We use the mean NDVI over the Photosynthetically Active Period (
NPAP) and the mean temperature from May to September (
TMS) to represent the inter-annual vegetation dynamics and the corresponding temperature changes [
5,
33] The tight coupling between temperature and vegetation growth can be found in the linear relationships between
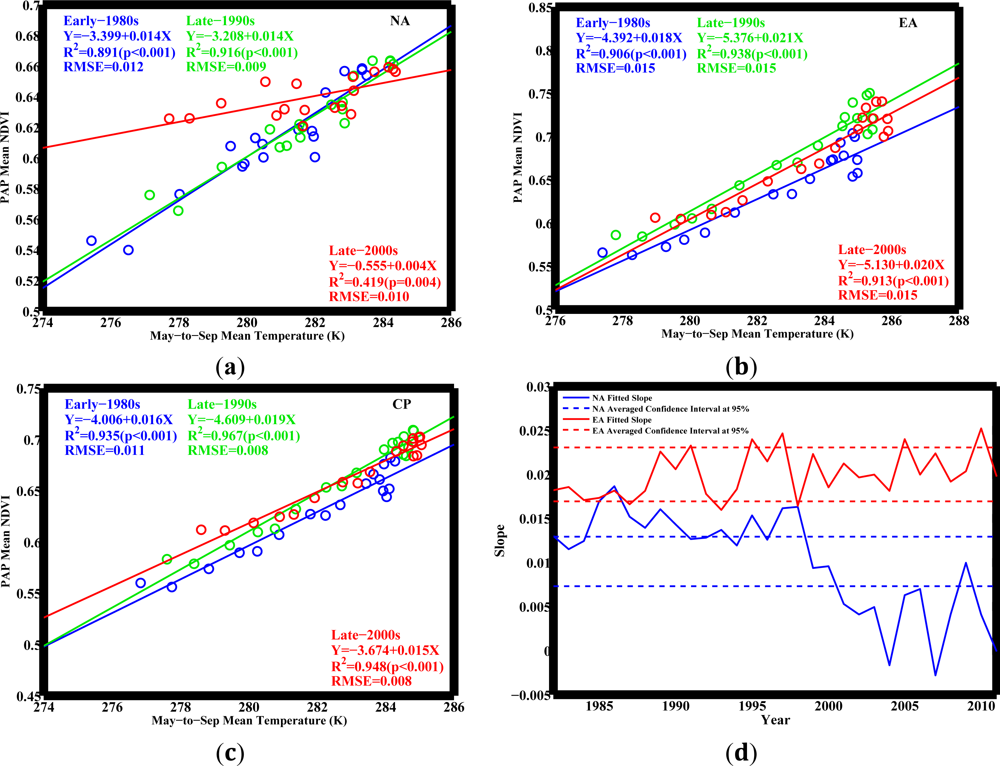
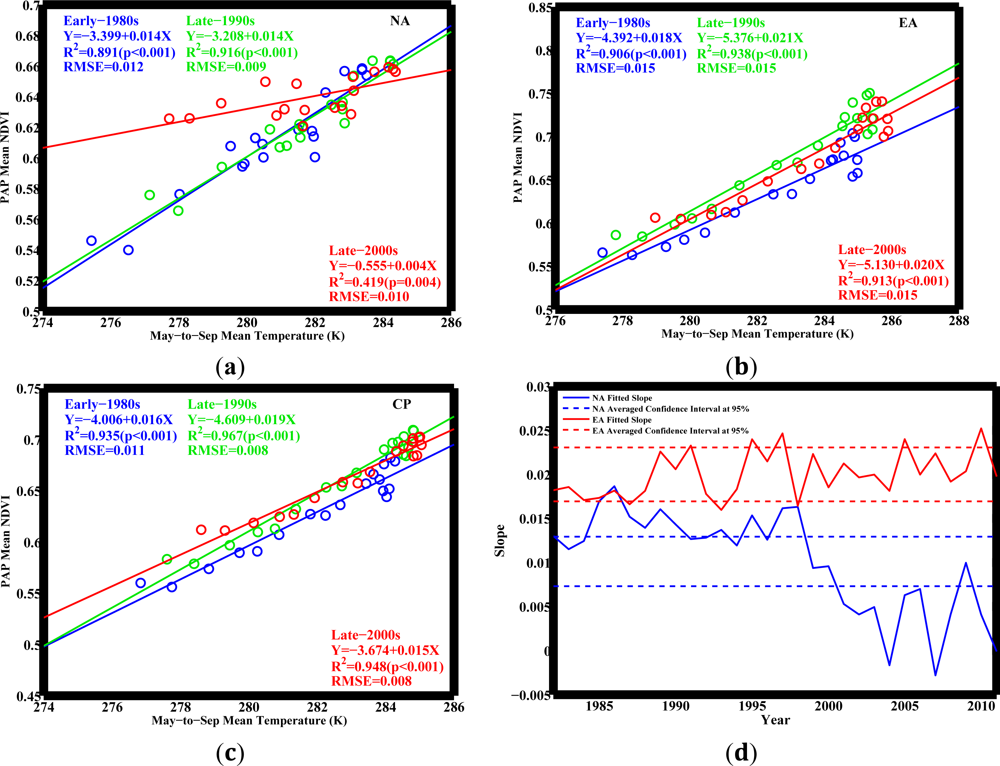
NPAP and
TMS across latitudes in both NA and EA for the past 30 years (
Figure 3). The circumpolar pattern follows EA, as the vegetated area in EA is 50% greater than that in NA. The consistent
NPAP−
TMS relationships across both NA and EA (and hence in CP) during the early-1980s and late-1990s indicate stable ecosystem response to temperature.
However, this relationship changes during the late-2000s in NA in a manner that the response of northerly vegetation to temperature no longer resembles that of southerly vegetation observed during early-1980s (
Figure 3(a)). This weakened relationship in NA has two implications: (1) The slope in the late-2000s (0.004) is 2/3 times smaller than the slope in the early-1980s (0.014). If future changes of temperature and vegetation growth follow the line of the late-2000s in NA, similar to the changes in the late-1990s following the line established in the early-1980s, the same amount of warming would cause 2/3 less greening compared to the changes in the early-1980s; (2) Taking a close look at the latitudinal points, these deviations are found mostly in the Arctic region of NA, where temperature is relatively low, but vegetation grows abnormally fast with rising temperatures in the period between the late-1990s and the late-2000s. Vegetation greening in this period is not subject to temperature change as before. Such deviations hint at novel vegetation responses, implying that temperature may no longer be the dominant factor as before governing vegetation growth in this region.
