On the Response of European Vegetation Phenology to Hydroclimatic Anomalies


Abstract
:1. Introduction
2. Materials and Methods
2.1. FAPAR Data and Computation of Anomalies
2.2. Anomalies of Rainfall and Temperature
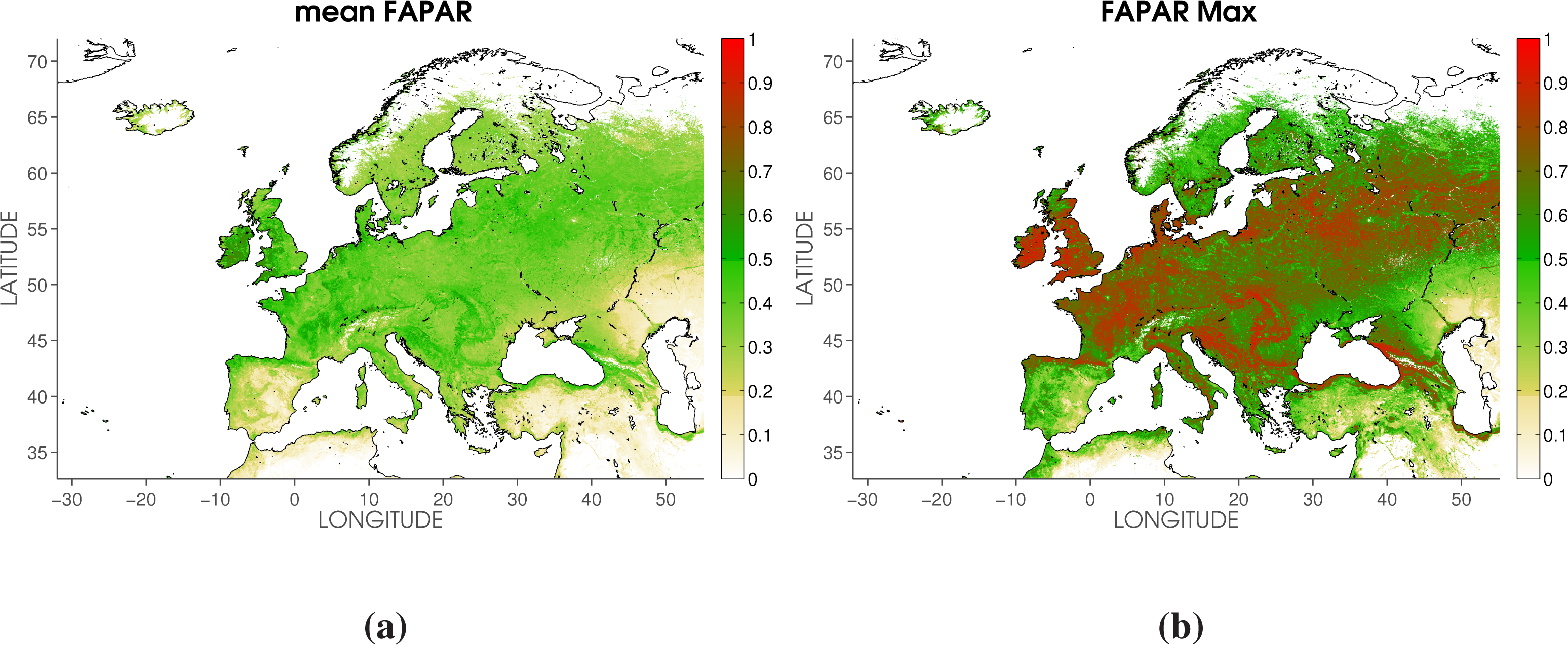
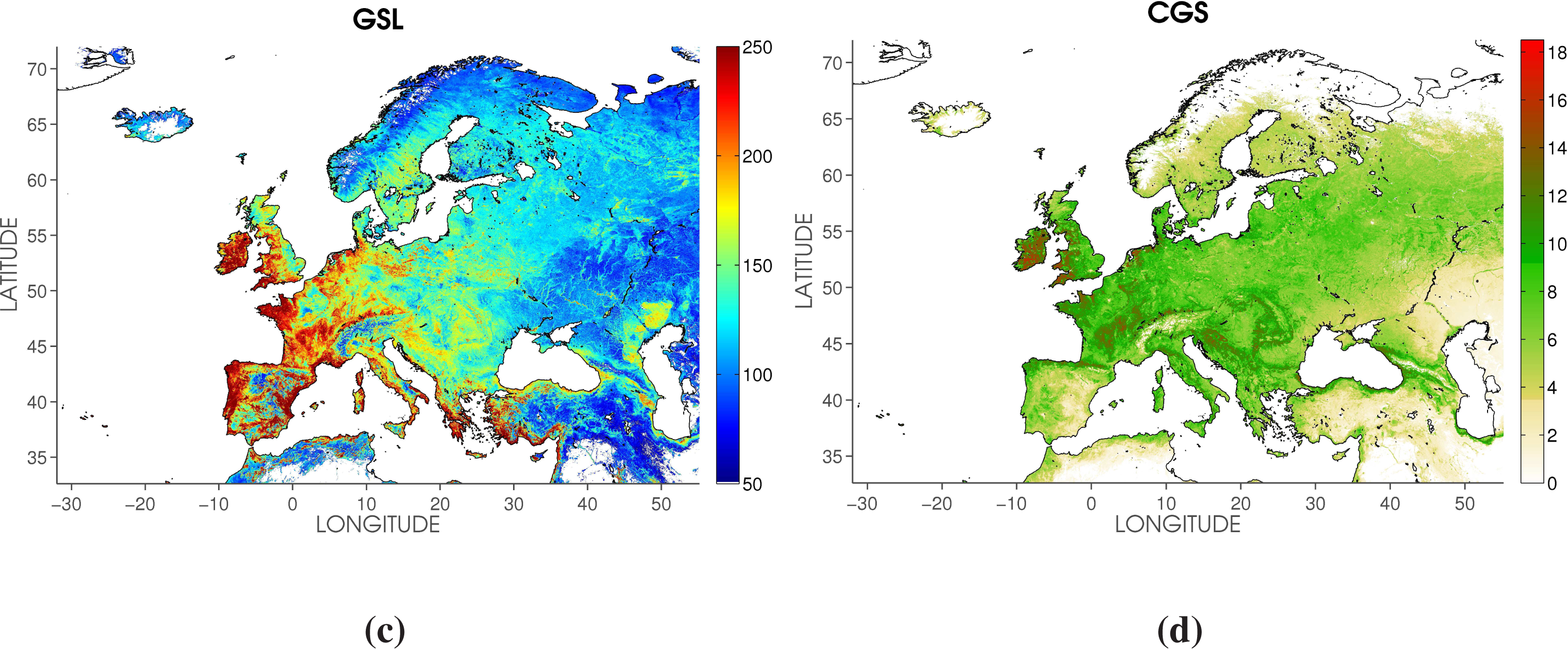
2.3. Phenology Metrics
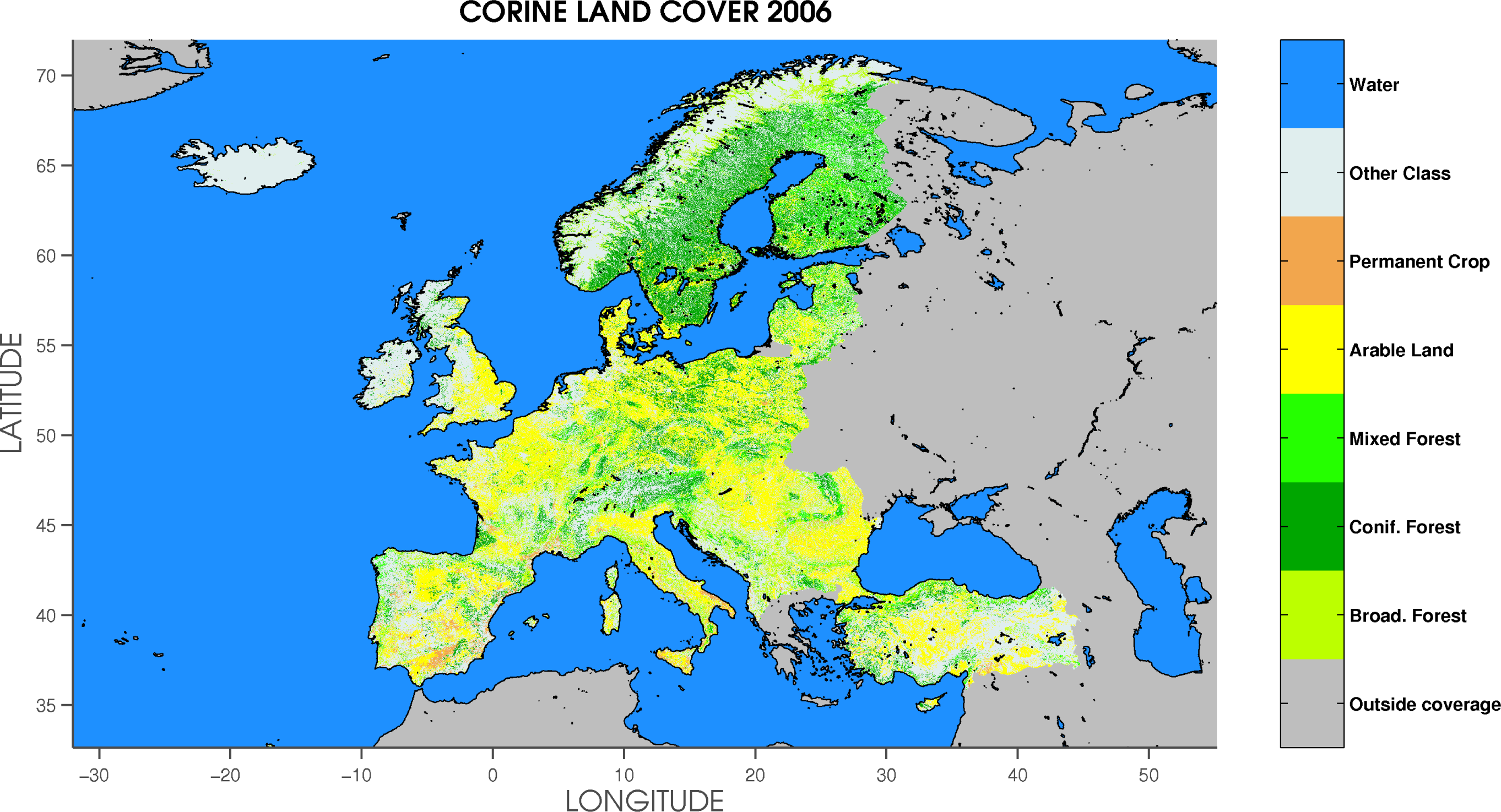
2.4. Land Cover
2.5. Correlation Analysis
3. Results
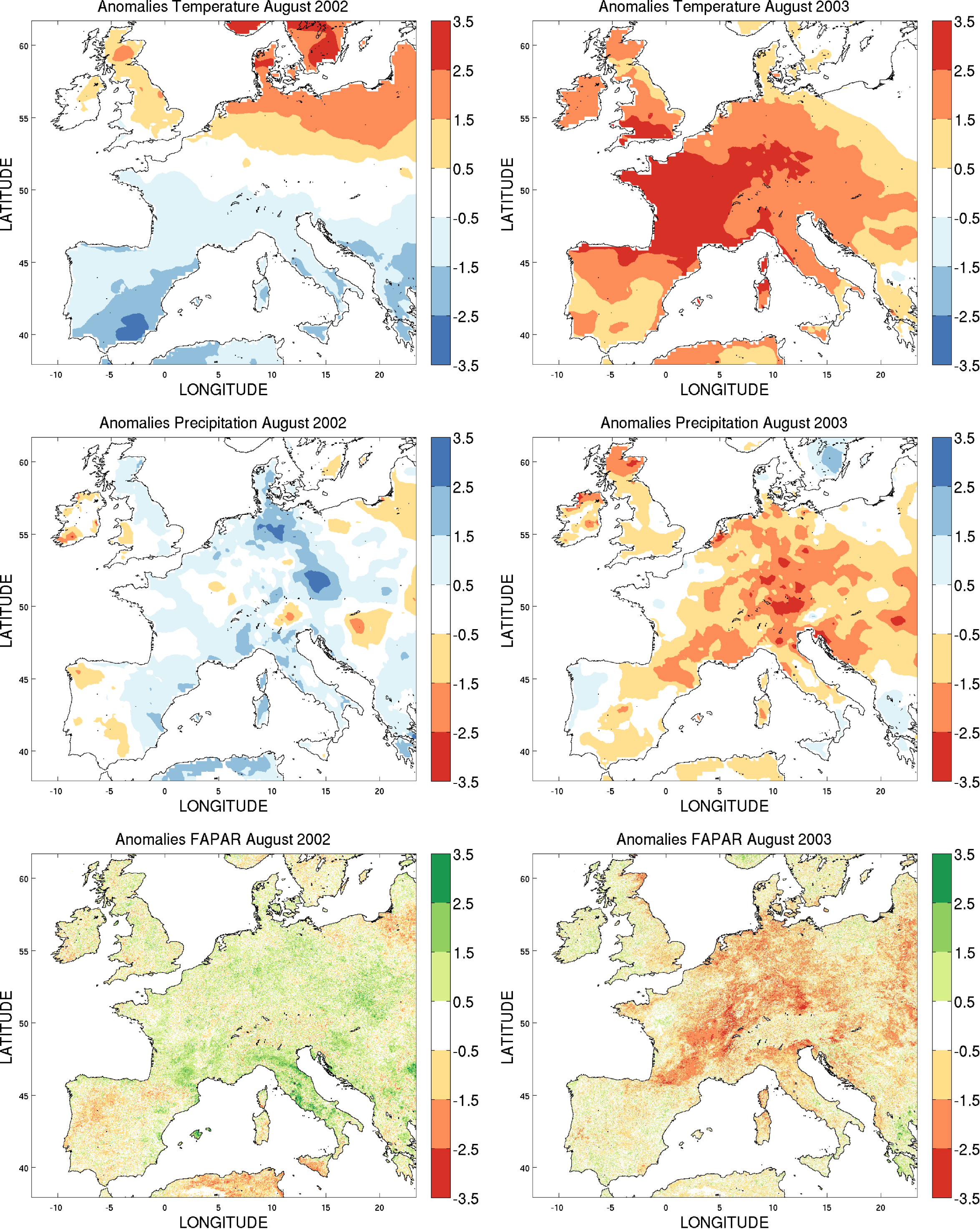
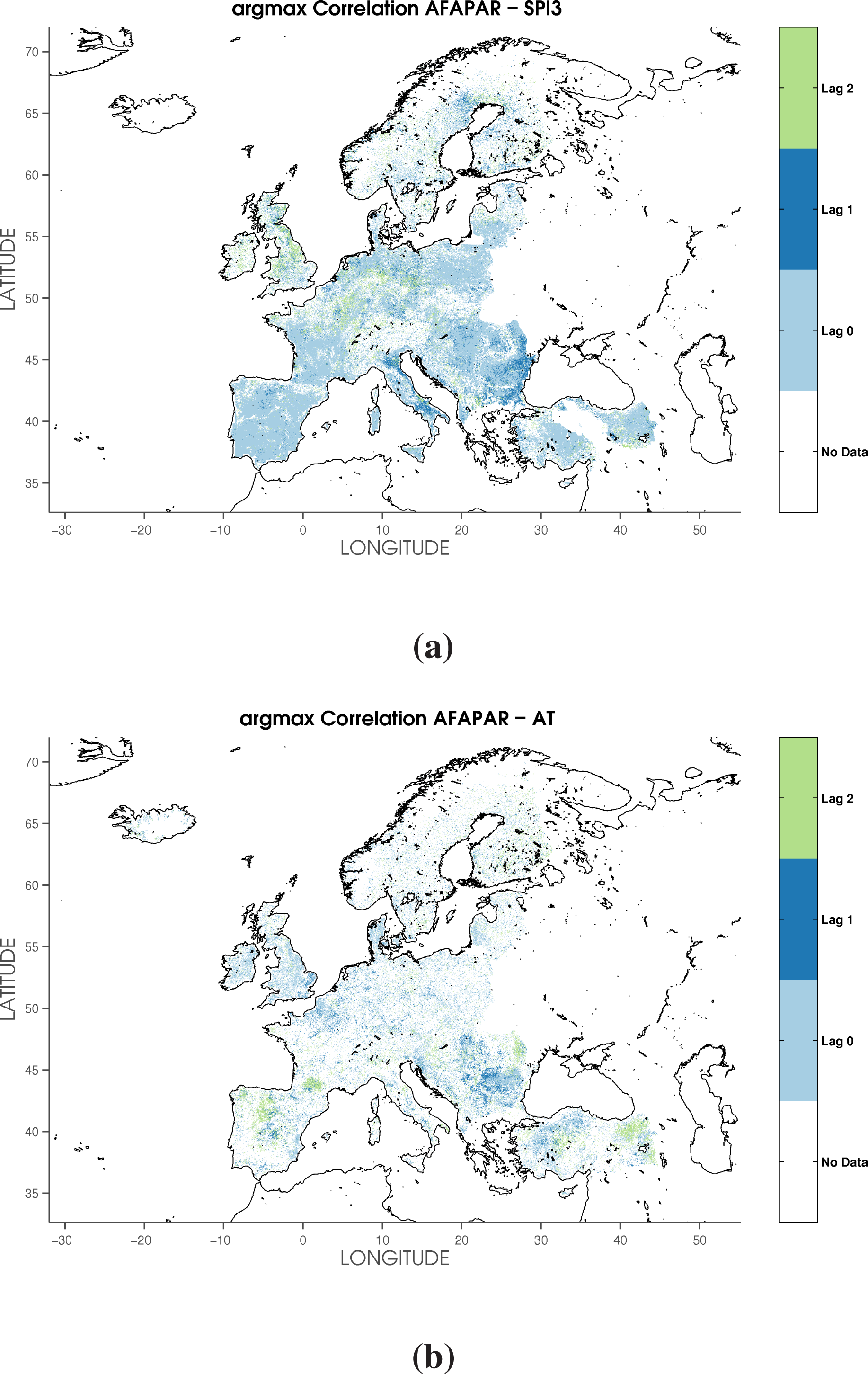
3.1. Cross-Correlation between FAPAR and Hydroclimatic Anomalies
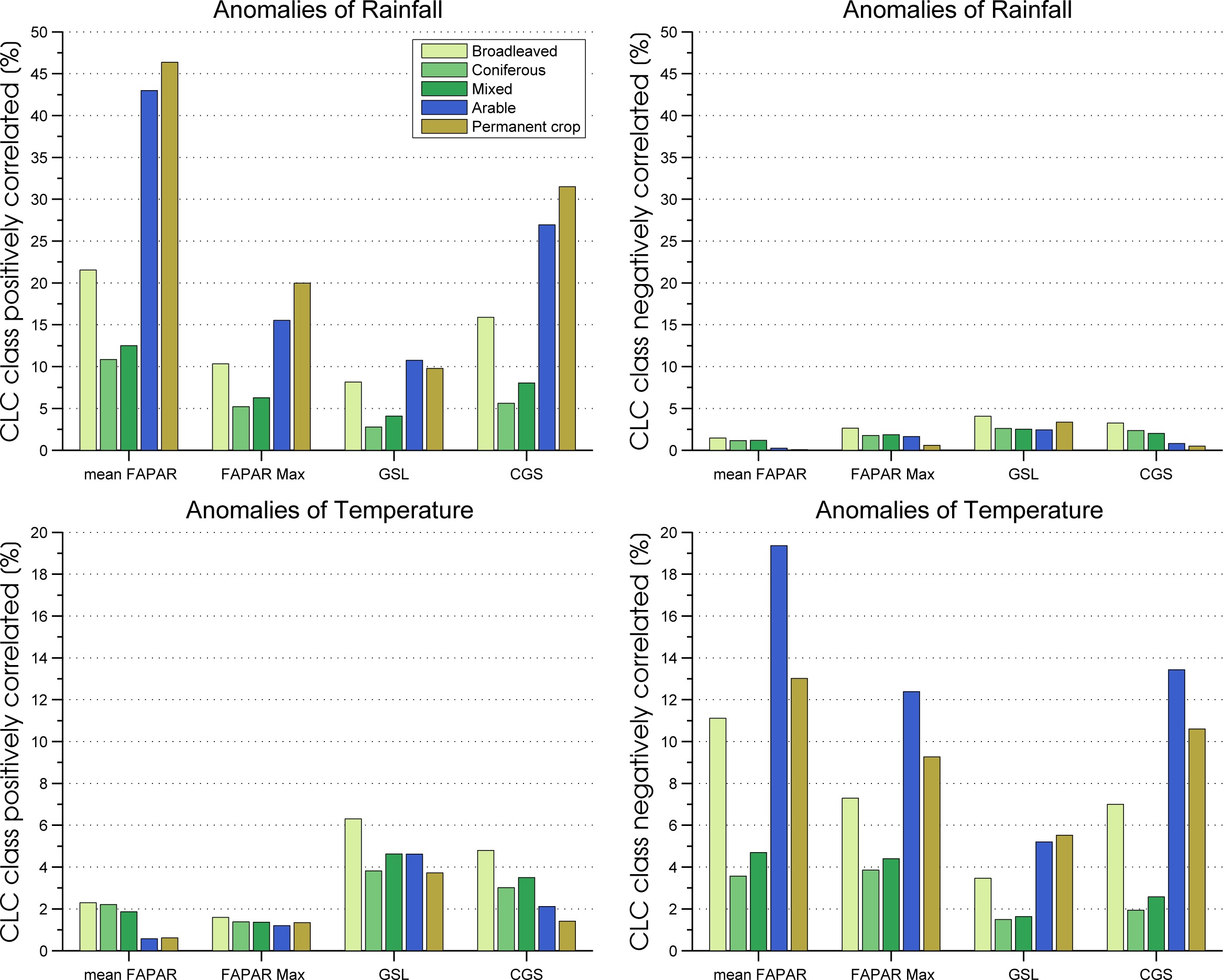
3.2. Correlation between Phenology Metrics and Hydroclimatic Anomalies
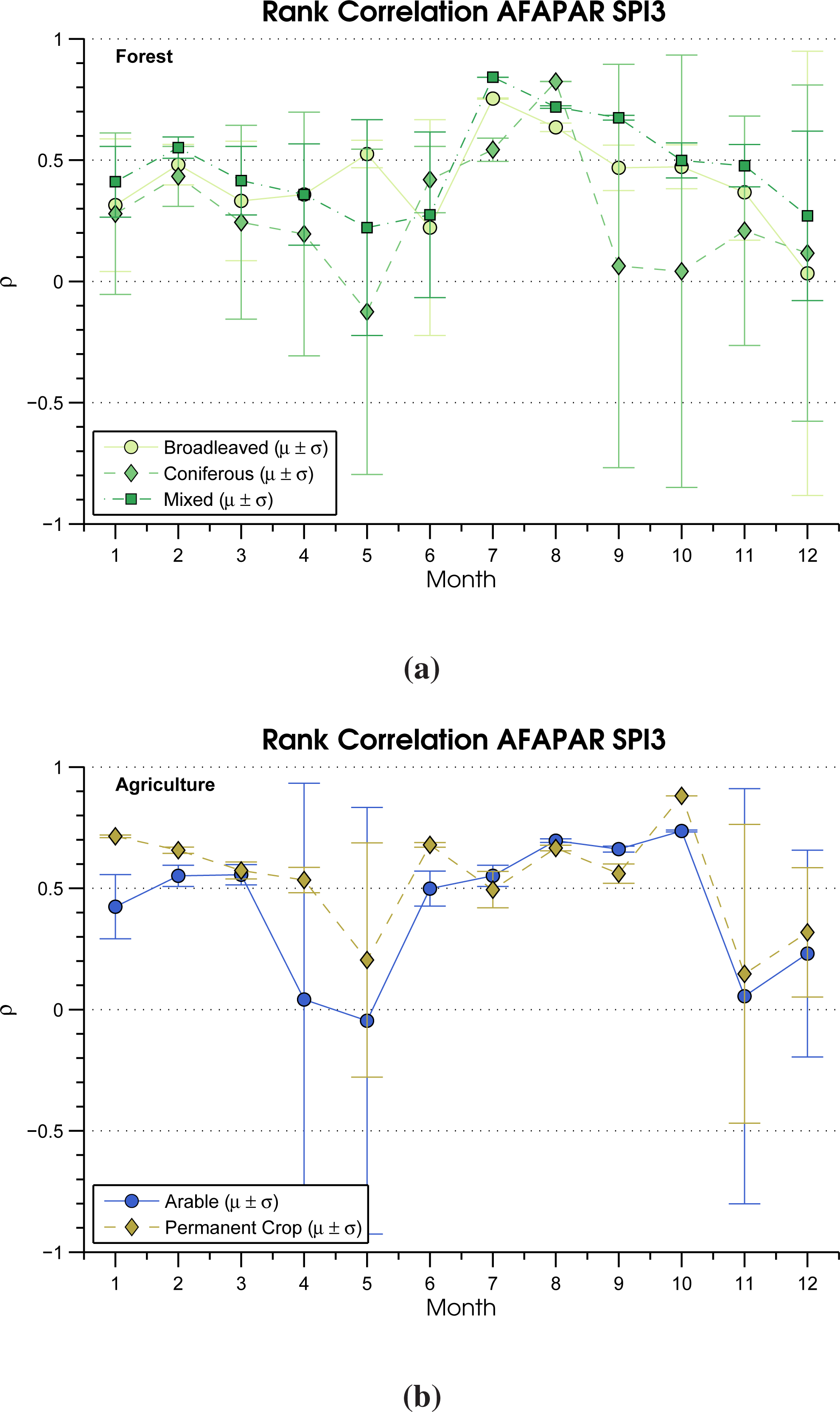
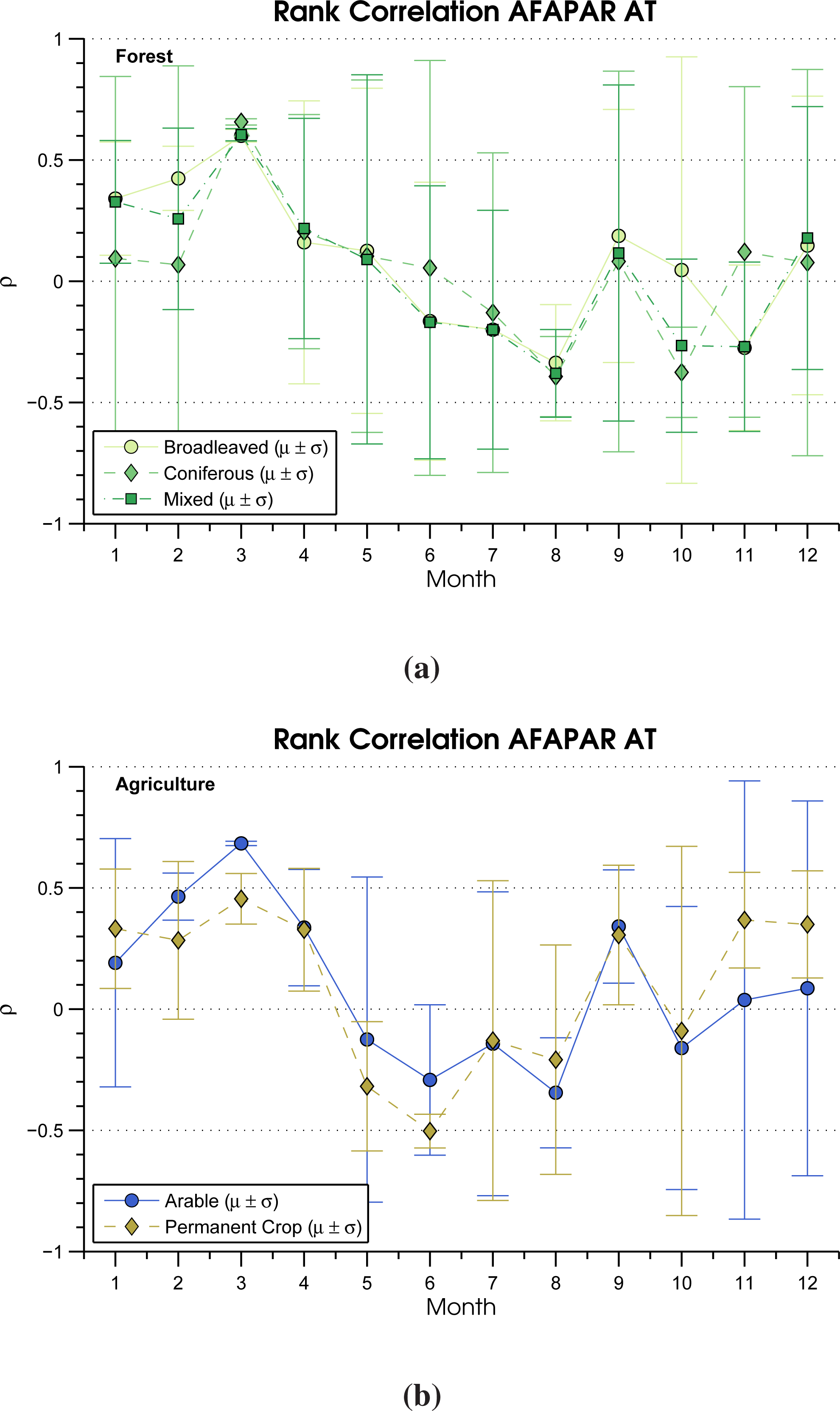
3.3. Rank Correlation
4. Discussion
5. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cox, P.; Betts, R.; Jones, C.; Spall, S.; Totterdell, I. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature 2000, 408, 184–187. [Google Scholar]
- Sánchez, E.; Gallardo, C.; Gaertner, M.; Arribas, A.; Castro, M. Future climate extreme events in the Mediterranean simulated by a regional climate model: A first approach. Glob. Planet. Chang 2004, 44, 163–180. [Google Scholar]
- Heimann, M.; Reichstein, M. Terrestrial ecosystem carbon dynamics and climate feedbacks. Nature 2008, 451, 289–292. [Google Scholar]
- Medvigy, D.; Wofsy, S.; Munger, J.; Moorcroft, P. Responses of terrestrial ecosystems and carbon budgets to current and future environmental variability. Proc. Natl. Acad. Sci. USA 2010, 107, 8275–8280. [Google Scholar]
- Ziehn, T.; Kattge, J.; Knorr, W.; Scholze, M. Improving the predictability of global CO2 assimilation rates under climate change. Geophys. Res. Lett 2011, 38. [Google Scholar] [CrossRef]
- Schwalm, C.; Williams, C.; Schaefer, K.; Baldocchi, D.; Black, T.; Goldstein, A.; Law, B.; Oechel, W.; Paw, U.K.; Scott, R. Reduction in carbon uptake during turn of the century drought in western North America. Nat. Geosci 2012, 5, 551–556. [Google Scholar] [Green Version]
- Parry, M. Climate Change 2007: Impacts, adaptation and vulnerability: Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Knorr, W.; Scholze, M.; Gobron, N.; Pinty, B.; Kaminski, T. Global-scale drought caused atmospheric CO2 increase. Eos Trans. Am. Geophys. Union 2005, 86, 178–181. [Google Scholar]
