
Monitoring the Distribution and Dynamics of an Invasive Grass in Tropical Savanna Using Airborne LiDAR

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
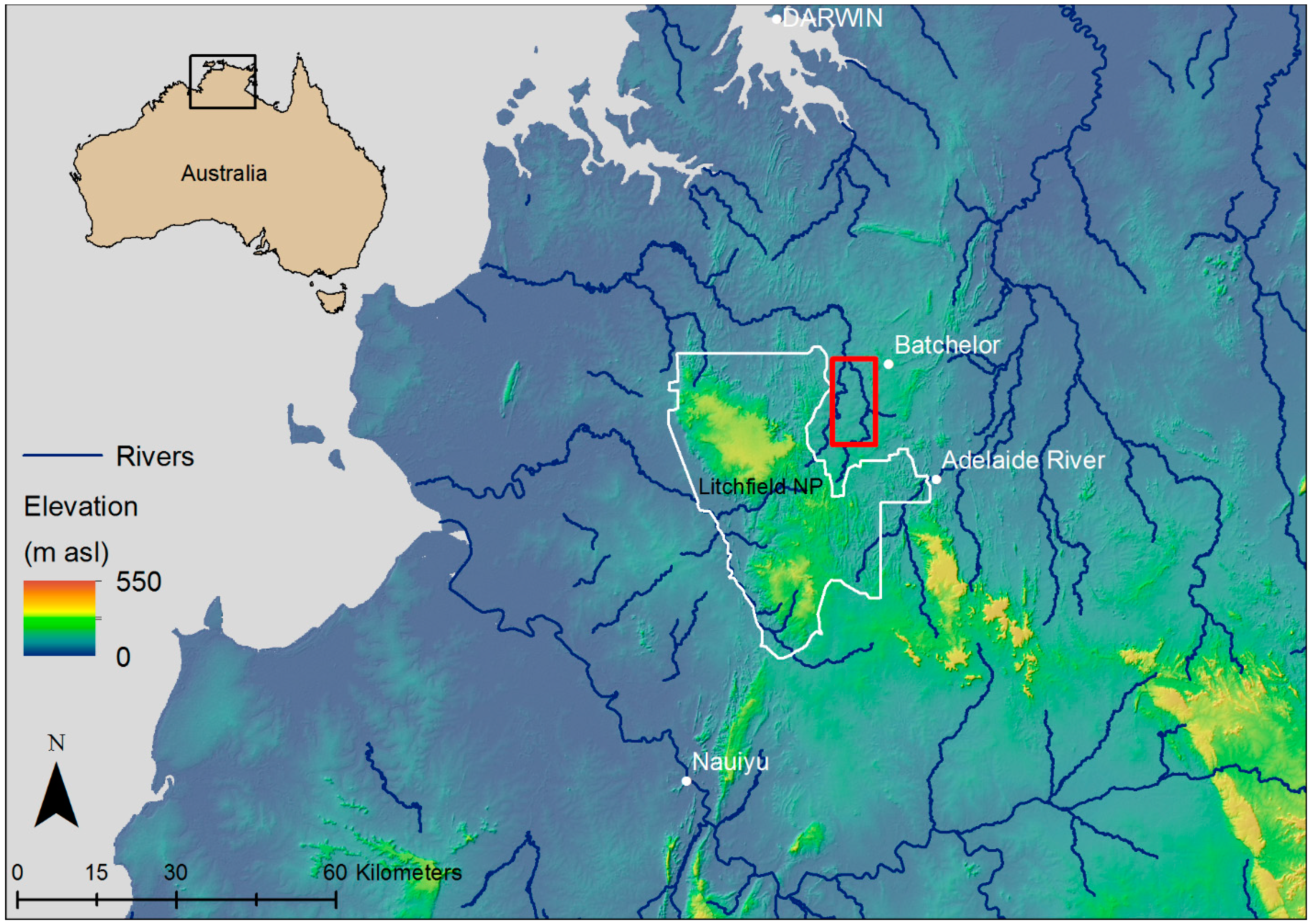
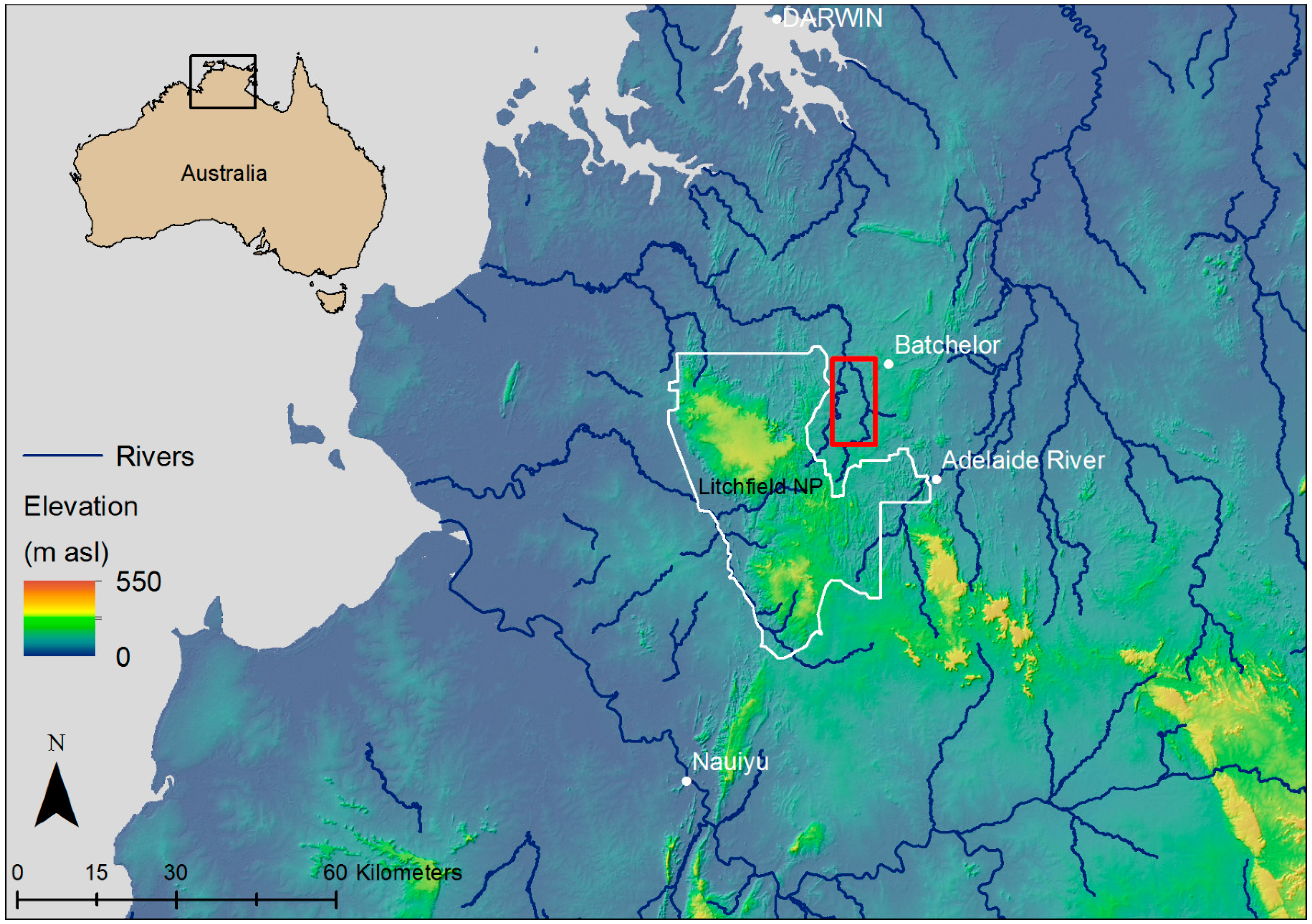
2.1. Study Species and Site Location

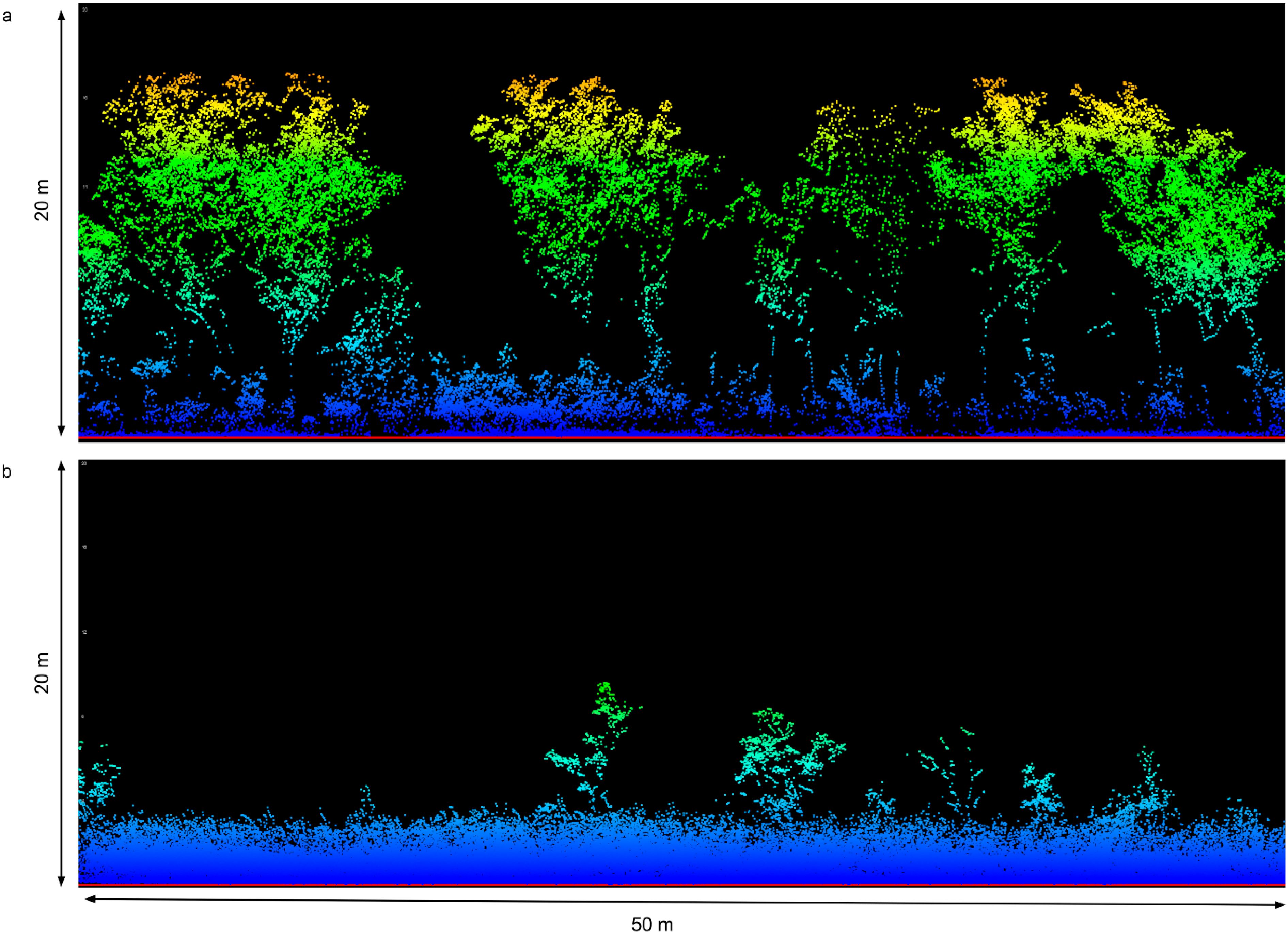
2.2. Airborne LiDAR Acquisition and Processing
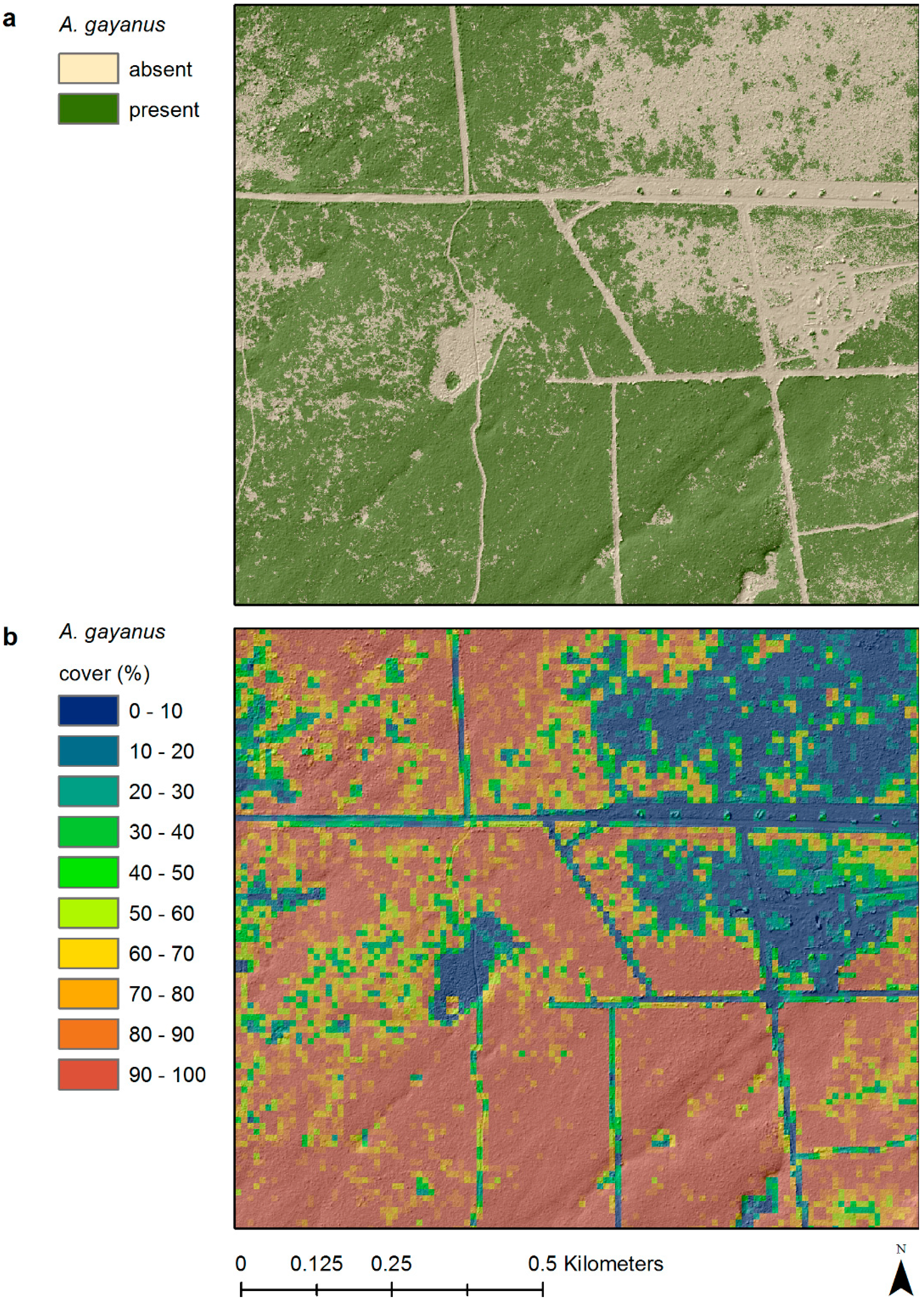
2.3. A. gayanus Classification and Cover Mapping
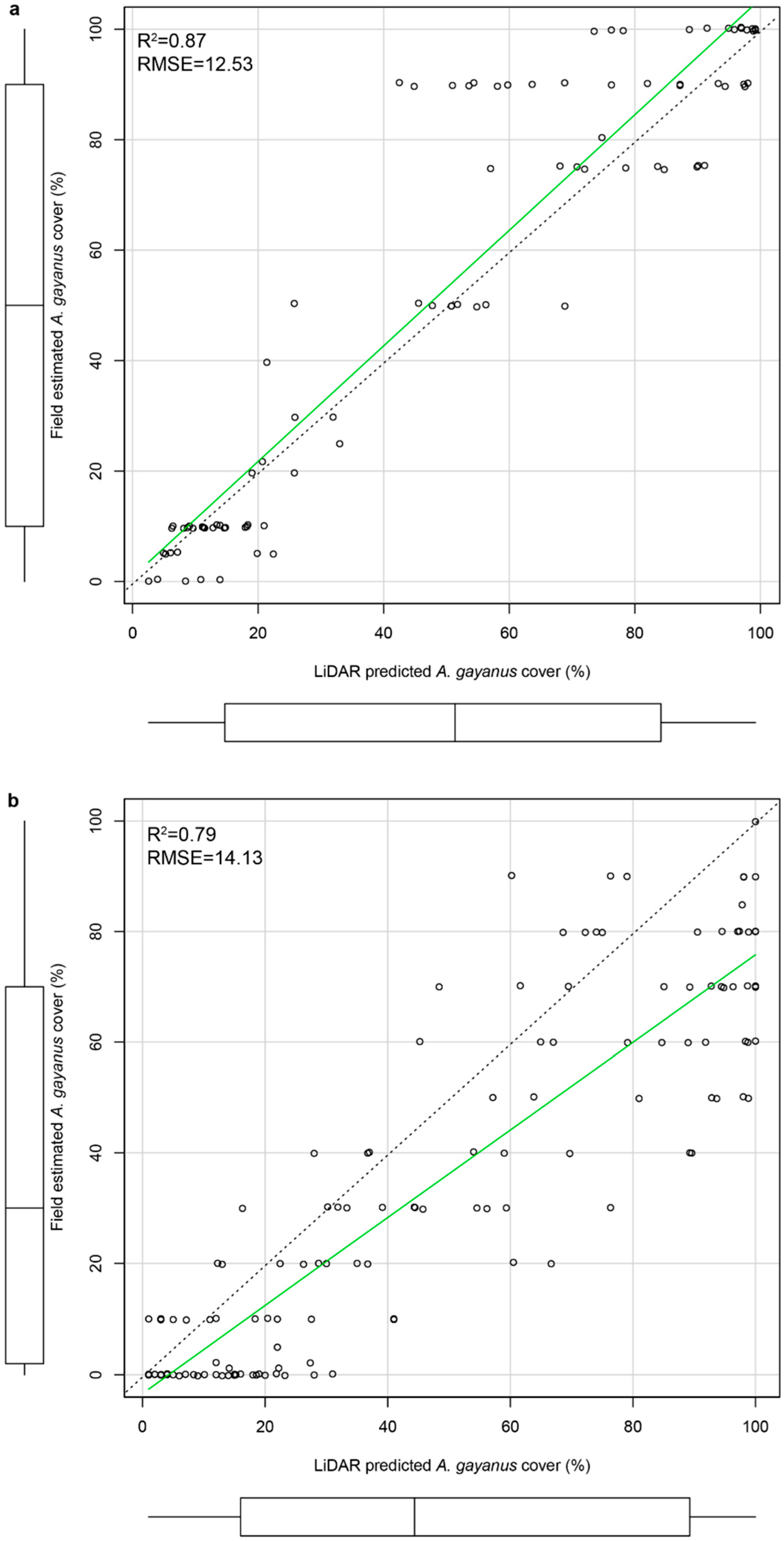
2.4. Field-Validation of LiDAR Predictions
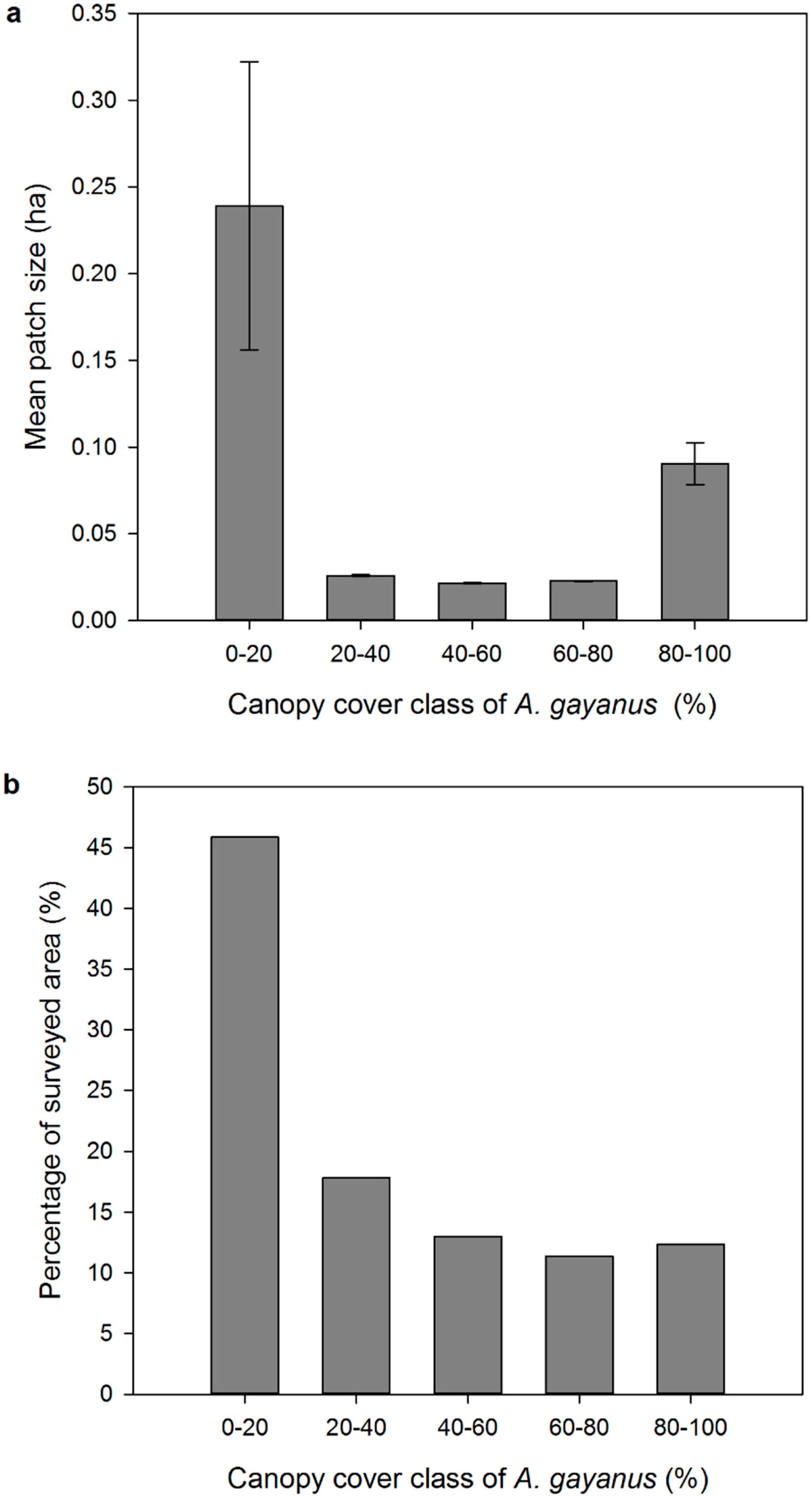
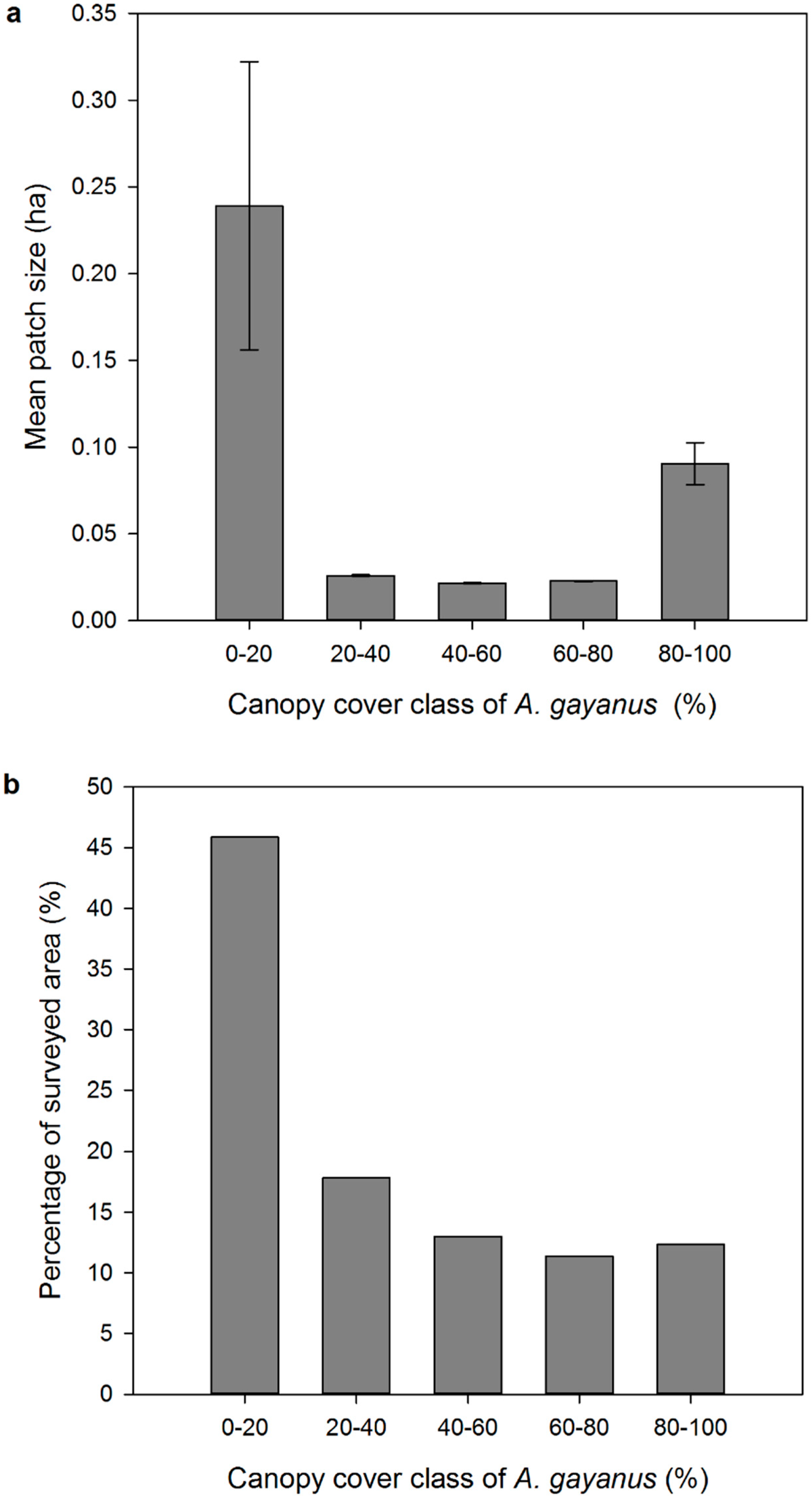
3. Results




4. Discussion
4.1. Airborne LiDAR Based Estimations of A. gayanus Cover
4.2. Implications for A. gayanus Invasion Ecology and Management

5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Grace, J.; José, J.S.; Meir, P.; Miranda, H.S.; Montes, R.A. Productivity and carbon fluxes of tropical savannas. J. Biogeogr. 2006, 33, 387–400. [Google Scholar] [CrossRef]
