Quantifying Ancient Maya Land Use Legacy Effects on Contemporary Rainforest Canopy Structure


Abstract
:
1. Background and Rationale
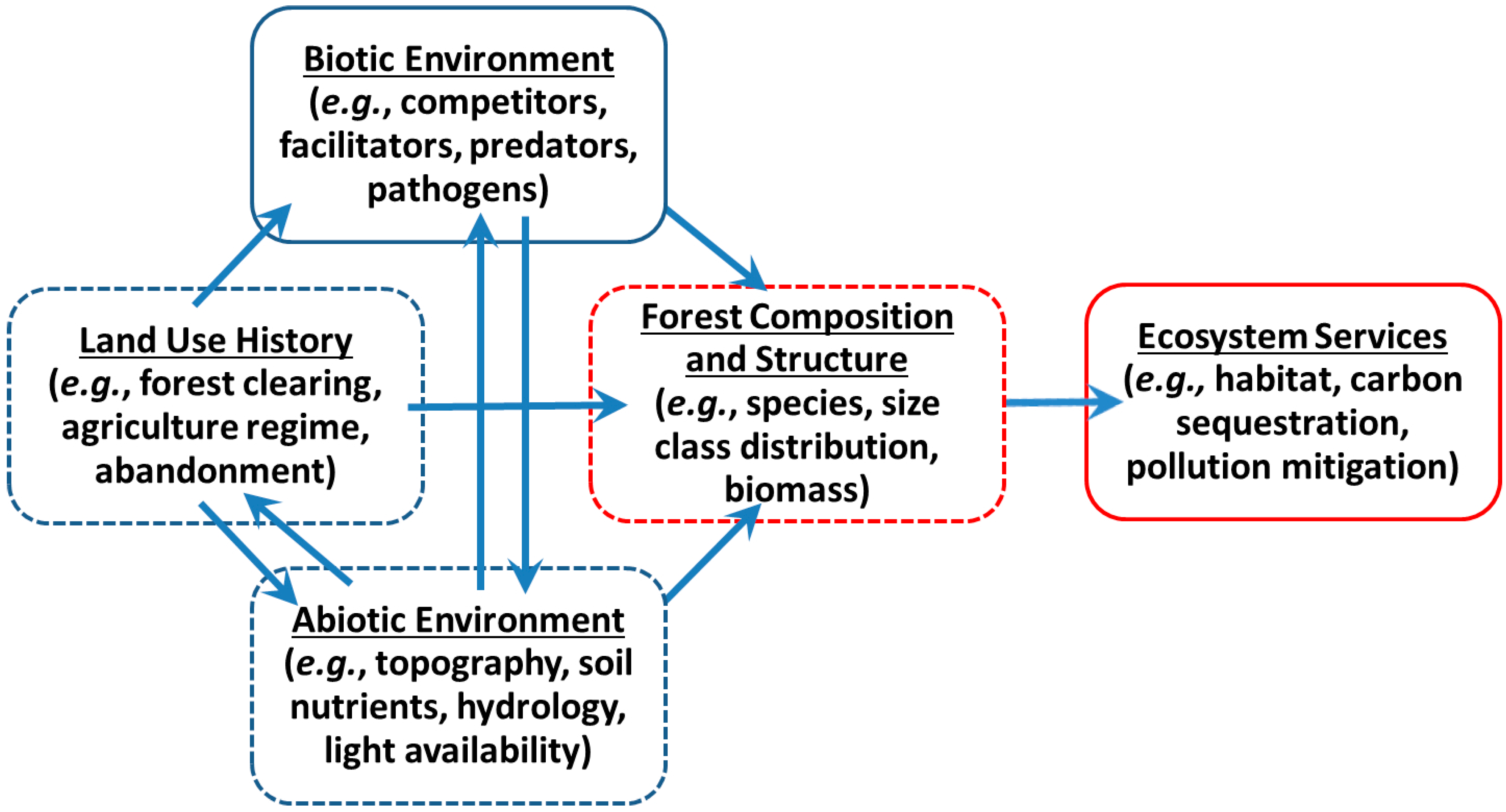
1.1. Human Land Use Legacies

1.2. Forest Canopy Structure
2. Objectives
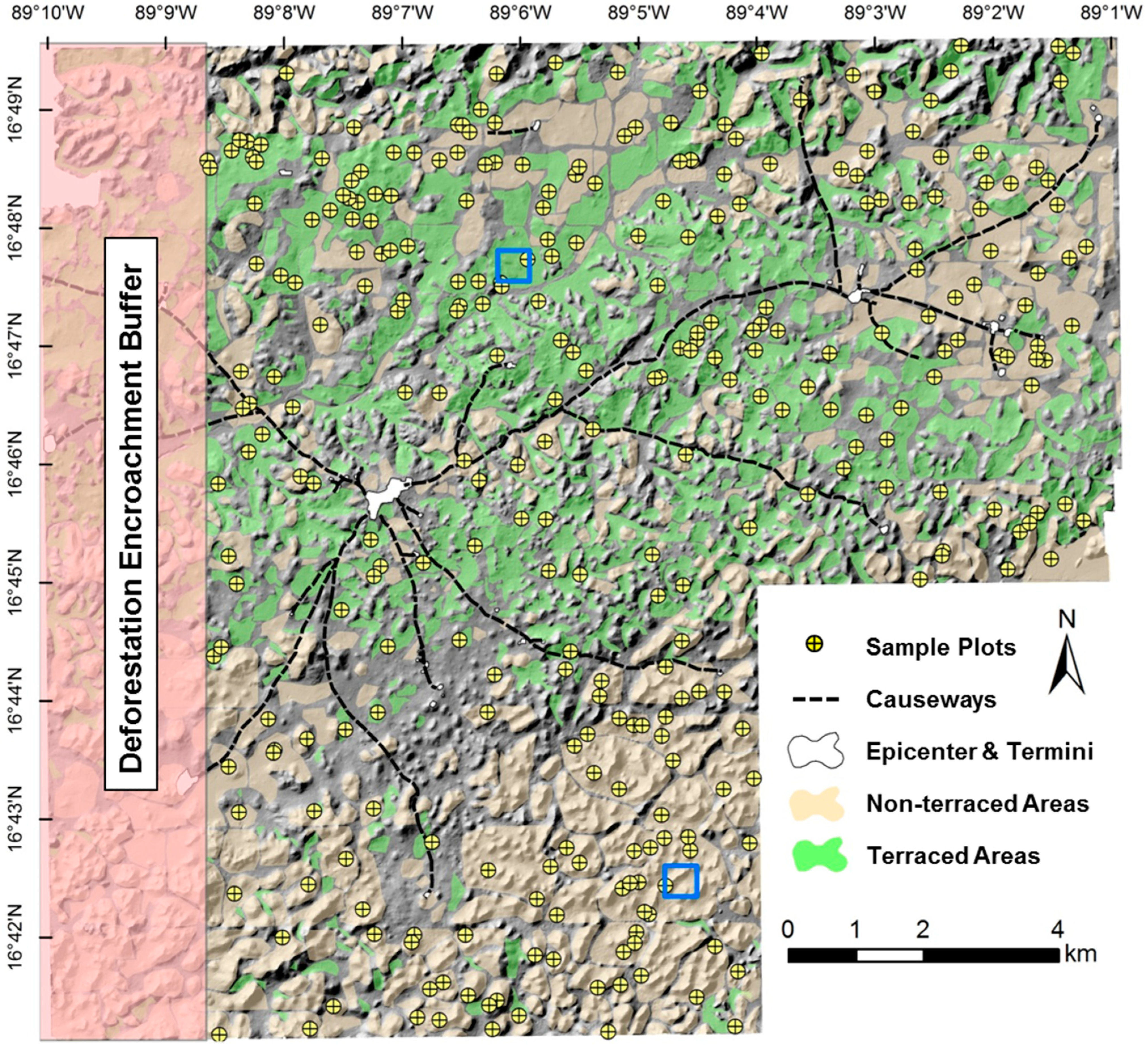
3. Study Area

4. Methods
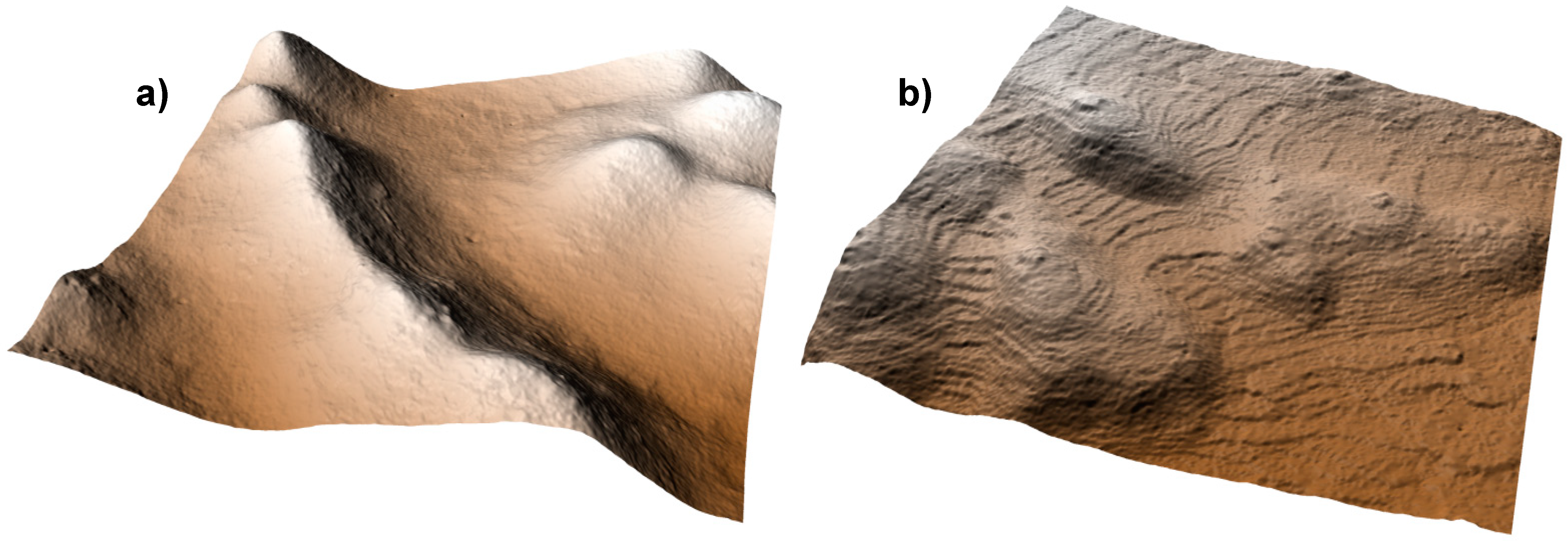
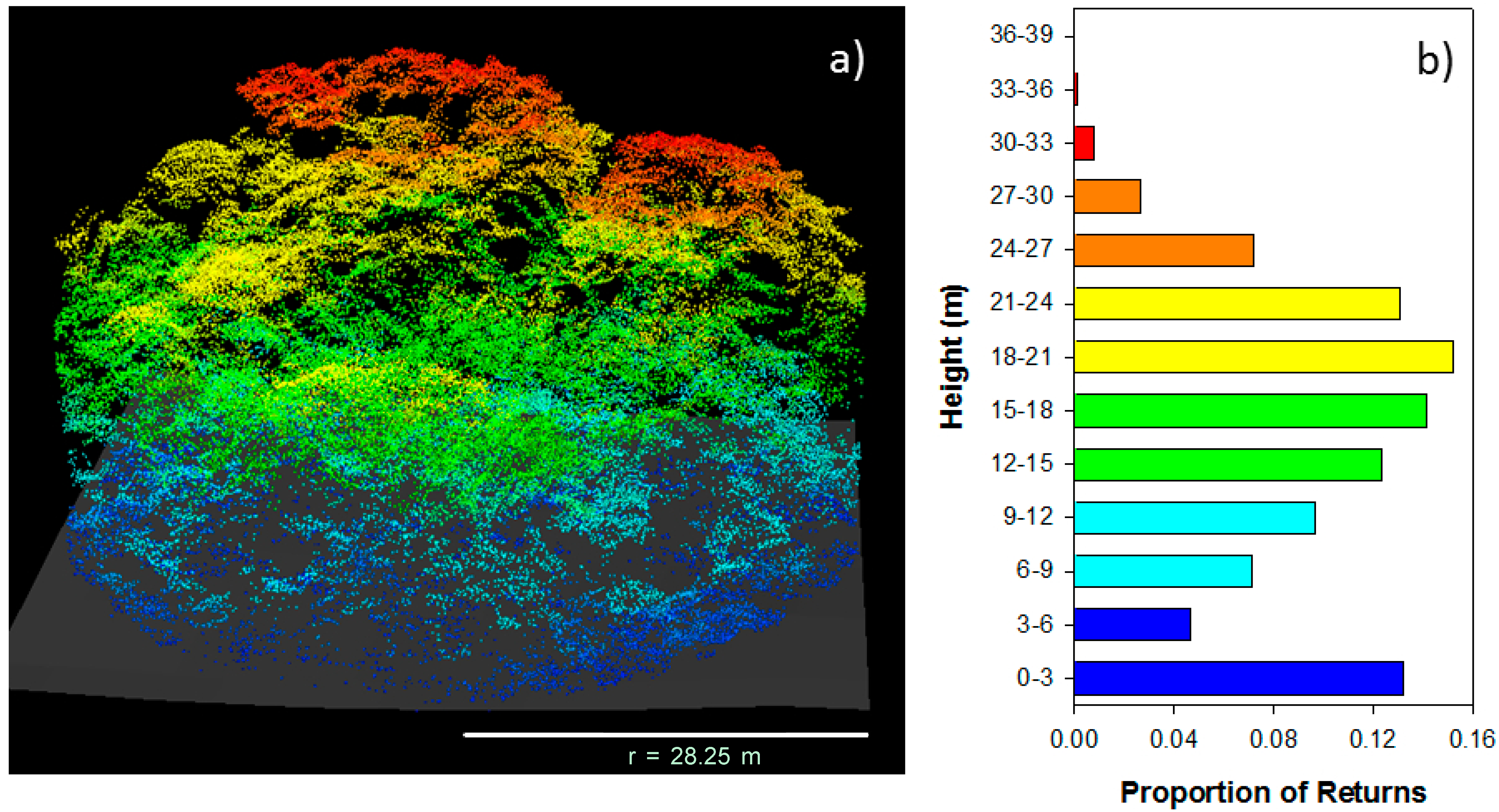
4.1. LiDAR Data Collection, Extraction, and Sampling


4.2. Statistical Analysis
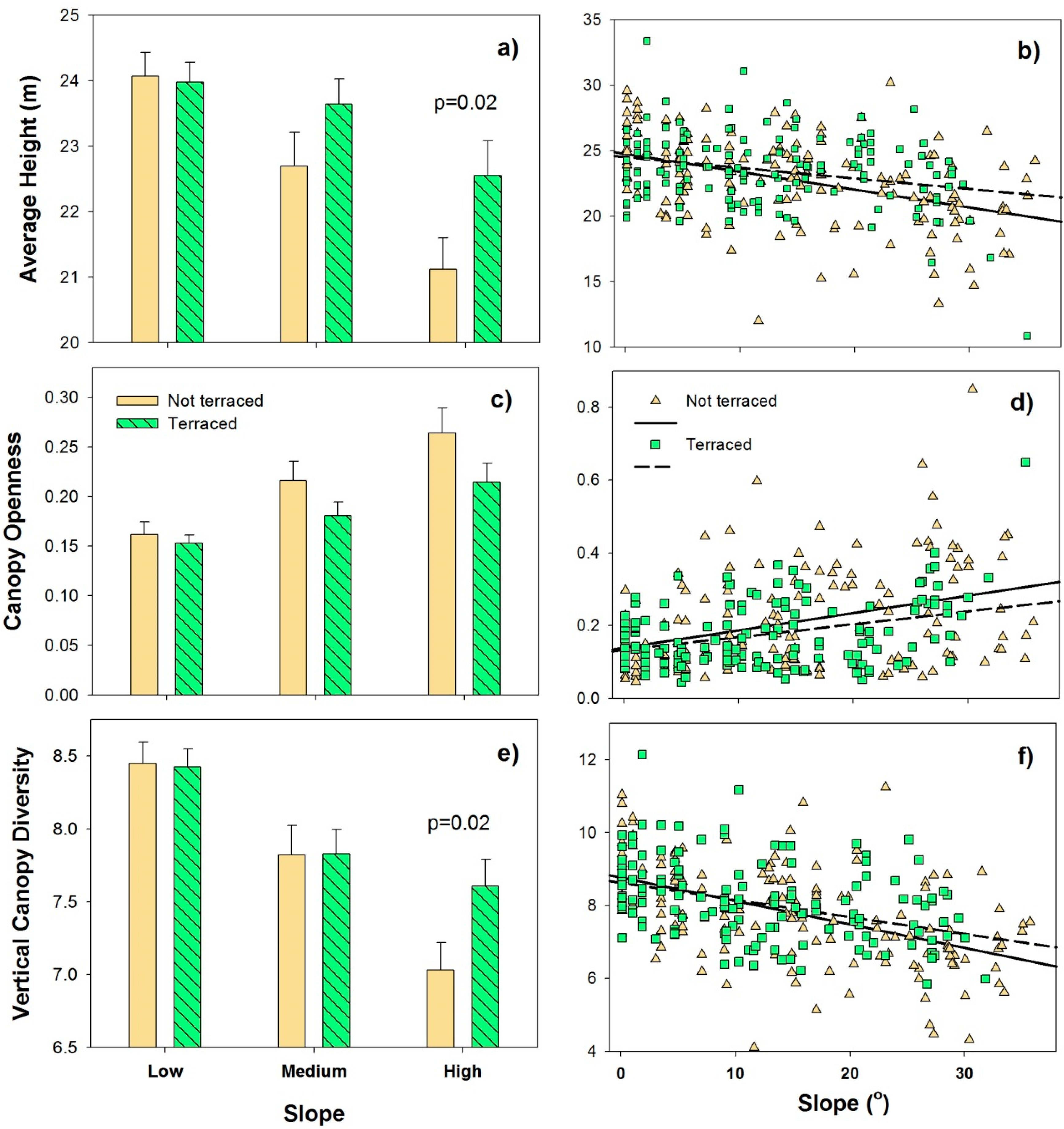
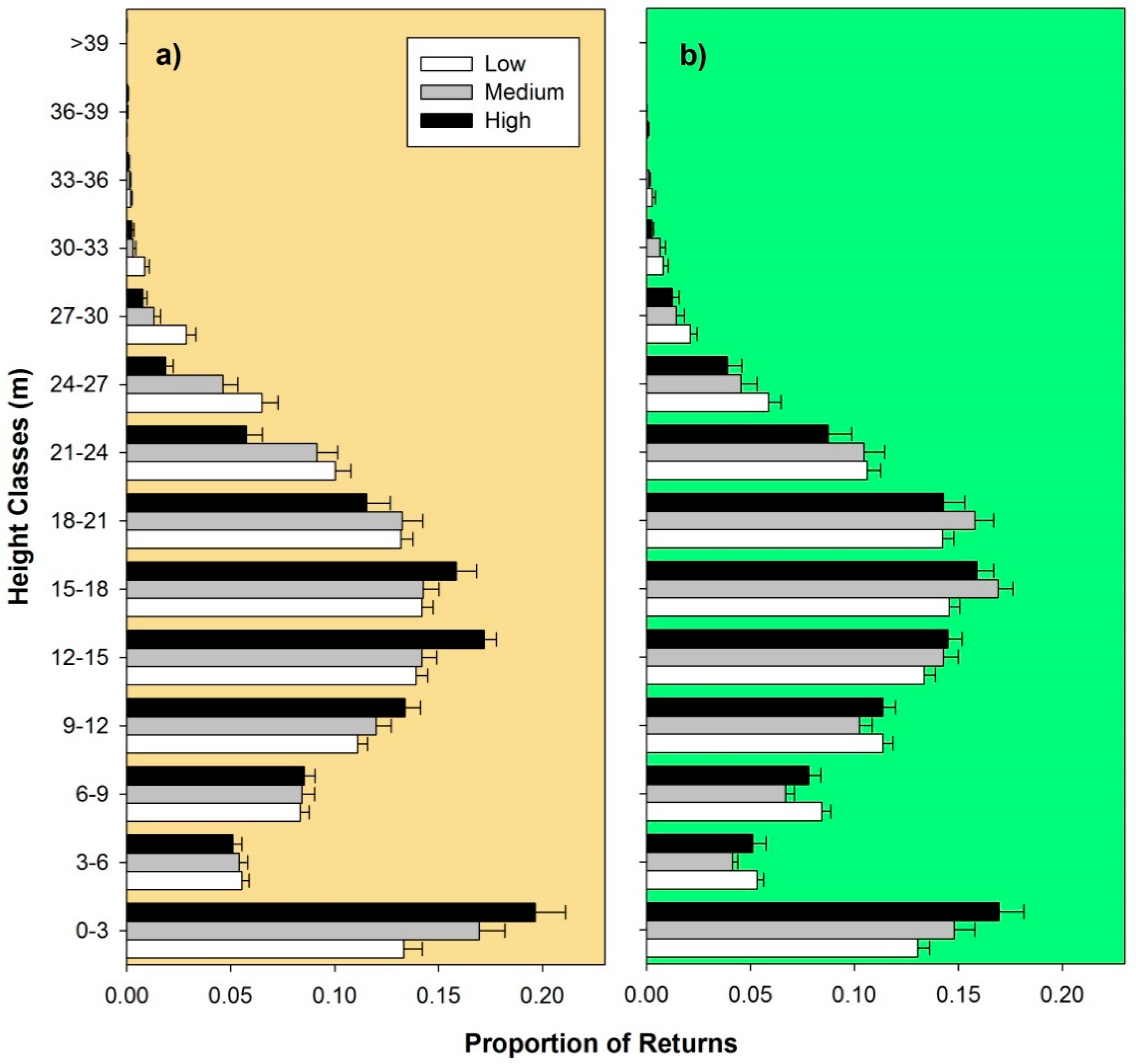
5. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources of Variation | df | Mean Square | F | r2 | p |
|---|---|---|---|---|---|
| Land Use Type * | 1 | 0.024 | 5.04 | 0.013 | 0.016 |
| Slope | 1 | 0.134 | 50.82 | 0.134 | 0.001 |
| Elevation | 1 | 0.028 | 10.74 | 0.028 | 0.001 |
| Slope: Aspect | 1 | 0.022 | 4.54 | 0.012 | 0.026 |
| Slope: Elevation | 1 | 0.042 | 15.78 | 0.041 | 0.001 |
| Residuals | 284 | 0.005 | 0.748 |
| Sources of Variation | df | Mean Square | F | r2 | p |
|---|---|---|---|---|---|
| Land Use Type * | 1 | 0.227 | 5.02 | 0.015 | 0.005 |
| Slope | 1 | 1.048 | 23.21 | 0.068 | 0.001 |
| Land Use Type: Slope | 1 | 0.170 | 3.76 | 0.011 | 0.018 |
| Slope: Elevation | 1 | 0.482 | 10.68 | 0.031 | 0.001 |
| Residuals | 284 | 0.045 | 0.083 |


6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. Perspect. Plant Ecol. 2003, 6, 51–71. [Google Scholar]
- Foster, D.R.; Swanson, F.; Aber, J.; Burke, I.; Brokaw, N.; Tilman, D.; Knapp, A. The importance of land-use legacies to ecology and conservation. BioScience 2003, 53, 77–88. [Google Scholar]
- Ruddiman, W.F. Earth Transformed; W.H. Freeman and Co.: New York, NY, USA, 2014. [Google Scholar]
- Flinn, K.M.; Vellend, M. Recovery of forest plant communities in post-agricultural landscapes. Front Ecol. Environ. 2005, 3, 243–250. [Google Scholar]
