427, (2) 442, (3) 495, (4), 514, (5) 522, (6) 530, (7) 563, (8) 589, (9) 618, (10) 634, (11) 665 and (12) 685. The heights of nine out of twelve peaks were significantly different between species (peaks at 514, 522 and 563 nm were not significantly different at α ≤ 0.05).
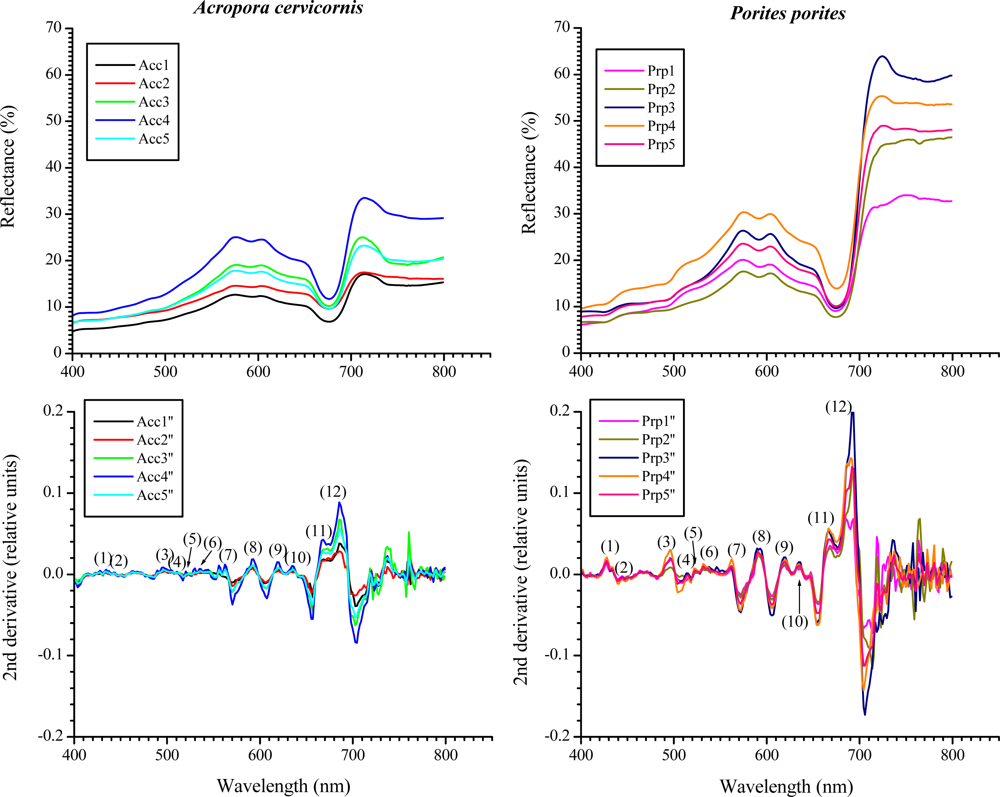
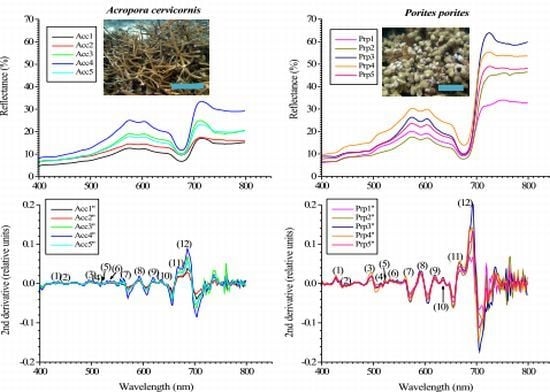
Figure 5.
Reflectance curves as measured from three locations of a branch (individual) of each studied species. The 2nd derivatives of each curve within a sample of each species show minimal intra-specimen variation of the spectral features independently of the difference in magnitudes in reflectance along a single branch. The numbers in parenthesis in the derivative graphs represent peaks at specific wavelengths (nm): (1) 427, (2) 442, (3) 495, (4), 514, (5) 522, (6) 530, (7) 563, (8) 589, (9) 618, (10) 634, (11) 665 and (12) 685.
Figure 5.
Reflectance curves as measured from three locations of a branch (individual) of each studied species. The 2nd derivatives of each curve within a sample of each species show minimal intra-specimen variation of the spectral features independently of the difference in magnitudes in reflectance along a single branch. The numbers in parenthesis in the derivative graphs represent peaks at specific wavelengths (nm): (1) 427, (2) 442, (3) 495, (4), 514, (5) 522, (6) 530, (7) 563, (8) 589, (9) 618, (10) 634, (11) 665 and (12) 685.