- Beringer, J.; Hutley, L.B.; Tapper, N.J.; Cernusak, L.A. Savanna fires and their impact on net ecosystem productivity in North Australia. Glob. Chang. Biol. 2007, 13, 990–1004. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed]
- D’Antonio, C.M.; Vitousek, P.M. Biological invasions by exotic grasses, the grass/fire cycle, and global change. Annu. Rev. Ecol. Syst. 1992, 23, 63–87. [Google Scholar]
- Rossiter-Rachor, N.A.; Setterfield, S.A.; Douglas, M.M.; Hutley, L.B.; Cook, G.D. Andropogon gayanus (gamba grass) invasion increases fire-mediated nitrogen losses in the tropical savannas of northern Australia. Ecosystems 2008, 11, 77–88. [Google Scholar] [CrossRef]
- Cook, G.D.; Grice, A.C. Historical perspectives on invasive grasses and their impact on wildlife in Australia. Wildl. Soc. Bull. 2013, 37, 469–477. [Google Scholar]
- Douglas, M.M.; O’Connor, R.A. Weed invasion changes fuel characteristics: Para Grass (Urochloa mutica (Forssk.) T.Q. Nguyen) on a tropical floodplain. Ecol. Manag. Restor. 2004, 5, 143–145. [Google Scholar] [CrossRef]
- Setterfield, S.A.; Rossiter-Rachor, N.A.; Douglas, M.M.; Wainger, L.; Petty, A.M.; Barrow, P.; Shepherd, I.J.; Ferdinands, K.B. Adding fuel to the fire: The impacts of non-native grass invasion on fire management at a regional scale. PLoS ONE 2013, 8, e59144. [Google Scholar] [CrossRef] [PubMed]
- Flores, T.A.; Setterfield, S.A.; Douglas, M.M. Seedling recruitment of the exotic grass Andropogon gayanus (Poaceae) in northern Australia. Aust. J. Bot. 2005, 53, 243–249. [Google Scholar] [CrossRef]
- Brooks, K.J.; Setterfield, S.A.; Douglas, M.M. Exotic grass invasions: Applying a conceptual framework to the dynamics of degradation and restoration in Australia’s tropical savannas. Restor. Ecol. 2010, 18, 188–197. [Google Scholar] [CrossRef]