- Sponsel, L.E.; Bailey, R.C.; Headland, T.N. Anthropological perspectives on the causes, consequences, and solutions of deforestation. In Tropical Deforestation: The Human Dimension; Sponsel, L.E., Headland, T.N., Bailey, R.C., Eds.; Columbia Univesity Press: New York, NY, USA, 1996; pp. 3–52. [Google Scholar]
- Laurance, W.F. Reflections of the tropical deforestation crisis. Biol. Conserv. 1999, 91, 109–117. [Google Scholar]
- Achard, F.; Eva, H.D.; Stibig, H.; Mayaux, P.; Gallego, J.; Richards, T.; Malingreau, J. Determination of deforestation rates of the world’s humid tropical forests. Science 2002, 297, 999–1002. [Google Scholar]
- Bellemare, J.; Motzkin, G.; Foster, D.R. Legacies of the agricultural past in the forested present: An assessment of historical land-use effects on rich mesic forests. J. Biogeogr. 2002, 29, 1401–1420. [Google Scholar]
- Sanford, R.L., Jr.; Horn, S.P. Holocene rain-forest wilderness: A neotropical perspective on humans as an exotic, invasive species. In USDA Forest Service RMRS-P-15-VOL-3, Proceedings of Wilderness Science in a Time of Change; McCool, S.F., Cole, D.N., Eds.; Rocky Mountain Research Station: Ogden, UT, USA, 2000; pp. 168–173. [Google Scholar]
- Gomez-Pompa, A.; Allen, M.F.; Fedick, S.L.; Jimenez-Osornio, J.J. The Maya Lowlands: A case study for the future? Conclusions. In The Lowland Maya Area: Three Millennia at the Human-Wildland Interface; Gomez-Pompa, A., Allen, M.F., Fedick, S.L., Jimenez-Osornio, J.J., Eds.; Hawthorn Press, Inc.: New York, NY, USA, 2003; pp. 623–631. [Google Scholar]
- Clark, D.B. Abolishing virginity. J. Trop. Ecol. 1996, 12, 735–739. [Google Scholar]
- Heckenberger, M.J.; Kuikuro, A.; Kuikuro, U.T.; Russell, J.C.; Schmidt, M.; Fausto, C.; Franchetto, B. Amazonia 1492: Pristine forest or cultural parkland. Science 2003, 301, 1710–1714. [Google Scholar]
- Delcourt, H.R. The impact of prehistoric agriculture and land occupation on natural vegetation. Trends Ecol. Evol. 1987, 2, 39–44. [Google Scholar]
- Butzer, K.W. Ecology in the long view: Settlement histories, agrosystemic strategies, and ecological performance. J. Field Archaeol. 1996, 23, 141–150. [Google Scholar]