3.1. Spatial Analysis of Long-Term Trend
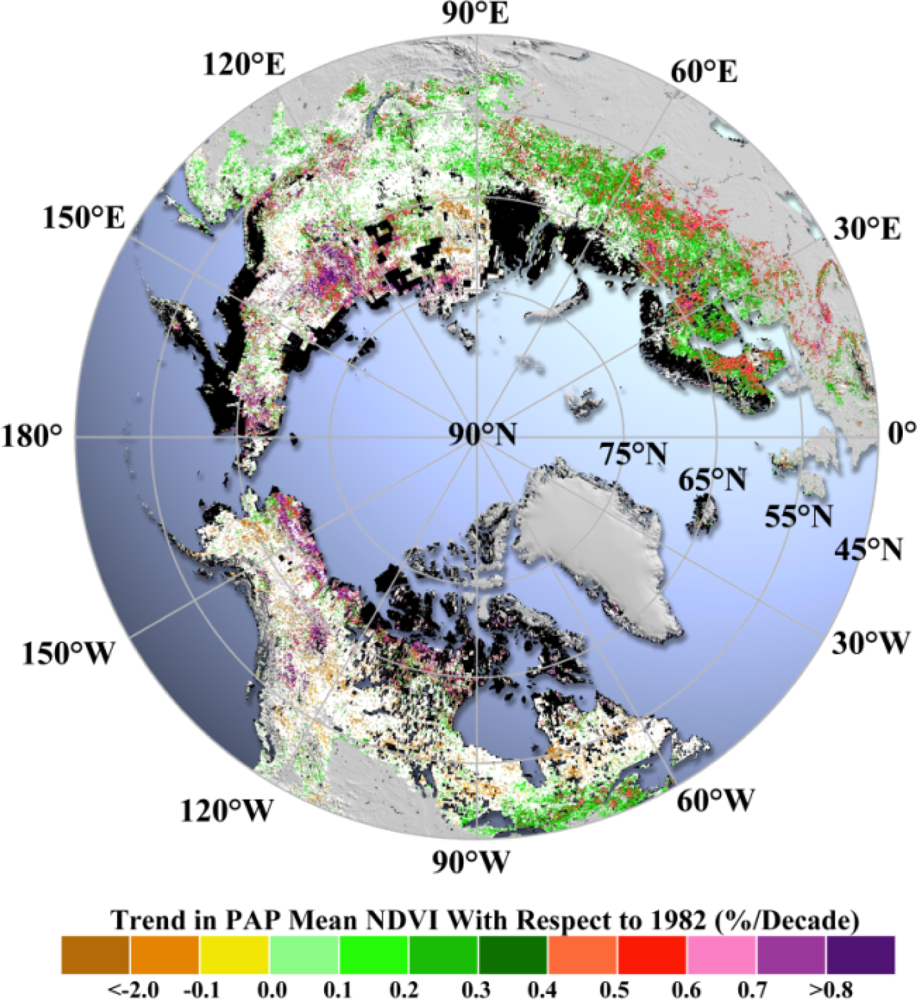
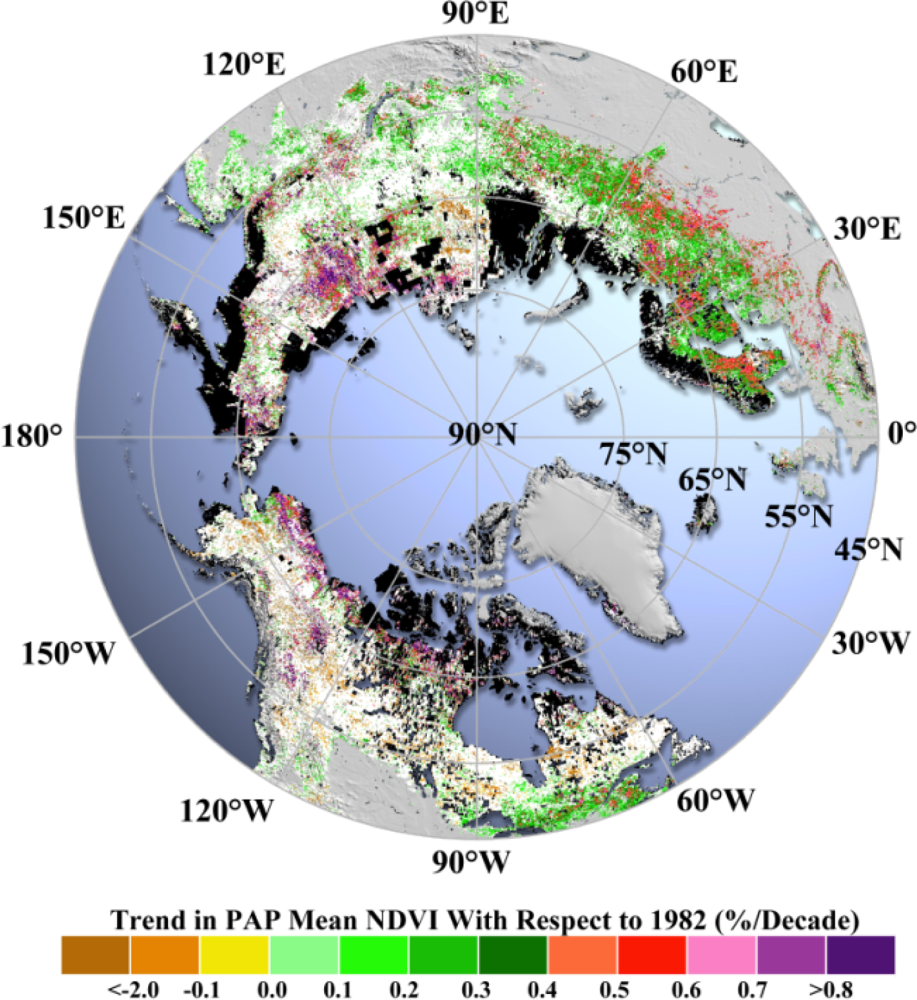
Pixel-wise application of the robust trend estimation model (
cf. Section 2.2.2) shows that increases in
NPAP for both NA and EA (
Figure 4) prevail during the periods from 1982 to 2011. Spatial patterns are also assessed to provide a general picture of the difference in vegetation growth between NA and EA.
EA shows significant greening,
i.e., about 45 times more greening (increase in
NPAP) area than the browning (decrease in
NPAP) area. By contrast, the greening area in NA is about two times larger than the browning area. Comparing the greening areas between EA and NA alone, the area in EA is 2.6 times larger than that in NA. In particular, the fraction of greening in the Arctic region of EA is similar to that of NA; and the fraction of greening in the Boreal region of EA is two times larger than that in NA (
Table 2). Within the Boreal region, forests and other natural vegetation in NA have similar fractions of greening (14%–16%). However, more than 70% of the boreal forests in EA show greening compared to 44% of greening for other natural vegetation in EA (
Table 3).
As for the browning trend, vegetation in NA has a larger area of browning compared to EA, especially in the Boreal region (
Figure 4). Browning area in NA is 4.2 times larger than that in EA. In particular, the fraction of browning in the Arctic region of EA is still comparable to that in NA. On the contrary, the fraction of browning in the Boreal region of NA is 20 times larger than that in EA. Within the NA region, the majority of browning area is located in the Boreal zone. The fraction of browning in the Boreal region of NA is two times larger than that in the Arctic of NA (