- Setterfield, S.A.; Rossiter-Rachor, N.A.; Hutley, L.B.; Douglas, M.M.; Williams, R.J. Turning up the heat: The impacts of Andropogon gayanus (gamba grass) invasion on fire behaviour in northern Australian savannas. Divers. Distrib. 2010, 16, 854–861. [Google Scholar] [CrossRef]
- Rossiter-Rachor, N.A.; Setterfield, S.A.; Douglas, M.M.; Hutley, L.B.; Cook, G.D.; Schmidt, S. Invasive Andropogon gayanus (gamba grass) is an ecosystem transformer of nitrogen relations in Australian savanna. Ecol. Appl. 2009, 19, 1546–1560. [Google Scholar] [CrossRef] [PubMed]
- Rossiter, N.A.; Setterfield, S.A.; Douglas, M.M.; Hutley, L.B. Testing the grass-fire cycle: Alien grass invasion in the tropical savannas of northern Australia. Divers. Distrib. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Adams, V.M.; Setterfield, S.A. Estimating the financial risks of Andropogon gayanus to greenhouse gas abatement projects in northern Australia. Environ. Res. Lett. 2013, 8, 025018. [Google Scholar] [CrossRef]
- Hutley, L.B.; Setterfield, S.A. Savanna. In Encyclopedia of Ecology; Jorgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 3143–3154. [Google Scholar]
- Shaw, D.R. Translation of remote sensing data into weed management decisions. Weed Sci. 2005, 53, 264–273. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balch, J.K.; Bradley, B.A.; D’Antonio, C.M.; Gómez-Dans, J. Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob. Chang. Biol. 2013, 19, 173–183. [Google Scholar] [CrossRef]
- Pettorelli, N.; Safi, K.; Turner, W. Satellite remote sensing, biodiversity research and conservation of the future. Philos. Trans. R. Society B: Biol. Sci. 2014, 369, 20130190. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Davis, A.J.; Meentemeyer, R.K. Detecting understory plant invasion in urban forests using LiDAR. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 267–279. [Google Scholar] [CrossRef]
- Lefsky, M.; Cohen, W.; Parker, G.; Harding, D. Lidar remote sensing for ecosystem studies. BioScience 2002, 52, 19–30. [Google Scholar] [CrossRef]