- Mann, C.C. 1491: New Revelations of the Americas before Columbus; Knopf Press: New York, NY, USA, 2005. [Google Scholar]
- Peterken, G.F.; Game, M. Historical factors affecting the number and distribution of vascular plant species in the woodlands of central Lincolnshire. J. Ecol. 1984, 72, 155–182. [Google Scholar]
- Motzkin, G.; Foster, D.; Allen, A.; Harrod, J.; Boone, R. Controlling site to evaluate history: Vegetation patterns of a New England sand plain. Ecol. Monogr. 1996, 66, 345–365. [Google Scholar]
- Briggs, J.M.; Spielmann, K.A.; Schaafsma, H.; Kintigh, K.W.; Kruse, M.; Morehouse, K.; Schollmeyer, K. Why ecology needs archaeologists and archaeology needs ecologists. Front. Ecol. Environ. 2006, 4, 180–188. [Google Scholar]
- Dambrine, E.; Dupouey, J.L.; Laut, L.; Humbert, L.; Thinon, M.; Beaufils, T.; Richard, H. Present forest biodiversity patterns in France related to former Roman agriculture. Ecology 2007, 88, 1430–1439. [Google Scholar]
- Turner, B.L., II; Klepeis, P.; Schneider, L.C. Three millennia in southern Yucatan peninsula: Implications for occupancy, use, and carrying capacity. In The Lowland Maya Area: Three Millennia at the Human-Wildland Interface; Gomez-Pompa, A., Allen, M.F., Fedick, S.L., Jimenez-Osornio, J.J., Eds.; Hawthorn Press, Inc.: New York, NY, USA, 2003; pp. 361–373. [Google Scholar]
- Gómez-Pompa, A.; Flores, J.S.; Sosa, V. The “pet kot”: A man-made tropical forest of the Maya. Interciencia 1987, 12, 10–15. [Google Scholar]
- Lambert, J.D.H.; Arnason, T. Distribution of vegetation on Maya ruins and its relationship to ancient land-use at Lamanai, Belize. Turrialba 1978, 28, 33–41. [Google Scholar]
- Lambert, J.D.H.; Arnason, J.T. Rámon and Maya ruins: An ecological, not an economic, relation. Science 1982, 216, 298–299. [Google Scholar]
- Harper, K.A.; MacDonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.; Jaiteh, M.S.; Esseen, P. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2010, 91, 2833–2849. [Google Scholar]