- Knorr, W.; Gobron, N.; Scholze, M.; Kaminski, T.; Schnur, R.; Pinty, B. Impact of terrestrial biosphere carbon exchanges on the anomalous CO2 increase in 2002–2003. Geophys. Res. Lett 2007, 34. [Google Scholar] [CrossRef]
- Delire, C.; de Noblet-Ducoudré, N.; Sima, A.; Gouirand, I. Vegetation dynamics enhancing long-term climate variability confirmed by two models. J. Clim 2011, 24, 2238–2257. [Google Scholar]
- Van Der Molen, M.; Dolman, A.; Ciais, P.; Eglin, T.; Gobron, N.; Law, B.; Meir, P.; Peters, W.; Phillips, O.; Reichstein, M.; et al. Drought and ecosystem carbon cycling. Agric. For. Meteorol 2011, 151, 765–773. [Google Scholar]
- Boisvenue, C.; Running, S. Impacts of climate change on natural forest productivity—Evidence since the middle of the 20th century. Glob. Chang. Biol 2006, 12, 862–882. [Google Scholar]
- Cañón, J.; Domínguez, F.; Valdes, J. Vegetation responses to precipitation and temperature: A spatiotemporal analysis of ecoregions in the Colorado River Basin. Int. J. Remote Sens 2011, 32, 5665–5687. [Google Scholar]
- Gessner, U.; Naeimi, V.; Klein, I.; Kuenzer, C.; Klein, D.; Dech, S. The relationship between precipitation anomalies and satellite-derived vegetation activity in Central Asia. Glob. Planet. Chang 2013, 110, 74–87. [Google Scholar]
- Hao, F.; Zhang, X.; Ouyang, W.; Skidmore, A.; Toxopeus, A. Vegetation NDVI linked to temperature and precipitation in the upper catchments of Yellow River. Environ. Model. Assess 2012, 17, 389–398. [Google Scholar]
- Sepulcre-Canto, G.; Horion, S.; Singleton, A.; Carrao, H.; Vogt, J. Development of a Combined Drought Indicator to detect agricultural drought in Europe. Nat. Hazards Earth Syst. Sci 2012, 12, 3519–3531. [Google Scholar]
- Weiss, M.; van den Hurk, B.; Haarsma, R.; Hazeleger, W. Impact of vegetation variability on potential predictability and skill of EC-Earth simulations. Clim. Dyn 2012, 39, 2733–2746. [Google Scholar]
- Horion, S.; Cornet, Y.; Erpicum, M.; Tychon, B. Studying interactions between climate variability and vegetation dynamic using a phenology based approach. Int. J. Appl. Earth Obs. Geoinform 2013, 20, 20–32. [Google Scholar]
- Woillez, M.N.; Kageyama, M.; Combourieu-Nebout, N.; Krinner, G. Simulating the vegetation response in western Europe to abrupt climate changes under glacial background conditions. Biogeosciences 2013, 10, 1561–1582. [Google Scholar]