- Levick, S.R.; Rogers, K.H. Structural biodiversity monitoring in savanna ecosystems: Integrating LiDAR and high resolution imagery through object-based image analysis. In Object-Based Image Analysis: Spatial Concepts for Knowledge-Driven Remote Sensing Applications; Blaschke, T., Lang, S., Hay, G., Eds.; Springer-Verlag: Berlin, Germany, 2008; pp. 477–492. [Google Scholar]
- Levick, S.R.; Baldeck, C.A.; Asner, G.P. Demographic legacies of fire history in an African savanna. Funct. Ecol, 2014, 29, 131–139. [Google Scholar] [CrossRef]
- Adams, V.M.; Setterfield, S.A. Optimal dynamic control of invasions: Applying a systematic conservation approach. Ecol. Appl. (in press).
- Popescu, S.C.; Zhao, K. A voxel-based lidar method for estimating crown base height for deciduous and pine trees. Remote Sens. Environ. 2008, 112, 767–781. [Google Scholar] [CrossRef]
- Asner, G.P.; Levick, S.R.; Kennedy-Bowdoin, T.; Knapp, D. E.; Emerson, R.; Jacobson, J.; Colgan, M.S.; Martin, R.E. Large-scale impacts of herbivores on the structural diversity of African savannas. Proc. Natl. Acad. Sci. USA 2009, 106, 4947–4952. [Google Scholar] [CrossRef] [PubMed]
- Levick, S.R.; Asner, G.P. The rate and spatial pattern of treefall in a savanna landscape. Biol. Conserv. 2013, 157, 121–127. [Google Scholar] [CrossRef]
- Lee, A.C.; Lucas, R.M. A LiDAR-derived canopy density model for tree stem and crown mapping in Australian forests. Remote Sens. Environ. 2007, 111, 493–518. [Google Scholar] [CrossRef]
- Asner, G. Hyperspectral remote sensing of canopy chemistry, physiology and diversity in tropical rainforests. In Hyperspectral Remote Sensing of Tropical and Sub-Tropical Forests; Kalacska, M., Sanchez-Azofeifa, G.A., Eds.; Taylor and Francis Group: Oxford, UK, 2008; pp. 261–288. [Google Scholar]