- Aide, M.T.; Zimmerman, J.K.; Pascarella, J.B.; River, L.; Marcano-Vega, H. Forest regeneration in a chronosequence of tropical abandoned pastures: Implication for restoration ecology. Restor. Ecol. 2000, 8, 328–338. [Google Scholar]
- Mueller, A.D.; Islebe, G.A.; Anselmetti, F.S.; Ariztegui, D.; Brenner, M.; Hodell, D.A.; Hajdas, I.; Hamann, Y.; Haug, G.H.; Kennett, D.J. Recovery of the forest ecosystem in the tropical lowlands of northern Guatemala after disintegration of Classic Maya polities. Geology 2010, 38, 523–526. [Google Scholar]
- Clark, D.B.; Clark, D.B. Landscape-scale variation in forest structure and biomass in a tropical rain forest. For. Ecol. Manag. 2000, 137, 185–198. [Google Scholar]
- Tateno, R.; Takeda, H. Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soil nitrogen and light at the forest floor. Ecol. Res. 2003, 18, 559–571. [Google Scholar]
- Spies, T.A. Forest structure: A key to the ecosystem. Northwest Sci. 1998, 72, 34–36. [Google Scholar]
- Alves, L.F.; Vieira, S.A.; Scaranello, M.A.; Camargo, P.B.; Santos, F.A.M.; Joly, C.A.; Martinelli, L.A. Forest structure and live aboveground biomass variation along an elevational gradient of tropical Atlantic moist forest (Brazil). For. Ecol. Manag. 2010, 260, 679–691. [Google Scholar]
- Turner, M.G. Landscape ecology: The effect of pattern on process. Annu. Rev. Ecol. Syst. 1989, 20, 171–197. [Google Scholar]
- MacArthur, R.H.; MacArthur, J.W. On bird species diversity. Ecology 1961, 42, 594–598. [Google Scholar]
- Verschuyl, J.P.; Hansen, A.J.; McWethy, D.B.; Sallabanks, R.; Hutto, R.L. Is the effect of forest structure on bird diversity modified by forest productivity. Ecol. Appl. 2008, 18, 1155–1170. [Google Scholar]
- Müller, J.; Bae, S.; Röder, J.; Chao, A.; Didham, R.K. Airborne LiDAR reveals context dependence in the effects of canopy architecture on arthropod diversity. For. Ecol. Manag. 2014, 312, 129–137. [Google Scholar]