Table 2). Within the Boreal region, the fraction of browning area of forests is comparable to that of other natural vegetation (8.2%
vs. 7.4%) in NA, while in EA almost no forests show browning (0.03%) compared to other natural vegetation (0.41%), even if both fractions are small (
Table 3).
The observed spatial changes (
Figure 4) are consistent with earlier studies reporting continued Arctic greening in NA and EA [
4,
5,
8–
11]. However, the previously reported Boreal browning in EA [
9] is not found here, possibly due to the improved data quality in the northerly lands of the new AVHRR NDVI3g product [
27]. However, the contrast between Boreal NA and EA, with more significant browning in NA, cannot be attributed to altered vegetation response to temperature, given that spatial patterns of vegetation and temperature trends are consistent [
14,
15,
21,
42]. Our analysis of temperature trends (
Table 4) shows more extensive cooling in Boreal NA compared to Boreal EA, about seven times higher, which could plausibly account for the browning trends found in Boreal NA.
On the other hand, we also find more extensive cooling in Arctic NA compared to Arctic EA—about over seven times higher—but with comparable vegetation changes. This implies more greening under conditions of relatively less warming in Arctic NA. Therefore, the observed loss of vegetation sensitivity to temperature in NA can be partly attributed to the spatial distribution of vegetation and temperature trends. If vegetation response to temperature has changed in the Arctic of NA, one would suspect that other variables such as precipitation, insolation and CO
2 concentration driving the changes of vegetation [
7,
24,
43] in the Arctic-Boreal region could play a bigger role.