- Asner, G.P.; Levick, S.R. Landscape-scale effects of herbivores on treefall in African savannas. Ecol. Lett. 2012, 15, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levick, S.R.; Setterfield, S.A.; Rossiter-Rachor, N.A.; Hutley, L.B.; McMaster, D.; Hacker, J.M. Monitoring the Distribution and Dynamics of an Invasive Grass in Tropical Savanna Using Airborne LiDAR. Remote Sens. 2015, 7, 5117-5132. https://doi.org/10.3390/rs70505117
Levick SR, Setterfield SA, Rossiter-Rachor NA, Hutley LB, McMaster D, Hacker JM. Monitoring the Distribution and Dynamics of an Invasive Grass in Tropical Savanna Using Airborne LiDAR. Remote Sensing. 2015; 7(5):5117-5132. https://doi.org/10.3390/rs70505117
Chicago/Turabian StyleLevick, Shaun R., Samantha A. Setterfield, Natalie A. Rossiter-Rachor, Lindsay B. Hutley, Damien McMaster, and Jorg M. Hacker. 2015. "Monitoring the Distribution and Dynamics of an Invasive Grass in Tropical Savanna Using Airborne LiDAR" Remote Sensing 7, no. 5: 5117-5132. https://doi.org/10.3390/rs70505117