- Alvarez, E.; Duque, A.; Saldarriaga, J.; Cabrera, K.; de las Salas, G.; del Valle, I.; Lema, A.; Moreno, F.; Orrego, S.; Rodríguez, L. Tree above-ground biomass allometries for carbon stocks estimation in the natural forests of Columbia. For. Ecol. Manag. 2012, 267, 297–308. [Google Scholar]
- Prentice, I.C.; Leemans, R. Pattern and process and the dynamics of forest structure: A simulation approach. J. Ecol. 1990, 78, 340–355. [Google Scholar]
- Brokaw, N.V.L. Gap-phase regeneration in a tropical forest. Ecology 1992, 66, 682–687. [Google Scholar]
- Pacala, S.W.; Deutschman, D.H. Details that matter: The spatial distribution of individual trees maintains forest ecosystem function. Oikos 1995, 74, 357–365. [Google Scholar]
- Phillips, O.L.; Baker, T.R.; Arroyo, L.; Higuchi, N.; Killeen, T.; Laurance, W.F.; Lewis, S.L.; Lloyd, J.; Malhi, Y.; Monteagudo, A.; et al. Pattern and process in Amazon forest dynamics, 1976–2001. Philos. Trans. R Soc. B 2004, 359, 381–407. [Google Scholar]
- Shugart, H.H.; Saatchi, S.; Hall, F.G. Importance of structure and its measurement in quatifying function of forest ecosystems. J. Geophys. Res. 2010, 115, G00E13. [Google Scholar] [CrossRef]
- Urquiza-Haas, T.; Dolman, P.M.; Peres, C.A. Regional scale variation in forest structure and biomass in the Yucatán Peninsula: Effects of forest disturbance. For. Ecol. Manag. 2007, 247, 80–90. [Google Scholar]
- Holdridge, L.R.; Grenke, W.C.; Hatheway, W.H.; Liang, T.; Tosi, J.A. Forest Environments in Tropical Life Zones: A Pilot Study; Pergamon: New York, NY, USA, 1971. [Google Scholar]
- Detto, M.; Muller-Landau, H.C.; Mascaro, J.; Asner, G.P. Hydrological networks and associated topographic variation as templates for the spatial organization of tropical forest vegetation. PLoS One 2013, 8, e76296. [Google Scholar] [CrossRef]
- Homeier, J.; Breckle, S.W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich Ecuadorian montane rain forest. Biotropica 2010, 42, 140–148. [Google Scholar]
- Weishampel, J.F.; Drake, J.B.; Cooper, A.; Blair, J.B.; Hofton, M. Forest canopy recovery from the 1938 hurricane and subsequent damage measured with airborne LiDAR. Remote Sens. Environ. 2007, 190, 142–153. [Google Scholar]