- Baldocchi, D.; Falge, E.; Gu, L.; Olson, R.; Hollinger, D.; Running, S.; Anthoni, P.; Bernhofer, C.; Davis, K.; Evans, R.; et al. FLUXNET: A new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bull. Am. Meteorol. Soc 2001, 82, 2415–2434. [Google Scholar]
- Schwartz, M.; Ault, T.; Betancourt, J. Spring onset variations and trends in the continental United States: Past and regional assessment using temperature-based indices. Int. J. Climatol 2013, 33, 2917–2922. [Google Scholar]
- Gonsamo, A.; Chen, J.; D’Odorico, P. Deriving land surface phenology indicators from CO2 eddy covariance measurements. Ecol. Indic 2013, 29, 203–207. [Google Scholar]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol 2013, 169, 156–173. [Google Scholar]
- Los, S.; Collatz, G.; Bounoua, L.; Sellers, P.; Tucker, C. Global interannual variations in sea surface temperature and land surface vegetation, air temperature, and precipitation. J. Clim 2001, 14, 1535–1549. [Google Scholar]
- Wang, W.; Anderson, B.; Entekhabi, D.; Huang, D.; Su, Y.; Kaufmann, R.; Myneni, R. Intraseasonal interactions between temperature and vegetation over the boreal forests. Earth Interact 2007, 11, 1–30. [Google Scholar]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.; Baldocchi, D.; Bonan, G.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar]
- Forzieri, G.; Vivoni, E.R.; Feyen, L. Ecosystem biophysical memory in the southwestern North America climate system. Environ. Res. Lett 2013, 8. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar]
- Gobron, N.; Pinty, B.; Mélin, F.; Taberner, M.; Verstraete, M.; Belward, A.; Lavergne, T.; Widlowski, J.L. The state of vegetation in Europe following the 2003 drought. Int. J. Remote Sens 2005, 26, 2013–2020. [Google Scholar]
- Diffenbaugh, N. Sensitivity of extreme climate events to CO2 induced biophysical atmosphere-vegetation feedbacks in the western United States. Geophys. Res. Lett 2005, 32, 1–4. [Google Scholar]
- Lorenz, R.; Davin, E.; Lawrence, D.; Stöckli, R.; Seneviratne, S. How important is vegetation phenology for European climate and heatwaves? J. Clim 2013, 26, 10077–10100. [Google Scholar]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.; Seneviratne, S.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar]