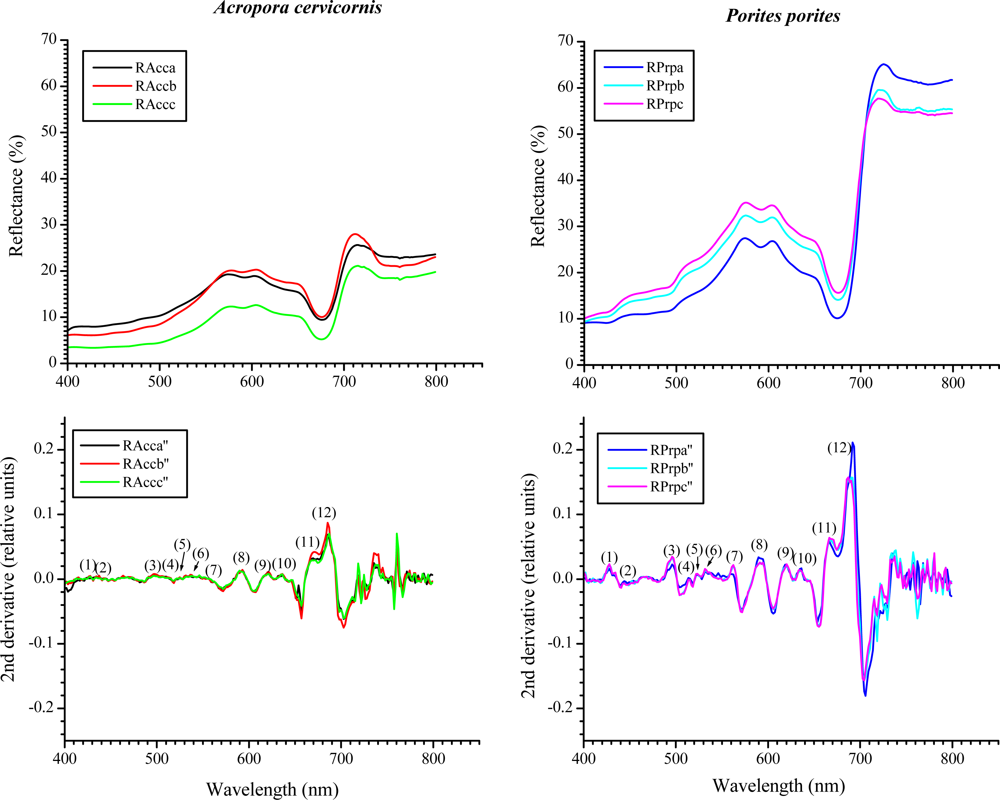
Figure 6.
Average absorption spectra (n = 5) for A. cervicornis and P. porites with their respective 2nd derivative. The numbers in parenthesis in the derivative graphs represent peaks at specific wavelengths (nm): (1) 424, (2) 452, (3) 468, (4), 490, (5) 507, (6) 518, (7) 534, (8) 551, (9) 561, (10) 575, (11) 586, (12) 604, (13) 616, (14) 632, (15) 649, (16) 658 (A. cervicornis) and 661 (P. porites) and (17) 680. Note the hypsochromic shift of absorption peaks (numbers in bold) that are also present in the coral’s reflectance curves shown in Figures 4 and 5.
Figure 6.
Average absorption spectra (n = 5) for A. cervicornis and P. porites with their respective 2nd derivative. The numbers in parenthesis in the derivative graphs represent peaks at specific wavelengths (nm): (1) 424, (2) 452, (3) 468, (4), 490, (5) 507, (6) 518, (7) 534, (8) 551, (9) 561, (10) 575, (11) 586, (12) 604, (13) 616, (14) 632, (15) 649, (16) 658 (A. cervicornis) and 661 (P. porites) and (17) 680. Note the hypsochromic shift of absorption peaks (numbers in bold) that are also present in the coral’s reflectance curves shown in Figures 4 and 5.