- Laurance, W.F.; Ferreira, L.V.; Rankin-de Merona, J.M.; Laurance, S.G. Rain forest fragmentation and the dynamics of Amazonian tree communities. Ecology 1998, 79, 2032–2040. [Google Scholar]
- Coultas, C.L.; Collins, M.C.; Chase, A.F. Some soils common to Caracol, Belize and their significance to ancient agriculture and land-use. In Studies in the Archaeology of Caracol, Belize; Chase, D., Chase, A., Eds.; Pre-Columbian Art Research Institute: San Francisco, CA, USA, 1994; Monograph 7; pp. 21–33. [Google Scholar]
- Chase, A.F.; Chase, D.Z. Scale and intensity in classic period Maya agriculture: Terracing and settlement at the “Garden City” of Caracol, Belize. Cult. Agric. 1998, 20, 60–77. [Google Scholar]
- Opitz, R.S.; Cowley, D.C. Interpreting Archaeological Topography: 3D Data; Visualisation and Observation Oxbow Books: Oxford, UK, 2013. [Google Scholar]
- Chase, A.F.; Chase, D.Z.; Weishampel, J.F. Lasers in the jungle: Airborne sensors reveal a vast Maya landscape. Archaeology 2010, 63, 27–29. [Google Scholar]
- Weishampel, J.F.; Hightower, J.N.; Chase, A.F.; Chase, D.Z. Remote sensing of below canopy land use features from the Maya polity of Caracol. In Understanding Landscapes, from Discovery to Their Spatial Organization; British Archaeological Reports S2541; Djinjian, F., Robert, S., Eds.; Archaeopress: Oxford, UK, 2013; pp. 131–136. [Google Scholar]
- Friesner, J. Hurricanes and the Forests of Belize; Forest Department, Ministry of Natural Resources and the Environment: Belmopan City, Belize, 1993; p. 20. [Google Scholar]
- Weishampel, J.F.; Hightower, J.N.; Chase, A.F.; Chase, D.Z. Use of airborne LiDAR to delineate canopy degradation and encroachment along the Guatemala—Belize border. Trop. Conserv. Biol. 2012, 5, 12–24. [Google Scholar]
- Webb, E.A.; Schwarcz, H.P.; Healy, P.F. Detection of ancient maize in lowland Maya soils using stable carbon isotopes: Evidence from Caracol, Belize. J. Archaeol. Sci. 2004, 31, 1039–1052. [Google Scholar]