We also analyzed precipitation trends (
Table 4), and found similar wetting/drying in NA and EA. For the Boreal region, the observed browning in NA could possibly be attributed to the more extensive cooling in the same region, when taking temperature as the dominant climatic factor, regardless of the similarities in precipitation. Based on the same reasoning, is it true that precipitation is becoming more important in the Arctic, as NDVI changes follow the precipitation spatial fractions (similar changes in both NA and EA), instead of temperature? It is difficult to judge merely based on the spatial patterns, as both warming and wetting are spatially more extensive than the observed vegetation changes in either NA or EA. Again,
Figure 3 gives a hint that vegetation tends to be in line with the climate change, and warming causes vegetation to resemble the southerly species, which is in favor of that climate condition. Therefore, it is necessary to investigate the spatial pattern of climate itself and how it shifts over time.
3.2. Analysis of Latitudinal Profiles of Temperature and Precipitation
Different spatial patterns of climate can result in unique distributions of ecosystems across space. For example, there are more tundra regions in the North America even to south of 65°N [
32], and most deciduous needleleaf forests are located in Eurasia [
31]. Under the rapid climate changes in the recent decades, vegetation must keep pace with the shifting climate for survival [
44]. In particular, temperature is most essential, as vegetation responses are expected to track the rate of isotherm migration over space [
41,
45,
46]. Ground surveys also show evidence that vegetation appears to have an upward shift in mountainous areas [
47,
48] and a northward shift in tundra areas [
16,
18,
19,
49,
50] responding to the temperature changes.
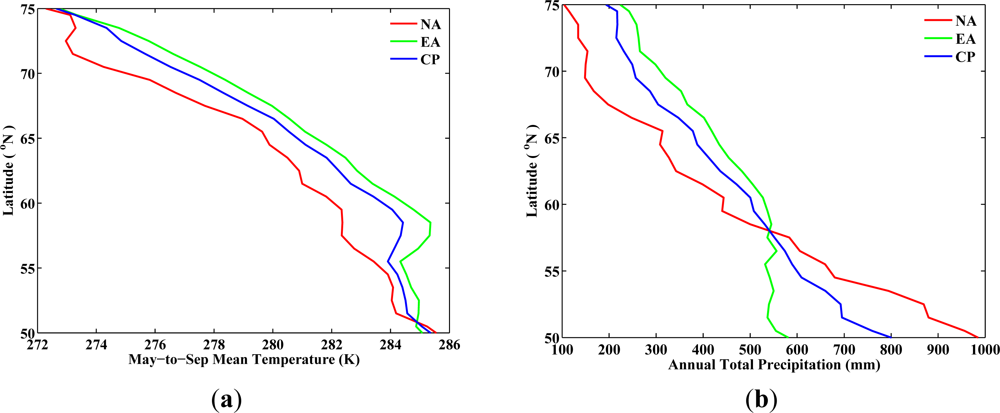
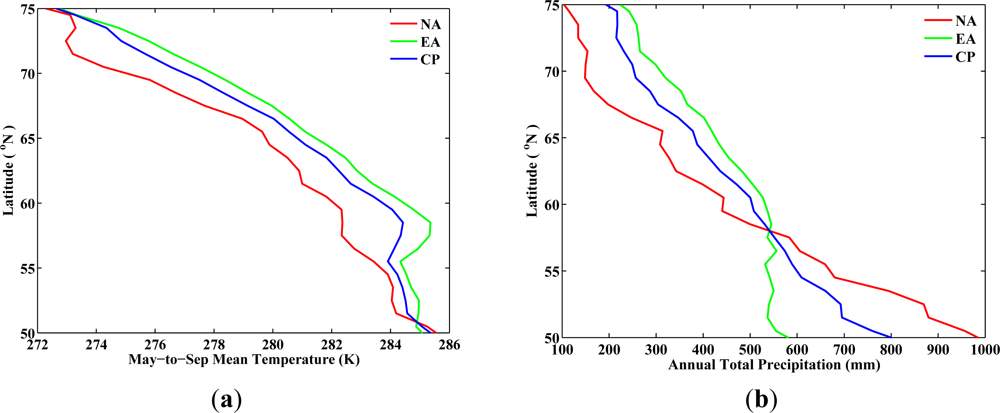
From the analysis for the period from 1982 to 1986 (baseline period), the latitudinal variations of
TMS for NA and EA show similar changes across latitudes (
Figure 2). Although the absolute values of
TMS in EA are one to two degrees higher than the one in NA between latitudes 55°N and 70°N, the average rate of decrease in