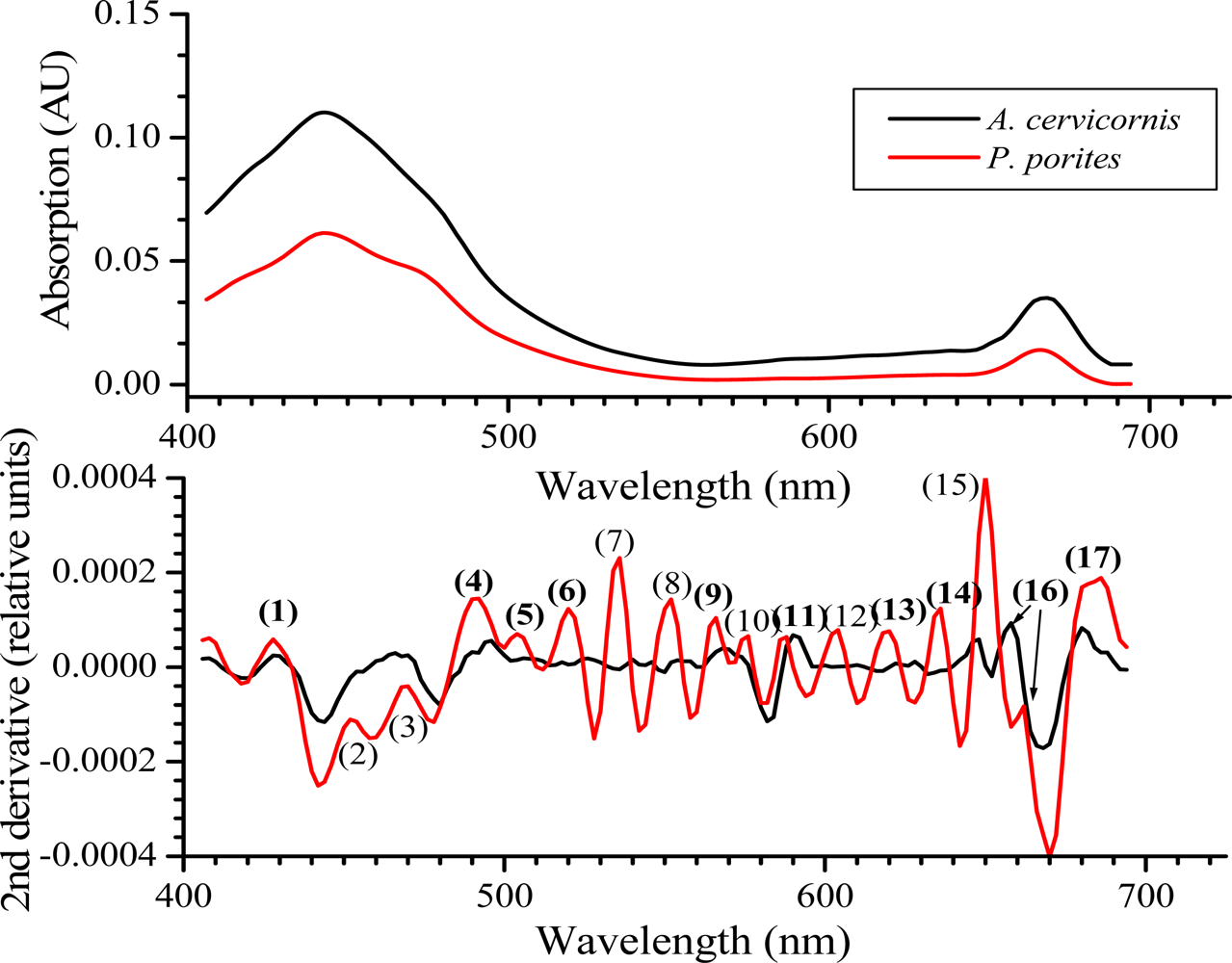
Figure 7.
Relationship between the area under the reflectance curve and the total pigment concentration in A. cervicornis and P. porites. The x-axis represents an integration of the reflectance curve from 400 to 700 nm.
Figure 7.
Relationship between the area under the reflectance curve and the total pigment concentration in A. cervicornis and P. porites. The x-axis represents an integration of the reflectance curve from 400 to 700 nm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of pigments found in the studied species. The concentrations are expressed as μg/cm2 ± 1SD based on 5 colonies sampled per species. Absorption peaks reported here may vary to others reported elsewhere depending on the solvent used and may be lower than actual in vivo absorption peaks as result of hypsochromic shift during the extraction process.