- Murtha, T. Land and Labor: Maya Terraced Agriculture: An Investigation of the Settlement Economy and Intensive Agricultural Landscape of Caracol, Belize; Verlag, D.M., Ed.; Muller: Saarbruckan, Germany, 2009. [Google Scholar]
- Chase, A.F.; Chase, D.Z.; Weishampel, J.F.; Drake, J.B.; Shrestha, R.L.; Slatton, K.C.; Awe, J.J.; Carter, W.E. Airborne LiDAR, archaeology, and the ancient Maya landscape at Caracol, Belize. J. Archaeol. Sci. 2011, 38, 387–398. [Google Scholar]
- Chase, A.F.; Chase, D.Z.; Fisher, C.T.; Leisz, S.J.; Weishampel, J.F. Geospatial revolution and remote sensing LiDAR in Mesoamerican archaeology. PNAS 2012, 109, 12837–12838. [Google Scholar]
- Beach, T.; Luzzadder-Beach, S.; Dunning, N.; Hageman, J.; Lohse, J. Upland agriculture in the Maya lowlands: Ancient Maya soil conservation in northwestern Belize. Geogr. Rev. 2002, 92, 372–397. [Google Scholar]
- Chase, A.S.Z.; Weishampel, J.F. Water capture and agricultural terracing at Caracol, Belize as revealed through the 2009 LiDAR campaign. 2014; in revision. [Google Scholar]
- Weishampel, J.F.; Chase, A.F.; Chase, D.Z.; Drake, J.B.; Shrestha, R.L.; Slatton, K.C.; Awe, J.J.; Hightower, J.; Angelo, J. Remote sensing of ancient Maya land use features at Caracol, Belize related to tropical rainforest structure. In Proceedings of the SpaceTimePlace: Third International Conference on Remote Sensing in Archaeology, Tamil Nadu, India, 17–21 August 2009; British Archaeological Reports S2118. Campana, S., Forte, M., Liuzza, C., Eds.; Archaeopress: Oxford, UK, 2010; pp. 45–52. [Google Scholar]
- Fernandez-Diaz, J.C.; Carter, W.E.; Shrestha, R.L.; Glennie, C.L. Now you see it…now you don’t: Understanding airborne mapping LiDAR collection and data product generation for archaeological research in Mesoamerica. Remote Sens. 2014, 6, 9951–10001. [Google Scholar]
- Beyer, H.L. Hawth’s Analysis Tools for ArcGIS. Available online: http://www.spatialecology.com/htools (accessed on 21 March 2012).