TMS per degree latitude toward north is 0.5 K for both NA and EA (
Table 5). However, vegetated regions in NA and EA do not share the same spatial pattern in
PAT across latitudes. The average rate of decrease in
PAT per degree latitude toward north in NA is more than two times faster than that in EA (35 mm per degree in NA
vs. 15 mm per degree in EA,
Table 5). This implies that at a given latitude, northward migration of vegetation would require greater precipitation changes in NA than in EA, so as to create a favorable temperature and precipitation environment for vegetation to the south. Precipitation may not be as essential as temperature in governing the growth of local vegetation in the north [
7], especially in the Arctic. It is still important for precipitation to keep the same pace as the temperature change in order to support the northward migration of structurally different vegetation, such as shrubs and trees, which need more water supplies. The choice of
PAT is based on the fact that winter biological processes can contribute to the positive feedback of vegetation growth related to winter snow accumulation [
22,
36], and summer precipitation is also responsible for the vegetation changes [
12,
14,
21,
49].
3.3. Velocity of Climate
Velocity of climate is defined as the ratio of the (time) trend to the baseline north-south gradient of a given variable (e.g.,
TMS,
PAT,
etc.). Although the velocity of precipitation is expected to have similar spatial patterns to temperature velocity, but with higher uncertainties [
46], previous studies have only focused on vegetation response to temperature velocity [
41]. We analyzed both temperature and precipitation velocities using
TMS and
PAT along the North-South direction for two time periods—the earlier 18-year period from 1982 to 1999, and the entire 30-year period from 1982 to 2011 (from 1982 to 2009 for precipitation)—with the emphasis on the spatial matching of temperature and precipitation velocities. Such a consistency in velocity is crucial to our hypothesis,
i.e., a favorable environment for vegetation migration is only available when the precipitation velocity keeps up with the temperature velocity.
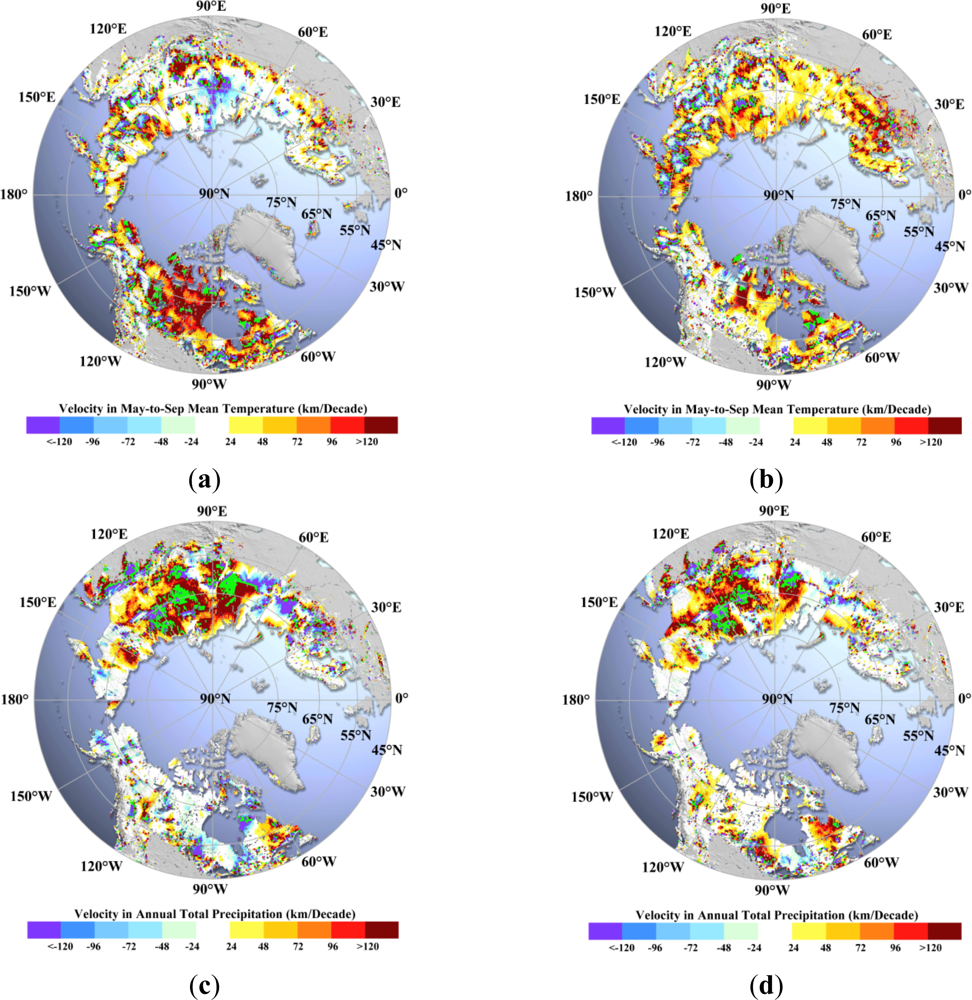
For the period from 1982 to 1999, both NA and EA show a majority of positive velocity of temperature change (