- Field, C.; Barros, V.; Stocker, T.; Qin, D.; Dokken, D.; Ebi, K.; Mastrandrea, M.; Mach, K.; Plattner, G.K.; Allen, S.; et al. Summary for Policymakers. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation; A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change; IPCC: Cambridge, UK, 2012; pp. 1–582. [Google Scholar]
- Bossard, M.; Feranec, J.; Otahel, J. CORINE Land Cover Technical Guide Addendum 2000; Technical Report 40; European Environment Agency (EEA): Copenhagen, Denmark, 2000. [Google Scholar]
- European Environment Agency (EEA). CLC2006 Technical Guidelines; Technical Report 17/2007; EEA: Copenhagen, Denmark, 2007. [Google Scholar]
- Gobron, N.; Pinty, B.; Verstraete, M.M.; Govaerts, Y. The MERIS Global Vegetation Index (MGVI): Description and preliminary application. Int. J. Remote Sens 1999, 20, 1917–1927. [Google Scholar]
- GCOS-92. Implementation Plan for the Global Observing System for Climate in Support of the UNFCCC, WMO/TD No. 1219 GCOS-92; World Meteorological Organization (WMO): Geneva, Switzerland, 2004. [Google Scholar]
- GCOS-107. Systematic Observation Requirements for Satellite-Based Products for Climate: Supplemental Details to the Satellite-Based Component of the Implementation Plan for the Global Observing System for Climate in Support of the UNFCCC; Rep. WMO/TD 1338 GCOS-107; World Meteorological Organization (WMO): Geneva, Switzerland, 2006. [Google Scholar]
- GTOS-52. Terrestrial Essential Climate Variables for Assessment, Mitigation and Adaptation; Rep. GTOS-52; Food and Agriculture Organization (FAO) (United Nations): Rome, Italy, 2008. [Google Scholar]
- Jung, M.; Verstraete, M.; Gobron, N.; Reichstein, M.; Papale, D.; Bondeau, A.; Robustelli, M.; Pinty, B. Diagnostic assessment of European gross primary production. Glob. Chang. Biol 2008, 14, 2349–2364. [Google Scholar]
- Knorr, W.; Kaminski, T.; Scholze, M.; Gobron, N.; Pinty, B.; Giering, R.; Mathieu, P.P. Carbon cycle data assimilation with a generic phenology model. J. Geophys. Res.-Biogeosci 2010, 115. [Google Scholar] [CrossRef]
- Kaminski, T.; Knorr, W.; Scholze, M.; Gobron, N.; Pinty, B.; Giering, R.; Mathieu, P.P. Consistent assimilation of MERIS FAPAR and atmospheric CO2 into a terrestrial vegetation model and interactive mission benefit analysis. Biogeosciences 2012, 9, 3173–3184. [Google Scholar]