| Pigments/Compounds | Acropora cervicornis | Porites porites | Abs. Peaks* | ||||
|---|---|---|---|---|---|---|---|
| Presence | [Pigm] | % | Presence | [Pigm] | % | ||
| MV-Chl c3 | + | 0.38 ± 0.13 | 0.9 | − | -- | -- | 447, 626 |
| Mg-DVP | + | 0.60 ± 0.26 | 1.4 | − | -- | -- | 437, 624 |
| Chl c2 | + | 2.82 ± 0.11 | 6.5 | + | 1.64 ± 0.70 | 9.5 | 452, 635 |
| Chl a allo | + | 0.29 ± 0.03 | 0.7 | − | -- | -- | 432, 665 |
| Chl a | + | 13.60 ± 1.92 | 31.4 | + | 9.46 ± 0.86 | 58.7 | 432, 665 |
| Chl a epi | + | 2.79 ± 0.27 | 6.4 | + | 0.41 ± 0.04 | 2.5 | 432, 665 |
| Per | + | 7.34 ± 0.21 | 16.9 | + | 2.38 ± 0.14 | 14.4 | 475 |
| β,β–car | + | 0.61 ± 0.05 | 1.4 | + | 0.28 ± 0.05 | 1.7 | 449, 475 |
| Nc | + | 1.72 ± 0.20 | 4.0 | + | 0.04 ± 0.01 | 0.3 | 424, 451 |
| P-457 | + | 1.43 ± 0.50 | 3.3 | + | 0.31 ± 0.18 | 2.0 | 457 |
| Dn | + | 0.22 ± 0.04 | 0.5 | − | -- | -- | 438, 467 |
| Dd | + | 6.50 ± 1.15 | 15.0 | − | -- | -- | 445, 476 |
| Dc I | + | 0.68 ± 0.06 | 1.6 | + | 0.81 ± 0.54 | 4.9 | 430, 457 |
| Dc II | + | 0.73 ± 0.12 | 1.7 | + | 0.25 ± 0.02 | 1.5 | 430, 457 |
| Fc | + | 0.69 ± 0.06 | 1.6 | − | -- | -- | 446, 475 |
| Zea | + | 1.34 ± 0.09 | 3.1 | − | -- | -- | 449, 475 |
| An | + | 0.50 ± 0.15 | 1.2 | − | -- | -- | 444, 472 |
| Lu** | + | 0.34 | 0.8 | − | -- | -- | 443, 470 |
| Gy** | + | 0.45 | 1.0 | − | -- | -- | 442, 470 |
| Dt | − | -- | -- | + | 0.23 ± 0.03 | 1.5 | 452, 478 |
Abbreviations: Monovinyl Chlorophyll c3 (MV-Chl c3), Magnesium 2,4-divinyl pheoporphyrin a monomethyl ester (MgDVP), Chlorophyll a (Chl a), Chlorophyll c2 (Chl c2), Peridinin (Per), Diadinoxanthin (Dd), Diatoxanthin (Dt), Dinoxanthin (Dn), Diadinochromes I and II (DcI and DcII, respectively), Neochrome (Nc), Fucoxanthin (Fc), Zeaxanthin (Zea), Antheraxanthin (An), Lutein (Lu), Gyroxanthin dodecanoate ethanoate (Gy), β,β-carotene (β,β-car).
*Pigments were extracted with a methanol:tetrahydrofuran (80:20, v/v) solution.
**found in only one sample.
Table 2.
Principal Components Analysis (PCA) for variations among the main pigments (or pigment groups) of A. cervicornis and P. porites based on 5 colonies sampled per species. Percentages of pigment groups are relative to each other. Eigenvectors in bold represent the highest values for PC1 and PC2.
| PC 1 | PC 2 | PC 3 | |
|---|---|---|---|
| Eigenvalues | 267 | 66.3 | 38.6 |
| % variation | 69.4 | 17.3 | 10.0 |
| Variables analyzed: | Eigenvectors | ||
| [Chl a] | −0.188 | −0.412 | −0.028 |
| [Chl c2] | −0.025 | −0.003 | −0.043 |
| [Per] | −0.027 | 0.057 | −0.037 |
| % Chl a | 0.547 | −0.540 | 0.324 |
| % Chl c2 | 0.189 | 0.088 | −0.326 |
| % Per | −0.043 | −0.083 | −0.730 |
| [total chlorophylls] | −0.442 | −0.684 | −0.256 |
| [total carotenes] | −0.020 | 0.063 | −0.034 |
| [total xanthophylls] | −0.169 | −0.057 | −0.002 |
| % chlorophylls | 0.514 | −0.120 | −0.333 |
| % carotenes | −0.201 | −0.051 | 0.068 |
| % xanthophylls | −0.313 | 0.171 | 0.265 |
Share and Cite
MDPI and ACS Style
Torres-Pérez, J.L.; Guild, L.S.; Armstrong, R.A. Hyperspectral Distinction of Two Caribbean Shallow-Water Corals Based on Their Pigments and Corresponding Reflectance. Remote Sens. 2012, 4, 3813-3832. https://doi.org/10.3390/rs4123813
AMA Style
Torres-Pérez JL, Guild LS, Armstrong RA. Hyperspectral Distinction of Two Caribbean Shallow-Water Corals Based on Their Pigments and Corresponding Reflectance. Remote Sensing. 2012; 4(12):3813-3832. https://doi.org/10.3390/rs4123813
Chicago/Turabian StyleTorres-Pérez, Juan L., Liane S. Guild, and Roy A. Armstrong. 2012. "Hyperspectral Distinction of Two Caribbean Shallow-Water Corals Based on Their Pigments and Corresponding Reflectance" Remote Sensing 4, no. 12: 3813-3832. https://doi.org/10.3390/rs4123813