Figure 5), while precipitation velocity in NA is mainly negative velocity in contrast to a mostly positive velocity in EA (
Figure 5(c)). Positive velocity here indicates a northward movement. More than 28% (11%) of the vegetated areas in NA (EA) have high rates of positive velocity in temperature (>100 km/decade,
Table 6). The fraction of areas with high rates of positive precipitation velocity (>100 km/decade) in EA is four times larger than that in NA (
Table 6). Although the precipitation velocity has different patterns between NA and EA, vegetation in both regions still shows the consistent response to temperature (
Figure 3). This can be due to the uncertainties due to the shorter time period, and lag in vegetation response to precipitation velocity [
14,
22,
36,
49].
For the period from 1982 to 2011, the velocity measurements are statistically more reliable due to the longer period. With both NA and EA showing a majority of positive velocity in both temperature and precipitation (
Figure 5(b,d)), NA exhibits a lack of high rates of positive precipitation velocity compared to temperature velocity, particularly in the Arctic. Fractions of vegetated areas with high rates of positive velocity in temperature (>100 km/decade) are comparable between NA (17%) and EA (17%) (
Table 6). However, the fraction of vegetated areas with a high rate of positive velocity of precipitation (>100 km/decade) is smaller in the Arctic of NA (4%) compared to temperature (17%), unlike in EA. This difference in velocities of temperature and precipitation in the Arctic of NA could account for dramatic (unpredictable) greening, but a later decrease in vegetation sensitivity to temperature changes. Concordant velocities of temperature and precipitation, such as in EA and in the Boreal of NA, support continued vegetation migration (greening).
The actual vegetation migration rates depend on other factors such as land cover types [
44] and the sizes and distributions of natural habitats [
51]. On the other hand, the inconsistent velocities of temperature and precipitation shifts in the Arctic of NA are creating new climate states, leading to unpredictable vegetation responses.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}