- Kato, T.; Knorr, W.; Scholze, M.; Veenendaal, E.; Kaminski, T.; Kattge, J.; Gobron, N. Simultaneous assimilation of satellite and eddy covariance data for improving terrestrial water and carbon simulations at a semi-arid woodland site in Botswana. Biogeosciences 2013, 10, 789–802. [Google Scholar] [Green Version]
- Jung, M.; Reichstein, M.; Ciais, P.; Seneviratne, S.; Sheffield, J.; Goulden, M.; Bonan, G.; Cescatti, A.; Chen, J.; de Jeu, R.; et al. Recent decline in the global land evapotranspiration trend due to limited moisture supply. Nature 2010, 467, 951–954. [Google Scholar]
- Rembold, F.; Atzberger, C.; Savin, I.; Rojas, O. Using low resolution satellite imagery for yield prediction and yield anomaly detection. Remote Sens 2013, 5, 1704–1733. [Google Scholar]
- Duveiller, G.; López-Lozano, R.; Baruth, B. Enhanced processing of 1-km spatial resolution fAPAR time series for sugarcane yield forecasting and monitoring. Remote Sens 2013, 5, 1091–1116. [Google Scholar]
- Gobron, N.; Pinty, B.; Aussedat, O.; Chen, J.; Cohen, W.; Fensholt, R.; Gond, V.; Huemmrich, K.; Lavergne, T.; Mélin, F.; et al. Evaluation of fraction of absorbed photosynthetically active radiation products for different canopy radiation transfer regimes: Methodology and results using Joint Research Center products derived from SeaWiFS against ground-based estimations. J. Geophys. Res. D: Atmos 2006, 111. [Google Scholar] [CrossRef]
- Gobron, N.; Pinty, B.; Aussedat, O.; Taberner, M.; Faber, O.; Mélin, F.; Lavergne, T.; Robustelli, M.; Snoeij, P. Uncertainty estimates for the FAPAR operational products derived from MERIS—Impact of top-of-atmosphere radiance uncertainties and validation with field data. Remote Sens. Environ 2008, 112, 1871–1883. [Google Scholar]
- Pinty, B.; Gobron, N.; Mélin, F.; Verstraete, M.M. Time Composite Algorithm Theoretical Basis Document; Eur Report, IES; Office for Official Publications of the European Communities: Luxembourg, 2002. [Google Scholar]
- Aussedat, O.; Gobron, N.; Pinty, B.; Taberner, M. MERIS Level 3 Land Surface Time Composite—Product File Description; Eur Report, IES; Office for Official Publications of the European Communities: Luxembourg, 2006. [Google Scholar]
- Ceccherini, G.; Gobron, N.; Robustelli, M. Harmonization of Fraction of Absorbed Photosynthetically Active Radiation (FAPAR) from Sea-ViewingWide Field-of-View Sensor (SeaWiFS) and Medium Resolution Imaging Spectrometer Instrument (MERIS). Remote Sens 2013, 5, 3357–3376. [Google Scholar] [Green Version]