- McGaughey, R.J. FUSION/LDV: Software for LIDAR Data Analysis and Visualization; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Seattle, WA, USA, 2009; p. 123. [Google Scholar]
- Angelo, J.J.; Duncan, B.W.; Weishampel, J.F. Using LiDAR-derived vegetation profiles to predict time since fire in an oak scrub landscape in east-central Florida. Remote Sens. 2010, 2, 514–525. [Google Scholar]
- Drake, J.B.; Dubayah, R.O.; Clark, D.B.; Knox, R.G.; Blair, J.B.; Hofton, M.A.; Chazdon, R.L.; Weishampel, J.F.; Prince, S. Estimation of tropical forest structural characteristics using large-footprint lidar. Remote Sens. Environ. 2002, 79, 305–319. [Google Scholar]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–365. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 1.17–9. Available online: http://CRAN.R-project.org/package=vegan (accessed on 21 March 2012).
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Healy, P.F.; Lambert, J.D.H.; Arnason, J.T.; Hebda, R.J. Caracol, Belize: Evidence of ancient Maya agricultural terraces. J. Field Archaeol. 1983, 10, 397–410. [Google Scholar]
- Hightower, J.N. Relating Ancient Maya Land Use Legacies to the Contemporary Forest of Caracol, Belize. Master’s Thesis, University of Central Florida, Orlando, FL, USA, 2012; p. 71. [Google Scholar]
- Brewer, S.T.; Rejmanek, M.; Webb, A.H.; Fine, P.V.A. Relationship of phytogeagraphy and diversity of tropical tree species with limestone topography in southern Belize. J. Biogeogr. 2003, 30, 1669–1688. [Google Scholar]
- White, D.A.; Hood, C.S. Vegetation patterns and environmental gradients in tropical dry forests of the northern Yucatan Peninsula. J. Veg. Sci. 2004, 15, 151–160. [Google Scholar]
- Furley, P.A.; Newey, W.W. Variations in plant communities with topography over tropical limestone soils. J. Biogeogr. 1979, 6, 1–15. [Google Scholar]
- DeClerck, F.A.J.; Chazdon, R.; Holl, K.D.; Milder, J.C.; Finegan, B.; Martinez-Salinas, A.; Imbach, P.; Canet, L.; Ramos, Z. Biodiversity conservation in human-modified landscapes of Mesoamerica: Past, present, and future. Biol. Conserv. 2010, 143, 2301–2313. [Google Scholar]
- Wyatt, A.R. Gardens on Hills: Ancient Maya Terracing and Agricultural Production at Chan, Belize. PhD Dissertation, University of Illinois, Chicago, IL, USA, 2008; p. 449. [Google Scholar]
- Beach, T.; Dunning, N.P. Ancient Maya terracing and modern conservation in the Peten rain forest of Guatemala. J. Soil Water Conserv. 1995, 50, 138–145. [Google Scholar]
- Ackermann, O.; Svoray, T.; Haiman, M. Nari (calcrete) outcrop contributions to ancient agricultural terraces in the Southern Shephelah, Israel: Insights from digital terrain analysis and a geoarchaeological field survey. J. Archaeol. Sci. 2008, 35, 930–941. [Google Scholar]
- Pretto, F.; Celesti-Grapow, L.; Carli, E.; Blasi, C. Influence of past land use and current human disturbance on non-native plants species on small Italian islands. Plant Ecol. 2010, 2010, 225–239. [Google Scholar]
- McCoy, M.D.; Asner, G.P.; Graves, M.W. Airborne lidar survey of irrigated agricultural landscapes: An application of the slope contrast method. J. Archaeol. Sci. 2011, 38, 2141–2154. [Google Scholar]
- Nassauer, J.I.; Raskin, J. Urban vacancy and land use legacies: A frontier for urban ecological research, design, and planning. Landsc. Urban Plan. 2014, 125, 245–253. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hightower, J.N.; Butterfield, A.C.; Weishampel, J.F. Quantifying Ancient Maya Land Use Legacy Effects on Contemporary Rainforest Canopy Structure. Remote Sens. 2014, 6, 10716-10732. https://doi.org/10.3390/rs61110716
Hightower JN, Butterfield AC, Weishampel JF. Quantifying Ancient Maya Land Use Legacy Effects on Contemporary Rainforest Canopy Structure. Remote Sensing. 2014; 6(11):10716-10732. https://doi.org/10.3390/rs61110716
Chicago/Turabian StyleHightower, Jessica N., A. Christine Butterfield, and John F. Weishampel. 2014. "Quantifying Ancient Maya Land Use Legacy Effects on Contemporary Rainforest Canopy Structure" Remote Sensing 6, no. 11: 10716-10732. https://doi.org/10.3390/rs61110716