- Haylock, M.; Hofstra, N.; Klein Tank, A.; Klok, E.; Jones, P.; New, M. A European daily high-resolution gridded data set of surface temperature and precipitation for 1950–2006. J. Geophys. Res. D: Atmos 2008, 113. [Google Scholar] [CrossRef]
- Van Der Linden, P.; Mitchell, J. ENSEMBLES: Climate Change and Its Impacts: Summary of Research and Results from the ENSEMBLES Project; Technical Report; Met Office Hadley Centre: Exeter, UK, 2009. [Google Scholar]
- McKee, T.B.; Doeskin, N.J.; Kleist, J. The Relationship of Drought Frequency and Duration to Time Scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; American Meteorological Society: Boston, MA, USA, 1993; pp. 179–184. [Google Scholar]
- Du, J.; Fang, J.; Xu, W.; Shi, P. Analysis of dry/wet conditions using the standardized precipitation index and its potential usefulness for drought/flood monitoring in Hunan Province, China. Stochastic Environ. Res. Risk Assess 2013, 27, 377–387. [Google Scholar]
- Seiler, R.; Hayes, M.; Bressan, L. Using the standardized precipitation index for flood risk monitoring. Int. J. Climatol 2002, 22, 1365–1376. [Google Scholar]
- Ji, L.; Peters, A. Assessing vegetation response to drought in the northern Great Plains using vegetation and drought indices. Remote Sens. Environ 2003, 87, 85–98. [Google Scholar]
- Ceccherini, G.; Gobron, N.; Migliavacca, M. Long-Term Measurements of Plant Phenology over Europe Derived from SeaWiFS and MERIS. In Proceedings of the ESA Living Planet Symposium 2013, Edinburgh, UK, 9–13 September 2013; Lacoste, H., Ouwehand, L., Eds.; European Space Agency Special Publication: Noordwijk, The Netherlands, 2013; Volume 722. [Google Scholar]
- Richardson, A.D.; Black, T.A.; Ciais, P.; Delbart, N.; Friedl, M.A.; Gobron, N.; Hollinger, D.Y.; Kutsch, W.L.; Longdoz, B.; Luyssaert, S.; et al. Influence of spring and autumn phenological transitions on forest ecosystem productivity. Philos. Trans. R. Soc. B: Biol. Sci 2010, 365, 3227–3246. [Google Scholar] [Green Version]
- Fensholt, R.; Sandholt, I.; Rasmussen, M.; Stisen, S.; Diouf, A. Evaluation of satellite based primary production modelling in the semi-arid Sahel. Remote Sens. Environ 2006, 105, 173–188. [Google Scholar]
- Kendall, M.; Gibbons, J.D. Rank Correlation Methods, 5th ed.; The Griffin: London, UK, 1990. [Google Scholar]
- Waple, A.; Lawrimore, J. State of the climate in 2002. Bull. Am. Meteorol. Soc 2003, 84, S1–S68. [Google Scholar]
- Levinson, D.; Waple, A. State of the climate in 2003. Bull. Am. Meteorol. Soc 2004, 85, S1–S72. [Google Scholar]
- Rossi, S.; Laguardia, G.; Kurnik, B.; Robustelli, M.; Niemeyer, S.; Gobron, N. Multisource Detection of Drought Events at the European Scale. In Proceedings of the 2nd MERIS/(A)ATSR Workshop (ESA-ESRIN), Frascati, Italy, 22–26 September 2008; Lacoste, H., Ouwehand, L., Eds.; European Space Agency Special Publication: Noordwijk, TheNetherlands, 2008; Volume 666. [Google Scholar]
- Peel, M.; Finlayson, B.; McMahon, T. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci 2007, 11, 1633–1644. [Google Scholar]
- Richardson, A.; Hollinger, D.; Dail, D.; Lee, J.; Munger, J.; O’ Keefe, J. Influence of spring phenology on seasonal and annual carbon balance in two contrasting New England forests. Tree Physiol 2009, 29, 321–331. [Google Scholar]
- Le Maire, G.; Delpierre, N.; Jung, M.; Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ibrom, A.; Kolari, P.; Longdoz, B.; et al. Detecting the critical periods that underpin interannual fluctuations in the carbon balance of European forests. J. Geophys. Res. G: Biogeosci 2010, 115, G00H03. [Google Scholar] [CrossRef]
- Augspurger, C. Spring 2007 warmth and frost: Phenology, damage and refoliation in a temperate deciduous forest. Funct. Ecol 2009, 23, 1031–1039. [Google Scholar]
- Walther, G. Two steps forward, one step back. Funct. Ecol 2009, 23, 1029–1030. [Google Scholar]
- Rosenzweig, C.; Tubiello, F.; Goldberg, R.; Mills, E.; Bloomfield, J. Increased crop damage in the US from excess precipitation under climate change. Glob. Environ. Chang 2002, 12, 197–202. [Google Scholar]
- Baruth, B.; Biavetti, I.; Bussay, A.; Ceglar, A.; Chukaliev, O.; Duveiller, G.; Fontana, G.; Garcia Condado, S.; Hooker, J.; Karetsos, S.; et al. Crop Monitoring in Europe—MARS Bulletin; Technical Report Volume 21 No. 6; EC Joint Research Centre: Ispra, Italy, 2013. [Google Scholar]
- Huete, A.; Didan, K.; Shimabukuro, Y.; Ratana, P.; Saleska, S.; Hutyra, L.; Yang, W.; Nemani, R.; Myneni, R. Amazon rainforests green-up with sunlight in dry season. Geophys. Res. Lett 2006, 33. [Google Scholar] [CrossRef]
- Caldararu, S.; Purves, D.; Palmer, P. Phenology as a strategy for carbon optimality: A global model. Biogeosciences 2014, 11, 763–778. [Google Scholar]
- Donlon, C.; Berruti, B.; Buongiorno, A.; Ferreira, M.H.; Féménias, P.; Frerick, J.; Goryl, P.; Klein, U.; Laur, H.; Mavrocordatos, C.; et al. The Global Monitoring for Environment and Security (GMES) Sentinel-3 mission. Remote Sens. Environ 2012, 120, 37–57. [Google Scholar]
- Baldocchi, D. Breathing’ of the terrestrial biosphere: Lessons learned from a global network of carbon dioxide flux measurement systems. Aust. J. Bot 2008, 56, 1–26. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Cover CLC2006 | SPI-3 (%) | AT (%) | ρ Pos. SPI-3 | ρ Neg. SPI-3 | ρ Pos. AT | ρ Neg. AT |
|---|---|---|---|---|---|---|
| Broadleaved forest | 38.37 | 20.48 | 0.33 | 0.23 | 0.23 | 0.28 |
| Coniferous forest | 28.57 | 14.22 | 0.29 | 0.23 | 0.21 | 0.23 |
| Mixed forest | 28.38 | 17.53 | 0.31 | 0.22 | 0.21 | 0.24 |
| Arable | 64.35 | 25.02 | 0.33 | 0.23 | 0.26 | 0.26 |
| Permanent crop | 73.07 | 18.80 | 0.36 | 0.21 | 0.32 | 0.25 |
| Land Cover CLC2006 | SPI-3 (%) | AT (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Lag 0 | Lag 1 | Lag 2 | Lag 3 | Lag 0 | Lag 1 | Lag 2 | Lag 3 | |
| Broadleaved forest | 59.38 | 27.09 | 13.54 | 0 | 80.25 | 10.83 | 8.92 | 0 |
| Coniferous forest | 68.17 | 18.51 | 13.31 | 0 | 71.69 | 11.59 | 16.72 | 0 |
| Mixed forest | 65.50 | 21.38 | 13.12 | 0 | 73.00 | 10.93 | 16.08 | 0 |
| Arable | 47.91 | 33.82 | 18.27 | 0 | 80.14 | 13.00 | 6.85 | 0 |
| Permanent crop | 48.90 | 37.72 | 13.39 | 0 | 78.19 | 10.40 | 11.41 | 0 |
| Metrics | Rain Pos. (%) | Rain Neg. (%) | Temperature Pos. (%) | Temperature Neg. (%) |
|---|---|---|---|---|
| Mean FAPAR | 26.8 | 0.1 | 1.5 | 11.6 |
| Maximum FAPAR | 10.8 | 1.9 | 1.3 | 8.1 |
| GSL | 7.3 | 2.9 | 4.7 | 3.5 |
| CGS | 17.0 | 1.9 | 3.0 | 7.8 |
© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ceccherini, G.; Gobron, N.; Migliavacca, M. On the Response of European Vegetation Phenology to Hydroclimatic Anomalies. Remote Sens. 2014, 6, 3143-3169. https://doi.org/10.3390/rs6043143
Ceccherini G, Gobron N, Migliavacca M. On the Response of European Vegetation Phenology to Hydroclimatic Anomalies. Remote Sensing. 2014; 6(4):3143-3169. https://doi.org/10.3390/rs6043143
Chicago/Turabian StyleCeccherini, Guido, Nadine Gobron, and Mirco Migliavacca. 2014. "On the Response of European Vegetation Phenology to Hydroclimatic Anomalies" Remote Sensing 6, no. 4: 3143-3169. https://doi.org/10.3390/rs6043143